Introduction
Ketamine, a noncompetitive N-methyl-D-aspartate
receptor (NMDAR) ion channel blocker, is a commonly used anesthetic
in pediatric patients (1).
Previous studies have suggested that ketamine induces
neuroapoptosis in developing animal brains (2,3) and
primary cultured neurons (4,5),
resulting in persistent cognitive deficits as the animal matures
(2,3). Thus far, the mechanisms underlying
ketamine-induced neuroapoptosis in the developing brain remain
elusive.
Steroid hormones and their metabolites present
within the central nervous system (CNS) are commonly defined as
neuro-active steroids or neurosteroids (6). They can be synthesized de novo
from cholesterol in the CNS by glial cells and neurons, or in the
periphery by the adrenal and gonadal glands. The concentration of
neurosteroids is higher in the CNS than in the periphery (6). They are regulators of CNS function,
and serve important roles in mood, behavior, reproduction and
cognition, and act as protective agents in models of injury and
disease, such as experimental models of traumatic brain injury,
including Alzheimer's disease (AD), autism, stroke and mood
disorders (6–8). In addition, certain neurosteroids
exert important regulatory and protective roles in the fetal brain
(9). Li et al (5) suggested that 17β-estradiol is able to
protect primary cultured cortical neurons against ketamine-induced
neuroapoptosis. Reduced concentrations or the absence of
neurosteroids during development and in adults may be associated
with neurodevelopmental, psychiatric or behavioral disorders or
neurodegeneration (10). A
previous study demonstrated that neurotoxicity induced by the NMDAR
blocker MK801 is associated with reduced levels of 17β-estradiol
(11). Ethanol, another NMDAR
blocker, has been reported to alter certain neurosteroid levels
with prenatal exposure, and this alteration may be associated with
the pathogenesis of ethanol-induced neurodevelopmental disorders
and fetal alcohol syndrome in the developing rat brain (12). Trickler et al (13), suggested that ketamine may
attenuate 17β-estradiol levels in the early life stages of
zebrafish, and ketamine has been indicated to be neurotoxic to
zebrafish embryos (14).
Therefore, it is reasonable to assume that
neurosteroid biosynthesis is an important mechanism underlying the
neuro-protection against ketamine-induced neuroapoptosis, and that
it may be regulated or markedly affected in primary cultured
cortical neurons. In the present study, the main aim was to
investigate the effects of ketamine exposure on the biosynthesis of
neurosteroids under ketamine-induced neuroapoptosis conditions in
primary cultured cortical neurons.
Materials and methods
Reagents
Gibco Dulbecco's modified Eagle's medium (DMEM),
fetal bovine serum (FBS), neurobasal medium and B27 supplement were
purchased from Thermo Fisher Scientific, Inc. (Waltham, MA, USA).
Ketamine hydrochloride was purchased from Fujian Gutian Yuanhang
Medical Company Ltd., Co. (Ningde, China). 17β-Estradiol,
pregnenolone (PREG), methyltestosterone (MT), dansyl chloride,
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
and dimethyl sulfoxide (DMSO) were purchased from Sigma-Aldrich
(St. Louis, MO, USA). Testosterone was provided by the National
Institute for Control of Pharmaceutical and Biological Products
(Beijing, China). Ethyl acetate-n-hexane was provided by Concord
Technology Co., Ltd. (Tianjin, China). Trypsin and Hoechst 33258
were purchased from Beijing Solarbio Science & Technology Co.,
Ltd. (Beijing, China). The in situ cell death detection kit
was from Roche Applied Science (Mannheim, Germany).
Primary culture of cortical neurons
Cortical neurons from rats were cultured as
previously described (15).
Briefly, cerebral cortices from newborn Sprague-Dawley rat pups
[obtained from Hebei Medical University (Shijiazhuang, China)],
<24 h old, were dissected and placed in ice-cold DMEM, then
mechanically dissociated and digested at 37°C for 15 min with
trypsin (1.25 g/l). The cells were distributed and seeded on
polylysine-coated plates (Beyotime Institute of Biotechnology,
Shanghai, China) in DMEM supplemented with 10% FBS at 37°C in a
humidified 5% CO2 air atmosphere incubator. DMEM was
replaced by neurobasal medium containing B27 supplement 24 h later.
From then, half the volume of culture medium was replaced with
fresh medium every other day. Neurons cultured for 7 days were used
for experimental purposes. The neurons were divided into four
groups as follows: Vehicle control group, treated with an equal
volume of neurobasal medium containing B27 supplement;
cholesterol-treated group, treated with 5 µM cholesterol;
ketamine-treated group, treated with 100 µM ketamine; and
cholesterol + ketamine-treated group, treated with 5 µM
cholesterol and 100 µM ketamine. All experiments were
performed in accordance with the Ethics Review Committee for Animal
Experimentation of Bethune International Peace Hospital of Chinese
PLA (Shijiazhuang, China).
Assessment of cell viability
The MTT assay was used to quantify the cell
viability, performed as previously described (5). In the current experiment, cells were
treated with ketamine (Fujian Gutian Yuanhang Medical Company Ltd.,
Co.) and/or cholesterol (Sigma-Aldrich) according to the
experimental design. Following treatment, 10 µl MTT (5
mg/ml) was added to each well and incubated for 4 h at 37°C. The
MTT culture medium was discarded and replaced with 200 µl
DMSO to dissolve the formazan crystals. The absorbance of each
sample was measured with a microplate reader (Molecular Devices
LLC, Sunnyvale, CA, USA) at 570 nm. Results were expressed as the
percentage of MTT reduction, assuming the absorbance of vehicle
control is 100%.
Apoptosis assays
Apoptosis was determined by using two different
assays. Firstly, apoptotic neurons were determined by the terminal
deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) method.
The TUNEL assay was performed using an in situ apoptosis
detection kit according to the manufacturer's instructions.
Briefly, neurons cultured on coverslips were rinsed with
phosphate-buffered saline [PBS (pH 7.2–7.4); Beijing noble Ryder
Technology Co., Ltd., Beijing, China) and fixed with 4%
paraformaldehyde (PFA) in PBS for 30 min at room temperature.
Terminal deoxynucleotidyl transferase (Thermo Fisher Scientific,
Inc., Waltham, MA, USA), a template-independent polymerase, was
used to incorporate nucleotides at the sites of DNA breaks. Nuclei
were stained with Invitrogen TO-PRO-3 [1:1,000 (1 µM) in
PBS; Thermo Fisher Scientific, Inc.). Then, the cells were
incubated with diaminobenzidine substrate (Shanghai Yesbio
Biological Technology Co., Ltd., Shangai, China) to produce a dark
brown precipitate. Apoptotic cells were identified according to the
following criteria: Condensed chromatin, reduced size and dark
brown nucleus. The TUNEL-positive and -negative cells were counted
in five randomly selected microscopic fields with a BX41 light
microscope (Olympus Corporation, Tokyo, Japan).
The second method used was Hoechst 33258 staining.
Neurons were stained with Hoechst 33258 nuclear dye according to
the manufacturer's instructions. Briefly, the cells were washed in
PBS, fixed with 4% PFA for 30 min at 4°C, then incubated with 10
µg/ml Hoechst 33258 for 8 min at room temperature and images
were captured under a BX41 fluorescence microscope (Olympus
Corporation). Morphological alterations in chromatin that are
characteristic of apoptosis, including condensation and
fragmentation, were observed. The percentage of positive cells was
calculated in five randomly selected microscopic fields using the
light microscope.
Quantification of neurosteroids
The samples (~1 ml) were subjected to MT (30 ng/ml,
internal standards) and were extracted three times with 2 ml ethyl
acetate-n-hexane (9:1, v/v). The organic phases were combined and
dried with a gentle stream of nitrogen in a water bath (50°C). The
samples were precipitated with dansyl chloride (Shanghai Huicheng
technology, Ltd., Shanghai, China) in a water bath (60°C, for 40
min), concentrated via centrifugation (16,000 × g, for 10 min at
4°C) and then transferred to autosampler vials prior to
high-performance liquid chromatography-tandem mass spectrometry
(HPLC-MS/MS) analysis.
The HPLC-MS/MS system (Thermo Fisher Scientific,
Inc.) consisted of a Surveyor MS Pump Plus, Surveyor AS Plus, TSQ
Quantum Access Triple-Quadrupole Mass Spectrometer and Xcalibur
software (version 2.1). Separation was achieved on an Eclipse XDB
C18 analytical column (4.6×50 mm; Agilent Technologies Inc, Palo
Alto, CA, USA) fitted with a XDB C18 guard column (4.6×12 mm). The
HPLC mobile phases were H2O/0.1% formic acid (solvent-A)
and MeOH/0.1% formic acid (solvent-B), and the gradient (flow rate
0.5 ml/min) was as follows: T0, 36, T6.5, 36,
T6.6, 30, T16, 10, T17, 10, and
T18, 36% solvent-A. The column temperature was 40°C, and
the injection volume was 30 µl. An MSD quadrupole mass
spectrometer equipped with an atmospheric pressure chemical
ionization source (Thermo Fisher Scientific, Inc.) was used for the
detection of analytes in the positive ion mode. The quantification
was performed using a multiple-reaction monitoring method with
transitions of 299.03-280.9 m/z for PREG, 254.97-158.9 m/z for
17β-estradiol, 289-97.2 m/z for testosterone and 303.1-109.06 m/z
for MT.
Statistical analysis
All data are expressed as the mean ± standard
deviation. Data analyses were performed with SPSS software, version
13.0 (SPSS, Inc., Chicago, IL, USA). Data were subjected to
statistical analysis using one-way analysis of variance followed by
the post-hoc Duncan's test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Cholesterol attenuates ketamine-induced
neuronal toxicity in cultured cortical neurons
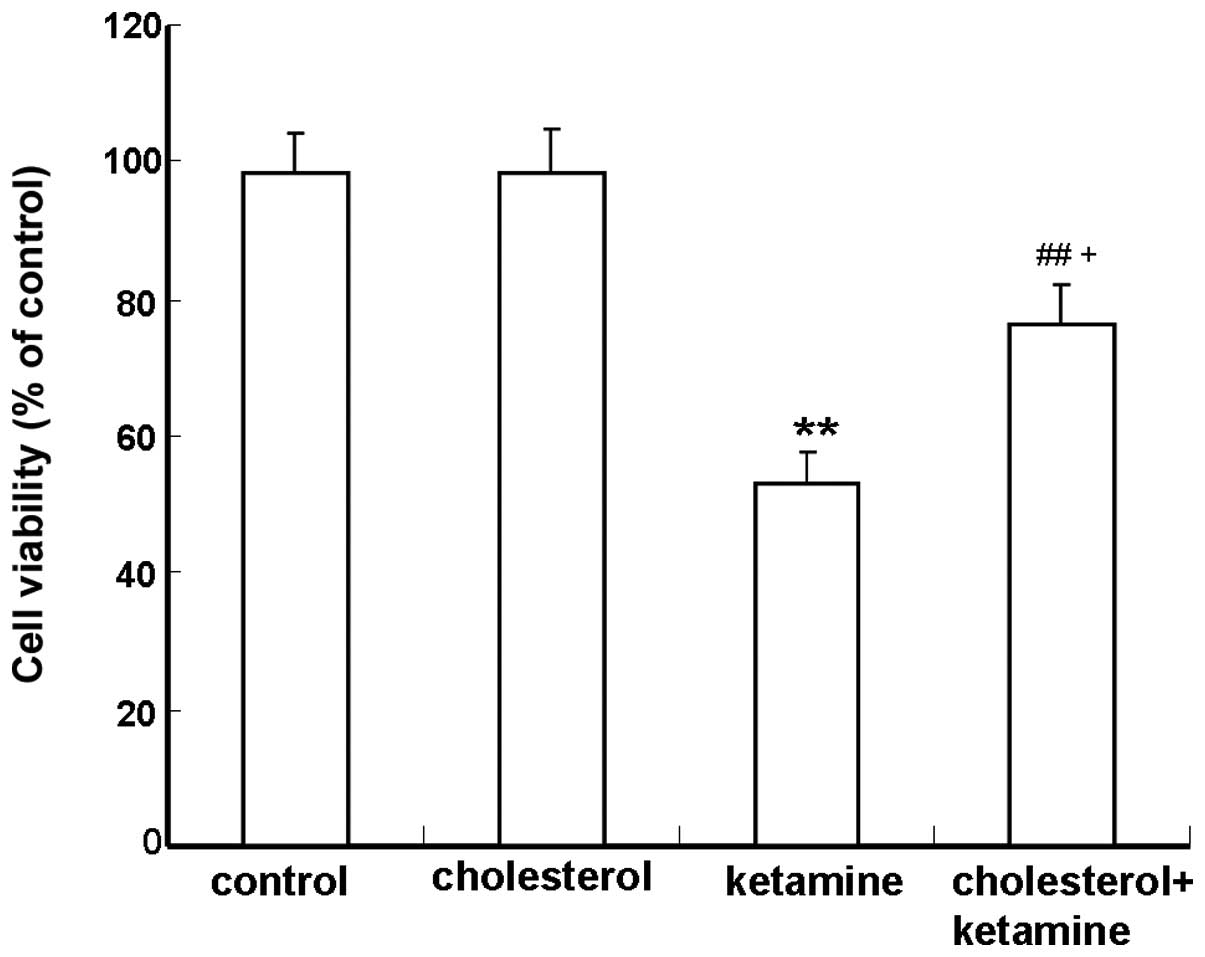
The MTT assay was performed to determine the
protective role of cholesterol against ketamine-induced neuronal
toxicity. Compared with the control group, neurons exposed to 100
µM ketamine exhibited a significant reduction in viability
(P<0.01; Fig. 1). The
combination of 100 µM ketamine and 5 µM cholesterol
treatments resulted in a significantly increased cell survival,
compared with the cholesterol-treated group (P<0.05; Fig. 1).
Cholesterol treatment reduces
ketamine-induced cell apoptosis in cultured cortical neurons
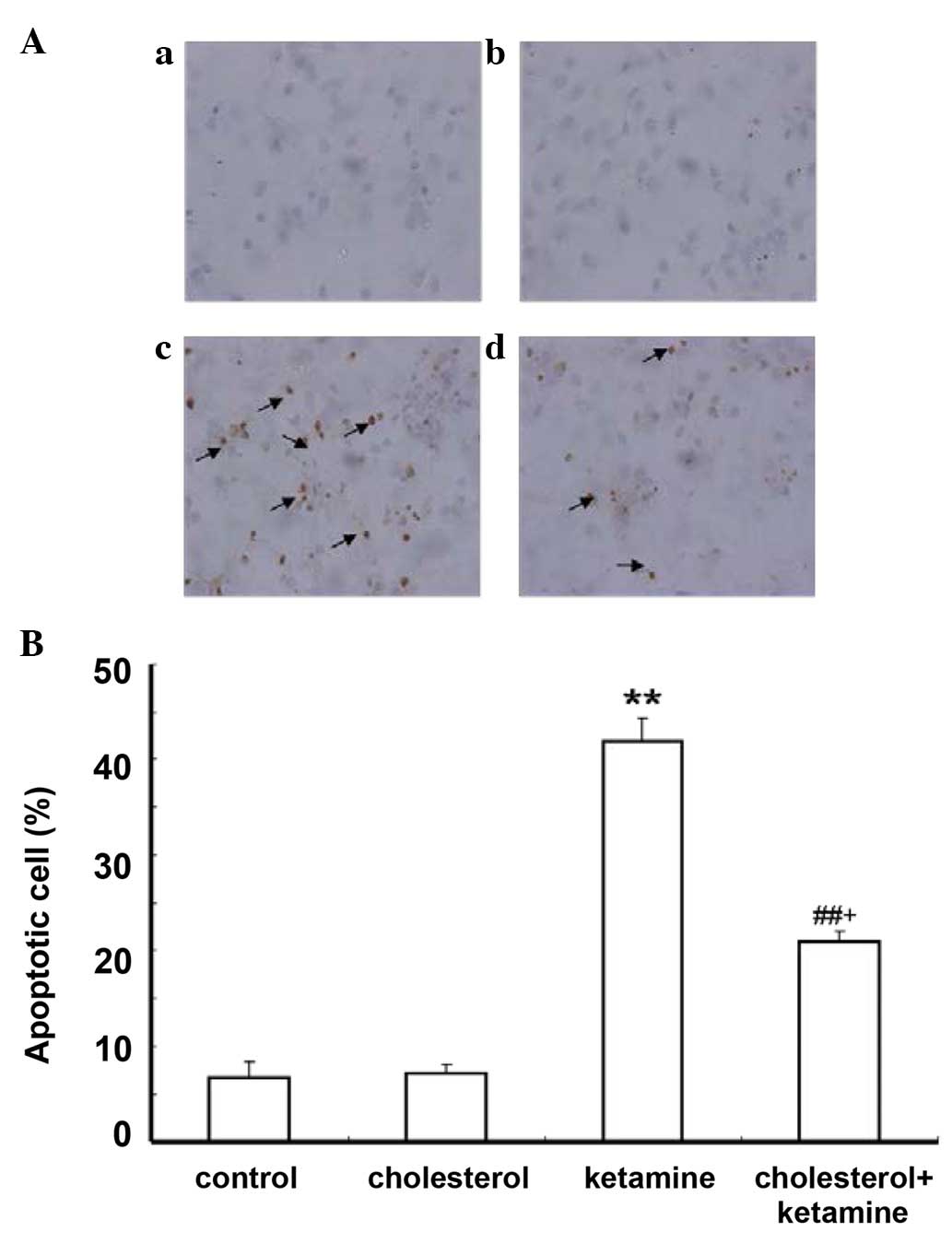
The TUNEL assay was utilized to detect whether the
ketamine-induced cell death is apoptotic. Apoptotic cells were
characterized by the appearance of intensely stained nuclei. The
results demonstrated that few apoptotic cells were present in the
control and cholesterol-treated groups (Fig. 2Aa and b, respectively), while the
number of apoptotic cells significantly increased in the
ketamine-treated group (Fig. 2Ac),
compared with the control group (P<0.01; Fig. 2B). Cholesterol treatment reduced
the number of apoptotic cells compared with the ketamine exposure
group (P<0.05; Fig. 2Ad and
B).
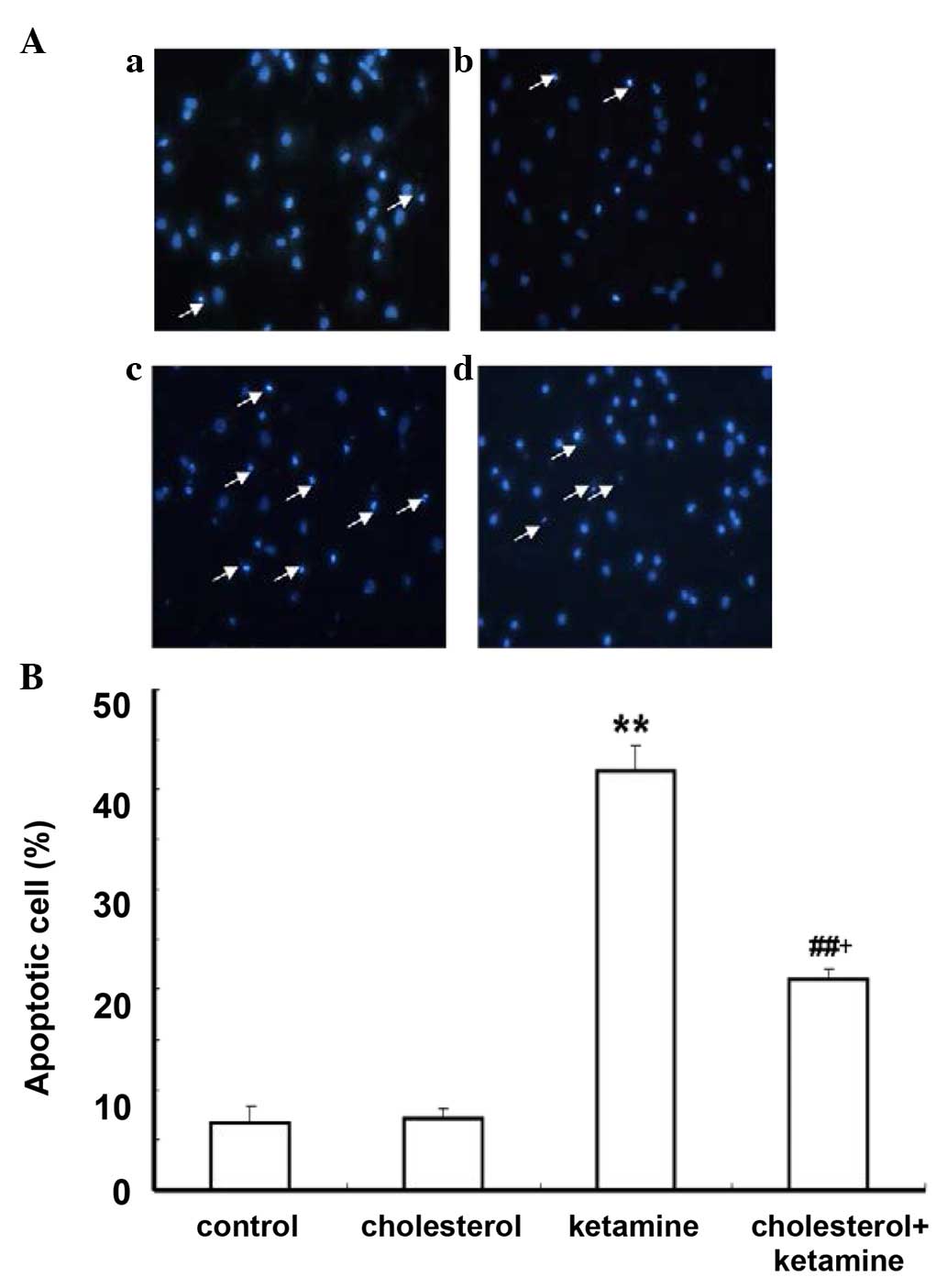
Hoechst 33258 staining was conducted to further
investigate the anti-apoptotic effect of cholesterol. As
demonstrated, in the control and cholesterol-treated groups, a
small number of apoptotic cells was present (Fig. 3Aa and b). The ketamine-treated
group (Fig. 3Ac) exhibited
increased numbers of apoptotic cells compared with the control
group (P<0.01; Fig. 3B). A
similar effect to the previous assay was observed for the
cholesterol and ketamine-treated group (Fig. 3A), compared with the
cholesterol-treated group (P<0.05; Fig. 3B).
Effect of ketamine on neurosteroid
biosynthesis under ketamine-induced neuroapoptosis condition in
primary cultured cortical neurons
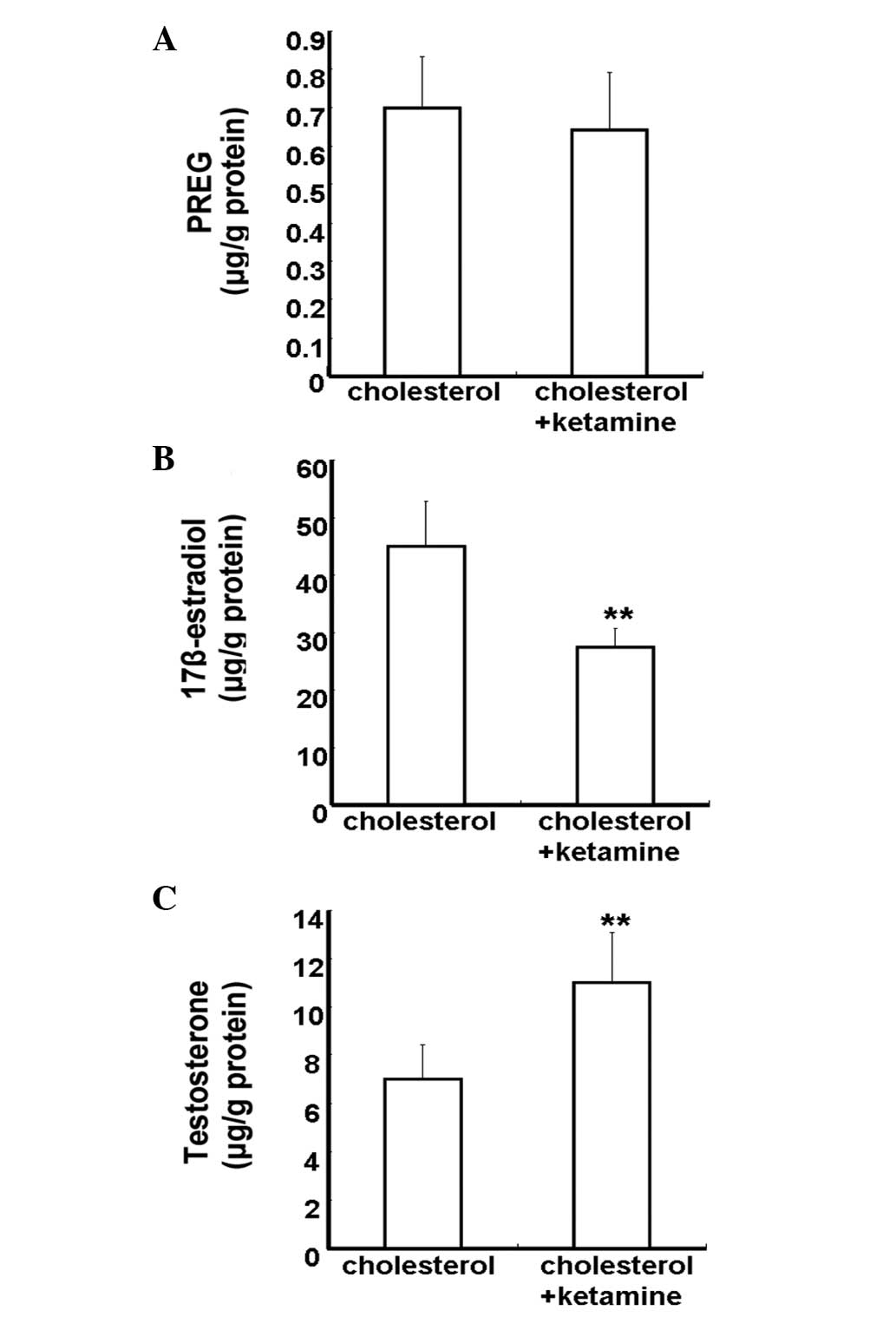
Neurosteroids are synthesized from cholesterol in
the CNS by means of a series of enzymatic processes, using the
pathway 'cholesterol to pregnenolone to dehydroepiandrosterone to
androstenedione to testosterone to estradiol' (16). To investigate the impact of
ketamine on neurosteroid biosynthesis, 5 µM cholesterol was
added into the culture medium. The concentration of neurosteroids
was detected by the HPLC-MS/MS assay. Neurosteroids were not
detected in the control and ketamine-treated groups, due to the
lack of a synthetic substrate for cholesterol (data not shown). The
cholesterol and cholesterol + ketamine-treated groups presented no
significant difference in PREG content levels (P>0.05; Fig. 4A). Compared with the
cholesterol-treated group, the 17β-estradiol content levels were
reduced in the cholesterol + ketamine group (P<0.01; Fig. 4B). In addition, the testosterone
content levels were increased in the dual treatment group, compared
with the cholesterol-only group (P<0.01; Fig. 4C). These results indicate that
ketamine is a potent suppressor of the conversion of testosterone
into 17β-estradiol.
Discussion
The results of the present study suggest that
cholesterol protects against ketamine-induced neuroapoptosis by
synthesizing certain neurosteroids. The results of the present
study indicate that cholesterol protects against ketamine-induced
neuro-apoptosis by synthesizing certain neurosteroids; in addition,
ketamine selectively regulates the pathway of neurosteroid
biosynthesis, which manifests in terms of a reduced level of
17β-estradiol and an increased level of testosterone. Data obtained
in the current study may provide the underlying mechanism of
ketamine-induced neuroapoptosis in primary cultured cortical
neurons and suggest an adjunctive neuroprotective measure for
ameliorating the toxicity of ketamine in the developing brain.
The effect of anesthetics on the developing brain is
of an important clinical and research interest. Although ketamine
is a widely used anesthetic, in certain instances, studies have
implicated that ketamine exposure can induce neuroapoptosis in the
developing brain (1,17,18).
In cases of severe damage, the developmental neuroapoptosis has
adverse effects on brain function and development as the animal
matures (2,3). Previously, numerous studies
investigated the mechanisms of, and protective strategies from
ketamine-induced neuroapoptosis (4,19,20).
It was postulated that the ketamine-induced neuroapoptosis may be
associated with a compensatory upregulation of the NMDA receptor
subunits and subsequent overstimulation of the glutamatergic system
by endogenous glutamate (3).
Although previous results have suggest a direct association between
blockade of NMDA receptors and neuroapoptosis (4), the underlying mechanisms remain
largely unclear.
Neurosteroids are synthesized de novo from
cholesterol in the brain (21).
Neurosteroids are produced locally in neuronal and glial cells,
which appear to modulate neurodevelopment by autocrine and/or
paracrine actions, and evidence indicates that neurosteroids are
neuroprotective and important during neurodevelopment (9). The imbalance of neurosteroid
androgens and estrogens is associated with the toxic effect of
MK801, another NMDA receptor blocker (11). A reduction of estradiol release
from astrocytes previously contributed to the neurodegeneration in
a model of Niemann-Pick disease type C (22). Previously, it was demonstrated that
ketamine attenuated 17β-estradiol levels in the early stages of
zebrafish life (13), and exerted
neurotoxic effects on the development of zebrafish embryos
(14). The main aim of the present
study was to investigate whether or not endogenous neurosteroid
synthesis was selectively regulated in primary cultured cortical
neurons during ketamine-induced neuroapoptosis. The results
suggested that cholesterol may protect against ketamine-induced
neuroapoptosis in vitro, which may be associated with the
conversion of cholesterol into certain neurosteroids, as
cholesterol is a steroid precursor and can increase the levels of
brain neurosteroids, including estradiol (16). Enzymes required for
neurosteroidogenesis are abundant in numerous brain regions
(23), which leads to the
possibility that cholesterol was locally converted into certain
neurosteroids. For example, 17β-estradiol can be synthesized in the
cortex region (24). Previous
studies suggested that continuous neurosteroid synthesis is
important for normal transmission in the hippocampus (25). In addition, cholesterol has been
suggested to be essential for synaptic activity (26,27)
and dendritic differentiation (28). Cholesterol may inhibit
stress-induced dendritic retraction through conversion into
17β-estradiol (29). One previous
study provided a similar conclusion; that chronic treatment with
17β-estradiol or cholesterol inhibited stress-induced hippocampal
CA3 dendritic retraction in ovariectomized female rats (30). In addition, another study
demonstrated that a high-cholesterol diet may remarkably increase
17β-estradiol serum levels and improve cognition deficits in
ovariectomized mice (31). Grewal
et al (32) reported that
carbamazepine exerted a neuroprotective effect against
ischemia-reperfusion injury, which was due to the increase in
synthesis of neurosteroids. Estrogens can exert neuroprotective
effects and enhance the survival of neurons (33). Previous studies demonstrated that
17β-estradiol protects against ketamine-induced neuroapoptosis in
primary cultured cortical neurons and in the developing rat brain
(3,5). These studies suggest that endogenous
neurosteroid biosyn-thesis, including synthesis of estradiol, may
be important to generate a protective and trophic environment for
cortical neurons.
Furthermore, the data suggested that the synthesis
of neurosteroid 17β-estradiol may be disrupted and 17β-estradiol
depletion contributes to the ketamine-induced neuroapoptosis in
primary cultured cortical neurons. A previous study demonstrated
that numerous pathogenic factors may induce neurodegenerative
injuries by downregulating the neuroprotective neurosteroid
biosynthesis in nerve cells (34).
A number of studies identified the levels of certain neurosteroids
in the brain to be markedly reduced in patients with AD and
Niemann-Pick disease type C (21,35,36).
Schaeffer et al (37)
reported that estradiol synthesis was selectively inhibited by
H2O2-treatment in SH-SY5Y cells and that
pretreatment with estradiol may protect against
H2O2-induced cell death. The present study
identified that ketamine-induced neuroapoptosis may be associated
with reduced endogenous 17β-estradiol secretion, and that treatment
with 17β-estradiol may protect against ketamine-induced
neuroapoptosis in the developing rat brain (3). Another study suggested that the
inhibition of neurosteroid estradiol biosynthesis resulted in the
reduction of long-term potentiation, dephosphorylation of cofilin
and resulting synapse loss (38).
In theory, testosterone can be converted into
estradiol through cytochrome p450 aromatase (16). Previous studies have demonstrated
that ketamine reduced cytochrome p450 aromatase activity (13,39).
However, the exact molecular mechanism underlying ketamine-induced
inhibition of the cytochrome p450 aromatase gene remains largely
unknown. In the brain, aromatase has been demonstrated to regulate
neural plasticity by stimulating cell growth and migration,
protecting against neurodegeneration and brain injury and
influencing learning and memory processes (40). Furthermore, a previous study
suggested that the inhibition of the cytochrome p450 aromatase
expression in the brain exacerbates neuronal death induced by
different forms of mild neurodegenerative stimuli (41). Morale et al (42) suggested that the inhibition of
cytochrome p450 aromatase function contributes to Parkinson's
disease. Others have demonstrated that female mice are infertile
when the aromatase gene is silenced, as their reproductive organs
cannot develop properly (43),
that the function of the amygdala and hypothalamus in these mice is
impaired (44) and that apoptosis
in the frontal cortex is increased (45). Numerous studies on aromatase
knockout mice have suggested that estrogen is important for
neurodevelopment (46). Another
study reported that aromatase suppression in hippocampal dispersed
cells inhibited the proliferation and induced apoptosis in granule
cells (47). One previous study
demonstrated that H2O2 induced SH-SY5Y cell
apoptosis by inhibiting aromatase activity and reducing endogenous
estradiol formation (37). Another
study suggested that an aromatase inhibitor had an effect on
cognitive performance in peripubertal boys who had been treated
over a period of two years with letrozole, a drug that inhibits
estrogen synthesis (48).
Furthermore, a previous study suggested that aromatase and
estradiol exerts a potential neuroprotective effect against kainic
acid-induced cytotoxicity (49–51).
The present study identified that among the neurosteroidogenic
pathways starting from the precursor, PREG and leading to the
distal metabolite, estradiol, only the estradiol biosynthesis was
markedly inhibited in ketamine-induced neuroapoptosis. These data
suggest that cytochrome p450 aromatase activity may be selectively
affected by ketamine, leading to the inhibition of estradiol
biosynthesis and inducing neuroapoptosis in primary cultured
cortical neurons. This should be further investigated to confirm
the inhibition of ketamine on cytochrome p450 aromatase
activity.
In conclusion, data from the present study provided
evidence that cholesterol may partially block ketamine-induced
neuroapoptosis in vitro by synthesizing numerous
neurosteroids. Additionally, ketamine reduced neurosteroid
17β-estradiol levels while elevating testosterone levels in primary
cultured cortical neurons. These data suggest that neurosteroid
17β-estradiol depletion may be associated with ketamine-induced
neuroapoptosis in primary cultured cortical neurons. Further
research is required to examine the potential neuroprotective
effects of neurosteroids, including 17β-estradiol, against
ketamine-induced neuroapoptosis in the developing brain, using a
well-designed animal model.
References
|
1
|
Mellon RD, Simone AF and Rappaport BA: Use
of anesthetic agents in neonates and young children. Anesth Analg.
104:509–520. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Brambrink AM, Evers AS, Avidan MS, Farber
NB, Smith DJ, Martin LD, Dissen GA, Creeley CE and Olney JW:
Ketamine-induced neuroapoptosis in the fetal and neonatal rhesus
macaque brain. Anesthesiology. 116:372–384. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li J, Wang B, Wu H, Yu Y, Xue G and Hou Y:
17β-estradiol attenuates ketamine-induced neuroapoptosis and
persistent cognitive deficits in the developing brain. Brain Res.
1593:30–39. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liu F, Patterson TA, Sadovova N, Zhang X,
Liu S, Zou X, Hanig JP, Paule MG, Slikker W Jr and Wang C:
Ketamine-induced neuronal damage and altered N-methyl-D-aspartate
receptor function in rat primary forebrain culture. Toxicol Sci.
131:548–557. 2013. View Article : Google Scholar :
|
|
5
|
Li J, Wu H, Xue G, Wang P and Hou Y:
17β-Oestradiol protects primary-cultured rat cortical neurons from
ketamine-induced apoptosis by activating PI3K/Akt/Bcl-2 signalling.
Basic Clin Pharmacol Toxicol. 113:411–418. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Melcangi RC, Garcia-Segura LM and
Mensah-Nyagan AG: Neuroactive steroids: State of the art and new
perspectives. Cell Mol Life Sci. 65:777–797. 2008. View Article : Google Scholar
|
|
7
|
Panzica GC, Balthazart J, Frye CA,
Garcia-Segura LM, Herbison E, Mensah-Nyagan AG, McCarthy MM and
Melcangi RC: Milestones on steroids and the nervous system: 10
years of basic and translational research. J Neuroendocrinol.
24:1–15. 2012. View Article : Google Scholar
|
|
8
|
Schumacher M, Hussain R, Gago N, Oudinet
JP, Mattern C and Ghoumari AM: Progesterone synthesis in the
nervous system: Implications for myelination and myelin repair.
Front Neurosci. 6:102012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hirst JJ, Kelleher MA, Walker DW and
Palliser HK: Neuroactive steroids in pregnancy: Key regulatory and
protective roles in the foetal brain. J Steroid Biochem Mol Biol.
139:144–153. 2014. View Article : Google Scholar
|
|
10
|
Mellon SH: Neurosteroid regulation of
central nervous system development. Pharmacol Ther. 116:107–124.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
De Olmos S, Bueno A, Bender C, Lorenzo A
and de Olmos J: Sex differences and influence of gonadal hormones
on MK801-induced neuronal degeneration in the granular
retro-splenial cortex of the rat. Brain Struct Funct. 213:229–238.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Caldeira JC, Wu Y, Mameli M, Purdy RH, Li
PK, Akwa Y, Savage DD, Engen JR and Valenzuela CF: Fetal alcohol
exposure alters neurosteroid levels in the developing rat brain. J
Neurochem. 90:1530–1539. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Trickler WJ, Guo X, Cuevas E, Ali SF,
Paule MG and Kanungo J: Ketamine attenuates cytochrome p450
aromatase gene expression and estradiol-17β levels in zebrafish
early life stages. J Appl Toxicol. 34:480–488. 2014. View Article : Google Scholar
|
|
14
|
Kanungo J, Cuevas E, Ali SF and Paule MG:
Ketamine induces motor neuron toxicity and alters neurogenic and
proneural gene expression in zebrafish. J Appl Toxicol. 33:410–417.
2013. View
Article : Google Scholar
|
|
15
|
Wang C, Sadovova N, Fu X, Schmued L,
Scallet A, Hanig J and Slikker W: The role of the
N-methyl-D-aspartate receptor in ketamine-induced apoptosis in rat
forebrain culture. Neuroscience. 132:967–977. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kawato S: Endocrine disrupters as
disrupters of brain function: A neurosteroid viewpoint. Environ
Sci. 11:1–14. 2004.
|
|
17
|
Hudson AE and Hemmings HC Jr: Are
anaesthetics toxic to the brain? Br J Anaesth. 107:30–37. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Vutskits L, Gascon E and Kiss JZ: Effects
of ketamine on the developing central nervous system. Ideggyogy Sz.
60:109–112. 2007.PubMed/NCBI
|
|
19
|
Liu F, Paule MG, Ali S and Wang C:
Ketamine-induced neurotoxicity and changes in gene expression in
the developing rat brain. Curr Neuropharmacol. 9:256–261. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Duan X, Li Y, Zhou C, Huang L and Dong Z:
Dexmedetomidine provides neuroprotection: impact on
ketamine-induced neuroapoptosis in the developing rat brain. Acta
Anaesthesiol Scand. 58:1121–1126. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Belelli D and Lambert JJ: Neurosteroids:
Endogenous regulators of the GABA(A) receptor. Nat Rev Neurosci.
6:565–575. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen G, Li HM, Chen YR, Gu XS and Duan S:
Decreased estradiol release from astrocytes contributes to the
neurodegeneration in a mouse model of Niemann-Pick disease type C.
Glia. 55:1509–1518. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nagarajan G, Aruna A and Chang CF:
Neurosteroidogenic enzymes and their regulation in the early brain
of the protogynous grouper Epinephelus coioides during gonadal sex
differentiation. Gen Comp Endocrinol. 181:271–287. 2013. View Article : Google Scholar
|
|
24
|
McCarthy MM: Estradiol and the developing
brain. Physiol Rev. 88:91–124. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tanaka M and Sokabe M: Continuous de novo
synthesis of neurosteroids is required for normal synaptic
transmission and plasticity in the dentate gyrus of the rat
hippocampus. Neuropharmacology. 62:2373–2387. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Suzuki S, Kiyosue K, Hazama S, Ogura A,
Kashihara M, Hara T, Koshimizu H and Kojima M: Brain-derived
neurotrophic factor regulates cholesterol metabolism for synapse
development. J Neurosci. 27:6417–6427. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Frank C, Rufini S, Tancredi V, Forcina R,
Grossi D and D'Arcangelo G: Cholesterol depletion inhibits synaptic
transmission and synaptic plasticity in rat hippocampus. Exp
Neurol. 12:407–414. 2008. View Article : Google Scholar
|
|
28
|
Goritz C, Mauch DH and Pfrieger FW:
Multiple mechanisms mediate cholesterol-induced synaptogenesis in a
CNS neuron. Mol Cell Neurosci. 29:190–201. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ortiz JB, McLaughlin KJ, Hamilton GF,
Baran SE, Campbell AN and Conrad CD: Cholesterol and perhaps
estradiol protect against corticosterone-induced hippocampal CA3
dendritic retraction in gonadectomized female and male rats.
Neuroscience. 246:409–421. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
McLaughlin KJ, Wilson JO, Harman J, Wright
RL, Wieczorek L, Gomez J, Korol DL and Conrad CD: Chronic
17beta-estradiol or cholesterol prevents stress-induced hippocampal
CA3 dendritic retraction in ovariectomized female rats: Possible
correspondence between CA1 spine properties and spatial
acquisition. Hippocampus. 20:768–786. 2010.
|
|
31
|
Li L, Xiao N, Yang X, Gao J, Ding J, Wang
T, Hu G and Xiao M: A high cholesterol diet ameliorates
hippocampus-related cognitive and pathological deficits in
ovariectomized mice. Behav Brain Res. 230:251–258. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Grewal AK, Jaggi AS, Rana AC and Singh N:
Effect of neurosteroid modulation on global
ischaemia-reperfusion-induced cerebral injury in mice. Korean J
Physiol Pharmacol. 17:485–491. 2013. View Article : Google Scholar
|
|
33
|
Fiocchetti M, Ascenzi P and Marino M:
Neuroprotective effects of 17β-estradiol rely on estrogen receptor
membrane initiated signals. Front Physiol. 3:732012. View Article : Google Scholar
|
|
34
|
Schaeffer V, Patte-Mensah C, Eckert A and
Mensah-Nyagan AG: Modulation of neurosteroid production in human
neuroblastoma cells by Alzheimer's disease key proteins. J
Neurobiol. 66:868–881. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Marx CE, Trost WT, Shampine LJ, Stevens
RD, Hulette CM, Steffens DC, Ervin JF, Butterfield MI, Blazer DG,
Massing MW, et al: The neurosteroid allopregnanolone is reduced in
prefrontal cortex in Alzheimer's disease. Biol Psychiatry.
60:1287–1294. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yue X, Lu M, Lancaster T, Cao P, Honda S,
Staufenbiel M, Harada N, Zhong Z, Shen Y and Li R: Brain estrogen
deficiency accelerates Abeta plaque formation in an Alzheimer's
disease animal model. Proc Natl Acad Sci USA. 102:19198–19203.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Schaeffer V, Patte-Mensah C, Eckert A and
Mensah-Nyagan AG: Selective regulation of neurosteroid biosynthesis
in human neuroblastoma cells under hydrogen peroxide-induced
oxidative stress condition. Neuroscience. 151:758–770. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Vierk R, Brandt N and Rune GM: Hippocampal
estradiol synthesis and its significance for hippocampal synaptic
stability in male and female animals. Neuroscience. 274:24–32.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lupp A, Kerst S and Karge E: Evaluation of
possible pro- or antioxidative properties and of the interaction
capacity with the microsomal cytochrome P450 system of different
NMDA-receptor ligands and of taurine in vitro. Exp Toxicol Pathol.
54:441–448. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lephart ED, Adlercreutz H and Lund TD:
Dietary soy phytoestrogen effects on brain structure and aromatase
in Long-Evans rats. Neuroreport. 12:3451–3455. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Garcia-Ovejero D, Azcoitia I, Doncarlos
LL, Melcangi RC and Garcia-Segura LM: Glia-neuron crosstalk in the
neuroprotective mechanisms of sex steroid hormones. Brain Res Brain
Res Rev. 48:273–286. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Morale MC, L'Episcopo F, Tirolo C,
Giaquinta G, Caniglia S, Testa N, Arcieri P, Serra PA, Lupo G,
Alberghina M, et al: Loss of aromatase cytochrome P450 function as
a risk factor for Parkinson's disease? Brain Res Brain Res Rev.
57:431–443. 2008. View Article : Google Scholar
|
|
43
|
Simpson ER: Models of aromatase
insufficiency. Semin Reprod Med. 22:25–30. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Pierman S, Sica M, Allieri F,
Viglietti-Panzica C, Panzica GC and Bakker J: Activational effects
of estradiol and dihydrotestosterone on social recognition and the
arginine-vasopressin immunoreactive system in male mice lacking a
functional aromatase gene. Horm Behav. 54:98–106. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hill RA, Simpson ER and Boon WC: Evidence
for the existence of an estrogen-responsive sexually dimorphic
group of cells in the medial preoptic area of the 129SvEv mouse
strain. Int J Impot Res. 20:315–323. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Sasahara K, Shikimi H, Haraguchi S,
Sakamoto H, Honda S, Harada N and Tsutsui K: Mode of action and
functional significance of estrogen-inducing dendritic growth,
spinogenesis, and synaptogenesis in the developing Purkinje cell. J
Neurosci. 27:7408–7417. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Fester L, Ribeiro-Gouveia V, Prange-Kiel
J, Von Schassen C, Böttner M, Jarry H and Rune GM: Proliferation
and apoptosis of hippocampal granule cells require local oestrogen
synthesis. J Neurochem. 97:1136–1144. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Hero M, Maury S, Luotoniemi E, Service E
and Dunkel L: Cognitive effects of aromatase inhibitor therapy in
peripubertal boys. Eur J Endocrinol. 163:149–155. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Garcia-Segura LM, Veiga S, Sierra A,
Melcangi RC and Azcoitia I: Aromatase: A neuroprotective enzyme.
Prog Neurobiol. 71:31–41. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Azcoitia I, Sierra A, Veiga S, Honda S,
Harada N and Garcia-Segura LM: Brain aromatase is neuroprotective.
J Neurobiol. 47:318–329. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Veiga S, Azcoitia I and Garcia-Segura LM:
Extragonadal synthesis of estradiol is protective against kainic
acid excitotoxic damage to the hippocampus. Neuroreport.
16:1599–1603. 2005. View Article : Google Scholar : PubMed/NCBI
|