Introduction
Due to its malignant biological characteristics,
including invasion and metastasis, patients with lung cancer
usually have a poor prognosis, and in 2012, 1.59 million succumbed
to lung cancer worldwide (1).
Therapies to combat the growth and metastasis of lung cancers are
urgently required. By elucidating the genetic and molecular
mechanisms underlying cancer cell proliferation and metastasis,
targets for anticancer therapy may be revealed.
Eukaryotic translation initiation factor 4E (eIF4E)
has an important role in the regulation of eukaryotic translation
initiation (2), and recognizes the
5′-cap of mRNA. The 5′-cap of mRNA has been reported to be
associated with mRNA transcription and translation, preventing the
degradation of mRNA precursors, and serving as a binding site for
regulatory factors, therefore, being involved in various RNA
metabolic processes (3). By
binding to the 5′-cap of mRNA, eIF4E forms a translation initiation
complex involved in protein trafficking and translation, thereby
regulating gene expression (4,5).
Previous studies have reported that eIF4E is
upregulated in various human malignant tumors, and that eIF4E
expression is closely associated with tumorigenesis, infiltration
and metastasis in solid tumors, including lung, breast and prostate
cancer (6–10). The overexpression of eIF4E is also
negatively correlated with survival in advanced cancer types,
including head and neck, breast, prostate, lung, and hematologic
malignancies (6,8–12).
In addition, eIF4E phosphorylation has been reported to be
specifically increased in advanced malignancies (7,12,13).
Our previous study demonstrated that the expression levels of eIF4E
were increased in the tumor, compared with adjacent tissue samples,
and its elevated expression was associated with lymph node
metastasis (13). Therefore, this
led to the hypothesis that eIF4E downregulation may lead to
biological function inhibition in lung cancer cells.
The present study sought to further elucidate the
mechanisms underlying the effects of eIF4E on cell proliferation,
apoptosis, invasion and the cell cycle. The expression of eIF4E was
inhibited in A549 lung adenocarcinoma cells using RNA interference,
and the impact of eIF4E knockdown on the rate of apoptosis, as well
as the capacity of these cells to proliferate, migrate and progress
through the cell cycle, was subsequently investigated in
vitro.
Materials and methods
Materials
DNA restriction enzymes (BbsI, PstI
and BamHI) and DNA ligase were purchased from Thermo Fisher
Scientific, Inc., (Waltham, MA, USA). A DNA gel purification kit
was purchased from Tiangen Biotech Co., Ltd. (Beijing, China), a
Plasmid Extraction kit was purchased from Hangzhou Axygen
Biotechnology, Ltd. (Hangzhou, China), and the Quantitative
fluorescent Polymerase Chain Reaction (PCR) kit was purchased from
Shanghai Bio-Tech Co., Ltd (Shanghai, China). Rabbit anti-eIF4E was
purchased from Abcam (Cambridge, MA, USA). DNA primers were
synthesized by GenePharma Co., Ltd. (Shanghai, China). A Gibco Cell
Apoptosis kit was purchased from Thermo Fisher Scientific, Inc.,
and the Cell Cycle kit from Multi Sciences (Lianke) Biotech Co.,
Ltd. (Hangzhou, China). The A549 lung adenocarcinoma cell line was
obtained from the Cell Bank of the Chinese Academy of Science
(Shanghai, China). The pGPU6/green fluorescent protein (GFP)/Neo
plasmid was obtained from GenePharma Co., Ltd.
Construction of an eIF4E-short hairpin
(sh)RNA plasmid
A total of three sets of primers for eIF4E-directed
shRNA and one set of primers for scrambled sequence shRNA were
designed by Oligo Designer 3.0 (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA), and synthesized by GenePharma Co., Ltd.
(Table I). The template contained
a TTCAAGAGA loop sequence, a T6 transcription termination sequence,
CACC was added to the 5′-end of the sense template to yield a
sticky end following BbsI digestion, and GATC was added to
the 5′-end of the anti-sense template to yield a complementary
sticky end following BamHI digestion. The four
oligonucleotides were ligated into a pGPU6/GFP/Neo plasmid
following annealing and were termed eIF4E-shRNA1, eIF4E-shRNA2,
eIF4E-shRNA3 and NC-shRNA. All plasmid sequences were confirmed by
restriction enzyme digestion and sequencing (Thermo Fisher
Scientific, Inc.).
 | Table IshRNA sequences used in the present
study. |
Table I
shRNA sequences used in the present
study.
| shRNA | Target
location | Target
sequence | shRNA synthetic
sequences |
|---|
| eIF4E-shRNA1 | 2094 |
CCAAAGATAGTGATTGGTTAT | Sense:
5′-CACCGCCAAAGATAGTGATTGGTTATTTCAAGAGAATAACCAATCACTATCTTTGGTTTTTTG-3′
Antisense:
5′-GATCCAAAAAACCAAAGATAGTGATTGGTTATTCTCTTGAAATAACCAATCACTATCTTTGGC-3′ |
| eIF4E-shRNA2 | 1849 |
GGAGGACGATGGCTAATTACA | Sense:
5′-CACCGGAGGACGATGGCTAATTACATTCAAGAGATGTAATTAGCCATCGTCCTCCTTTTTTG-3′
Antisense:
5′-GATCCAAAAAAGGAGGACGATGGCTAATTACATCTCTTGAATGTAATTAGCCATCGTCCTCC-3′ |
| eIF4E-shRNA3 | 1970 |
GTGGCGCTGTTGTTAATGTTA | Sense:
5′-CACCGTGGCGCTGTTGTTAATGTTATTCAAGAGATAACATTAACAACAGCGCCACTTTTTTG-3′
Antisense:
5′-GATCCAAAAAAGTGGCGCTGTTGTTAATGTTATCTCTTGAATAACATTAACAACAGCGCCAC-3′ |
| NC-shRNA | | | Sense:
5′-CACCGTTCTCCGAACGTGTCACGTCAAGAGATTACGTGACACGTTCGG AGA
ATTTTTTG-3′
Antisense:
5′-GATCCAAAAAATTCTCCGAACGTGTCACGTAATCTCTTGACGTGACACGTTCGGAGAAC-3′ |
Cell culture and transfection
A549 lung adenocarcinoma cells were cultured in
RPMI-1640 media (HyClone Laboratories; GE Healthcare Life Sciences,
Logan, UT, USA), supplemented with 10% fetal bovine serum (FBS;
HyClone Laboratories; GE Healthcare Life Sciences), at 37°C in an
atmosphere containing 5% CO2. The cells were washed with
phosphate-buffered saline (PBS) and 0.25% trypsin digestion (Gibco;
Thermo Fisher Scientific, Inc.) for passaging. The cells were
transfected with Invitrogen Lipofectamine® 2000 (Thermo
Fisher Scientific, Inc.) alone (negative control), or one of the
three eIF4E-targeting shRNA: eIF4E-shRNA1, eIF4E-shRNA2 or
eIF4E-shRNA3, according to the manufacturer's protocol. Transfected
cells were selected by addition of G418 (Amresco LLC, Solon, OH,
USA) to the culture media.
Measurement of the mRNA expression of
eIF4E by reverse transcription-quantitative polymerase chain
reaction (RT-qPCR)
The total RNA was extracted from the A549 cells
using TRIzol® reagent (Thermo Fisher Scientific, Inc.),
according to the manufacturer's protocol. The cDNA was reverse
transcribed, according to the Fermentas M-MLV reverse transcriptase
protocol (Thermo Fisher Scientific, Inc.). The primer sequences
were as follows: eIF4E, forward: 5′-ACGGAATCTAATCAGGAC-3′ and
reverse: 5′-CCCACATAGGCAATAC-3′; β-actin, forward:
5′-AAGATGACCCAGATCATGTTTGA-3′ and reverse:
5′-TTAATGTCACGCACGATTTCC-3′. The product length was 287 bp. PCR was
conducted using a CFX Connect Real-Time PCR system (Bio-Rad
Laboratories, Inc.). The PCR cycling conditions were as follows: 35
cycles of 94°C for 5 min, 94°C for 10 sec and 60°C for 30 sec, and
60°C for 10 min. The expression levels were measured by
SYBR® Green (Applied Biosystems; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol.
Measurement of the protein expression of
eIF4E by western blotting
The cells (1×107 cells/ml) were lysed
using lysis buffer (pH 7.4, 50 mM Tris-base, 150 mM NaCl, 1 mM
ethylenediaminetetraacetic acid, 1% Triton X-100, 1 mM
phenylmethylsulfonyl fluoride). The homogenate was subsequently
centrifuged at 12,000 × g for 15 min at 4°C, and the supernatant
was retained and preserved at −80°C until further use. A total of
30 µg protein was separated by 10% SDS-PAGE (Beyotime
Institute of Biotechnology, Haimen, China) according to the
manufacturer's instructions and was subsequently transferred onto
nitrocellulose membranes (EMD Millipore, Billerica, MA, USA) prior
to blocking with 5% non-fat milk at room temperature for 2 h. The
membranes were subsequently incubated with a rabbit anti-human
eIF4E primary antibody (1:1,000; cat. no. AB1126; Abcam) overnight
at 4°C, and were washed three times with Tris-buffered saline
containing 0.1% Tween-20 (Amresco LLC). The membranes were then
incubated with a horseradish peroxidase-conjugated secondary
antibody (1:1,000; cat. no. BS13278; Bioworld Technology, Inc., St.
Louis Park, MN, USA) for 2 h at room temperature. The protein bands
were detected using an Enhanced Chemiluminescence kit (Beyotime
Institute of Biotechnology) and X-ray exposure (Beyotime Institute
of Biotechnology). Densitometry was quantified by Quantity One
software (ChemiDoc XRS+ system; Bio-Rad Laboratories, Inc.) and the
eIF4E/GAPDH expression level ratio was calculated.
Measurement of cell viability using a
Cell Counting kit-8 assay
A549 cells in the logarithmic phase were resuspended
and seeded into a 96-well plate at 2×103 cells/well and
incubated at 37°C in an atmosphere containing 5% CO2.
After 24 h, the cells were transfected, as previously described
(14). The transfected cells were
cultured and after 48 and 72 h, cell viability was assessed using a
Cell Counting kit-8 (Beyotime Institute of Biotechnology), and
calculated using the following formula: Viability (%) =
(Aexperimental / Acontrol) ×100%.
Measurement of apoptosis using annexin V
staining
The transfected A549 cells in the logarithmic phase
were resuspended and seeded into a 6-well plate at 2×105
cells/well. After 24 h, serum-free RPMI-1640 media was added and
following a further 24, 48 or 72 h incubation, the cells were
resuspended with trypsin and centrifuged at 400 x g for 5 min at
room temperature. The cell pellet was washed twice with cold PBS
and resuspended in 100 µl 1X Binding Buffer (Taizhou
Hospital Laboratory, Linhai, China) to which 5 µl
fluorescein isothiocyanate-conjugated annexin V antibody and 1
µl propidium iodide (Gibco; Thermo Fisher Scientific, Inc.)
was added. The mixture was incubated in the dark for 15 min at 4°C
and cell apoptosis was examined by flow cytometry (Accuri C6; BD
Biosciences, San Jose, CA, USA).
Assessment of cell cycle distribution by
flow cytometry
Transfected A549 cells in the logarithmic phase were
resuspended and seeded into a 6-well plate at 2×105
cells/well. After 24 h, serum-free RPMI-1640 media was added.
Following a further 24, 48 or 72 h incubation, the cells were
resuspended in trypsin and centrifuged at 400 × g for 5 min at room
temperature. The cell pellet was resuspended and fixed in 75%
ethanol at 4°C for 24 h. The sample was subsequently centrifuged at
400 x g for 5 min at room temperature and the cell pellet was air
dried. The dry pellet was washed twice with PBS and resuspended in
1 ml reagent A [Multi Sciences (Lianke) Biotech Co., Ltd.,
Hangzhou, China], and vortexed for 5–10 sec. After 30 min, the cell
cycle was examined by flow cytometry (Accuri C6; BD
Biosciences).
Assessment of cell invasion using a
Transwell assay
Matrigel (BD Biosciences) dissolved at 4°C was
diluted in cold PBS. A total of 100 µl diluted gel (~25
µg) was added to the upper chamber of each well of a 24-well
Transwell plate. The plate was incubated overnight at 37°C for
basal lamina coating and washed with serum-free RPMI-1640 media.
The transfected cells were resuspended in RPMI-1640, supplemented
with 1% FBS at 5×104/ml, and a 200 µl suspension
was added into the upper chamber and 600 µl culture medium
was added to the lower chamber. The plate was incubated for 24 h at
37°C in an atmosphere containing 5% CO2. Non-migrating
cells in the upper chamber were removed with a cotton swab. The
Transwells were removed and the plate was inverted and air dried. A
total of 500 µl 0.1% crystal violet solution (Sigma-Aldrich,
St. Louis, MO, USA) was added to each well for 30 min at 37°C and
the Transwell plate was subsequently washed with PBS. Four fields
were selected for cell counting.
Statistical analysis
The data were presented as the mean ± standard
deviation. Frequencies were compared by Student's t-test, and
comparisons between groups were made by one-way analysis of
variance. The data were analyzed using the SPSS 17.0 software
package (SPSS, Inc., Chicago, IL, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
Effects of shRNA transfection on the mRNA
and protein expression levels of eIF4E
A549 lung adenocarcinoma cells were transfected with
one of three eIF4E-targeting shRNAs or scrambled shRNA. RT-qPCR
revealed that the mRNA expression of eIF4E in the cells transfected
with scramble shRNA (0.905±0.068 relative to β-actin mRNA) revealed
no significant difference compared with those in the mock
transfected control cells (0.918±0.150). However, the relative mRNA
expression levels of eIF4E in cells transfected with eIF4E-shRNA1,
eIF4E-shRNA2 and eIF4E-shRNA3 were significantly reduced to
0.5±0.2, 0.4±0.2, 0.3±0.1, respectively (P<0.01; Table II).
| Table IImRNA and protein expression levels of
eIF4E in transfected cells. |
Table II
mRNA and protein expression levels of
eIF4E in transfected cells.
| Treatment | Relative expression
levels
|
|---|
| mRNA | Protein |
|---|
| Mock | 0.918±0.150 | 0.857±0.069 |
| NC | 0.905±0.068 | 0.829±0.102 |
| eIF4E-shRNA1 |
0.541±0.092ab |
0.495±0.093ab |
| eIF4E-shRNA2 |
0.428±0.137ab |
0.339±0.080ab |
| eIF4E-shRNA3 |
0.361±0.083ab |
0.254±0.086ab |
Western blotting revealed that the reduction in mRNA
eIF4E expression levels corresponded to a significant reduction in
the protein expression levels of eIF4E in cells transfected with
all three eIF4E-targeting shRNAs (P<0.01; Table II). eIF4E-shRNA3 reduced the eIF4E
protein expression levels most significantly relative to those of
GAPDH, from 0.857±0.069 in the mock transfected cells, to
0.254±0.086 (P<0.01; Table
II). Therefore, eIF4E-shRNA3 was used for further
experimentation.
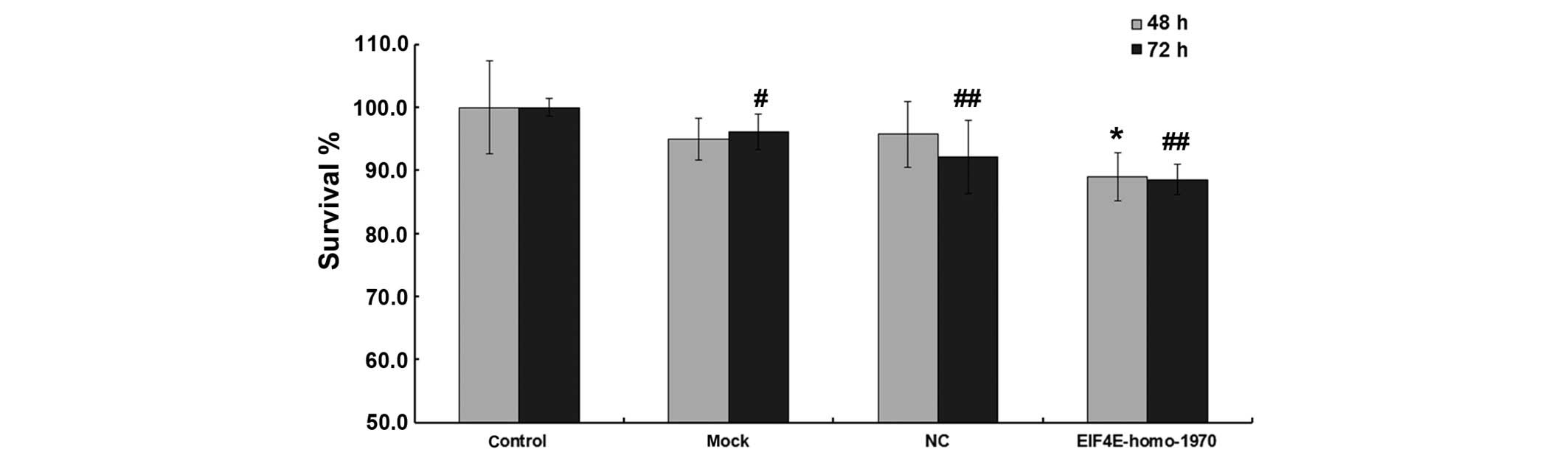
Effects of eIF4E-shRNA on cell
viability
Although mock or scrambled shRNA transfection caused
no impact on cell viability, the viability of
eIF4E-shRNA-transfected cells was significantly lower compared with
that of mock transfected cells after 48 and 72 h (48 h
post-transfection, 94.9±3.3%, 89.0±3.8%, P<0.05; 72 h
post-transfection, 96.1±2.8%, 88.6±2.4%, P<0.05; Fig. 1).
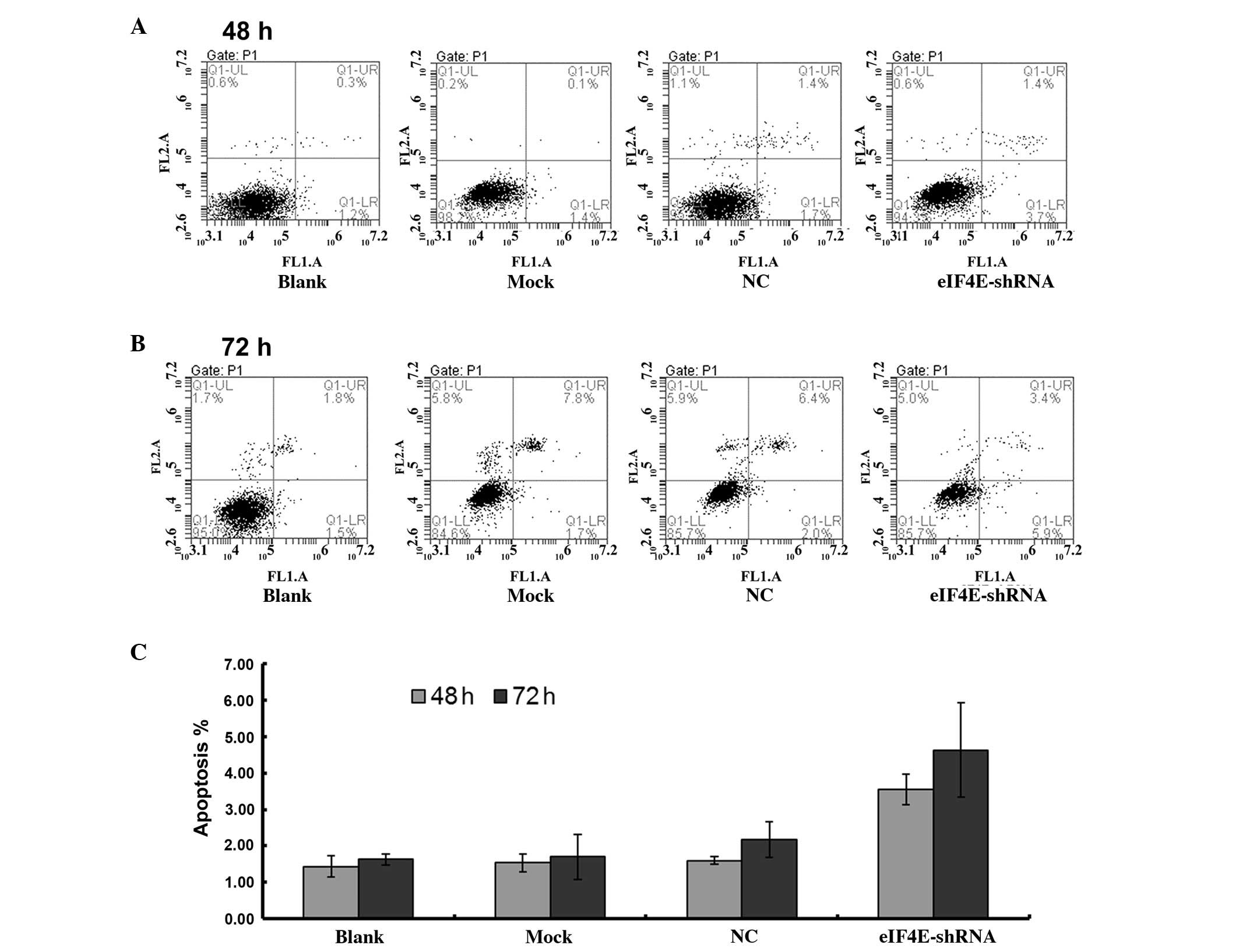
Transfection of scrambled shRNA significantly
increased the rate of apoptosis from 1.43±0.29% in the untreated
cells and 1.53±0.25% in the mock transfected cells to 1.60±0.10% 48
h post-transfection with scrambled shRNA. However, transfection
with eIF4E-shRNA induced a significantly higher rate of apoptosis
(3.57±0.42% after 48 h and 4.63±1.30 after 72 h; P<0.05;
Fig. 2).
Effects of eIF4E-shRNA on the cell
cycle
Although mock and scrambled shRNA transfection
caused no impact on the cell cycle, the fraction of eIF4E-shRNA
transfected cells in the G0/G1 phase was
significantly increased, whereas the percentage of cells in the S
phase was reduced, indicating G0/G1 arrest.
The fraction of cells in the G0/G1 phase
increased from 49.08±2.57 in the scrambled shRNA-transfected cells
to 57.14±0.59 after 48 h. This changed from 64.87±2.45 before to
73.95±6.00 after 72 h (P<0.05; Table III). The fraction of cells in the
S phase decreased from 45.81±1.32 in the scrambled
shRNA-transfected cells to 23.81±0.83 after 48 h, and from
25.75±2.07 to 14.29±1.75 after 72 h (P<0.05, Table III).
| Table IIICell cycle distribution following
transfection. |
Table III
Cell cycle distribution following
transfection.
| Treatment | 48 h
post-transfection
| 72 h
post-transfection
|
|---|
|
G0/G1 (%) | S (%) | G2/M
(%) |
G0/G1 (%) | S (%) | G2/M
(%) |
|---|
| Untreated | 46.88±1.67 | 38.06±2.10 | 15.06±1.18 | 63.10±2.03 | 28.29±2.83 | 8.61±1.21 |
| Mock | 48.84±1.64 | 41.64±1.57 | 9.52±1.92 | 63.33±1.28 | 27.98±1.09 | 8.69±2.34 |
| NC | 49.08±2.57 | 45.81±1.32 | 5.11±0.96 | 64.87±2.45 | 25.75±2.07 | 9.38±1.70 |
| eIF4E-shRNA | 57.14±0.59a | 23.81±0.83a | 19.05±1.78 | 73.95±6.00a | 14.29±1.75a | 11.76±2.74 |
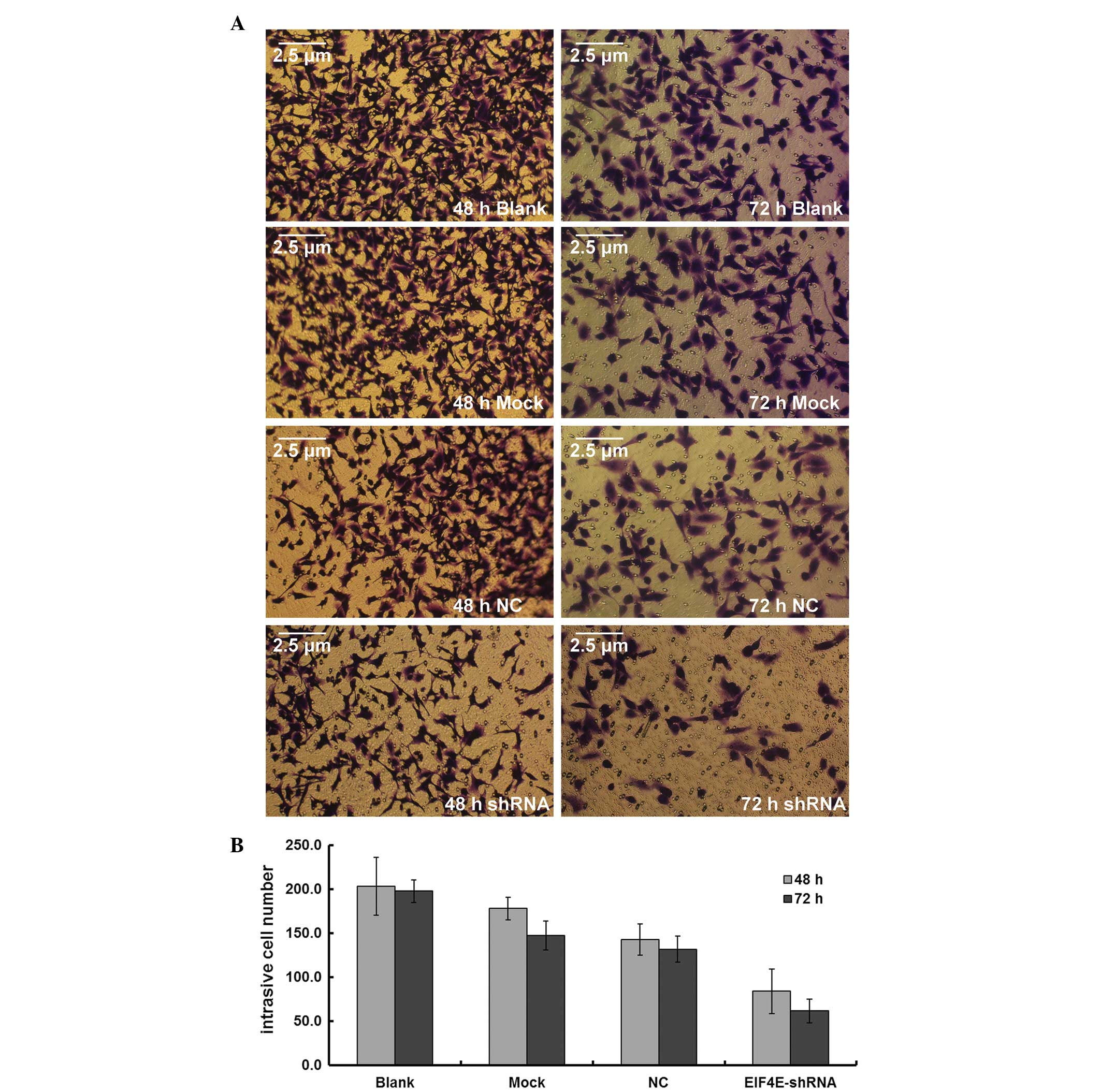
Effects of eIF4E-shRNA on cell
invasion
Although mock and scrambled shRNA transfection
revealed no impact on the number of cells transversing a Transwell
insert, the fraction of eIF4E-shRNA-transfected cells migrating was
significantly lower. The fraction of cells migrating decreased from
182.5±14.0 in the scrambled shRNA-transfected cells to 83.8±25.4 in
the eIF4E-shRNA-transfected cells after 48 h, and from 167.5±30.4
to 61.5±13.5 after 72 h (P<0.01; Fig. 3).
Discussion
Due to propensity of lung cancer to invade and
metastasize, patients with lung cancer usually have a poor
prognosis (1,15). Lung cancer is a major cause of
mortality worldwide (1), and
therefore, investigations into the genetic contributors to the cell
proliferation and metastasis of lung cancer are urgently
required.
Previous studies reported that the expression of
eIF4E was upregulated in malignant human tumors (2,6). The
present study aimed to further elucidate the mechanisms underlying
the effects of eIF4E on cell proliferation, apoptosis, invasion and
cell cycle. Several constructs were successfully cloned to harbor
eIF4E shRNAs targeting various eIF4E domains. Transfection of A549
cells with eIF4E-targeting shRNA reduced the mRNA and protein
expression levels of eIF4E by >70%.
A549 cells transfected with eIF4E-targeting shRNA
were significantly less viable compared with A549 cells transfected
with scrambled shRNA. The rate of apoptosis was also significantly
increased, and significantly more cells were in the
G0/G1 phase and fewer were in the S phase,
indicating cell cycle arrest. These observations supported previous
studies that demonstrated that eIF4E selectively enhanced the
translation and nucleocytoplasmic transport of mRNAs containing
long, highly structured untranslated regions, generally encoding
proteins involved in the cell growth (cyclin D1, c-myc),
angiogenesis [vascular endothelial growth factor (VEGF), fibroblast
growth factor-2], invasion [matrix metalloproteinase (MMP)-9,
heparanase], and survival (survivin, B cell lymphoma 2) (6,11,16–19).
The fraction of transfected cells migrating across the Transwell
inserts was also reduced, likely due to the regulation of cell
migration-associated genes, including Cyclin D1, VEGF, and MMP-9 by
eIF4E (6). These results suggested
that the overexpression of eIF4E contributes to the proliferative
phenotype of the A549 lung adenocarcinoma cell line, and that eIF4E
is crucial for cell cycle progression and invasion.
In vitro, eIF4E phosphorylation appears to be
important for the proliferation of numerous tumor cell lines
(20–22), and the upregulation of eIF4E
expression in transgenic mice increased the incidence of lymphoma,
lung adenocarcinoma, angiosarcoma and hepatoma (23,24).
Conversely, disruption of eIF4E phosphorylation inhibits tumor
development in mouse lymphoma and prostate cancer models (25,26).
As the activity levels of eIF4E are elevated in
cancer cells, cancer may be preferentially susceptible to
eIF4E-targeted therapy. Several eIF4E inhibitors have recently been
investigated for clinical use, including an eIF4E-targeting
antisense oligonucleotide (27,28),
and small molecular inhibitors of eIF4E phosphorylation or activity
(29,30), including mRNA cap structure binding
(31,32) and eIF4E:eIF4G binding (33).
In conclusion, the results of the present study
indicated that the inhibition of eIF4E may suppress A549 cell
growth and invasion, and induce apoptosis and cell cycle arrest
in vitro, demonstrating a critical role for eIF4E in lung
cancer cell growth. As numerous studies (34–36)
have now implicated eIF-4E overexpressison in a wide range of human
tumor types, and eIF-4E overexpression has been associated with
disease progression, strategies to decrease eIF4E expression may
represent a potential therapeutic technique in numerous cancer
types. These findings indicated that strategies to decrease eIF4E
expression or inhibit eIF4E function represent a promising strategy
for reducing the rate of lung adenocarcinoma proliferation and
metastasis.
Acknowledgments
The present study was supported by the Zhejiang
Medical and Health Science and Technology Plan Project (no.
2014KYB310) and the Taizhou Science and Technology Plan Project
(no. 11ky09).
References
|
1
|
WHO international agency for research on
cancer: Globocan 2012: Estimated cancer incidence, mortality and
prevalence worldwide in 2012. 2012.
|
|
2
|
Joshi B, Cameron A and Jagus R:
Characterization of mammalian eIF4E-family members. Eur J Biochem.
271:2189–2203. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ziemniak M, Strenkowska M, Kowalska J and
Jemielity J: Potential therapeutic applications of RNA cap analogs.
Future Med Chem. 5:1141–1172. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Culjkovic B, Topisirovic I and Borden KL:
Controlling gene expression through RNA regulons: The role of the
eukaryotic translation initiation factor eIF4E. Cell Cycle.
6:65–69. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jackson RJ, Hellen CU and Pestova TV: The
mechanism of eukaryotic translation initiation and principles of
its regulation. Nat Rev Mol Cell Biol. 11:113–127. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
De Benedetti A and Graff JR: eIF-4E
expression and its role in malignancies and metastases. Oncogene.
23:3189–3199. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yoshizawa A, Fukuoka J, Shimizu S, Shilo
K, Franks TJ, Hewitt SM, Fujii T, Cordon-Cardo C, Jen J and Travis
WD: Overexpression of phospho-eIF4E is associated with survival
through AKT pathway in non-small cell lung cancer. Clin Cancer Res.
16:240–248. 2010. View Article : Google Scholar
|
|
8
|
Armengol G, Rojo F, Castellví J, Iglesias
C, Cuatrecasas M, Pons B, Baselga J and Ramón y Cajal S: 4E-binding
protein 1: A key molecular 'funnel factor' in human cancer with
clinical implications. Cancer Res. 67:7551–7555. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rojo F, Najera L, Lirola J, Jiménez J,
Guzmán M, Sabadell MD, Baselga J and Ramon y Cajal S: 4E-binding
protein 1, a cell signaling hallmark in breast cancer that
correlates with pathologic grade and prognosis. Clin Cancer Res.
13:81–89. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Graff JR, Konicek BW, Lynch RL, Dumstorf
CA, Dowless MS, McNulty AM, Parsons SH, Brail LH, Colligan BM, Koop
JW, et al: eIF4E activation is commonly elevated in advanced human
prostate cancers and significantly related to reduced patient
survival. Cancer Res. 69:3866–3873. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Graff JR, Konicek BW, Carter JH and
Marcusson EG: Targeting the eukaryotic translation initiation
factor 4E for cancer therapy. Cancer Res. 68:631–634. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fan S, Ramalingam SS, Kauh J, Xu Z, Khuri
FR and Sun SY: Phosphorylated eukaryotic translation initiation
factor 4 (eIF4E) is elevated in human cancer tissues. Cancer Biol
Ther. 8:1463–1469. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang B, Zhu C, Chen B, Zhang X, Ye M and
Lin A: Expression and its clinical significance of eIF4E in
non-small cell lung cancer. Zhongguo Fei Ai Za Zhi. 13:1132–1135.
2010.In Chinese. PubMed/NCBI
|
|
14
|
Li L, Lin M, Li L, Wang R, Zhang C, Qi G,
Xu M, Rong R and Zhu T: Renal telocytes contribute to the repair of
ischemically injured renal tubules. J Cell Mol Med. 18:1144–1156.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Crinò L, Weder W and van Meerbeeck J:
Early stage and locally advanced (non-metastatic) non-small-cell
lung cancer: ESMO Clinical Practice Guidelines for diagnosis,
treatment and follow-up. Ann Oncol. 21(Suppl 5): v103–v115. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sonenberg N and Hinnebusch AG: Regulation
of translation initiation in eukaryotes: Mechanisms and biological
targets. Cell. 136:731–745. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rousseau D, Kaspar R, Rosenwald I, Gehrke
L and Sonenberg N: Translation initiation of ornithine
decarboxylase and nucleocytoplasmic transport of cyclin D1 mRNA are
increased in cells overexpressing eukaryotic initiation factor 4E.
Proc Natl Acad Sci USA. 93:1065–1070. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Scapinello A, D'Amore ES, Cavazzana AO,
Gramegna V and Ninfo V: Retroperitoneal cystic neuroendocrine
tumor. A case report. Pathologica. 87:544–547. 1995.PubMed/NCBI
|
|
19
|
Graff JR and Zimmer SG: Translational
control and metastatic progression: Enhanced activity of the mRNA
cap-binding protein eIF-4E selectively enhances translation of
metastasis-related mRNAs. Clin Exp Metastasis. 20:265–273. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Topisirovic I, Ruiz-Gutierrez M and Borden
KL: Phosphorylation of the eukaryotic translation initiation factor
eIF4E contributes to its transformation and mRNA transport
activities. Cancer Res. 64:8639–8642. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Phillips A and Blaydes JP: MNK1 and EIF4E
are downstream effectors of MEKs in the regulation of the nuclear
export of HDM2 mRNA. Oncogene. 27:1645–1649. 2008. View Article : Google Scholar
|
|
22
|
Wendel HG, Silva RL, Malina A, Mills JR,
Zhu H, Ueda T, Watanabe-Fukunaga R, Fukunaga R, Teruya-Feldstein J,
Pelletier J and Lowe SW: Dissecting eIF4E action in tumorigenesis.
Genes Dev. 21:3232–3237. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ruggero D, Montanaro L, Ma L, Xu W, Londei
P, Cordon-Cardo C and Pandolfi PP: The translation factor eIF-4E
promotes tumor formation and cooperates with c-Myc in
lymphomagenesis. Nat Med. 10:484–486. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wendel HG, De Stanchina E, Fridman JS,
Malina A, Ray S, Kogan S, Cordon-Cardo C, Pelletier J and Lowe SW:
Survival signalling by Akt and eIF4E in oncogenesis and cancer
therapy. Nature. 428:332–337. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bremaud A, West DC and Thomson AM:
Binomial parameters differ across neocortical layers and with
different classes of connections in adult rat and cat neocortex.
Proc Natl Acad Sci USA. 104:14134–14139. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ueda T, Sasaki M, Elia AJ, Chio II, Hamada
K, Fukunaga R and Mak TW: Combined deficiency for MAP
kinase-interacting kinase 1 and 2 (Mnk1 and Mnk2) delays tumor
development. Proc Natl Acad Sci USA. 107:13984–13990. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Graff JR, Konicek BW, Vincent TM, Lynch
RL, Monteith D, Weir SN, Schwier P, Capen A, Goode RL, Dowless MS,
et al: Therapeutic suppression of translation initiation factor
eIF4E expression reduces tumor growth without toxicity. J Clin
Invest. 117:2638–2648. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jacobson BA, Thumma SC, Jay-Dixon J, Patel
MR, Dubear Kroening K, Kratzke MG, Etchison RG, Konicek BW, Graff
JR and Kratzke RA: Targeting eukaryotic translation in mesothelioma
cells with an eIF4E-specific antisense oligonucleotide. PLoS One.
8:e816692013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ramalingam S, Gediya L, Kwegyir-Afful AK,
Ramamurthy VP, Purushottamachar P, Mbatia H and Njar VC: First MNKs
degrading agents block phosphorylation of eIF4E, induce apoptosis,
inhibit cell growth, migration and invasion in triple negative and
Her2-overexpressing breast cancer cell lines. Oncotarget.
5:530–543. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Konicek BW, Stephens JR, McNulty AM,
Robichaud N, Peery RB, Dumstorf CA, Dowless MS, Iversen PW, Parsons
S, Ellis KE, et al: Therapeutic inhibition of MAP kinase
interacting kinase blocks eukaryotic initiation factor 4E
phosphorylation and suppresses outgrowth of experimental lung
metastases. Cancer Res. 71:1849–1857. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kentsis A, Topisirovic I, Culjkovic B,
Shao L and Borden KL: Ribavirin suppresses eIF4E-mediated oncogenic
transformation by physical mimicry of the 7-methyl guanosine mRNA
cap. Proc Natl Acad Sci USA. 101:18105–18110. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Assouline S, Culjkovic B, Cocolakis E,
Rousseau C, Beslu N, Amri A, Caplan S, Leber B, Roy DC, Miller WH
Jr and Borden KL: Molecular targeting of the oncogene eIF4E in
acute myeloid leukemia (AML): A proof-of-principle clinical trial
with ribavirin. Blood. 114:257–260. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Moerke NJ, Aktas H, Chen H, Cantel S,
Reibarkh MY, Fahmy A, Gross JD, Degterev A, Yuan J, Chorev M, et
al: Small-molecule inhibition of the interaction between the
translation initiation factors eIF4E and eIF4G. Cell. 128:257–267.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yang SX, Hewitt SM, Steinberg SM, Liewehr
DJ and Swain SM: Expression levels of eIF4E, VEGF, and cyclin D1,
and correlation of eIF4E with VEGF and cyclin D1 in multitumor
tissue microarray. Oncol Rep. 17:281–287. 2007.PubMed/NCBI
|
|
35
|
Helkkinen T, Korpela T, Fagerholm R, Khan
S, Aittomäki K, Heikkilä P, Blomqvist C, Carpén O and Nevanlinna H:
Eukayrotic translation initiation factor 4E (eIF4E) expression is
associated with breast cancer treatment phenotype and predicts
survival after anthracycline chemotherapy treatment. Breast Cancer
Res Treat. 141:79–88. 2013. View Article : Google Scholar
|
|
36
|
Liang S, Guo R, Zhang Z, Liu D, Xu H, Xu
Z, Wang X and Yang L: Upregulation of the eIF4E signaling pathway
contributes to the progression of gastric cancer, and targeting
eIF4E by perifosine inhibits cell growth. Oncol Rep. 29:2422–2430.
2013.PubMed/NCBI
|