Introduction
In 2008, there were a 989,600 new cases of gastric
cancer (GC) and a GC-associated mortality rate of 738,000 worldwide
(1). Although the rate of
GC-associated mortality has declined in the last 50 years (2), it remains the second leading cause of
cancer-associated mortality worldwide, with an incidence rate
ranking fourth following lung, breast and colorectal cancer
(3). Systemic chemotherapy remains
critical in the treatment of GC.
DOX is a well-known chemotherapeutic treatment used
in Hodgkin's lymphoma, non-Hodgkin's lymphoma, multiple myeloma,
acute leukemia, Kaposi's sarcoma, Ewing's sarcoma, Wilms' tumor and
in solid tumors of the breast, adrenal cortex, prostate, thyroid,
head and neck, endometrium, lung and ovary (4). DOX is widely used to treat GC in
combination regimens with etoposide, cisplatin and 5-fluorouracil.
However, drug resistance to DOX significantly limits its efficacy.
Resistance to DOX is often associated with the expression of
p-glycoprotein (P-gp) (4).
Multidrug resistance (MDR) is a complex process due
to several mechanisms. Numerous resistance-associated proteins,
including P-gp, have been found in different cancer cells, and
overexpression of this transmembrane glycoprotein causes
accelerated efflux of several lipid-soluble chemotherapeutic agents
from the cells, decreasing their efficacy and leading to treatment
failure (5). The difficulty of
reversing this mechanism underlies its complexity.
Arsenic trioxide (As2O3)
appears to achieve complete remission in 70% of patients with acute
promyelocytic leukemia (APL) by inducing cancer cell apoptosis
(6). Additional studies have
confirmed that low doses of As2O3 induces
complete remission in 90% of relapsed patients with APL (7–9). To
date, a number of studies have provided novel insights into the
pathogenesis of this agent and indicate that
As2O3 may be useful in treating other types
of cancer (10,11). It has become evident that the
effects of As2O3 are not restricted to APL
cells, but that As2O3 may also be effective
in other malignant cells in several of types of cancer, including
chronic myeloid leukemia, multiple myeloma, and solid prostate,
esophageal and ovarian tumors (12–15).
A number of mechanisms are associated with the effects of
As2O3, including the induction of apoptosis,
inhibition of growth, inhibition of differentiation and
angiogenesis, and inhibition of mitochondrial respiration (16–18).
In addition, several signaling pathways are involved in the
antitumoral activity of As2O3, including
inactivation of the Notch, c-Jun NH2-terminal kinase (JNK) and
extracellular signal-regulated kinase (ERK) signaling pathways
(19–21). Human GC cells have been found to be
sensitive to As2O3 (22).
Few studies have investigated the involvement of Ras
and P-gp in MDR in GC cancer. The present study was designed to
assess As2O3 in reversing DIX-induced MDR in
GC cancer cell lines. The association between the antitumor effects
of As2O3 and the Ras/phosphorylated
(p-)ERK1/2 signaling pathway was also examined. The results of the
present study aimed to provide novel insights into the treatment of
gastric cancer.
Materials and methods
Chemicals and reagents
In the present study, As2O3
and 3-(4, 5-Dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide
(MTT) were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Cyclosporin A (CsA) was purchased from Novartis (Bale,
Switzerland). RPMI 1640, fetal bovine serum (FBS), and dimethyl
sulfoxide (DMSO) were purchased from Thermo Fisher Scientific
(Hyclone, Waltham, MA, USA). DOX was purchased from Zhejiang
Haizheng Pharmaceutical Co. (Taizhou, China). The recombinant human
granulocyte colony stimulating factor (rhG-CSF) was purchased from
Qilu Pharmaceutical (Jinan, China). The mouse anti-P-gp (P-170),
Ras (cat. no. ZA-0343), p-ERK1/2 (cat. no. sc-7383), horseradish
peroxidase (HRP)-conjugated goat-anti-mouse secondary antibody and
the corresponding immunohisto-chemistry kits were purchased from
Beijing Golden Bridge Biotechnology Co. (Beijing, China).
Cell culture
The parental SGC7901/S human GC cell line and the
DOX-resistant SGC7901/ADM cell line were obtained from the Key
Laboratory of Cancer Biology and Xijing Hospital of Digestive
Diseases, Fourth Military Medical University (Xi'an, China). The
SGC7901/ADM cells were obtained from the SGC7901/S cells, based on
a previously described method (23). In brief, the SGC7901/ADM cells were
induced in vitro by continuous exposure of SGC7901 parent
cells to adriamycin (ADM) at concentrations of 0.25 µg/l to
25 µg/l. Cell lines capable of sustained growth in medium
containing 25 µg/l ADM were considered to be resistant after
3 months. The SGC7901/S cells were cultured in RPMI 1640 medium
supplemented with 100 U/ml penicillin, 100 µg/ml
streptomycin and 10% (v/v) heat-inactivated FBS. The SGC7901/ADM
cells were cultured in the same medium with 1 µg/ml DOX, and
were grown without DOX for 2 weeks prior to the experiments. All
the cells were cultured at 37°C in a 5% CO2 humidified
atmosphere and analyses were performed in the logarithmic growth
phase.
Cytotoxicity and sensitivity of SGC7901/S
and SGC7901/ADM cells to As2O3
The SGC7901/S and SGC7901/ADM cells were
trypsinized, centrifuged at 1,000 × g for 5 min, harvested and
seeded into 96-well plates, in triplicate, at a density of
1×104 cells/well (100 µl) at 37°C. Following
overnight incubation, the cells became adherent. The culture medium
was removed and As2O3 was added at 12
concentrations (0.01, 0.05, 0.10, 0.25, 0.50, 0.75, 1.0, 2.5, 5.0,
10.0, 15.0 or 20.0 mM). The cells (1×104 cells/well)
were incubated for 24 h, 48 h and 72 h, and were further incubated
for an additional 4 h in the presence of 10 µl/well of MTT
reagent, containing 5.00 mg/ml in phosphate-buffered saline (PBS).
Following aspiration of the medium, the resulting formazan was
dissolved with DMSO (100 µl/cell). The plates were agitated
mechanically for 60 sec, and the optical density (OD) or absorbance
was measured immediately at 490 nm on a microplate reader (AR2010;
Bio-Rad Laboratories, Inc., Hercules, CA, USA). The cell survival
rate was calculated as: (OD of the assessment group / OD of the
control group) × 100%. The inhibition rate and half maximal
inhibitory concentration of the reagent (IC50) were
calculated based on the survival rate (SR) (24,25).
The dose of As2O3, which resulted in a SR of
>95% was considered non-toxic, and the dose resulting in a SR
>85% was considered mildly-toxic.
Resistance of SGC7901/ADM cells and the
resistance-reversing effect of As2O3
The present study included five groups, which were
as follows: SGC7901/ADM, SGC7901/S,
SGC7901/ADM+As2O3 (0.10 µM),
SGC7901/ADM+As2O3 (0.50 µM) and
SGC7901/ADM+CsA (4.00 µg/ml). These two concentrations of
As2O3 were selected as the non-toxic and the
mildly-toxic doses in the experiment described above. To
investigate the drug resistance-reversing effect of
As2O3, DOX (0.1, 0.5, 1.0, 2.5, 5.0, 10.0,
15.0 and 30.0 µg/ml) was added to the cell culture
(1×104 cells/well), rather than
As2O3. The cells were then incubated for 24 h
and 48 h at 37°C. The drug resistance ratio was calculated as the
IC50 of the drug-resistant cells divided by the
IC50 of the parental cells. The reversal ratio was
calculated as the IC50 of the drug-resistant cells
divided by the IC50 of the drug-resistant cells treated
with reversal agents.
Reversing effect of
As2O3 following treatment with rhG-CSF
The SGC7901/ADM, SGC7901/S, SGC7901/ADM+0.10
µM As2O3, SGC7901/ADM+0.50 µM
As2O3, and SGC7901/ADM+4.00 µg/ml CsA
cells (1×104 cells/well) were incubated with rhG-CSF
(1.50 µg/ml) for 3 days at 37°C, and then cultured with DOX
at different concentrations (0.1, 0.5, 1.0, 2.5, 5.0, 10.0, 30.0,
40.0 and 50.0 µM) for a further 2 days at 37°C, following
which the reversing effect of As2O3 was
assessed. The blank control contained no cells in the culture
medium, while the negative control contained medium with no
reagent.
Analyzing the expression of P-gp using
western blotting
The SGC7901/ADM and SGC7901/S cells
(1×104 cells/well) were grouped, as described above, and
were pretreated with non- and mildly-toxic concentrations of
As2O3 for 24, 48 and 72 h (18). The cells were collected by
centrifugation at 1,000 × g for 20 min at 4°C and washed with
ice-cold PBS. The cell pellets were then resuspended in lysis
buffer (Invitrogen Life Technologies, Carlsbad, CA, USA),
containing 50 mM Tris-chloride (pH 7.4), 150 mM sodium chloride,
0.1% sodium dodecyl sulfate, 1% Triton-100 and 1 mM
ethylenediaminetetraacetic acid (pH 8.0). Following incubation at
4°C for 40 min, the samples were centrifuged at 15,400 × g for 20
min. The supernatants containing cytosolic proteins were separated
by polyacrylamide gel (Bio-Rad Laboratories, Inc.) electrophoresis
at 200 V for 1–2 h, and were transferred onto a nitrocellulose
membrane (EMD Millipore, Billerica, MA, USA) at 120 V for 3 h. The
membrane was blocked with 5% skim milk powder at room temperature
for 2 h, followed by incubation with primary monoclonal mouse
anti-human antibody (P-gp, 1:50) for 2 h at room temperature.
Following washing with blocking buffer, the membranes were
incubated with HRP-conjugated goat-anti-mouse secondary antibody
(1:1,000) for 2 h at room temperature, followed by
chemiluminescence detection (Pierce Biotechnology, Inc., Rockford,
IL, USA). Western blot analysis was performed using an image
analysis system (VIDAS System; Biomérieux, Marcy l'Étoile,
France).
Immunocytochemical assays
Expression levels of Ras and p-ERK1/2
in cells treated with As2O3
A total of six treatment groups were used to assess
the levels of Ras and p-ERK1/2:
SGC7901/ADM+As2O3 (0.10, 0.50, and 0.75
µM), SGC7901/ADM+CsA (4.00 µg/ml), SGC7901/ADM and
SGC7901/S. The cells (1×104 cells/well) were treated, as
described above, and incubated for 24, 48, and 72 h. As previously
described (13), the cells were
fixed in ice-cold acetone for 15 min. Following incubating with 3%
hydrogen peroxide-carbinol for 10 min, the cells were incubated at
37°C for 2 h with the following primary antibodies:
Mouse-anti-human monoclonal Ras or p-ERK1/2. Following incubation
with Poly Helper (50 µl; Santa Cruz Biotechnology, Inc.,
Santa Cruz, CA, USA) at 37°C for 30 min, the HRP-conjugated
goat-anti-mouse IgG antibody solution was added and incubated again
at 37°C for 30 min. Finally, staining with 3,3′-diaminobenzidine
(Sigma-Aldrich) and hematoxylin (Sigma-Aldrich) were performed. The
labeled cells were analyzed using an image analysis system (VIDAS,
Option Company).
Expression levels of Ras and p-ERK1/2
following treatment with an ERK activator
rhG-CSF is a cytokine, which activates a number of
cellular pathways, including the ERK pathway (26,27).
Therefore, the SGC7901/ADM and SGC7901/S cells (1×104
cells/well) were treated with rhG-CSF (1.50 µg/ml) for 3
days at 37°C, and then divided into the five treatment groups,
described above. The cells were then incubated with the primary
antibody. The experiments were repeated three times.
Statistical analysis
Data are presented as the mean ± standard deviation
of three independent experiments. Student's t-test and analysis of
variance were used to analyze the differences between groups, with
a Student-Newman-Keuls (q-test) post hoc test. P<0.05 was
considered to indicate a statistically significant difference. All
data were analyzed using SPSS 17.0 (SPSS Inc., Chicago, IL,
USA).
Results
Sensitivity of SGC7901/ADM and SGC7901/S
cells to As2O3
In the present study, SGC7901/ADM and SGC7901/S
cells were treated with different concentrations of
As2O3 for 24, 48 and 72 h, following which
the OD at 490 nm was measured and the IC50 and survival
rates were calculated (Table I).
The IC50 values of the two cell types to
As2O3 decreased gradually with increasing
treatment duration. However, no difference was observed between the
two cell lines at any time-point (all P>0.05). Doses of 0.1 and
0.5 mM were considered non-toxic and mildly-toxic doses,
respectively (Table I), and
subsequent experiments were performed using these doses. These
results revealed that the SGC7901/ADM and SGC7901/S cells had the
same sensitivity to As2O3, which occurred in
a time- and dose-dependent manner.
 | Table IIC50 volume of non- and
mildly-toxic doses of arsenic trioxide at different
time-points. |
Table I
IC50 volume of non- and
mildly-toxic doses of arsenic trioxide at different
time-points.
| Cell line |
IC50µMa
| 95%
>SR>85%b
| SR>95%c
|
|---|
| 24 h | 48 h | 72 h | 24 h | 48 h | 72 h | 24 h | 48 h | 72 h |
|---|
| SGC7901/ADM | 5.57±0.36 | 2.63±0.18 | 1.42±0.09 | 0.10–0.08 | 0.10–0.75 | 0.05–0.25 | 0.01–0.01 | 0.01–0.01 | 0.01–0.05 |
| SGC7901/S | 5.94±0.54 | 3.06±0.28 | 1.59±0.12 | 0.25–1.30 | 0.07–0.08 | 0.04–0.25 | 0.01–0.25 | 0.01–0.07 | 0.01–0.04 |
As2O3 reduces drug
resistance of SGC7901/ADM cells to DOX, possibly through the
p-ERK1/2 pathway
The IC50 to DOX was assessed in the five
groups of cells incubated for 24 and 48 h. The experimental groups
were: SGC7901/ADM+0.10 µM As2O3 and
SGC7901/ADM+0.50 µM As2O3. The
positive control group was SGC7901/ADM+4.00 µg/ml CsA. The
negative control groups were SGC7901/ADM and SGC7901/S. The results
revealed that the level of drug resistance between the resistant
and parental cells were 6.63- and 6.85-fold at 24 h and 48 h,
respectively (Table II). The
IC50 values to DOX in the experimental groups were
significantly lower than that in the SGC7901/ADM group when treated
with 0.10 µM at 24 and 48 h (P<0.01) and with 0.50
µM at 24 and 48 h (P<0.01). The reversal in the CsA group
was ~4.5-fold at different time points, while in the experimental
groups, the effect was time-dependent (Table II). The IC50 of
As2O3 at 0.50 µM was markedly lower
than that at As2O3 at 0.10 µM,
particularly at 48 h. The value was close to that in the SGC7901/S
group and lower than that in the CsA group, but without statistical
significance (P>0.05).
| Table IIComparison of the IC50
values of DOX and drug-reversing ratio of
As2O3 at different time-points. |
Table II
Comparison of the IC50
values of DOX and drug-reversing ratio of
As2O3 at different time-points.
| Factor | Treatment | −rhG-CSF
| +rhG-CSF
|
|---|
| 24 h | 48 h | 48 h |
|---|
| IC50 to
DOXa |
As2O3 (0.1
µM) | 23.46±3.30 | 14.35±1.84 | 19.28±2.16 |
|
As2O3 (0.5
µM) | 9.52±1.02 | 4.28±0.55 | 6.85±1.31 |
| CsA (4
µg/ml) | 7.62±0.83 | 5.98±0.71 | 7.73±2.24 |
| SGC7901/ADM | 34.31±4.38 | 26.94±3.67 | 32.39±4.52 |
| SGC7901/S | 5.17±0.46 | 3.93±0.42 | 5.49±0.68 |
| Drug-resistance
ratiob | | 6.63 | 6.85 | 5.90 |
| Drug-reversal
ratioc |
As2O3 (0.1
µM) | 1.46 | 1.87 | 1.68 |
|
As2O3 (0.5
µM) | 3.60 | 6.29 | 4.72 |
| CsA (4
µg/ml) | 4.50 | 4.51 | 4.19 |
Following treatment with rhG-CSF, a cytokine that
activates the ERK pathway, the IC50 at 48 h was assessed
again (Table II). Compared with
the cells without rhG-CSF treatment, the IC50 to DOX of
As2O3 (0.50 µM) was significantly
higher (P<0.05), suggesting that the ERK activator had the
ability to partially reduce the drug resistance-reversing effect of
As2O3.
These findings suggested that the SGC7901/ADM cells
were resistant to DOX, and that As2O3 had a
drug resistance-reversing effect, in a time- and dose-dependent
manner. In addition, p-ERK1/2 may be involved in this mechanism,
which requires further investigation.
Reduction in the expression of P-gp
The expression of P-gp was detected using western
blotting (Fig. 1). The expression
of P-gp was more marked in the SGC7901/ADM cells than in the
SGC7901/S cells (0.38±0.025 vs. 0.19±0.012, respectively;
P<0.001), without significant variation in with time (Table III). In the SGC7901/ADM cells,
As2O3 at the non-toxic concentration (0.10
µM) at 24 h did not decrease the protein expression of P-gp,
however, the increase in duration significantly decreased the
levels of P-gp (P<0.01). Unlike the non-toxic concentration, the
mildly-toxic concentration (0.50 µM) reduced the levels of
P-gp even at 24 h (P<0.01). In the positive control cells, the
levels of P-gp were close to that in the SGC7901/S cells, without
significant changes with time (Table
III). These results revealed that the expression of P-gp in the
drug-resistant cells was higher than in the parental cells. CsA
appeared to reduce the protein expression of P-gp, and
As2O3 decreased the levels of P-gp, even at
non- or mildly-toxic concentrations, in a time- and dose-dependent
manner.
| Table IIIComparison of the OD values of P-gp,
measured using western blot analysis, and the expression levels of
Ras and p-ERK1/2, measured using immunocytochemistry, at different
time-points. |
Table III
Comparison of the OD values of P-gp,
measured using western blot analysis, and the expression levels of
Ras and p-ERK1/2, measured using immunocytochemistry, at different
time-points.
| Protein | Treatment | OD value
|
|---|
| 24 h | 48 h | 72 h |
|---|
| P-gp |
As2O3 (0.1
µM) | 0.349±0.001 | 0.32±0.003 | 0.29±0.002 |
|
As2O3 (0.5
µM) | 0.30±0.002 | 0.28±0.002 | 0.16±0.001 |
| CsA (4
µg/ml) | 0.16±0.004 | 0.16±0.001 | 0.15±0.002 |
| SGC7901/ADM | 0.32±0.061 | 0.38±0.010 | 0.38±0.005 |
| SGC7901/S | 0.14±0.003 | 0.14±0.003 | 0.14±0.002 |
| Ras |
As2O3 (0.1
µM) | 0.091±0.004 | 0.084±0.001 | 0.080±0.002 |
|
As2O3 (0.5
µM) | 0.089±0.003 | 0.078±0.003 | 0.076±0.003 |
|
As2O3 (0.75
µM) | 0.081±0.004 | 0.073±0.002 | 0.073±0.002 |
| CsA (4
µg/ml) | 0.091±0.003 | 0.093±0.003 | 0.084±0.005 |
| SGC7901/ADM | 0.093±0.003 | 0.090±0.003 | 0.091±0.004 |
| SGC7901/S | 0.068±0.003 | 0.071±0.005 | 0.066±0.008 |
| p-ERK1/2 | As2O3
(0.1µM) | 0.099±0.009 | 0.086±0.002 | 0.083±0.004 |
|
As2O3 (0.5
µM) | 0.086±0.007 | 0.080±0.002 | 0.070±0.001 |
|
As2O3 (0.75
µM) | 0.08±0.004 | 0.078±0.005 | 0.067±0.002 |
| CsA (4
µg/ml) | 0.098±0.009 | 0.11±0.018 | 0.11±0.025 |
| SGC7901/ADM | 0.11±0.008 | 0.11±0.012 | 0.11±0.014 |
| SGC7901/S | 0.10±0.016 | 0.090±0.010 | 0.10±0.027 |
Expression of Ras and p-ERK1/2
expression, determined using immunocytochemistry
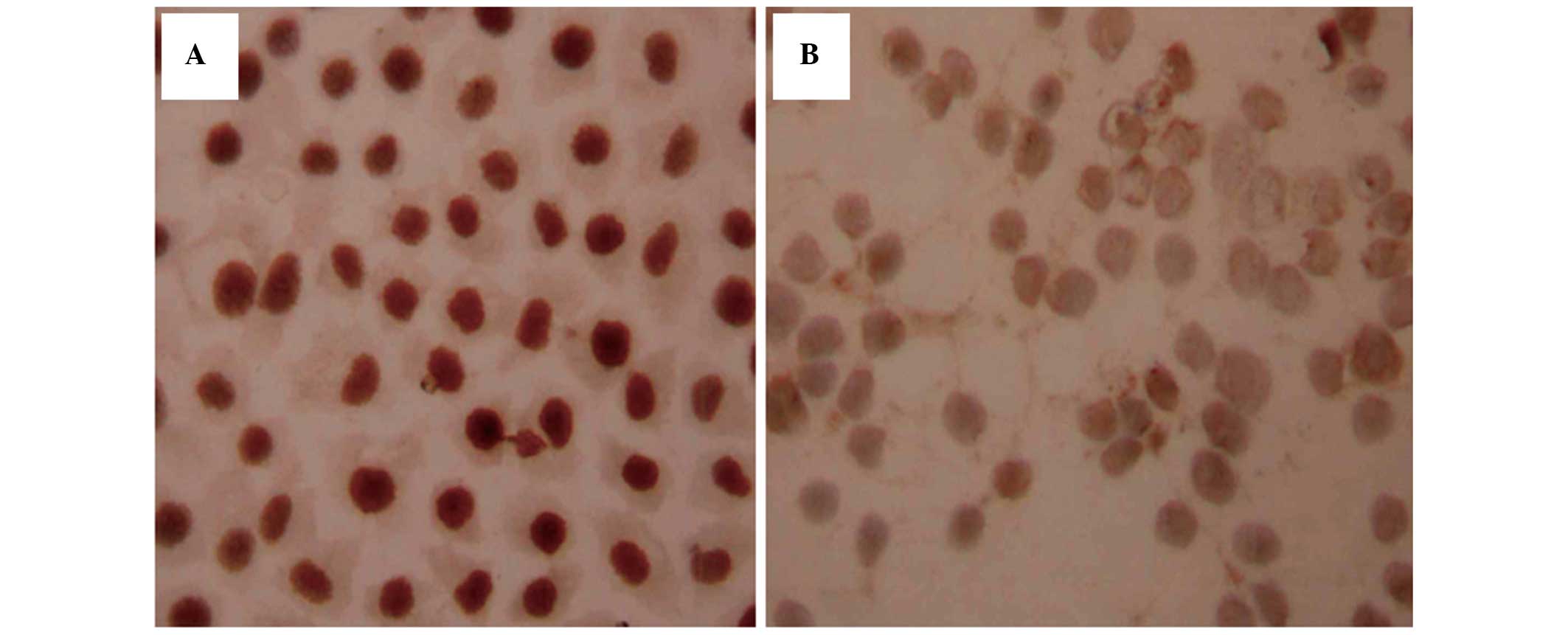
Ras protein was predominantly localized in the
membrane and partially translocated in the cytoplasm close to the
membrane (Fig. 2A). The expression
of Ras was higher in the SGC7901/ADM cells than in the SGC7901/S
cells (P<0.01). Following treatment with
As2O3, the levels of Ras were reduced in a
time- and dose-dependent manner (Table III; Fig. 2B). At 24 h, only the 0.75 µM
group exhibited lower expression than the SGC7901/ADM cells
(P<0.01). At 48 and 72 h, the cell groups treated with
As2O3 had significantly lower levels of Ras
(P<0.05). The expression of Ras in the three control groups
remained stable over time. These results suggested that
drug-resistant cells may have a higher expression levels of Ras
expression, and that As2O3 decreases these
levels.
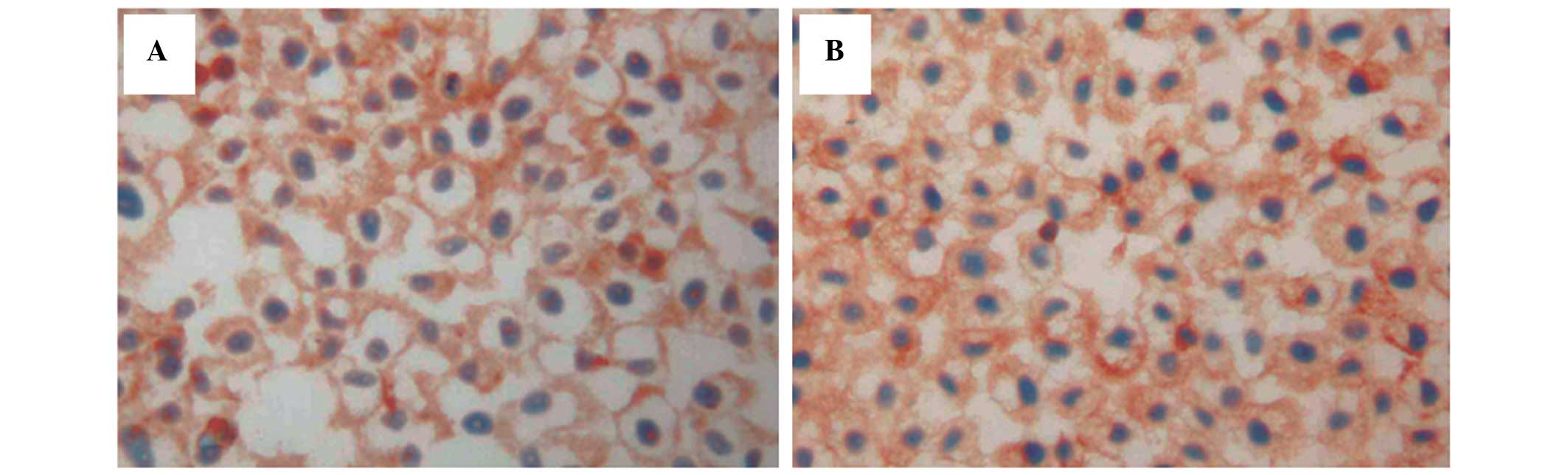
Expression of p-ERK1/2 expression was detected in
the cytoplasm surrounding the nucleus of cells (Fig. 3A and B). No significant differences
were identified in the levels of p-ERK1/2 between the SGC7901/ADM,
SGC7901/S and CsA-treated cells. Following treatment with
As2O3, the protein expression of p-ERK1/2
decreased (Table III). At 24 h,
the levels of p-ERK1/2 were decreased at all
As2O3 concentrations, without statistical
significance (P>0.05). However, in the 0.50 and 0.75 µM
As2O3 groups, the expression of p-ERK1/2 was
decreased, particularly in the As2O3 0.75
µM group at 72 h (P<0.01). These results suggested that
the Ras/p-ERK1/2 signaling pathway may be involved in the mechanism
of reducing drug resistance to DOX by As2O3
in GC cells.
Ras/p-ERK1/2 signaling pathway is
involved in reducing drug resistance
The cells were incubated with non- and mildly-toxic
concentrations of As2O3 for 48 h and,
following pretreatment with rhG-CSF, cytokines activating the ERK
pathway, Ras and p-ERK1/2 were assessed using
immunocytochemistry.
Following rhG-CSF treatment, the expression of Ras
was similar in all groups (P>0.05). No difference was identified
in the levels of Ras between the SGC7901/ADM and CsA-treated cells
(P>0.05; Table IV; Fig. 4A and B). Compared with the cells
without rhG-CSF treatment, the levels of Ras were not different
(P=0.12) in the 0.1 µM As2O3-treated
groups, but were higher in the As2O3 0.5
µM As2O3-treated groups
(P<0.01).
| Table IVProtein expression levels of Ras and
p-ERK1/2 with or without treatment of rhG-CSF, determined using
immunocytochemistry (optical density). |
Table IV
Protein expression levels of Ras and
p-ERK1/2 with or without treatment of rhG-CSF, determined using
immunocytochemistry (optical density).
| Protein | Treatment | rhG-CSF/− | rhG-CSF/+ |
|---|
| Ras b |
As2O3 (0.1
µM)b | 0.084±0.001 | 0.093±0.008 |
|
As2O3 (0.5
µM) | 0.078±0.003 | 0.089±0.001 |
| CsA (4
µg/ml) | 0.093±0.003 | 0.099±0.005 |
| SGC7901/ADM | 0.094±0.006 | 0.10±0.008 |
| p-ERK1/2a |
As2O3 (0.1
µM) | 0.086±0.002 | 0.093±0.002 |
|
As2O3 (0.5
µM) | 0.080±0.002 | 0.091±0.001 |
| CsA (4
µg/ml) | 0.11±0.018 | 0.12±0.019 |
| SGC7901/ADMc | 0.10±0.010 | 0.14±0.003 |
The levels of p-ERK1/2 in all the cells increased
following rhG-CSF treatment (Table
IV; P<0.01). Treatment with 0.10 and 0.50 µM
As2O3 reduced the levels of p-ERK1/2,
compared with the negative control group (P<0.05; Fig. 5A and B), whereas the levels of
p-ERK1/2 increased significantly at the same doses of
As2O3 following rhF-CSF treatment
(P<0.01). Similar to Ras, these results suggested that the
expression of p-ERK1/2 decreased following treatment with the ERK
activator.
Discussion
GC is one of the most common types of cancer, and
generally has a poor prognosis. DOX is used as one of the key
components in a number of chemotherapeutic regimens against GC,
however, MDR eventually limits its use. Several types of cancer
cell, including non-acute myeloid leukemia, multiple myeloma,
breast cancer and APL, have been found to be sensitive to
As2O3 through cell growth inhibition and
apoptosis (12,20,28,29).
However, few studies have examined the effects of
As2O3 in GC cells, and these studies reported
a number of different mechanisms (30–32).
In the present study, the human GC cell lines, SGC7901/S and
SGC7901/ADM, were investigated. The latter was induced by long-term
continuous exposure to DOX in a step-wise increment of various
concentrations. The high IC50 of SGC7901/ADM, but not of
SGC7901/S, confirmed the resistance to DOX. The doses of 0.10 and
0.50 µM of As2O3 were defined as non-
and mildly-toxic concentrations, respectively. This was different
from the study by Zhao et al (22), which reported that 2 µM was
the non-toxic concentration. In the MTT assay for the detection of
cytotoxicity and sensitivity to As2O3, cell
growth was inhibited without any treatment at 72 h. This was
possibly the result of the toxic metabolic accumulation and
nutrient deficiency. Previously, it was found that
As2O3 induced APL cells to enter apoptosis
(6). The present study
demonstrated that As2O3 partially reduced
in vitro drug-resistance, even at a non-toxic dose (0.10
µM), in a time- and dose-dependent manner.
A number of mechanisms are involved in MDR, and the
ovexpression of P-gp is one of the key mechanisms. Agents to reduce
MDR have been investigated for a number of years (33,34).
A novel inhibitor of P-gp-mediated MDR, OC144-093, was reported in
2000 (28). In the present study,
the expression of P-gp in SGC7901/ADM was higher than in the
parental cells and, following As2O3
treatment, the levels of P-gp decreased significantly. The non- and
mildly-toxic doses of As2O3 were selected
with the purpose of using them in clinical practice without serious
side effects.
Several studies have reported the in vitro
effects of As2O3 in the inhibition of
different types of solid tumor, through a number of different
mechanisms. Eguchi et al (19) demonstrated that apoptosis induced
in human mesothelioma cells is accompanied by the activation of
JNK1/2 and ERK1/2 (19). In breast
cancer, As2O3 inhibits cell growth through
the inactivation of the Notch signaling pathway (20). To investigate the role of the
Ras/p-ERK1/2 signaling pathway in the mechanism of
As2O3, the rhG-CSF cytokine, which activates
the ERK pathway was used to treat the cells.
Ras is a monomeric GTP-coupled protein encoded by
the Ras gene, which is important in cell growth regulation
(35). It was reported in 1993
that Ras has direct interactions with the RAF1 serine/threonine
kinase, which was the first mammalian Ras effector to be identified
(36). ERK1 and ERK2, also termed
mitogen-activated protein kinase (MAPK)1 and MAPK2 are MAPK
isomers, which are widely expressed in eukaryotic cells. In
addition, the identification of B-Raf mutations in cancer emphasize
the importance for aberrant Raf-MEK-ERK signaling in oncogenesis
(36). In the present study, the
levels of Ras and p-ERK were determined using immunocytochemistry.
The levels of Ras were higher in the SGC7901/ADM cells than in the
SGC7901/S cells, however, no significant difference was observed in
the levels of p-ERK1/2. The results suggested that p-ERK1/2 was not
involved in the MDR mechanisms in these cells, but indicated that
Ras was involved. Following treatment with
As2O3, the levels of Ras levels reduced, and
the protein levels of p-ERK1/2 decreased until intervention with
mildly-toxic concentrations of As2O3 (0.50
and 0.75 µM).
Following pretreatment with rhG-CSF, no change was
observed in the levels of Ras in cells from the control groups,
however, levels increased in the 0.50 µM
As2O3 treatment group. The expression of
p-ERK1/2 increased in all cells, suggesting that rhG-CSF activated
p-ERK1/2. As2O3 reduced the levels of
p-ERK1/2, compared with the negative control group. Thus, a p-ERK
activator may partially inhibit the drug resistance-reversing
effects of As2O3. A phase II trial revealed
that As2O3 is inactive in patients with
pancreatic cancer, who develop a progressive disease following
gemcitabine treatment (37). This
failure may be due to the lack of co-treatment with other
chemotherapeutics. As2O3 has been
demonstrated to be effective in treating APL without significant
side effects (8,9).
The present study involved the examination of MDR
induced by DOX. There are several other chemotherapeutic agents,
which are actively used in the treatment of GC in a number of
combination regimens (38).
Further investigations are required to examine the effect of
As2O3 in reversing the MDR induced by these
other agents. In addition, it is important to further investigate
the effects of As2O3 in GC, compared with
other types of cancer. Although the present study clearly suggested
the involvement of Ras in the MDR of GC, further investigations are
required to clarify whether mutations in Raf are involved in
drug-resistant cells. Finally, the rhG-CSF cytokine, used in the
present study to activate the ERK pathway, also activates a number
of other cell mechanisms, which require further investigations,
including the PI3K/AKT pathway (39,40).
In conclusion, the results of the present study
revealed that As2O3 had the ability to
reverse MDR in human GC cells. This mechanism may be relevant to
reduce the expression of P-gp. Drug-resistant cells may have higher
expression levels of Ras, and As2O3 may
decrease these levels. The Ras/p-ERK1/2 signal transduction pathway
may be involved in this mechanism. Further investigations,
involving a combination of chemotherapeutics and arsenic trioxide,
are essential.
Acknowledgments
This study was supported by ths Qingdao Municipal
Science and Technology Commission (grant no. 09-1-3-75-jch). The
authors would like to thank Dr Jing Dong for his critical reading
of the manuscript, and the Central Laboratory of the Affiliated
Hospital of the Medical College Qingdao University for providing
cell lines.
References
|
1
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar
|
|
2
|
Matsuda A and Machii R: Trends in stomach
cancer mortality rates in Japan, USA, UK, France and Korea based on
the WHO mortality database. Jpn J Clin Oncol. 42:1542012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Global cancer statistics, 2002. CA Cancer J Clin. 55:74–108. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zheng LH, Bao YL, Wu Y, Yu CL, Meng X and
Li YX: Cantharidin reverses multidrug resistance of human hepatoma
HepG2/ADM cells via down-regulation of P-glycoprotein expression.
Cancer Lett. 272:102–109. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Volm M and Mattern J: Resistance
mechanisms and their regulation in lung cancer. Crit Rev Oncog.
7:227–244. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mervis J: Ancient remedy performs new
tricks. Science. 273:5781996. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhu J, Chen Z, Lallemand-Breitenbach V and
de Thé H: How acute promyelocytic leukaemia revived arsenic. Nat
Rev Cancer. 2:705–713. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang TD, Chen GQ, Wang ZG, Wang ZY, Chen
SJ and Chen Z: Arsenic trioxide, a therapeutic agent for APL.
Oncogene. 20:7146–7153. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lo-Coco F, Avvisati G, Vignetti M, Thiede
C, Orlando SM, Iacobelli S, Ferrara F, Fazi P, Cicconi L, Di Bona
E, et al: Retinoic acid and arsenic trioxide for acute
promyelocytic leukemia. N Engl J Med. 369:111–121. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Murgo AJ: Clinical trials of arsenic
trioxide in hematologic and solid tumors: Overview of the National
Cancer Institute Cooperative Research and Development Studies.
Oncologist. 6(Suppl 2): 22–28. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lu J, Chew EH and Holmgren A: Targeting
thioredoxin reductase is a basis for cancer therapy by arsenic
trioxide. Proc Natl Acad Sci USA. 104:12288–12293. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Munshi NC: Arsenic trioxide: An emerging
therapy for multiple myeloma. Oncologist. (6 Suppl 2): 17–21. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Maeda H, Hori S, Nishitoh H, Ichijo H,
Ogawa O, Kakehi Y and Kakizuka A: Tumor growth inhibition by
arsenic trioxide (As2O3) in the orthotopic metastasis model of
androgen-independent prostate cancer. Cancer Res. 61:5432–5440.
2001.PubMed/NCBI
|
|
14
|
Shen ZY, Zhang Y, Chen JY, Chen MH, Shen
J, Luo WH and Zeng Y: Intratumoral injection of arsenic to enhance
antitumor efficacy in human esophageal carcinoma cell xenografts.
Oncol Rep. 11:155–159. 2004.
|
|
15
|
Bornstein J, Sagi S, Haj A, Harroch J and
Fares F: Arsenic Trioxide inhibits the growth of human ovarian
carcinoma cell line. Gynecol Oncol. 99:726–729. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Woo SH, Park IC, Park MJ, Lee HC, Lee SJ,
Chun YJ, Lee SH, Hong SI and Rhee CH: Arsenic trioxide induces
apoptosis through a reactive oxygen species-dependent pathway and
loss of mitochondrial membrane potential in HeLa cells. Int J
Oncol. 21:57–63. 2002.PubMed/NCBI
|
|
17
|
Miller WH Jr, Schipper HM, Lee JS, Singer
J and Waxman S: Mechanisms of action of arsenic trioxide. Cancer
Res. 62:3893–3903. 2002.PubMed/NCBI
|
|
18
|
Pelicano H, Feng L, Zhou Y, Carew JS,
Hileman EO, Plunkett W, Keating MJ and Huang P: Inhibition of
mitochondrial respiration: a novel strategy to enhance drug-induced
apoptosis in human leukemia cells by a reactive oxygen
species-mediated mechanism. J Biol Chem. 278:37832–37839. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Eguchi R, Fujimori Y, Takeda H, Tabata C,
Ohta T, Kuribayashi K, Fukuoka K and Nakano T: Arsenic trioxide
induces apoptosis through JNK and ERK in human mesothelioma cells.
J Cell Physiol. 226:762–768. 2011. View Article : Google Scholar
|
|
20
|
Xia J, Li Y, Yang Q, Mei C, Chen Z, Bao B,
Ahmad A, Miele L, Sarkar FH and Wang Z: Arsenic trioxide inhibits
cell growth and induces apoptosis through inactivation of notch
signaling pathway in breast cancer. Int J Mol Sci. 13:9627–9641.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xiao Z, Ding N, Xiao G, Wang S, Wu Y and
Tang L: Reversal of multidrug resistance by gefitinib via RAF1/ERK
pathway in pancreatic cancer cell line. Anat Rec (Hoboken).
295:2122–2128. 2012. View Article : Google Scholar
|
|
22
|
Zhao D, Jiang Y, Dong X, Liu Z, Qu B,
Zhang Y, Ma N and Han Q: Arsenic trioxide reduces drug resistance
to adriamycin in leukemic K562/A02 cells via multiple mechanisms.
Biomed Pharmacother. 65:354–358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhao Y, You H, Liu F, An H, Shi Y, Yu Q
and Fan D: Differentially expressed gene profiles between multidrug
resistant gastric adenocarcinoma cells and their parental cells.
Cancer Lett. 185:211–218. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Che XF, Nakajima Y, Sumizawa T, Ikeda R,
Ren XQ, Zheng CL, Mukai M, Furukawa T, Haraguchi M, Gao H, et al:
Reversal of P-glycoprotein mediated multidrug resistance by a newly
synthesized 1,4-benzothiazipine derivative, JTV-519. Cancer Lett.
187:111–119. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang R, Xia L, Gabrilove J, Waxman S and
Jing Y: Downregulation of Mcl-1 through GSK-3β activation
contributes to arsenic trioxide-induced apoptosis in acute myeloid
leukemia cells. Leukemia. 27:315–324. 2013. View Article : Google Scholar
|
|
26
|
Brandstetter T, Ninci E, Falken U, Wagner
E, Hess R and Bauknecht T: rhG-CSF affects genes involved in
mitogen signalling and early gene expression in the ovarian cancer
cell line HEY. Int J Cancer. 75:847–854. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ricciardi MR, McQueen T, Chism D, Milella
M, Estey E, Kaldjian E, Sebolt-Leopold J, Konopleva M and Andreeff
M: Quantitative single cell determination of ERK phosphorylation
and regulation in relapsed and refractory primary acute myeloid
leukemia. Leukemia. 19:1543–1549. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Newman MJ, Rodarte JC, Benbatoul KD,
Romano SJ, Zhang C, Krane S, Moran EJ, Uyeda RT, Dixon R, Guns ES,
et al: Discovery and characterization of OC144-093, a novel
inhibitor of P-glycoprotein-mediated multidrug resistance. Cancer
Res. 60:2964–2972. 2000.PubMed/NCBI
|
|
29
|
Li Y, Qu X, Qu J, Zhang Y, Liu J, Teng Y,
Hu X, Hou K and Liu Y: Arsenic trioxide induces apoptosis and G2/M
phase arrest by inducing Cbl to inhibit PI3K/Akt signaling and
thereby regulate p53 activation. Cancer Lett. 284:208–215. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chen X, Zhang M and Liu LX: The
overexpression of multidrug resistance-associated proteins and
gankyrin contribute to arsenic trioxide resistance in liver and
gastric cancer cells. Oncol Rep. 22:73–80. 2009.PubMed/NCBI
|
|
31
|
Liu Y, Zhang W, Zhang X, Qi Y, Huang D and
Zhang Y: Arsenic trioxide inhibits invasion/migration in SGC-7901
cells by activating the reactive oxygen species-dependent
cyclo-oxygenase-2/matrix metalloproteinase-2 pathway. Exp Biol Med
(Maywood). 236:592–597. 2011. View Article : Google Scholar
|
|
32
|
Zhang G, Liu J, Zhang Y, Qu J, Xu L, Zheng
H, Liu Y and Qu X: Cbl-b-dependent degradation of FLIP(L) is
involved in ATO-induced autophagy in leukemic K562 and gastric
cancer cells. FEBS Lett. 586:3104–3110. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Baguley BC: Multiple drug resistance
mechanisms in cancer. Mol Biotechnol. 46:308–316. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gillet JP and Gottesman MM: Mechanisms of
multidrug resistance in cancer. Methods Mol Biol. 596:47–76. 2010.
View Article : Google Scholar
|
|
35
|
Wong KA, Russo A, Wang X, Chen YJ, Lavie A
and O'Bryan JP: A new dimension to Ras function: A novel role for
nucleotide-free Ras in Class II phosphatidylinositol 3-kinase beta
(PI3KC2β) regulation. PLoS One. 7:e453602012. View Article : Google Scholar
|
|
36
|
Karnoub AE and Weinberg RA: Ras oncogenes:
Split personalities. Nat Rev Mol Cell Biol. 9:517–531. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kindler HL, Aklilu M, Nattam S and Vokes
EE: Arsenic trioxide in patients with adenocarcinoma of the
pancreas refractory to gemcitabine: A phase II trial of the
University of Chicago Phase II Consortium. Am J Clin Oncol.
31:553–556. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
NCCN Clinical Practice Guidelines in
Oncology (NCCN Guidelines)Gastric Cancer. 3rd edition. National
Comprehensive Cancer Network; Fort Washington: 2015
|
|
39
|
Guo H, Sun F, Huang W, Liu Z, Zhang S,
Zhou Q and Liang C: The effect of rhG-CSF on spleen transcriptome
in mouse leukopenia model induced by cyclophosphamide.
Immunopharmacol Immunotoxicol. 36:114–123. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kumar J, Fraser FW, Riley C, Ahmed N,
McCulloch DR and Ward AC: Granulocyte colony-stimulating factor
receptor signalling via Janus kinase 2/signal transducer and
activator of transcription 3 in ovarian cancer. Br J Cancer.
110:133–145. 2014. View Article : Google Scholar :
|