Introduction
Pelvic organ prolapse (POP) is a global health
problem, which adversely affects 50% of women >50 years old, in
terms of poor life quality and increasing economic burden (1–3), and
has become one of the most common indications for gynecological
surgery in elderly women (4).
However, the etiology and pathophysiological mechanisms remain to
be fully elucidated. POP is considered to be a multifactorial
etiological disease (5,6). Aging and vaginal childbirth are well
established risk factors, and other factors, including obesity,
chronic constipation and declined hormonal status (menopause), have
also been reported to be associated with prolapse (6). Previous studies have demonstrated
pelvic muscle injuries (7), loss
of extracellular matrix (ECM) (8–10)
and hyperfunction of matrix metalloproteinases (MMPs) (11,12),
together with upregulated cell apoptosis and downregulated
proliferation in pelvic connective tissue in women with POP
(13). The loss of ECM proteins is
considered to be the molecular basis facilitating the incidence and
development of POP (10).
OS reflects a status of imbalance, in which
excessive accumulated reactive oxygen species (ROS) exceeds the
neutralizing ability of the redox system in cells, resulting in
oxidative damage to cell organisms, including deoxyribonucleic acid
(DNA), proteins and lipids. In addition, ROS act as cellular
messengers modulating specific signaling pathways (14). Previous studies have suggested that
OS is involved in cardiovascular remodeling and fibrotic disorders
by activating MMPs, and reducing collagen synthesis in fibroblasts
and smooth muscle cells (15–17).
Thus, the present study hypothesized that OS may contribute to
collagen metabolic disorder in the pathogenesis of POP. If this is
the case, it may assist in developing novel clinic therapeutic
strategies.
In the present study, this hypothesis was tested
through a number of experiments. Preliminarily, specimens of human
uterosacral ligament (USL) were collected to identify the oxidative
damage, and assess whether there are differences between women with
and without POP. Furthermore, the cytotoxic effect of hydrogen
peroxide (H2O2) on fibroblasts derived from
normal human USL tissues was evaluated. A cell model was also
established to mimic intracellular OS, in order to investigate the
exact role of OS in collagen metabolism and the associated
mechanisms.
Materials and methods
Patient grouping and sample
collection
The study was approved by the ethics committee of
Renim Hospital of Wuhan University (Wuhan, China), and each
participant provided written informed consent, according to the
Declaration of Helsinki (18). A
total of 56 menopausal women were enrolled from the Department of
Obstetrics and Gynecology, Renmin Hospital of Wuhan University
between March 2011 and June 2013. Menopause was defined as the
cessation of menses for at least 1 year. The study group consisted
of 20 women, who underwent hysterectomy as part of pelvic
reconstruction surgery for POP stage II, III or IV, according to
POP quantitative examination (POP-Q) (19). In addition, 20 women without POP
(non-POP group), who underwent hysterectomy for benign indications,
including cervical intraepithelial neoplasia and dysfunctional
uterine bleeding, served as the control group. Another 16 non-POP
cases were used to develop primary and passage cultures of human
uterosacral ligamental fibroblasts (hUSLFs). In order to minimize
the effects of confounding factors on the expression of OS
biomarkers, women who had pelvic infectious diseases, leiomyoma,
adenomyosis, endometriosis, history of pelvic surgery, malignant
gynecological tumors, systemic autoimmune diseases, cardiovascular
diseases, diabetes mellitus or other endocrine disorders, and
neuromuscular degenerative diseases were excluded from either
group. In addition, women who received hormone replacement therapy
or antioxidant supplementation were not included in either
group.
Tissue sample biopsies measuring 0.5×1.0 cm were
obtained from the USL, close to the cervix, during surgery,
following which the specimens were prepared according to the
respective following protocols for the different
investigations.
Immunohistochemistry
The tissue specimens were fixed with 4%
paraformaldehyde and embedded in paraffin (both purchased from
Maxim Biotechnology Development Co., Ltd., Fuzhou, China), and then
sliced into 4 µm sections for immunohistochemical staining.
An Elivision™ super HRP IHC kit (cat. no. Kit-9921; Fuzhou Maxim
Biotechnology Development Co., Ltd.), was used for
immunohistochemistry. The paraffin-embedded sections were incubated
for 30 min at 60°C, and then deparaffinized and rehydrated in a
graded alcohol series. Antigen retrieval was performed by heat
mediation in citrate buffer (pH<6) or EDTA buffer (pH>9),
according to the product datasheets for the primary antibodies. The
sections were then incubated with 3% H2O2
(Maxim Biotechnology Development Co., Ltd.) for 10 min at room
temperature to inactivate endogenous peroxidase, and blocked with
5% goat serum for 15 min. The sections were then incubated with
primary antibodies overnight at 4°C. Following incubation with
streptavidin peroxidase (Maxim Biotechnology Development Co., Ltd.)
for 10 min at room temperature, secondary antibodies were added for
30 min at 37°C. The immune reaction was then visualized using DAB
(Fuzhou Maxim Biotechnology Development Co., Ltd., Fuzhou, China).
The specimens were washed with phosphate-buffered saline (PBS)
following each step in the protocol. For the negative controls, the
primary antibody was replaced with PBS. The following antibodies
were used: Mouse monoclonal anti-8-hydroxyguanosine (8-OHdG)
antibody (1:100 dilution; ab62623; Abcam, Cambridge, UK), rabbit
polyclonal anti-4-hydroxynonenal (4-HNE) antibody (1:200 dilution;
ab46545; Abcam) and rabbit polyclonal anti-TGF-β1 antibody (1:100
dilution; ab53169; Abcam). Secondary antibodies included goat
anti-rabbit polyclonal horseradish peroxidase (HRP)-conjugated IgG
(1:1,000 dilution; ab97051; Abcam) and goat anti-mouse polyclonal
HRP-conjugated IgG (1:1,000 dilution; ab97023; Abcam).
Immunoreactivity was quantified by the integrated optical density
value, obtained from immunohistochemical images using Image-Pro
Plus 6.0 software.
Primary cell culture and passaging
hUSLFs were developed from the fresh USL biopsy
tissues, according to Gibco (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) protocols for primary culture. Briefly, the
tissue samples were cut into small sections (~1 mm3) and
washed twice in PBS, followed by digestion with collagenase I
(Invitrogen; Thermo Fisher Scientific, Inc.) and trypsase
(Sigma-Aldrich, St. Louis, MO, USA) successively. The isolated
cells were routinely cultured in Dulbecco's modified Eagle's medium
(GE Healthcare Life Sciences. Logan, UT, USA) supplemented with 10%
fetal bovine serum, 1% penicillin (100 U/ml) and streptomycin (100
ug/ml) (Gibco; Thermo Fisher Scientific, Inc.) in a 5%
CO2-humidified atmosphere at 37°C. The cells were
subcultured at a confluence of 80%. To identify the cells as
fibroblasts, immunocytochemical analyses were performed.
Cytokeratin is a specific biomarker for epithelial original cells,
while vimentin is a specific biomarker for mesenchymal original
cells (20). Cells were seeded
into a six-well plate containing a pre-placed coverslip at a
density of 1×105 cells/ml, and incubated at 37°C for 48
h. The cells were harvested at 70% confluence, washed with PBS and
fixed in 4% paraformaldehyde for 20 min at room temperature, and
then permeabilized using 0.3% Triton X100 buffer (Sigma-Aldrich)
for 10 min at room temperature. After three washes with PBS, the
cells were incubated with 3% H2O2 for 10 min
at room temperature to inactivate the endogenous peroxidase. After
being washed thrice with PBS, the slides were blocked with 1%
bovine serum albumin (Sigma-Aldrich) in PBS for 30 min at room
temperature. Next, the cells were incubated with mouse monoclonal
vimentin (1:200 dilution; sc6260; Santa Cruz Biotechnology, Inc.,
Santa Cruz, CA, USA), and mouse monoclonal cytokeratin-19 (1:200
dilution; sc6278; Santa Cruz Biotechnology, Inc.) antibodies
overnight at 4°C followed by three washes in PBS. Subsequently, the
cells were incubated with secondary antibodies (k5007; Dako,
Glostrup, Denmark) at room temperature for 1 h followed by three
washes in PBS, and visualized using 3,3′-diaminobenzidine (k5007;
Dako). The immunocytochemical staining slides were evaluated using
a microscope (BX51; Olympus Corporation, Tokyo, Japan).
Cell Counting Kit-8 (CCK-8) assay for
assessment of cytotoxicity
Cytotoxicity was measured using a CCK-8 assay
(Beyotime Institute of Biotechnology, Haimen, China). Cells
suspended with Dulbecco's modified Eagle medium (100 µl;
Gibco; Thermo Fisher Scientific) were inoculated into a 96-well
plate (5,000 cells/well) and incubated for 24 h at 37°C, following
which the spent culture medium was removed and 100 µl of
prepared media containing various concentrations of
H2O2 (0, 0.2, 0.4, 0.8 and 1.6 mM) was added.
Each sample was loaded in triplicate. Following incubation at 37°C
for 4 and 24 h respectively, 10 µl CCK-8 solution was added
into each well and incubated at 37°C for 4 h. The absorbance was
measured at 450 nm using a microplate spectrophotometer (Victor3
1420 Multilable Counter; PerkinElmer, Inc., Waltham, MA, USA). The
viability of the treated group was expressed as a percentage of the
untreated control group, which was designated as 100%.
Flow cytometric assay for cell
apoptosis
Annexin V-fluorescein isothiocyanate
(FITC)/propidium iodide (PI) double staining was used to assay cell
apoptosis, in strict accordance with the manufacturer's protocols
(Beyotime Institute of Biotechnology). In brief, the cells were
resuspended at a density of 1×106/ml and incubated with
10 µl Annexin V-FITC at room temperature for 30 min in the
dark. Subsequently, 5 µl PI was added for 5 min, following
which 400 µl 1X binding buffer was added into each tube. A
flow cytometer (BD FACSCalibur; BD Biosciences, San Jose, CA, USA)
was finally used to detect the labeled cells.
Direct immunofluorescent assay for
intracellular ROS
Following exposure to different concentrations of
H2O2, The cells were incubated with 10
µM 2′,7′-dichlorofluorescein diacetate (DCF-DA; Applygen
Technologies, Inc., Beijing, China), a peroxide sensitive
fluorescent probe, for 30 min at 37°C, according to the
manufacturer's protocol. Images were captured with a fluorescent
microscope (BC51; Olympus Corporation) and analyzed using Image-pro
Plus 6.0 software (Media Cybernetics, Inc., Rockville, MD,
USA).
Indirect immunofluorescent assay for
intracellular 8-OHdG
Following treatment with H2O2
solution or the placebo, the plated cells were fixed and incubated
with mouse monoclonal anti-8-OHdG (1:100 dilution; ab62623; Abcam)
overnight at 4°C, and were then successively incubated with
fluorescent-labeled goat anti-mouse polyclonal IgG secondary
antibody (1:500 dilution; ab150117; Abcam) and DAPI (2
µg/ml; Beyotime Institute of Biotechnology) at room
temperature for 20 min, according to routine protocols. The cells
were observed under a fluorescent microscope (Olympus Corporation)
and analyzed using Image-pro Plus 6.0 software.
Western blot analysis
Total protein was extracted from the USL cells using
radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology) containing phenylmethylsulfonyl fluoride, and the
quantity was determined using a Bicinchoninic Acid Protein Assay
kit (Beyotime Institute of Biotechnology). The protein samples (20
µg) were separated by electrophoresis on SDS-PAGE gels (10%;
Beyotime Institute of Biotechnology) and transferred onto
polyvinylidene fluoride (PVDF) membranes (EMD Millipore, Billerica,
MA, USA). Following blocking, the membranes were incubated
overnight at 4°C with diluted primary antibody. The PVDF membranes
were then washed three times with Tris-buffered saline with 0.1%
Tween 20 (Beyotime Institute of Biotechnology), and the blots were
incubated with IRDye 800CW goat anti-rabbit/mouse secondary
antibodies (1:10,000 dilution; LI-COR Biosciences, Lincoln, NE,
USA) at room temperature for 1 h. Finally, the protein intensity
were scanned as a fluorescent signal and quantified using an
Odyssey imaging system (LI-COR Biosciences, Lincoln, NE, USA).
Experiments were performed in triplicate. The following primary
antibodies were used: Polyclonal rabbit collagen, type 1, α1
(COL1A1) antibody (1:400 dilution, sc-8784-R), polyclonal rabbit
MMP-2 antibody (1:400 dilution; sc-10736), polyclonal rabbit TIMP-2
antibody (1:500 dilution; sc-5539) (all purchased from Santa Cruz
Biotechnology, Inc.) and polyclonal rabbit TGF-β1 antibody (1:1,000
dilution; ab53169; Abcam). GAPDH antibody (1:1000 dilution;
sc-25778; Santa Cruz Biotechnology, Inc.) served as an endogenous
reference.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted using TRIzol regent
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. cDNA synthesis was performed with 2
µg of total RNA in a reaction volume of 20 µl using a
RevertAid First Strand cDNA Synthesis kit (Fermentas; Thermo Fisher
Scientific, Inc.) containing 6 µl total RNA, 1 µl
oligo 18 primer, 5 µL diethylpyrocarbonate water, 4
µl 5X reaction buffer, 1 µl Ribolock RNAse inhibitor,
2 µl 10 mM dNTP and 1 µl RevertAid M-MuLV Reverse
Transcriptase. SYBR Green labeled probes (Takara Bio, Inc., Tokyo,
Japan) were used to detect gene expression levels in an ABI 7500
Real-Time PCR system (Applied Biosystems; Thermo Fisher Scientific,
Inc.). The expression levels of the target mRNAs were calculated
and normalized to the mRNA level of GAPDH. The following primers
(Sangon Biotech Co., Ltd., Shanghai, China) were used: GAPDH (used
for normalization), forward 5′-GAAGGTGAAGGTCGGAGT C-3′ and reverse
5′-GAAGATGGTGATGGGATTTC-3′; COL1A1, forward
5′-CAAGACGAAGACATCCCACCAATC-3′ and reverse
5′-ACAGATCACGTCATCGCACAACA-3′; MMP-2, forward
5′-AGTTTCCATTCCGCTTCCAG-3′ and reverse 5′-CGGTCGTAGTCCTCAGTGGT-3′;
TIMP-2, forward 5′-TCTGGAAACGACATTTATGG-3′ and reverse
5′-GTTGGAGGCCTGCTTATGGG-3′; and TGF-β1, forward
5′-TATTGAGCACCTTGGGCACT-3′ and reverse 5′-ACCTCTCTGGGCTTGTTTCC-3′.
The thermal cycling conditions were as follows: 30 Sec at 95°C,
followed by 40 cycles of 5 sec at 95°C and 34 sec at 60°C, and
finally 15 sec at 95°C, 1 min at 60°C, 15 sec at 95°C and 15 sec at
60°C. mRNA levels were subsequently quantified using the
2−ΔΔCq method (21).
Statistical analysis
The experiments were repeated at least three times
and data are partially presented as the mean ± standard deviation.
A Mann-Whitney U test was used for comparison between two
groups, One-way/two-way analysis of variance (ANOVA) was used to
analyze the statistical difference among groups. GraphPad Prism
5.01 (GraphPad Software, Inc.) and SPSS 16.0 (SPSS, Inc., Chicago,
IL, USA) were used for statistical analyses and graph plotting.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Demographic and clinical
characteristics
All 40 subjects among the two groups were well
matched in terms of demographic and clinical characteristics. No
significant differences in age, parity, body mass index or
postmenopausal duration were observed between the POP group and the
control group (Table I).
 | Table IDemographic and clinical
characteristics of all participants. |
Table I
Demographic and clinical
characteristics of all participants.
| Parameter | POP group
(n=20) | Control group
(n=20) | P-value |
|---|
| Age (years) | 55.1±4.9 | 53.6±4.1 | NSa |
| Parity | 2.2±1.3 | 1.9±1.1 | NSa |
| Body mass index
(Kg/m2) | 26.2±5.3 | 25.2 ±4.9 | NSa |
| Postmenopausal
duration (months) | 58.6±8.9 | 60.3±9.4 | NSa |
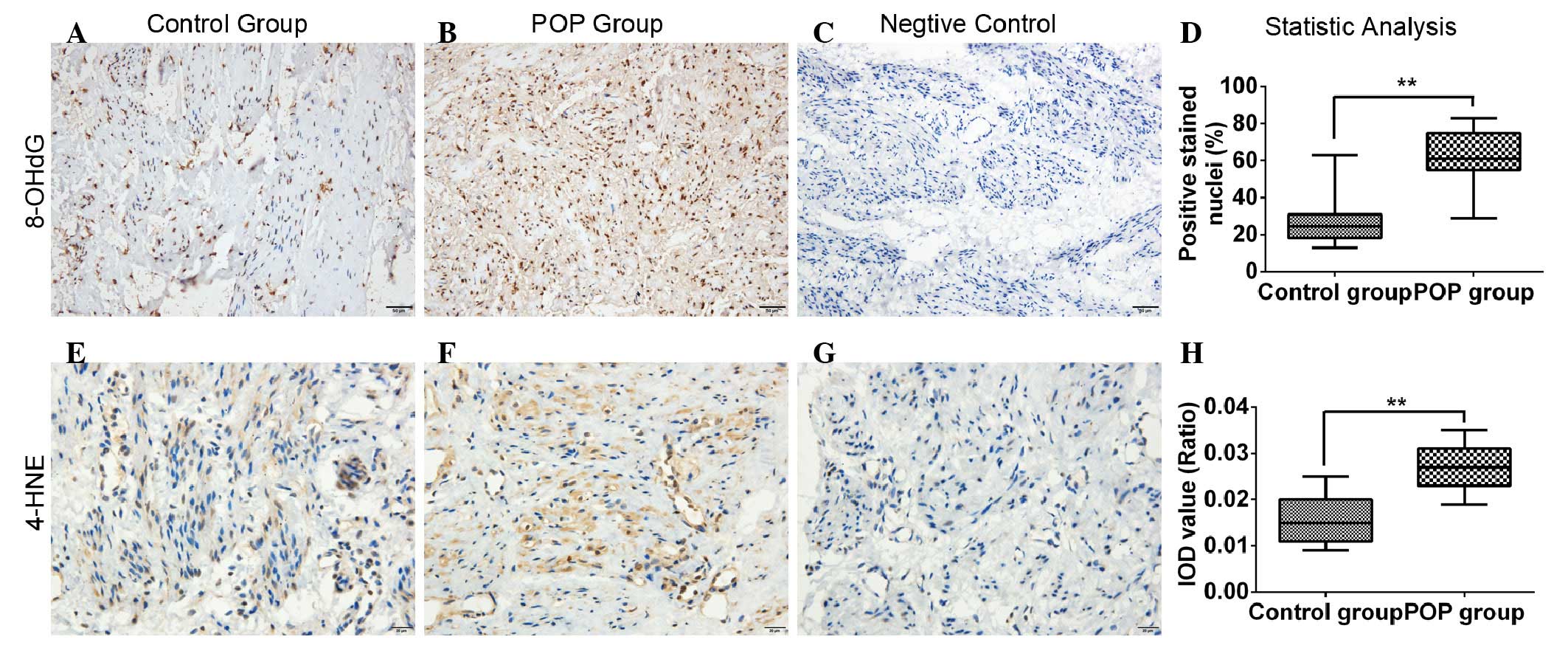
Expression of OS biomarkers are
upregulated in POP USL tissues
Through immunohistochemical investigations, the
present study found that the immunoreactivity of 8-OHdG in the POP
group was significantly higher, compared with that in the control
group (0.625±0.145, vs. 0.263±0.117, respectively; P<0.01). The
same results were observed for 4-HNE between the POP and control
groups (0.027±0.006, vs. 0.016±0.006, respectively; P<0.01;
Fig. 1).
Cytotoxic effects are induced by
exogenous H2O2 in fibroblasts
To evaluate the cytotoxicity of exogenous
H2O2 on the hUSLFs, the fibroblasts were
treated with graded H2O2 at concentrations of
0, 0.1, 0.2, 0.4, 0.6, 0.8 and 1.0 mM for 2, 4, 6, 8, 12 and 24 h,
respectively, following which cell viability was examined using a
CCK-8 assay. As shown in Fig. 2,
cell viability decreased following H2O2
treatment in time-dependent and concentration-dependent manner.
Based on two-way ANOVA, no statistically significant difference was
observed between the 0.1 mM group and the untreated control group.
At concentrations >0.2 mM, the cell viability in the treated
groups were significantly different from that of the control group
(P<0.05; Fig. 2A). Following
comparison of the cell viability at 4 h (Fig. 2B) with that of 24 h (Fig. 2C), significant inter-group
differences were found among the groups with concentrations of 0.1,
0.2 and 0.4 mM (one-way ANOVA; P<0.05). Additionally, following
treatment for 24 h, concentrations ≥0.4 mM led to a significant
decrease in cell viability, however, no significant differences
were observed at the 0.4, 0.6, 0.8 and 1.0 mM concentrations. The
above data suggested that 0.1 mM may be a sub-toxic dose, which
induces no significant cytotoxicity, whereas 0.2 mM is mildly to
moderately toxic and concentrations ≥0.4 mM are severely toxic in a
treatment duration of 24 h. Immunocytochemical analyses
demonstrated that the cells were anti-vimentin-positive and
anti-keratin-negative.
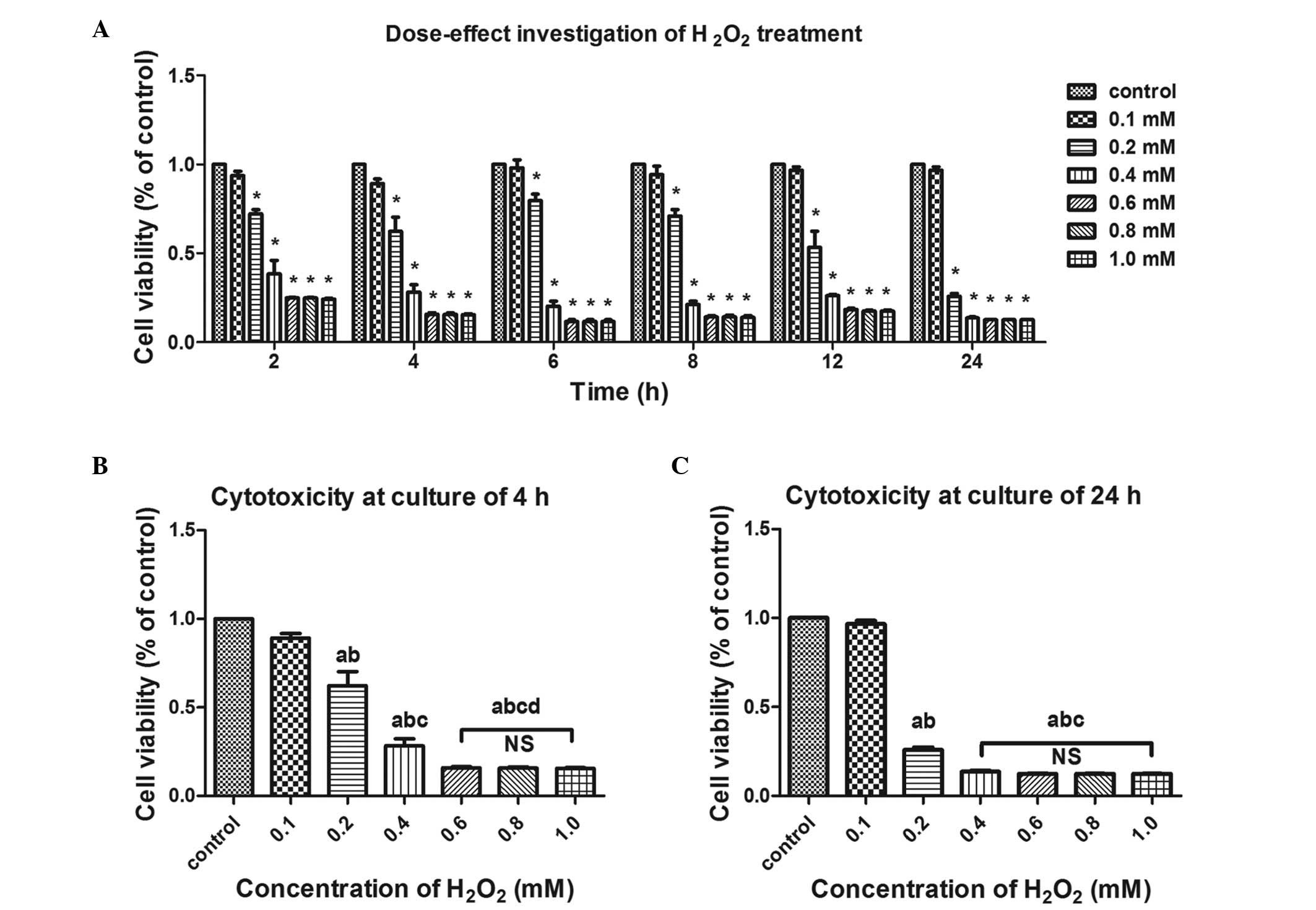 | Figure 2Effects of exogenous
H2O2 on the cell viability of human
uterosacral ligament fibroblasts. Cell viability was examined using
a Cell Counting Kit-8 assay and data are presented as apercentage
of the untreated control group. (A) Cells were treated with the
indicated concentrations of H2O2 for 2, 4, 6,
8, 12 and 24 h for determination of concentration-dependence. Cell
viability decreased in a concentration-dependent and time-dependent
manner under H2O2 exposure.
*P<0.05, vs. control group (two-way ANOVA. The cells
were treated with the indicated concentrations of
H2O2 for (B) 4 h and (C) 24 h for
determination of concentration-dependence. One-way ANOVA was
performed, and data are presented as the median ± standard error of
the mean (n=3). a, P<0.05, vs. unstreated control; b, P<0.05,
vs. 0.1 mM; c, P<0.05, vs. 0.2 mM; d, P<0.05, vs. 0.4 mM; NS,
no significance; ANOVA, analysis of variance;
H2O2, hydrogen peroxide. |
Cell apoptosis is induced by
H2O2 treatment
Based on the results of the above cytotoxicity
investigations, the hUSLFs cells were treated for 24 h with
H2O2 at concentrations ranging between 0.1
and 0.6 mM, to investigate the effects on cell apoptosis. As shown
in Fig. 3, as the concentration of
H2O2 increased, the apoptotic rates increased
gradually. No significant difference was observed between the
control group and the 0.1, 0.4 or 0.6 mM groups, however, there
were significant differences between the 0.2 mM group and the other
groups (P<0.05).
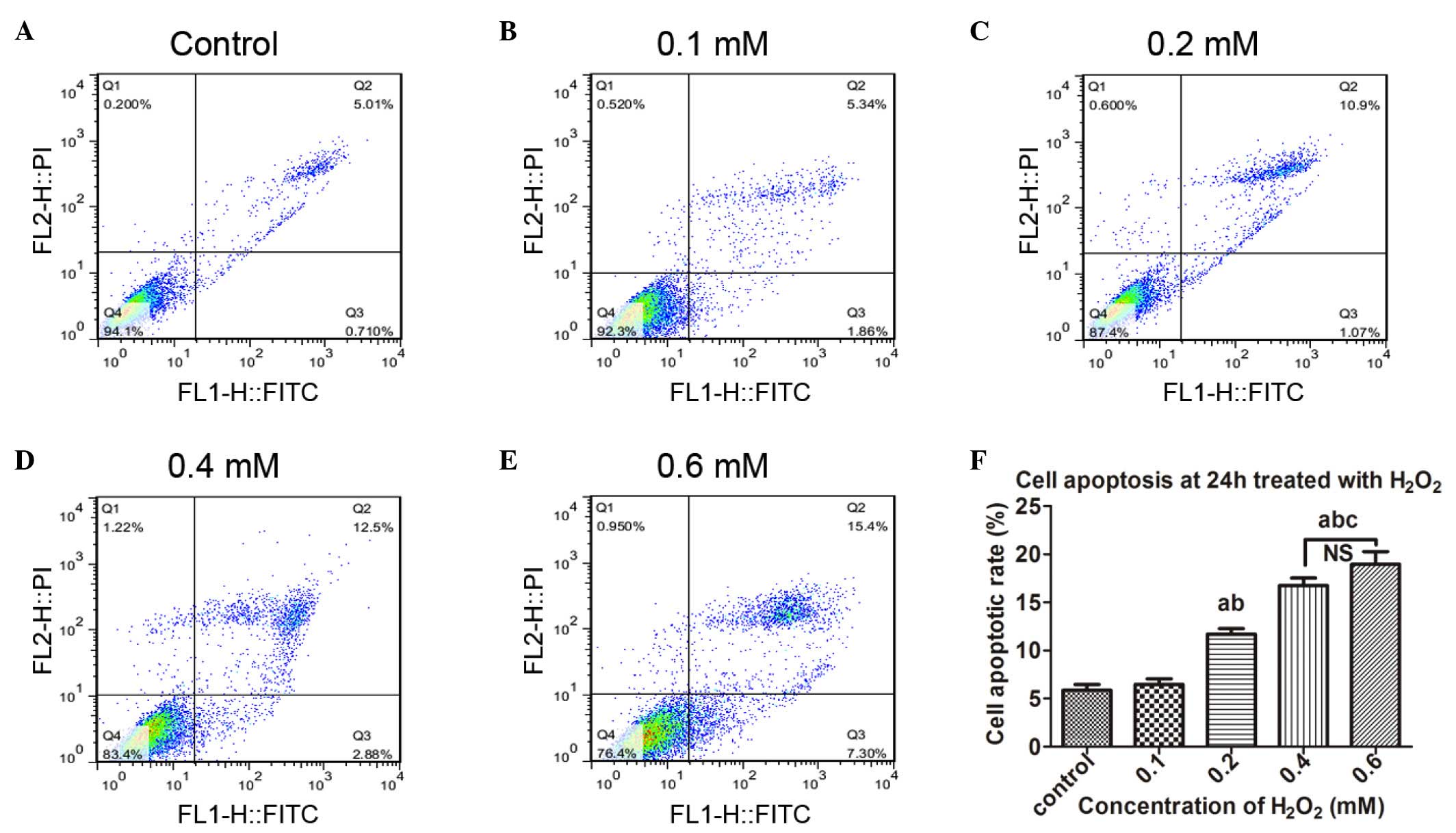 | Figure 3Cell apoptosis is induced by
exogenous H2O2. Following treatment with
different concentrations of H2O2 for 24 h,
the human uterosacral ligament fibroblasts were stained with
Annexin V/propidium iodide and then assayed using flow cytometry.
Cell apoptosis following exposure to (A) 0, (B) 0.1, (C) 0.2, (D)
0.4 and (E) 0.6 mM H2O2. (F) Statistical
analysis of the apoptotic rates. Data are presented as the median ±
standard error of the mean. One-way analysis of variance was
performed, followed by an unpaired t-test. Data are presented as
the median ± standard error of the mean (n=3). a, P<0.05, vs.
untreated control; b, P<0.05, vs. 0.1 mM; c, P<0.05, vs. 0.2
mM; NS, no significance; H2O2, hydrogen
peroxid; PI, propidium iodide; FITC, fluorescein isothocyanate. |
Intracellular ROS generation is induced
by incubation with H2O2
To confirm the increase of intracellular ROS in
hUSLFs, a DCF-DA assay was performed following treatment with
H2O2 at different concentrations ranging
between 0 and 0.6 mM for 24 h. As shown in Fig. 4, with increasing concentrations of
H2O2, the fluorescence intensity of the
oxidized DCF, which indicates the induction of intracellular ROS,
gradually increased. There were significant differences among all
pairs of groups (P<0.05), with the exception of 0.4 and 0.6 mM.
These results revealed that H2O2
significantly elevated the levels of intracellular ROS in the
hUSLFs following 24 h exposure at concentrations ranging between 0
and 0.4 mM.
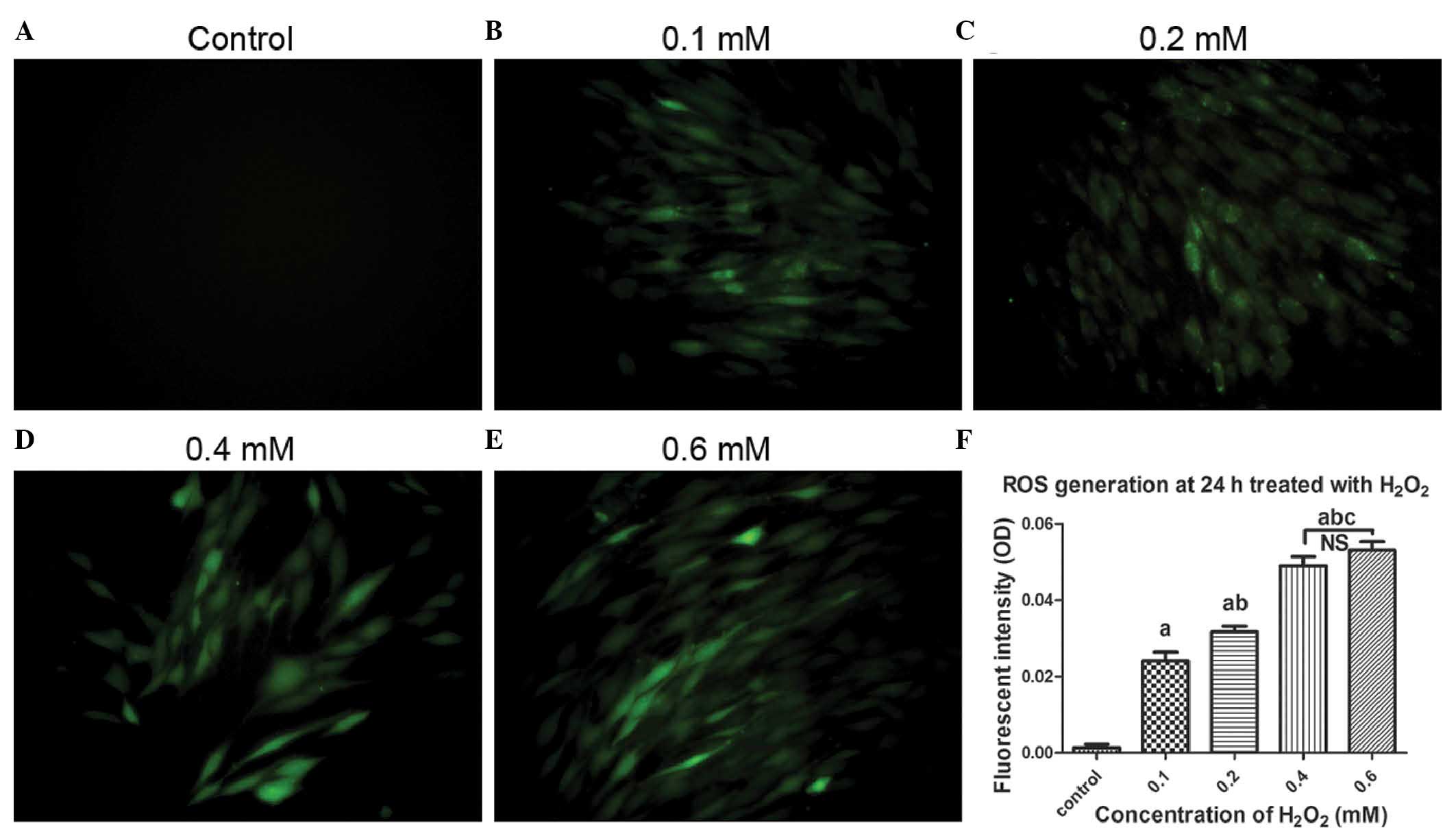 | Figure 4Microscopic images of ROS generation
induced by H2O2 treatment using DCF-DA
staining. The cells were pre-treated with exogenous
H2O2 at concentrations of (A) 0, (B) 0.1, (C)
0.2, (D) 0.4 and (E) 0.6 mM, and then incubated with DCF-DA. The
cells were observed under a fluorescent microscope (magnification,
×200). (F) Quantitative analysis based on fluorescence intensity,
obtained using Image-pro Plus 6.0 software. Data are presented as
the median ± standard error of the mean. One-way analysis of
variance was performed, followed by an unpaired t-test. Data are
presented as the median ± standard error of the mean (n=3). a,
P<0.05, vs. untreated control; b, P<0.05, vs. 0.1 mM; c,
P<0.05, vs. 0.2 mM; NS, no significance; ROS, reactive oxygen
species; OD, optical density; H2O2, hydrogen
peroxide; DCF-DA, 2′,7′-dichlorofluorescein diacetate. |
Production of 8-OHdG is a biomarker of
oxidative damage
To clarify the oxidative damage secondary to the
increase of intracellular ROS, the production of 8-OHdG was
examined using an indirect immunofluorescent assay. As shown in
Fig. 5, as the concentration of
H2O2 increased between 0 and 0.4 mM, the
fluorescence intensity of 8-OHdG, a biomarker of oxidative damage,
gradually increased, and the intergroup differences were
statistically significant (P<0.05).
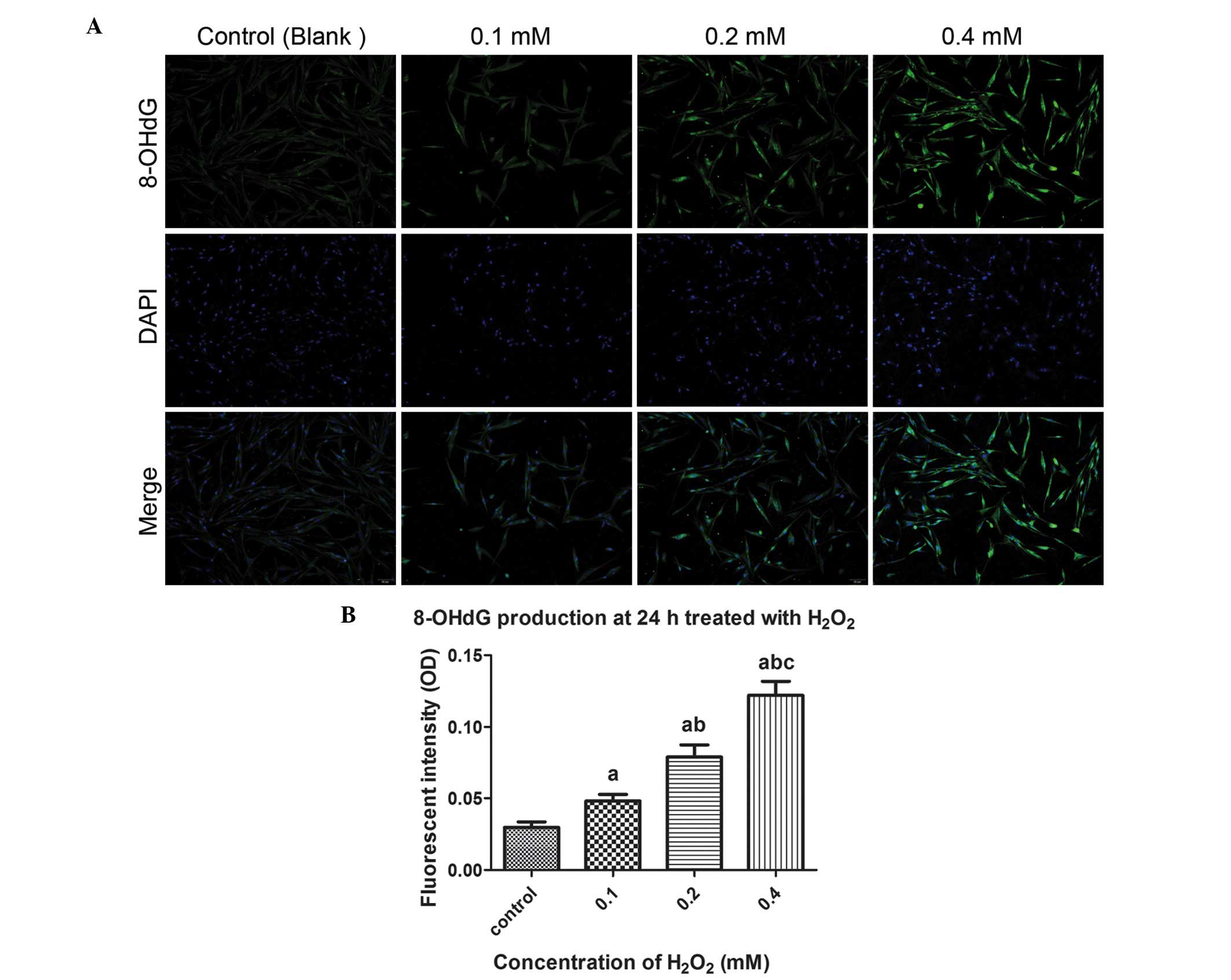 | Figure 5Microscopic images of 8-OHdG
production induced by H2O2 treatment using an
immunofluorescent assay. (A) Cells were pre-treated with exogenous
H2O2 (0, 0.1, 0.2, and 0.4 mM) for 24 h,
followed by incubation with 8-OHdG antibodies and staining with
DAPI. The cells were observed under a fluorescent microscope
(magnification, ×200). (B) Quantitative analysis based on
fluorescence intensity, obtained using image-pro plus 6.0 software.
One-way analysis of variance was performed, followed by an unpaired
t-test. Data are presented as the median ± standard error of the
mean (n=3). a, P<0.05, vs. untreated control; b, P<0.05, vs.
0.1 mM; c, P<0.05, vs. 0.2 mM; NS, no significance; 8-OHdg,
8-hydroxyguanosine; OD, optical density;
H2O2, hydrogen peroxide. |
H2O2 treatment
regulates collagen metabolism in fibroblasts
In order to clarify the effects of
H2O2 treatment on collagen metabolism, and
determine the potential mechanism, the present study examined the
expression levels of COL1A1, MMP-2, TIMP-2 and TGF-β1 by Western
blot and RT-qPCR analyses. As shown in Fig. 6A and B), as the concentration of
H2O2 increased between from 0 and 0.4 mM, the
protein synthesis of COL1A1 reduced following an initial increase,
and the intergroup differences were statistically significant
(P<0.05), with the exception of that between the 0.1 and 0.2 mM
groups. The protein level of MMP-2 was gradually and significantly
increased as the H2O2 concentration increased
(P<0.05). By contrast, there was a sharp decline in the protein
expression of TIMP-2 when the concentration of
H2O2 increased between 0.1 and 0.4 mM, and
the difference among these groups were significant (P<0.05).
Notably, TGF-β1 showed a similar change to that of COL1A1. Based on
the RT-qPCR data for COL1A1, MMP-2, TIMP-2 and TGF-β1 (Fig. 6C–F), it was confirmed that the
changes in the mRNA expression levels were consistent with those of
the proteins.
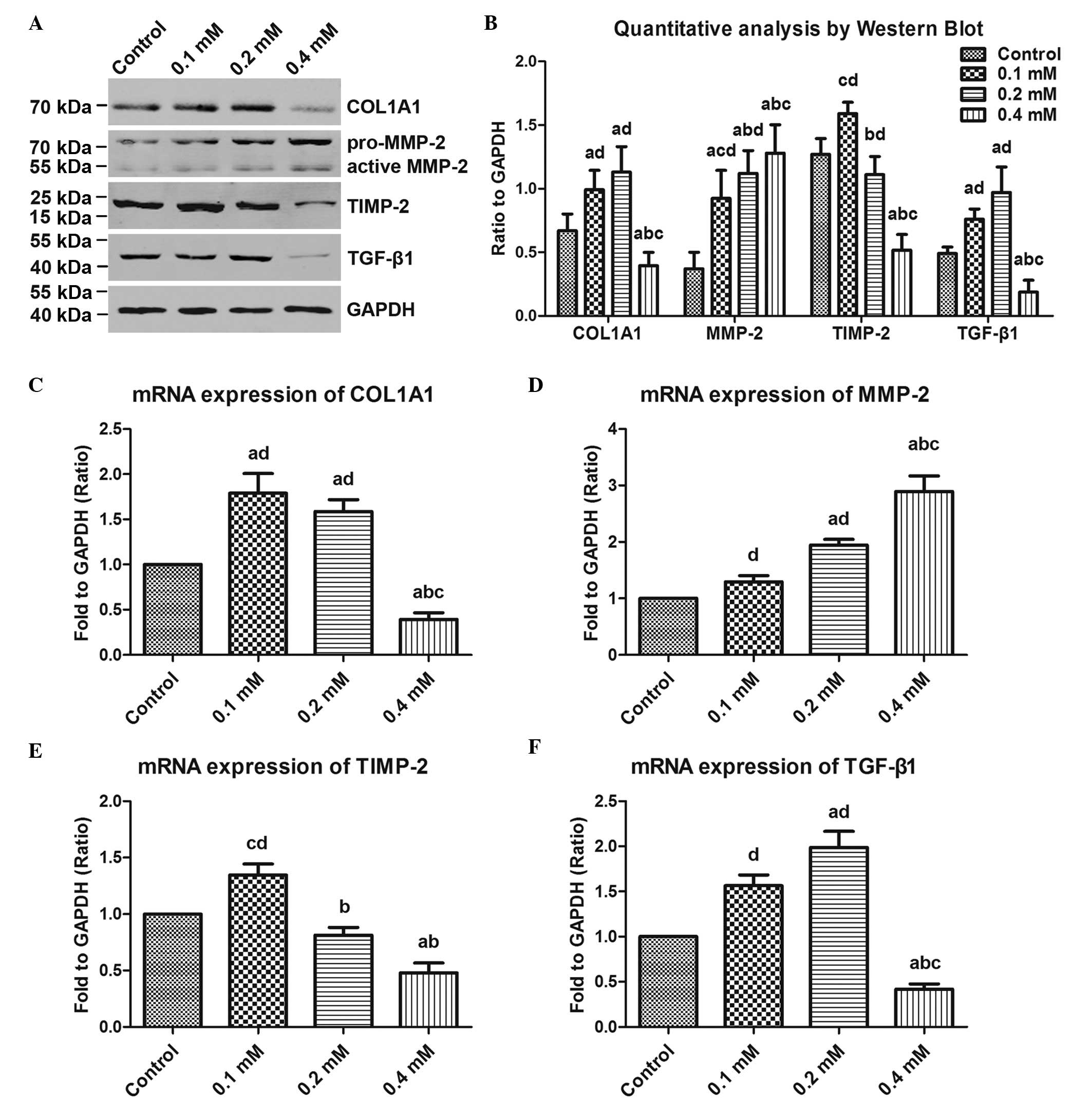 | Figure 6Effects of exogenous
H2O2 on COL1A1 metabolism in human
uterosacral ligament fibroblasts. The cells were pre-treated with
the indicated concentrations of H2O2 (0, 0.1,
0.2 and 0.4 mM) for 24 h, and then assayed by Western blot and
RT-qPCR analyses. (A) COL1A1, MMP-2, TIMP-2 and TGF-β1 were
examined using Western blot analysis at the protein level. (B)
Quantitative analysis based on the bands of the Western blot. The
mRNA expression levels of (C) COL1A1, (D) MMP-2, (E) TIMP-2 and (F)
TGF-β1 were determined by RT-qPCR. One-way analysis of variance was
performed and data are presented as the median ± standard error of
the mean (n=3). a, P<0.05, vs. untreated control; b, P<0.05,
vs. 0.1 mM; c, P<0.05, vs. 0.2 mM; d, P<0/05, vs. 0.4 mM; NS,
no significance; COL1A1, collagen, type 1, α1; MMP-2, matrix
metalloproteinase-2, TIMP-2, tissue inhibitor of
metalloproteinase-2; TGF-β1, transforming growth factor-β1;
H2O2, hydrogen peroxide. |
Discussion
Previous reports have indicated that metabolic
disorder of the ECM, characterized by reduced collagen anabolism
and hyperfunction of MMPs, is the pathological molecular basis of
POP (8–12). Aging, vaginal delivery, chronic
constipation, obesity and declined hormone status, are
well-recognized risk factors (5,6). A
number of notable commonalities have been identified regarding the
similarities in these risk factors. According to the Free Radical
Theory of Aging (22), cell aging
is a consequence of oxidative injury in a sense. To recognize the
damage to the pelvic floor caused by vaginal delivery, aside from
direct trauma at childbirth (23),
chronic nerve injuries of the levator ani muscle during pregnancy
and postpartum periods have been repeatedly reported (24,25).
In addition, obesity and constipation can cause increased
intra-abdominal pressure (IAP), which is currently viewed as an
exacerbating factor of POP (26),
as IAP may exert chronic mechanical strain on the pelvic support
structures. In vitro, it has been demonstrated that cyclic
mechanical stretches cause OS in several types of cell (27–29).
Considering these previous findings, the present study hypothesized
that OS may mediate the pathogenesis of POP.
According to established experiment design, all
participants in the present study were well matched in age, parity,
body mass index and postmenopausal duration (Table I), therefore, investigation was
performed to confirm whether more serious oxidative injury was
present in the women with POP, compared with the normal control
group. 8-OHdG is a modified base, which occurs in DNA due to attack
by hydroxyl radicals that are formed as by-products and
intermediates of aerobic metabolism and during OS (30). 8-OHdG is well correlated with OS
and damage to DNA, which leads to degenerative disease states. As a
result, 8-OHdG has become increasingly used as a sensitive, stable
and integral marker of oxidative damage in cellular DNA. 4-HNE, as
a stable product of lipid peroxidation, has been implicated in the
etiology of pathological changes under OS, as a key mediator of
OS-induced cell death. Through immunohistochemical examination of
USL sections, the present study found significantly high levels of
8-OHdG and 4-HNE immunoreactivity in the POP group, compared with
those in the control group (Fig.
1). Although the number of cases was insufficient to determine
whether there was a linear correlation between the severity of
oxidative injury and POP staging, the data obtained confirmed the
presence of OS in prolapsed USL, which was partially in accordance
with previous conclusions (31,32).
In order to determine the exact role of OS in the
pathogenesis of POP and the associated mechanisms, it is necessary
to establish an OS cell model in fibroblasts derived from USL
tissue of non-POP women (hUSLFs). As described above, a series of
dose-effect investigations were performed in the present study to
determine the appropriate concentration and incubation duration
(Figs. 2 and 3). In addition, the rationale of the cell
model was verified from two aspects, the generation of
intracellular ROS (Fig. 4) and the
production of 8-OHdG (Fig. 5). To
the best of our knowledge, the present study is the first to
successfully establish an OS model in hUSLFs via
H2O2 incubation, and this cell model may
facilitate further investigations involving OS in POP. Due to time
and experimental technology constraints, the present study was
limited, resulting in examination of the cell model with
immunofluorescence only. The addition of Western blot or PCR data
is required to provide more systemic and persuasive
conclusions.
The primary aim of the present study was to
elucidate the effects of OS on collagen metabolism in hUSLFs. The
results of the present study revealed that exogenous
H2O2 had two-way regulatory effects on
collagen metabolism (Fig. 6).
Following incubation for 24 h, lower concentrations of
H2O2 stimulated the anabolism of COL1A1,
whereas higher concentration promoted catabolism. The specific
effect was dependent on the severity of OS. Therefore, it was
concluded that OS contributed to collagen metabolic disorder in the
human pelvic fibroblasts. To examine the associated mechanisms,
MMP-2, TIMP-2 and TGF-β1 were examined. MMP-2, as a key proteinase
responsible for degradation of collagen, has been well demonstrated
in previous reports, and TIMP-2 acts as a metallopeptidase
inhibitor. According to reports on fibrotic diseases, TGF-β1, a 25
kDa polypeptide tissue growth factor, has been identified as an
important cytokine, which promotes fibrosis by inducing fibroblast
differentiation, stimulating synthesis of ECM and inhibiting its
degradation. The TGF-β1/small mothers against decapentaplegic 3
signaling pathway is currently viewed as an important regulator
that is widely involved in fibrosis and degenerative fibrotic
diseases (33–36). However, TGF-β1 is rarely discussed
in previous reports discussing the pathophysiology of POP. Moalli
et al (37) reported that
exogenous TGF-β1 stimulates the expression of MMP-2 in human pelvic
fibroblasts (32). In the present
study, the expression levels of MMP2, TIMP2 and TGFβ1 corresponded
to the levels of OS. Although the present study did not verify
whether the changes in TGF-β1 were primary or secondary to COL1A1,
MMP-2 or TIMP-2, TGF-β1 may be involved in collagen metabolic
disorder by regulating MMP-2 and/or TIMP-2. Further investigation
is warranted, and may further assist in further elucidating the
pathophysiology of POP.
Elevated oxidative injury is one of the
characteristics of POP, and OS contributes to collagen metabolic
disorder in a severity-dependent manner in hUSLFs. The present
study hypothesized that OS may be involved in the pathophysiology
of POP, either by inhibiting the anabolism of collagen or,
alternatively, by promoting catabolism indirectly through the
regulation of TGF-β1 and proteolytic enzymes, including MMPs.
Further investigation is required to improve current understanding
of the exact mechanism and to elucidate the pathophysiology of POP,
which may be beneficial in preventing or disrupting the progression
of POP.
Acknowledgments
The authors would like to thank Xu Xue-Xian, Li
Yan-Bo, Cheng Yan-Xiang, Luo Ruo-Yu (Department of Gynecology and
Obstetrics, Renmin Hospital of Wuhan University) for specimen
biopsy assistance. This study was funded by the National Natural
Science Foundation of China (grant nos. H0407-81270684 and
H0407-81471442).
References
|
1
|
Barber MD and Maher C: Epidemiology and
outcome assessment of pelvic organ prolapse. Int Urogynecol J.
24:1783–1790. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Olsen AL, Smith VJ, Bergstrom JO, Colling
JC and Clark AL: Epidemiology of surgically managed pelvic organ
prolapse and urinary incontinence. Obstet Gynecol. 89:501–506.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Subak LL, Waetjen LE, van den Eeden S,
Thom DH, Vittinghoff E and Brown JS: Cost of pelvic organ prolapse
surgery in the United States. Obstet Gynecol. 98:646–651.
2001.PubMed/NCBI
|
|
4
|
Merrill RM: Hysterectomy surveillance in
the United States, 1997 through 2005. Med Sci Monit. 14:CR24–CR31.
2008.
|
|
5
|
Weber AM, Buchsbaum GM, Chen B, Clark AL,
Damaser MS, Daneshgari F, Davis G, DeLancey J, Kenton K, Weidner AC
and Word RA: Basic science and translational research in female
pelvic floor disorders: Proceedings of an NIH-sponsored meeting.
Neurourol Urodyn. 23:288–301. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jelovsek JE, Maher C and Barber MD: Pelvic
organ prolapse. The Lancet. 369:1027–1038. 2007. View Article : Google Scholar
|
|
7
|
Yiou R, Authier FJ, Gherardi R and Abbou
C: Evidence of mitochondrial damage in the levator ani muscle of
women with pelvic organ prolapse. Eur Urol. 55:1241–1243. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jackson SR, Avery NC, Tarlton JF, Eckford
SD, Abrams P and Bailey AJ: Changes in metabolism of collagen in
genitourinary prolapse. Lancet. 347:1658–1661. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Klutke J, Ji Q, Campeau J, Starcher B,
Felix JC, Stanczyk FZ and Klutke C: Decreased endopelvic fascia
elastin content in uterine prolapse. Acta Obstet Gynecol Scand.
87:111–115. 2008. View Article : Google Scholar
|
|
10
|
Chen B and Yeh J: Alterations in
connective tissue metabolism in stress incontinence and prolapse. J
Urol. 186:1768–1772. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gabriel B, Watermann D, Hancke K, Gitsch
G, Werner M, Tempfer C and zur Hausen A: Increased expression of
matrix metalloproteinase 2 in uterosacral ligaments is associated
with pelvic organ prolapse. Int Urogynecol J Pelvic Floor Dysfunct.
17:478–482. 2006. View Article : Google Scholar
|
|
12
|
Strinic T, Vulic M, Tomic S, Capkun V,
Stipic I and Alujevic I: Matrix metalloproteinases-1, -2 expression
in uterosacral ligaments from women with pelvic organ prolapse.
Maturitas. 64:132–135. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Takacs P, Nassiri M, Gualtieri M,
Candiotti K and Medina CA: Uterosacral ligament smooth muscle cell
apoptosis is increased in women with uterine prolapse. Reprod Sci.
16:447–452. 2009. View Article : Google Scholar
|
|
14
|
Sampson N, Berger P and Zenzmaier C: Redox
signaling as a therapeutic target to inhibit myofibroblast
activation in degenerative fibrotic disease. Biomed Red Int.
2014:1–14. 2014. View Article : Google Scholar
|
|
15
|
Fisher GJ, Wang ZQ, Datta SC, Varani J,
Kang S and Voorhees JJ: Pathophysiology of premature skin aging
induced by ultraviolet light. N Engl J Med. 337:1419–1428. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Siwik DA, Pagano PJ and Colucci WS:
Oxidative stress regulates collagen synthesis and matrix
metalloproteinase activity in cardiac fibroblasts. Am J Physiol
Cell Physiol. 280:C53–C60. 2001.
|
|
17
|
Akhtar K, Broekelmann TJ, Miao M, Keeley
FW, Starcher BC, Pierce RA, Mecham RP and Adair-Kirk TL: Oxidative
and nitrosative modifications of tropoelastin prevent elastic fiber
assembly in vitro. J Biol Chem. 285:37396–37404. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
World Medical Association: World Medical
Association Declaration of Helsinki: Ethical principles for medical
research involving human subjects. JAMA. 318:2191–2194. 2013.
|
|
19
|
Bump RC, Mattiasson A, Bø K, Brubaker LP,
DeLancey JO, Klarskov P, Shull BL and Smith AR: The standardization
of terminology of female pelvic organ prolapse and pelvic floor
dysfunction. Am J Obstet Gynecol. 175:10–17. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hong S, Li H, Wu D, Li B, Liu C, Guo W,
Min J, Hu M, Zhao Y and Yang Q: Oxidative damage to human
parametrial ligament fibroblasts induced by mechanical stress. Mol
Med Rep. 12:5342–5348. 2015.PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCt method. Methods. 25:402–408. 2001. View Article : Google Scholar
|
|
22
|
Harman D: Aging: A theory based on free
radical and radiation chemistry. J Gerontol. 11:298–300. 1956.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
DeLancey JO, Kearney R, Chou Q, Speights S
and Binno S: The appearance of levator ani muscle abnormalities in
magnetic resonance images after vaginal delivery. Obstet Gynecol.
101:46–53. 2003.PubMed/NCBI
|
|
24
|
Weidner AC, Jamison MG, Branham V, South
MM, Borawski KM and Romero AA: Neuropathic injury to the levator
ani occurs in 1 in 4 primiparous women. Am J Obstet Gynecol.
195:1851–1856. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lubowski DZ, Swash M, Nicholls RJ and
Henry MM: Increase in pudendal nerve terminal motor latency with
defaecation straining. Br J Surg. 75:1095–1097. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Spence-Jones C, Kamm MA, Henry MM and
Hudson CN: Bowel dysfunction: A pathogenic factor in uterovaginal
prolapse and urinary stress incontinence. Br J Obstet Gynaecol.
101:147–152. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pimentel DR, Amin JK, Xiao L, Miller T,
Viereck J, Oliver-Krasinski J, Baliga R, Wang J, Siwik DA, Singh K,
et al: Reactive oxygen species mediate amplitude-dependent
hypertrophic and apoptotic responses to mechanical stretch in
cardiac myocytes. Circ Res. 89:453–460. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Davidovich N, DiPaolo BC, Lawrence GG,
Chhour P, Yehya N and Margulies SS: Cyclic stretch-induced
oxidative stress increases pulmonary alveolar epithelial
permeability. Am J Respir Cell Mol Biol. 49:156–164. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rodríguez AI, Csányi G, Ranayhossaini DJ,
Feck DM, Blose KJ, Assatourian L, Vorp DA and Pagano PJ: MEF2B-Nox1
signaling is critical for stretch-induced phenotypic modulation of
vascular smooth muscle cells. Arterioscler Thromb Vasc Biol.
35:430–438. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kroese LJ and Scheffer PG:
8-hydroxy-2′-deoxyguanosine and cardiovascular disease: A
systematic review. Curr Atheroscler Rep. 16:4522014. View Article : Google Scholar
|
|
31
|
Kim EJ, Chung N, Park SH, Lee KH, Kim SW,
Kim JY, Bai SW and Jeon MJ: Involvement of oxidative stress and
mitochondrial apoptosis in the pathogenesis of pelvic organ
prolapse. J Urol. 189:588–594. 2013. View Article : Google Scholar
|
|
32
|
Ewies A and Elshafie M: High isoprostane
level in cardinal ligament-derived fibroblasts and urine sample of
women with uterine prolapse. BJOG. 116:126–127; author reply
127–128. 2009. View Article : Google Scholar
|
|
33
|
Hinz B: The extracellular matrix and
transforming growth factor-beta1: Tale of a strained relationship.
Matrix Biol. 47:54–65. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Rodríguez-Vita J, Sánchez-Galán E,
Santamaría B, Sánchez-López E, Rodrigues-Díez R, Blanco-Colio LM,
Egido J, Ortiz A and Ruiz-Ortega M: Essential role of TGF-beta/Smad
pathway on statin dependent vascular smooth muscle cell regulation.
PLoS One. 3:e39592008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gordon KJ and Blobe GC: Role of
transforming growth factor-beta superfamily signaling pathways in
human disease. Biochim Biophys Acta. 1782:197–228. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yang J, Zheng J, Wu L, Shi M, Zhang H,
Wang X, Xia N, Wang D, Liu X, Yao L, et al: NDRG2 ameliorates
hepatic fibrosis by inhibiting the TGF-β1/Smad pathway and altering
the MMP2/TIMP2 ratio in rats. PLoS One. 6:e277102011. View Article : Google Scholar
|
|
37
|
Moalli PA, Klingensmith WL, Meyn LA and
Zyczynski HM: Regulation of matrix metalloproteinase expression by
estrogen in fibroblasts that are derived from the pelvic floor. Am
J Obstet Gynecol. 187:72–79. 2002. View Article : Google Scholar : PubMed/NCBI
|