Introduction
Liver transplantation is currently the only
effective therapy for end-stage liver disease and severe acute
liver failure, however, its use is limited due to high costs and a
shortage of allografts. Numerous studies have shown that hepatocyte
transplantation may provide an alternative to liver
transplantation, which is safer and more tolerable for patients
suffering from end-stage liver dysfunction (1–3).
However, the major limitations of using human primary hepatocytes
as a cell source for transplantation are the requirement for a
substantial number of cells and the low proliferative capacity of
primary cells (4). Therefore, the
differentiation of human stem cells into hepatocytes may provide an
alternative cell source for liver-based cell therapies.
Previous in vitro studies have shown that
mesenchymal stem cells are readily obtained from human bone marrow,
adipose tissue and the umbilical cord, and can be differentiated
into hepatocyte-like cells (5–7).
Human adipose-derived stem cells (hADSCs) have been found to be a
more practical candidate for autologous cell transplantation,
compared with bone marrow stem cells (BMSCs) in regenerative
medicine. ADSCs may be obtained repeatedly, and in large quantities
through a convenient, safe and less painful procedure than that
used to obtain BMSCs (8–10). Previously, ADSCs have been found to
differentiate into hepatocytes in vitro and in vivo
(11). Therefore, ADSCs represent
a promising source of autologous adult stem cells for cell therapy
in the treatment of liver disease.
The mechanism underlying the ability of ADSCs, as
somatic cells, to preserve their capacity for hepatic
differentiation remains to be fully elucidated. Caveolin-1 (Cav-1),
a 21–24 kDa membrane structural protein, is a scaffolding protein
of cholesterol-rich caveolae lipid rafts in the plasma membrane,
which is important in cellular processes, including transport,
signaling and tumor suppression (12,13).
It has also been implicated in the regulation of liver regeneration
(14,15). Cav-1 is also involved in epithelial
to mesenchymal transition and in cell differentiation via the AKT,
mitogen-activated protein kinase (MAPK) and transforming growth
factor β-small mothers against decapentaplegic signaling pathways
in pancreatic cancer cells (16).
Cav-1 can also inhibit neuronal differentiation via the
downregulation of the vascular endothelial growth factor,
p44/42MAPK, AKT and signal transducer and activator of
transcription 3 signaling pathways, thus contributing to the
modulation of neuronal differentiation of neural progenitor cells
or bone mesenchymal stem cells (17). Furthermore, Cav-1 is a valuable
marker in the diagnosis of rhabdomyosarcoma, as these cells exhibit
a low degree of differentiation and a mechanical association to the
extracellular signal-regulated kinase/MAPK signaling pathways
(18). Whether Cav-1 is involved
in the differentiation of hADSCs into hepatic cells remains to be
elucidated, as do the signal transduction pathways underlying these
effects.
The present study aimed to determine whether Cav-1
is involved in the differentiation of hADSCs into hepatic cells and
to dissect the signal transduction pathways underlying these
effects. This was investigated by initially separating and
identifying hADSCs from human adipose tissue, followed by assessing
the effects of Cav-1 through analysis of the signal transduction
pathways involved in the process of hADSC differentiation into
hepatocytes. Cav-1 and its associated signaling pathway appear
promising for use in stem cell-based clinical applications of the
treatment of liver cancer.
Materials and methods
Isolation and culture of hADSCs
Human adipose tissue was obtained from five healthy
donors (two male and three female; age, 20–45 years; 10 ml tissue
samples/individual) at the First Affiliated Hospital of Dalian
Medical University (Dalian, China), and the study was approved by
the ethics committee of the First Affiliated Hospital of Dalian
Medical University. Written informed consent was obtained from the
donors. The hADSCs were isolated from raw liposuction aspirate, as
described previously (19).
Briefly, the raw liposuction aspirate was washed extensively three
times with equal volumes (10 ml) of sterile phosphate-buffered
saline (PBS) containing 1% penicillin and streptomycin (Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA). The
extracellular matrix was digested with 1% type I collagenase
(Worthington Biochemical Corporation, Lakewood, NJ, USA) at 37°C
for 60 min. The cells were separated through a 70 µm mesh
filter (BD Biosciences, San Jose, CA, USA) and the cell sap was
centrifuged at 180 × g for 5 min. The hADSC pellet was suspended in
Dulbecco's modified Eagle's medium (Gibco; Thermo Fisher
Scientific, Inc.) containing 10% fetal bovine serum (Gibco; Thermo
Fisher Scientific, Inc.) and maintained at 37°C in 5%
CO2 (all incubation conditions for regular cell culture
in this study were 37°C in 5% CO2). Cell cultures at a
confluence of 80–90% were used for subsequent experiments. To
quantify cell growth, the hADSCs were seeded in triplicate at a
density of 4,000 cells per well in 24-well plates. The cells were
digested and counted using a hemocytometer (XB-K-25; Anxin Co.,
Shanghai, China) every day for 7 days. Three wells were used for
each cell count and the analysis was repeated in triplicate.
Fluorescence immunocytochemistry
The hADSCs were seeded at a density of 4,000
cells/well in 24-well plates. The cells were then rinsed with PBS
three times and fixed in 4% paraformaldehyde (Tianjin Bodi Chemical
Co., Ltd., Tianjin, China) at room temperature for 1 h. Following
washing, 2 ml PBS mixed with 1% Triton X-100 (Beijing Solarbio
Science & Technology Co., Ltd., Beijing, China) was added to
each well for 10 min to increase the permeability of the cell
membranes. The samples were incubated with mouse monoclonal
anti-human Cav-1 antibody (1:100; cat. no. sc-53564; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA), mouse monoclonal anti-human
albumin (ALB; 1:70; cat. no. sc-271604; Santa Cruz Biotechnology,
Inc.), mouse monoclonal anti-human α-fetoprotein (AFP; 1:70; cat.
no. sc-166335; Santa Cruz Biotechnology, Inc.) at 4°C overnight.
Following washing three times with PBS, FITC-labeled goat
anti-mouse immuno-globulin (Ig)G (1:70; cat. no. F0382;
Sigma-Aldrich, St. Louis, MO, USA) was added for 1 h at room
temperature, followed by washing with PBS. Hoechst 33258 (1:500;
Sigma-Aldrich) was then added for nuclear staining.
Flow cytometric analysis
Flow cytometry (BD FACSAria II; BD Pharmingen, San
Diego, CA, USA) was performed on the hADSCs. The cells were washed
twice with PBS and counted under a microscope (Leica Microsystems
GmbH, Wetzlar, Germany). Cells were analyzed using conjugated mouse
monoclonal antibodies. The cells were washed once in flow wash
buffer (1X Dulbecco's BPS, 0.5% bovine serum albumin and 0.1%
sodium azide), resuspended in blocking buffer (wash buffer with 25
µg/ml mouse IgG), and incubated for 10 min at room
temperature. A total of 100 µl cell suspension
(~1×105 cells) was added to each tube, and labeled
monoclonal antibodies were added for tricolor analysis (FITC and
PE). Isotype control combinations were performed to reflect
monoclonal isotype combinations. The following antibodies:
Monoclonal mouse anti-human CD13 PE (cat. no. 347837), CD34 PE
(cat. no. 348057), CD45 FITC (cat. no. 347463), CD71 FITC (cat. no.
347513), CD90 FITC (cat. no. 555595) and CD106 PE (cat. no. 555647)
were purchased from BD Pharmingen. CD90 was diluted at 1:100 and
the remaining antibodies were diluted at 1:10. All tubes were
incubated and protected from the light for 30 min. Following
incubation, the cells were washed once with wash buffer and fixed
in 200 µl 4% paraformaldehyde. Data analysis was performed
using BD FACSDiva Software v 6.1.3 (BD Pharmingen).
Adipogenic differentiation of hADSCs
For adipogenic differentiation, the hADSCs were
seeded into a 6-well plate at 1×104 cells per well in
complete medium. Following 24 h incubation, the cells were cultured
in AdipoDiff medium, comprising H-DMEM with 10% FBS, 0.5 mmol/l
1,3-isobutyl methylxanthine (IBMX; Sigma-Aldrich),
1×10−6 M dexamethasone (Dex; Sigma-Aldrich), 10
µg/ml insulin (Sigma-Aldrich) and 0.2 mmol/l indometacin
(Sigma-Aldrich), for 2 weeks, with medium replaced with fresh
medium every 3 days. The induction of adipogenic differentiation
was detected using Oil Red O staining (Sigma-Aldrich). The formalin
fixed cells were washed three times with PBS and stained with 0.5%
Oil Red O for 5 min. Following counterstaining with hematoxylin
(Beyotime Institute of Biotechnology, Shanghai, China), the red
colored lipids were detected under a light microscope (DMI4000B;
Leica Microsystems GmbH).
Osteogenic differentiation of hADSCs
For osteogenic differentiation, the hADSCs were
seeded into a 6-well plate at 1×104 cells per well in
complete medium for 24 h. Subsequently, OsteoDiff medium was added
(H-DMEM with 10% FBS, 1×10−7 M Dex, 10 mmol/l β-sodium
glycerophosphate and 50 µmol/l vitamin C; both from
Sigma-Aldrich) to induce osteogenesis for 3 weeks. The osteogenic
differentiation was evaluated by the calcium accumulation using Von
Kossa staining. Briefly, the cells were rinsed with PBS and fixed
with 95% ethanol at room temperature for 10 min. Following
treatment with 2% silver nitrate (Tianjin Bodi Chemical Co., Ltd.)
and exposure to sunlight for 15–60 min, the cells were washed and
then reduced by 2% sodium thiosulfate (Tianjin Bodi Chemical Co.,
Ltd.) for 1 h. Eosin (Beyotime Institute of Biotechnology) was used
for counterstaining, and the cells were observed using a microscope
(Leica DMI4000B).
Hepatic induction
Passage four cells were seeded into 6-well plates at
a density of 8,000 cells per well in complete medium for 24 h. The
complete medium was replaced with serum-free H-DMEM with 10 ng/ml
basic fibroblast growth factor (bFGF; PeproTech, Inc., Rocky Hill,
NJ) and 20 ng/ml human epidermal growth factor (PeproTech, Inc.)
for 48 h. The cells were then cultured in H-DMEM with 10 ng/ml
bFGF, 20 ng/ml hepatocye growth factor (Sigma-Aldrich), 15% FBS and
1×10−7 M Dex for 7, 14 and 21 days.
ELISA analysis
Culture supernatants were collected for the
measurement of urea content using an ELISA kit (cat. no. MAK077;
Sigma-Aldrich) according to the manufacturer's instructions.
Absorbance was read at 450 nm using an ELx808 Absorbance Reader
(Bio-Tek Instruments, Inc., Winooski, VT, USA) and analyzed using
Gen5 software (Bio-Tek Instruments, Inc.).
Transient transfection of Cav-1 siRNA
into hADSCs
For transfection, 8×103 cells per well
were seeded into 6-well plates for 24 h and induced towards hepatic
differentiation. 2 µg Cav-1 siRNA was transiently
transfected into the cells on days 5 and 12 using Lipofectmine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. The cells were incubated in compete medium
4 h following transfection and were collected for examination after
48 h.
Semiquantitative reverse
transcription-polymerase chain reaction (RT-PCR)
Total RNA was extracted from the cells using RNAiso
Plus (Takara Bio, Inc., Otsu, Japan), according to the
manufacturer's protocol. RNA (5 µg) was reverse transcribed
using MMLV Reverse Transcriptase (Takara Bio, Inc.) and oligo dT.
The primers (Takara Bio, Inc.) used for RT-PCR are listed in
Table I. A PCR kit (Premix Taq™;
cat. no. RR901A; Takara Bio, Inc.) was used according to the
manufacturer's instructions. The reaction system contained cDNA
(2.5 µl), Premix Taq (25 µl), primers (2 µl)
and sterile purified water (≤50 µl). The samples were
amplified with an initial denaturation step of 94°C for 5 min,
followed by 30 cycles of 94°C for 30 sec, 55°C for 30 sec, 72°C for
1 min and a final extension step of 72°C for 10 min, using a PCR
thermocycler (Techne TC-512; Bibby Scientific Ltd., Staffordshire,
UK). The PCR products (15 µl) were separated by
electrophoresis on 1% agarose gels (Invitrogen; Thermo Fisher
Scientific, Inc.) and visualized with ethidium bromide
(Sigma-Aldrich; final concentration 0.5 µg/ml) staining
under ultraviolet illumination (24–25-PR; DNR Bio-Imaging Systems
Ltd., Jerusalem, Israel).
 | Table IList of primer sequences used for
semiquantitative reverse transcription-polymerase chain reaction
analysis. |
Table I
List of primer sequences used for
semiquantitative reverse transcription-polymerase chain reaction
analysis.
| Gene (Accession;
length) | Primer
sequence |
|---|
| Oct
(NM_001173531.2; 433 bp) | Forward
5′-GGGGTTCTATTTGGGAAGGTATT-3′
Reverse 5′-CAGAGTGGTGACGGAGACAGG-3′ |
| Nanog
(NM_024865.2; 367 bp) | Forward
5′-ACCTATGCCTGTGATTTGTGGG-3′
Reverse 5′-GGGTTGTTTGCCTTTGGGAC-3′ |
| ALB
(NM_000477.5; 598 bp) | Forward
5′-GTGTTGATTGCCTTTGCTCAGTAT-3′
Reverse 5′-GGAGGTTTGGGTTGTCATCTTTGT-3′ |
| AFP
(NM_001134.2; 174 bp) | Forward
5′-ATTGAGAAACCCACTGGAGATGA-3′
Reverse 5′-TAGCGAGCAGCCCAAAGAAG-3′ |
| CYP1A1
(NM_000499.3; 147 bp) | Forward
5′-TCCCTATTCTTCGCTACCTACCC-3′
Reverse 5′-TCAGGCTGTCTGTGATGTCCC-3′ |
| HNF1A
(NM_000545.5; 208 bp) | Forward
5′-GAGGACGAGACGGACGACGAT-3′
Reverse 5′-GGAGTGCCCTTGTTGAGGTGTT-3′ |
| HNF1B
(NM_000458.2; 152 bp) | Forward
5′-CCCCTATGAAGACCCAGAAGC-3′
Reverse 5′-GACACGGACCTCAGTGACCAA-3′ |
| GAPDH
(NM_001256799.2; 700 bp) | Forward
5′-TGACCTGCCGTCTAGAAAACC-3′
Reverse 5′-GTGGGTGTCGCTGTTGAAGTC-3′ |
Western blot analysis
The cells were disrupted with
radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology), containing 50 mM Tris-HCl (pH 8.0), 150 mM NaCl,
0.25 mM EDTA (pH 8.0), 0.1% SDS, 1% Triton X-100, 50 mM NaF and a
protease inhibitor cocktail. The lysates were boiled in gel-loading
buffer (Takara Bio, Inc.) and separated on 10% SDS-PAGE gels
(Beijing Solarbio Science & Technology Co., Ltd.). Following
PAGE, the proteins were transferred onto a polyvinylidene fluoride
membrane (EMD Millipore, Billerica, MA, USA). The membrane was
blocked with 5% non-fat dry milk (dissolved by PBS with Tween 20;
Gibco; Thermo Fisher Scientific, Inc.) at room temperature for 1 h,
prior to being probed with the indicated antibodies and visualized
with horseradish peroxidase-conjugated polyclonal goat anti-mouse
(1:5,000; cat. no. 31430; Thermo Fisher Scientific, Inc.) and
polyclonal goat anti-rabbit (1:5,000; cat. no. 31460; Thermo Fisher
Scientific, Inc.) secondary antibodies and enhanced
chemiluminescence reagents (GE Healthcare Life Sciences,
Piscataway, NJ, USA). The phosphorylated monoclonal mouse
anti-human (p)-MAPK (1:2,000; cat. no. 9106) and polyclonal rabbit
anti-human total MAPK (1:1,000; cat. no. 9102) antibodies were
purchased from Cell Signaling Technology, Inc.
Statistical analysis
All values are expressed as the mean ± standard
deviation. Student's t-test was used to identify significant
differences using SPSS 14.0 software (SPSS, Inc., Chicago, IL,
USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
hADSCs isolation and characteristic
identification
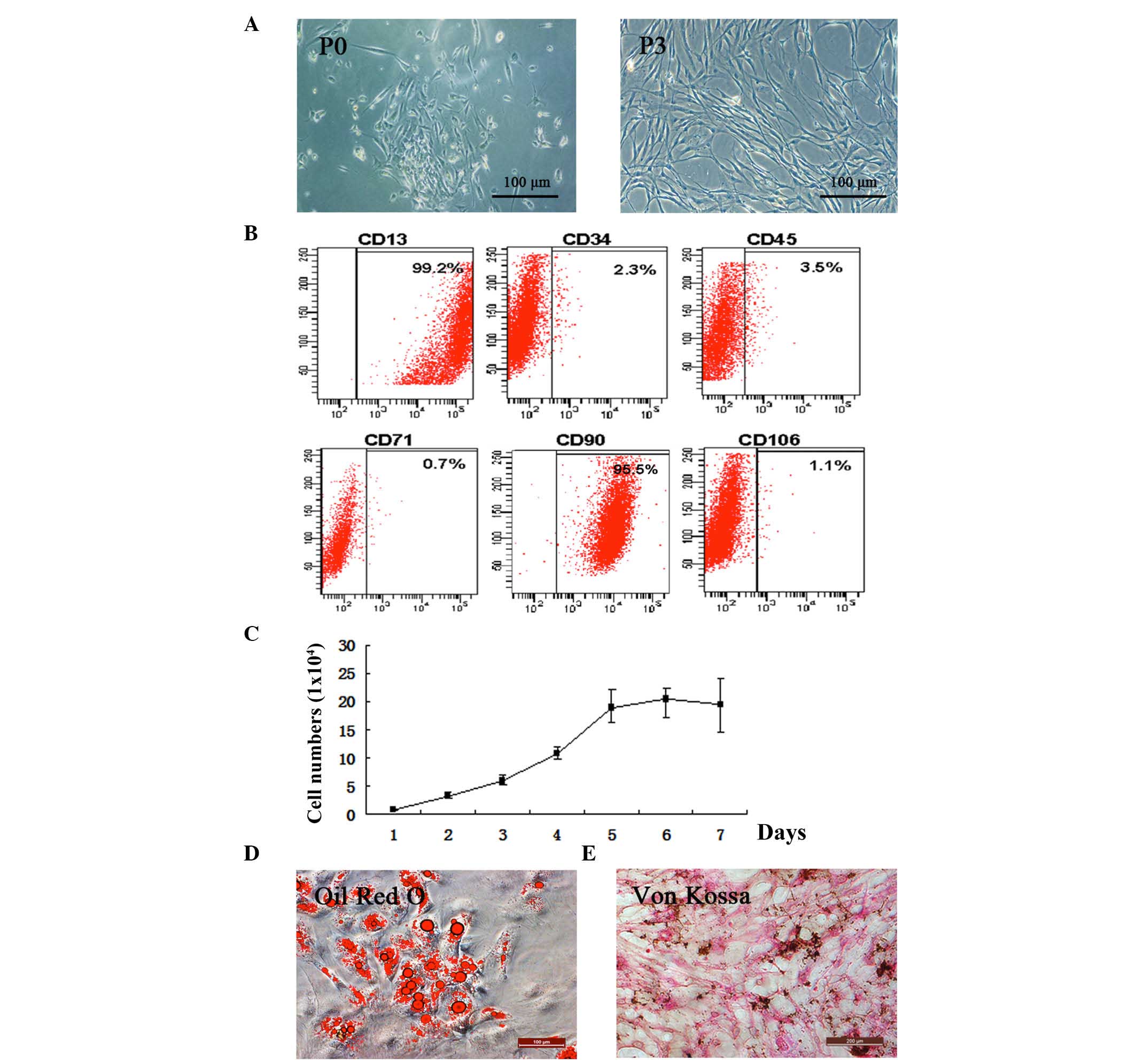
The present study isolated hADSCs from adipose
tissue (Fig. 1A). The cells began
to attach to flasks and proliferate to passage three. To determine
whether the isolated cells preserved ADSC characteristics, the
expression levels of the CD13 and CD90 stem cell surface markers
were quantified and were found to be 99.2 and 95.5%, respectively.
CD34, CD45, CD71 and CD106 were not expressed. This indicated that
isolated hADSCs remained in an undifferentiated state (Fig. 1B). The results of the proliferation
assay showed that the isolated hADSCs were capable of
proliferation, an essential property of ADSCs (20) (Fig.
1C).
Two weeks following exposure to AdipoDiff medium,
intracellular lipid droplets were observed within the
adipogenic-differentiated hADSCs using Oil Red O staining (Fig. 1D). Von Kossa staining of the hADSCs
was maintained in the OsteoDiff medium during differentiation
(Fig. 1E).
Hepatic differentiation capacity of
hADSCs
To evaluate whether isolated and primary cultured
hADSCs retained hepatic differentiation ability, hepatocyte
inducing factors were added, and hepatocyte marker proteins and
genes were detected. α-fetoprotein (AFP) and albumin (ALB) were
expressed at high levels on days 7 and 14, but were undetectable in
the untreated hADSCs, indicating that the hADSCs underwent
successful hepatic differentiation (Fig. 2A and B). To confirm this,
semi-quantitative RT-PCR was performed to detect markers of gene
transcription. The ADSC marker genes, Oct4 and Nanog,
were activated in the absence of induction, and were gradually
inactivated as the hADSCs differentiated into hepatocytes.
Oct4 and Nanog were almost undetectable on day 21,
indicating the complete differentiation of hADSCs into hepatocytes
(Fig. 2C). The present study also
assessed the hepatic marker genes, ALB, CYP1A1,
HNF1A and HNF1B, which gradually increased and were
most prevalent on day 21. AFP, a gene associated with the
major plasma protein produced by the yolk sac and liver during
fetal development, decreased as hADSCs underwent differentiation,
which was in accordance with liver development (Fig. 2C). Additionally, urea synthesis,
known as an important function of hepatocytes, significantly
increased with the duration of hepatic differentiation (Fig. 2D). These results suggested that
isolated hADSCs differentiated into hepatocyte-like cells.
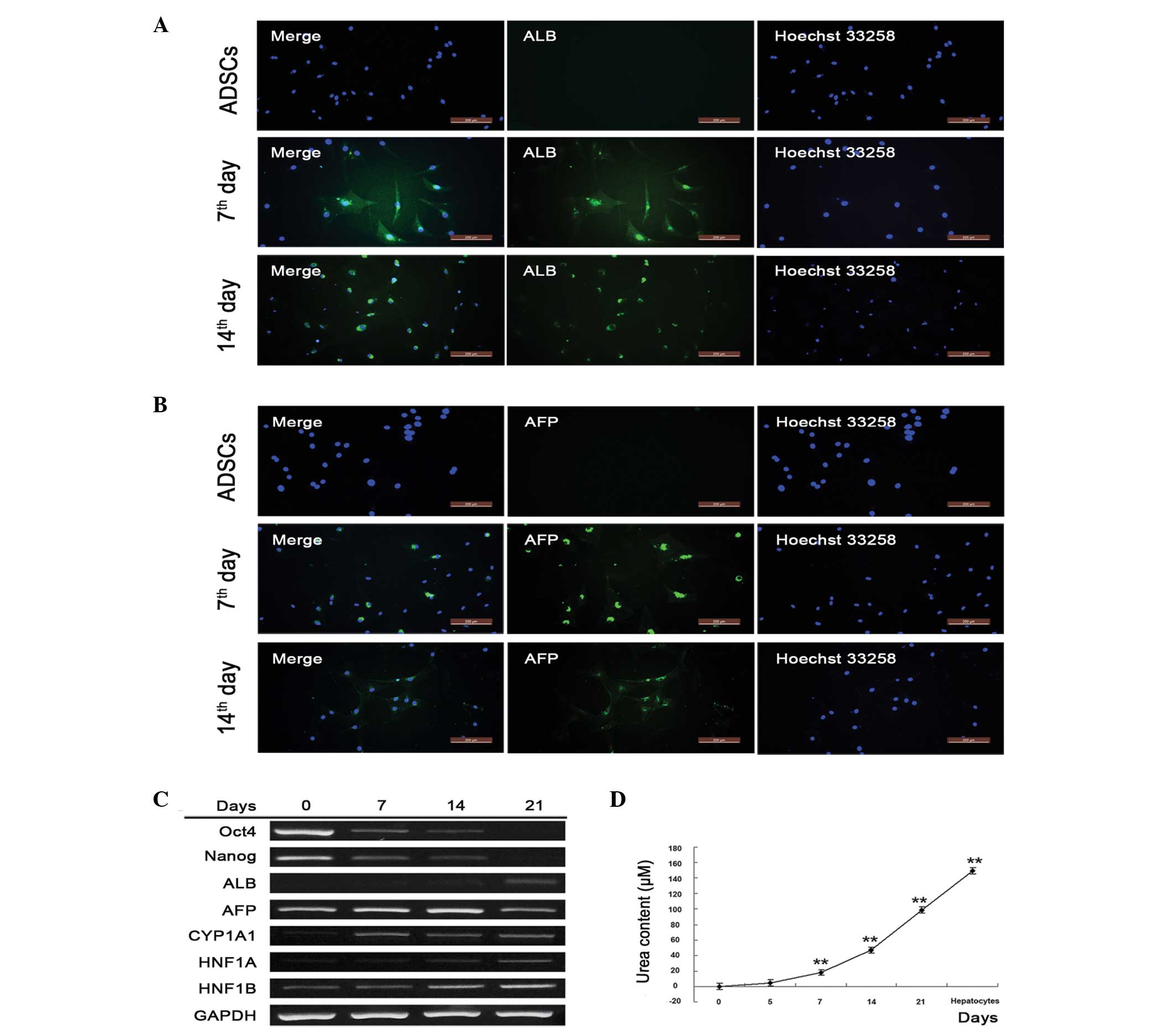 | Figure 2Isolated hADSCs retain hepatic
differentiation capacity. Expression levels of (A) ALB and (B) AFP
were detected using immunofluorescence following 7 and 14 days of
hADSC differentiation. The nuclei were stained with Hoechst (right
panel) and the target proteins (ALB and AFP) were stained with
relative antibodies (middle panel). The merged images are shown in
the left panel (scale bar=200 µm). (C) Stem cell- and
hepatocyte-specific gene transcription was detected using
semiquantitative reverse transcription-polymerase chain reaction
analysis following 7, 14 and 21 days of hADSC differentiation. (D)
Urea content in the culture medium of the induction group.
Supernatants were collected and stored at −20°C. For detection,
samples were processed according to the manufacturer's protocol,
and the optical density was detected using a spectrometer at 450 nm
within 15 min following the addition of stop buffer.
*P<0.05 vs. untreated cells; **P<0.01
vs. untreated cells (n≥3). hADSCs, human adipose-derived stem
cells; ALB, albumin; AFP, α-fetoprotein; Oct4, octamer-binding
transcription factor 4; CYP1A1, cytochrome P450, family 1, member
A1; HNF1A hepatocyte nuclear factor-1-α; HNF1B, hepatocyte nuclear
factor-1-β; GAPDH, glyceraldehyde 3-phosphate dehydrogenase. |
Downregulation of Cav-1 inhibits hepatic
marker proteins by attenuating the MEK/MAPK pathway during the
induction of hepatocyte-like cells from hADSCs
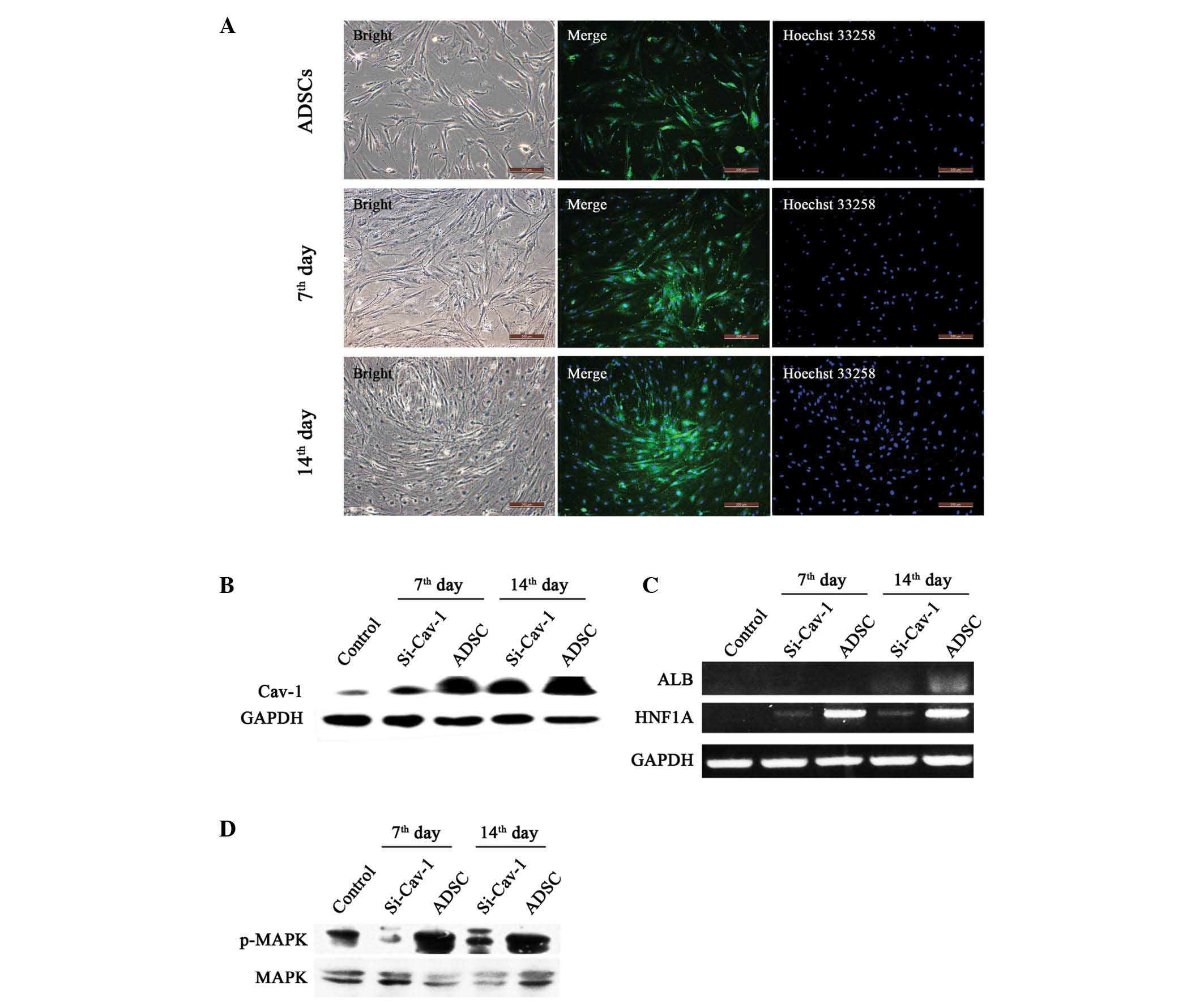
On days 7 and 14 of differentiation, the results of
the immunostaining and western blot analyses showed that the
expression of Cav-1 was significantly increased, compared with that
in the control, indicating a positive role in hepatocyte
differentiation (Fig. 3A and B).
To assess whether Cav-1 has a central role in hepatic
differentiation, the present study transiently knocked down
Cav-1 on days 5 and 12 (Fig.
3B). With the suppression of Cav-1, the gene expression
levels of ALB and HNF1A were significantly reduced
(Fig. 3C), indicating attenuation
of the hepatic differentiation process following Cav-1
knockdown. Furthermore, to identify the signaling pathway involved
in differentiation, the present study examined the MAPK pathway and
found that it was significantly activated during early
differentiation. In addition, the MAPK pathway was largely
attenuated following Cav-1 knockdown (Fig. 3D).
Discussion
hADSCs are widely present in the adult human body
and have been suggested as an emerging and attractive tool in
regenerative medicine, with vast applications. They are easily
obtained, hold multiple potency (21), and have been suggested for use in
liver transplantation to prolong patient survival, with ADSC
transplantation shown to promote liver recovery in mice with liver
injury (22,23). In the present study, hADSCs were
successfully isolated from human adipose tissue and it was found
that these cells were able to differentiate into hepatocyte-like
cells following incubation in hepatocyte growing conditions, which
was consistent with previous studies (24–27).
The hADSCs separated from the adipose tissue
remained pluripotent and retained CD13 and CD90 stem cell surface
markers. In addition they demonstrated the ability of adipogenic
differentiation and osteogenic differentiation. These results
suggested the potential of hADSCs for use in stem-cell-based
therapies. Furthermore, ADSCs may be obtained repeatedly and in
large quantities, through a convenient and safe procedure, thus
presenting a practical alternative to BMSCs for clinical
applications (28). Although BMSCs
have the same biological properties as ADSCs, the numbers of hMSCs,
obtained through marrow aspiration, decline with age (29). In addition, there is an age-related
decline in overall BMSC differentiation potency, which poses a
problem for their use in cell-based therapies. By contrast, the
differentiation capacity of ADSCs is minimally affected by donor
age (30).
The mechanism underlying hepatic ADSC
differentiation remains to be fully elucidated. Previous studies
have focused on the clinical and tissue engineering applications of
ADSCs in regenerative medicine (4,31),
however, they have not investigated the capacity of ADSCs, as
somatic cells, to retain multipotent differentiation functionality.
Several of these studies have focussed on a description of the
phenomenon, rather than examination of the mechanism (32).
Our previous study investigated the role of Cav-1 in
human hepatocyte proliferation using Chang liver cells (CHL), and
found that the downregulation of Cav-1 induced biphasic
change in growth rate, associated with AKT and MAPK signaling
(33). In addition, Cav-1 has been
shown to be involved in the progress of liver regeneration and
liver development (14,34,35).
Previous results have also indicated that the expression of Cav-1
in the mouse liver increased in a time-dependent manner across
developmental stages from natal to 3 months post-natal. However,
whether Cav-1 has the same effect in the hepatic differentiation of
hADSCs has received little attention. In the present study, hADSCs
were successfully induced into hepatocyte-like cells, and the
observed trend of expression of Cav-1 in the hADSCs during hepatic
differentiation was the same as that observed in mouse liver cells
during development, with undifferentiated hADSCs exhibiting low
expression levels of Cav-1, which gradually increased as hepatic
differentiation commences. This is not an unexpected result, as
cholesterol and lipid metabolism are acquired during liver
development (36,37) and Cav-1 has been shown to be key in
this process (38,39). In addition, the present study found
that Cav-1 knockdown was accompanied by a decrease in HNF1A
and ALB, indicating that Cav-1 is essential in the hepatic
differentiation process of hADSCs. Although it was observed that
the expression of Cav-1 increased significantly on day 14 of
hepatic differentiation, this expression level was low compared
with those in Chang liver cells, suggesting that preservation of
certain stem cell properties in the hADSCs.
Liver regeneration also requires hepatocyte
proliferation, and the MAPK signaling pathway has a major role in
this process (40). A previous
study reported that the MAPK signaling pathway is important for
ADSC transplantation into the damaged liver (41). The results of the present study
demonstrated that p-MAPK significantly decreases during
Cav-1 knockdown, resulting in a subsequent decrease in the
levels of ALB and HNF1A, an indication of failed
hepatic differentiation. Taken together, these results suggested
that the Cav-1-MAPK signaling pathway is important in hADSC hepatic
differentiation.
The present study showed for the first time, to the
best of our knowledge, that Cav-1 is essential for hADSC hepatic
differentiation, and the mechanism underlying this differentiation
was further characterized. The results suggested that the
regulation of Cav-1 and its associated pathway presents a promising
avenue for further investigation, with the ultimate aim to increase
the efficiency of hADSC hepatic differentiation for use in
practical stem cell-based clinical applications.
Acknowledgments
This study was supported by the Chinese National
Natural Science Foundation (grant nos. 81071009 and 81271412), the
International S&T Cooperation Project of the Ministry of
S&T of China, (grant no. 2010DFR30850), the Science and
Technology Plan Projects in Liaoning Province (grant no. 201501522)
and the Scientific Research Foundation for the Returned Overseas
Chinese Scholars, State Education Ministry.
References
|
1
|
Glanemann M, Gaebelein G, Nussler N, Hao
L, Kronbach Z, Shi B, Neuhaus P and Nussler AK: Transplantation of
monocyte-derived hepatocyte-like cells (NeoHeps) improves survival
in a model of acute liver failure. Ann Surg. 249:149–154. 2009.
View Article : Google Scholar
|
|
2
|
Hossein Aghdaie M, Geramizadeh B, Azarpira
N, Esfandiari E, Darai M, Rahsaz M, Nikeghbalian S and
Malekhosseini SA: Hepatocyte isolation from unused/rejected livers
for transplantation: Initial step toward hepatocyte
transplantation, the first experience from iran. Hepat Mon.
13:e103972013.PubMed/NCBI
|
|
3
|
Forbes SJ and Alison MR: Regenerative
medicine: Knocking on the door to successful hepatocyte
transplantation. Nat Rev Gastroenterol Hepatol. 11:277–278. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zheng MH, Ye C, Braddock M and Chen YP:
Liver tissue engineering: Promises and prospects of new technology.
Cytotherapy. 12:349–360. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Williams AR and Hare JM: Mesenchymal stem
cells: Biology, pathophysiology, translational findings and
therapeutic implications for cardiac disease. Circ Res.
109:923–940. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Aurich H, Sgodda M, Kaltwasser P, Vetter
M, Weise A, Liehr T, Brulport M, Hengstler JG, Dollinger MM, Fleig
WE and Christ B: Hepatocyte differentiation of mesenchymal stem
cells from human adipose tissue in vitro promotes hepatic
integration in vivo. Gut. 58:570–581. 2009. View Article : Google Scholar
|
|
7
|
Anzalone R, Lo Iacono M, Corrao S, Magno
F, Loria T, Cappello F, Zummo G, Farina F and La Rocca G: New
emerging potentials for human Wharton's jelly mesenchymal stem
cells: Immunological features and hepatocyte-like differentiative
capacity. Stem Cells Dev. 19:423–438. 2010. View Article : Google Scholar
|
|
8
|
Al Battah F, De Kock J, Vanhaecke T and
Rogiers V: Current status of human adipose-derived stem cells:
Differentiation into hepatocyte-like cells. Scientific World
Journal. 11:1568–1581. 2011. View Article : Google Scholar
|
|
9
|
Zhu Y, Liu T, Song K, Fan X, Ma X and Cui
Z: Adipose-derived stem cell: A better stem cell than BMSC. Cell
Biochem Funct. 26:664–675. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Huang SJ, Fu RH, Shyu WC, Liu SP, Jong GP,
Chiu YW, Wu HS, Tsou YA, Cheng CW and Lin SZ: Adipose-derived stem
cells: Isolation, characterization and differentiation potential.
Cell Transplant. 22:701–709. 2013. View Article : Google Scholar
|
|
11
|
Ruiz JC, Ludlow JW, Sherwood S, Yu G, Wu X
and Gimble JM: Differentiated human adipose-derived stem cells
exhibit hepatogenic capability in vitro and in vivo. J Cell
Physiol. 225:429–436. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang X, Shen P, Coleman M, Zou W, Loggie
BW, Smith LM and Wang Z: Caveolin-1 down-regulation activates
estrogen receptor alpha expression and leads to
17beta-estradiol-stimulated mammary tumorigenesis. Anticancer Res.
25:369–375. 2005.PubMed/NCBI
|
|
13
|
Wang XX, Wu Z, Huang HF, Han C, Zou W and
Liu J: Caveolin-1, through its ability to negatively regulate TLR4,
is a crucial determinant of MAPK activation in LPS-challenged
mammary epithelial cells. Asian Pac J Cancer Prev. 14:2295–2299.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Frank PG and Lisanti MP: Caveolin-1 and
liver regeneration: role in proliferation and lipogenesis. Cell
Cycle. 6:115–116. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fernández-Rojo MA, Gongora M, Fitzsimmons
RL, Martel N, Martin SD, Nixon SJ, Brooks AJ, Ikonomopoulou MP,
Martin S, Lo HP, et al: Caveolin-1 is necessary for hepatic
oxidative lipid metabolism: Evidence for crosstalk between
caveolin-1 and bile acid signaling. Cell Rep. 4:238–247. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Salem AF, Bonuccelli G, Bevilacqua G,
Arafat H, Pestell RG, Sotgia F and Lisanti MP: Caveolin-1 promotes
pancreatic cancer cell differentiation and restores membranous
E-cadherin via suppression of the epithelial-mesenchymal
transition. Cell Cycle. 10:3692–3700. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li Y, Luo J, Lau WM, Zheng G, Fu S, Wang
TT, Zeng HP, So KF, Chung SK, Tong Y, et al: Caveolin-1 plays a
crucial role in inhibiting neuronal differentiation of neural
stem/progenitor cells via VEGF signaling-dependent pathway. PLoS
One. 6:e229012011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rossi S, Poliani PL, Cominelli M, Bozzato
A, Vescovi R, Monti E and Fanzani A: Caveolin 1 is a marker of poor
differentiation in Rhabdomyosarcoma. Eur J Cancer. 47:761–772.
2011. View Article : Google Scholar
|
|
19
|
Locke M, Windsor J and Dunbar PR: Human
adipose-derived stem cells: Isolation, characterization and
applications in surgery. ANZ J Surg. 79:235–244. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bunnell BA, Flaat M, Gagliardi C, Patel B
and Ripoll C: Adipose-derived stem cells: Isolation, expansion and
differentiation. Methods. 45:115–120. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gimble JM, Katz AJ and Bunnell BA:
Adipose-derived stem cells for regenerative medicine. Circ Res.
100:1249–1260. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Banas A, Teratani T, Yamamoto Y, Tokuhara
M, Takeshita F, Osaki M, Kawamata M, Kato T, Okochi H and Ochiya T:
IFATS collection: In vivo therapeutic potential of human adipose
tissue mesenchymal stem cells after transplantation into mice with
liver injury. Stem Cells. 26:2705–2712. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Banas A, Teratani T, Yamamoto Y, Tokuhara
M, Takeshita F, Osaki M, Kato T, Okochi H and Ochiya T: Rapid
hepatic fate specification of adipose-derived stem cells and their
therapeutic potential for liver failure. J Gastroenterol Hepatol.
24:70–77. 2009. View Article : Google Scholar
|
|
24
|
Banas A, Teratani T, Yamamoto Y, Tokuhara
M, Takeshita F, Quinn G, Okochi H and Ochiya T: Adipose
tissue-derived mesenchymal stem cells as a source of human
hepatocytes. Hepatology. 46:219–228. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sgodda M, Aurich H, Kleist S, Aurich I,
König S, Dollinger MM, Fleig WE and Christ B: Hepatocyte
differentiation of mesenchymal stem cells from rat peritoneal
adipose tissue in vitro and in vivo. Exp Cell Res. 313:2875–2886.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen X, Zhang S, Liu T, Liu Y and Wang Y:
Maintenance of rat hepatocytes under inflammation by coculture with
human orbital fat-derived stem cells. Cell Mol Biol Lett.
17:182–195. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li X, Yuan J, Li W, Liu S, Hua M, Lu X and
Zhang H: Direct differentiation of homogeneous human adipose stem
cells into functional hepatocytes by mimicking liver embryogenesis.
J Cell Physiol. 229:801–812. 2014. View Article : Google Scholar
|
|
28
|
Mimeault M and Batra SK: Recent progress
on tissue-resident adult stem cell biology and their therapeutic
implications. Stem Cell Rev. 4:27–49. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Stolzing A, Jones E, McGonagle D and Scutt
A: Age-related changes in human bone marrow-derived mesenchymal
stem cells: Consequences for cell therapies. Mech Ageing Dev.
129:163–173. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shi YY, Nacamuli RP, Salim A and Longaker
MT: The osteogenic potential of adipose-derived mesenchymal cells
is maintained with aging. Plast Reconstr Surg. 116:1686–1696. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mailey B, Hosseini A, Baker J, Young A,
Alfonso Z, Hicok K, Wallace AM and Cohen SR: Adipose-derived stem
cells: Methods for isolation and applications for clinical use.
Methods Mol Biol. 1210:161–181. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sterodimas A, de Faria J, Nicaretta B and
Pitanguy I: Tissue engineering with adipose-derived stem cells
(ADSCs): Current and future applications. J Plast Reconstr Aesthet
Surg. 63:1886–1892. 2010. View Article : Google Scholar
|
|
33
|
Ren G, Liu Y, Wang XM, Zhao CH and Zou W:
Role of caveolin-1 down-regulation by iRNA in human hepatocyte
proliferation. Zhonghua Gan Zang Bing Za Zhi. 16:379–382. 2008.In
Chinese. PubMed/NCBI
|
|
34
|
Fernández MA, Albor C, Ingelmo-Torres M,
Nixon SJ, Ferguson C, Kurzchalia T, Tebar F, Enrich C, Parton RG
and Pol A: Caveolin-1 is essential for liver regeneration. Science.
313:1628–1632. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mayoral R, Fernández-Martínez A, Roy R,
Boscá L and Martín-Sanz P: Dispensability and dynamics of
caveolin-1 during liver regeneration and in isolated hepatic cells.
Hepatology. 46:813–822. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bosch M, Marí M, Herms A, Fernández A,
Fajardo A, Kassan A, Giralt A, Colell A, Balgoma D, Barbero E, et
al: Caveolin-1 deficiency causes cholesterol-dependent
mitochondrial dysfunction and apoptotic susceptibility. Curr Biol.
21:681–686. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Baker N and Tuan RS: The
less-often-traveled surface of stem cells: Caveolin-1 and caveolae
in stem cells, tissue repair and regeneration. Stem Cell Res Ther.
4:902013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Frank PG, Pavlides S, Cheung MW, Daumer K
and Lisanti MP: Role of caveolin-1 in the regulation of lipoprotein
metabolism. Am J Physiol Cell Physiol. 295:C242–C248. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Fernández-Rojo MA, Restall C, Ferguson C,
Martel N, Martin S, Bosch M, Kassan A, Leong GM, Martin SD, McGee
SL, et al: Caveolin-1 orchestrates the balance between glucose and
lipid-dependent energy metabolism: Implications for liver
regeneration. Hepatology. 55:1574–1584. 2012. View Article : Google Scholar
|
|
40
|
Campbell JS, Argast GM, Yuen SY, Hayes B
and Fausto N: Inactivation of p38 MAPK during liver regeneration.
Int J Biochem Cell Biol. 43:180–188. 2011. View Article : Google Scholar :
|
|
41
|
Liang L, Ma T, Chen W, Hu J, Bai X, Li J
and Liang T: Therapeutic potential and related signal pathway of
adipose-derived stem cell transplantation for rat liver injury.
Hepatol Res. 39:822–832. 2009. View Article : Google Scholar : PubMed/NCBI
|