Introduction
Human dental mesenchymal stem cells (hDMSCs) can be
isolated from various types of dental tissues, such as periodontal
ligament, dental pulp, alveolar bone marrow and periapical
follicle, and have the ability to differentiate into different cell
types, including odontoblasts, osteoblasts, adipocytes and
chondrocytes, with the purpose of cell based tissue engineering
(1,2). Previous studies have used hDMSCs for
dental tissue regeneration (3).
Dental pulp cells (DPCs) can be obtained from extracted third molar
teeth, which are clinically regarded as ‘waste tissue’, and can
differentiate into odontoblasts that serve as the precursor cells
for dentin formation. However, the precise mechanism of DPCs
odontoblast differentiation is still unclear.
In recent years, epigenetic regulation had been
proven to serve an important role in the differentiation of many
cells. Epigenetic modification of chromatin does not result in
changes to the DNA sequence, but instead modifies the structure and
the accessibility of chromatin, leading to heritable alterations of
gene expression during cell division (4). Epigenetic modification of gene
expression explicitly operates through chromatin remodelling, such
as nucleosome structural modification. Acetylation and
deacetylation controlled by histone acetyltransferases (HATs) and
histone deacetylases (HDACs) are the common methods of
post-translational histone modification (5). HDAC inhibitors (HDACi) have been
widely used in oncotherapy due to the potent effects of inducing
cell cycle arrest, apoptosis and differentiation of tumor cell
lines (6). Furthermore, it had
been demonstrated that HDACi can promote osteoblastic
differentiation and bone formation by increasing the expression of
osteogenic-associated proteins, including alkaline phosphatase
(ALP), osteocalcin (OCN), osterix and runt-related transcription
factor 2 (Runx2), and accelerating mineralization via HDAC
inhibition (7,8). Class I and class II HDACs have been
demonstrated to be highly expressed in human periodontal ligaments,
while HDAC3 expression is decreased gradually during osteogenic
differentiation. Meanwhile, inhibition of HDAC3 by trichostatin A
(TSA) might accelerate osteoblast differentiation in periodontal
ligament cells through hyperacetylation of Histone H3 (9). Osteoblast differentiation could also
be stimulated by HDAC1 inhibition via the downregulation of osterix
and OCN (10). In contrast,
long-term use of VPA, an HDAC inhibitor thatis also used as an
antiepileptic drug, reduces bone mineral density as well as content
in vivo, which is possibly due to the systemic use of VPA
and a relatively high dose (11).
Therefore, the appropriate usage and dosage of HDACi for activating
cell differentiation needs to be determined before use.
Previous studies regarding HDAC expression in human
dental tissue, especially dental pulp, indicated that HDAC1, 2, 3,
4 and 5 were highly expressed in odontoblasts, and class IHDACi has
been demonstrated to promote differentiation and increase
mineralization at relatively low concentrations (12,13).
Kwon et al (14)
demonstrated that HDAC inhibition enhanced odontoblast
differentiation and increased dentin sialophosphoprotein (DSPP)
expression in odontoblast-like cells partially by increasing the
expression of nuclear factor 1 C-type. Class IIHDACs such as HDAC4
and HDAC5 are closely associated with osteoblast differentiation.
It was demonstrated that acetylation of osteoblast-specific
transcription factor osterix (Osx) was essential for osteoblast
differentiation of C2C12 cells, while deacetylation of Osx mediated
by HDAC4 may serve the opposite role (15). Through inhibiting HDAC5 during
osteogenic differentiation of vascular smooth muscle cells,
microRNA-2861 could upregulate Runx2 protein expression levels
(16). However, the effects of
HDAC4, HDAC5and their specific inhibitor on odontoblast
differentiation of DPCs, and the precise molecular and epigenetic
mechanisms behind this process, remain unclear. LMK-235 is a human
specific HDAC4 and HDAC5 inhibitor (17,18),
and the present study aimed to investigate how LMK-235 affects the
proliferation and differentiation of DPCs in vitro, and
identify its possible molecular mechanism. This study may provide
novel ideas for odontoblast differentiation of dental pulp derived
cells and dentin regeneration.
Materials and methods
Primary cell culture
Human third molars were collected from healthy young
men (18–25 years of age) at the Department of Oral and
Maxillofacial Surgery, Nanfang Hospital, Southern Medical
University (Guangzhou, China). This study was approved by the
Institutional Review Board at the Department of Stomatology,
Nanfang Hospital, Southern Medical University. Written informed
consent was obtained from the patients prior to the study. The pulp
tissue was gently separated from the extracted third molars and
subsequently digested in a solution of 3 mg/ml collagenase type I
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) at 37°C for 30 min.
The pulp tissue was cultured in 10% fetal calf serum with 100 U/ml
penicillin and 100 mg/ml streptomycin (all from Hyclone, Logan, UT,
USA) and cultured at 37°C in 5% CO2. The medium was
changed 24 h later and then changed every 3 days. The primary cells
were used at passage 3–5 in this study.
Cell treatment
To evaluate the appropriate concentration of LMK-235
(Selleck Chemicals, Houston, TX, USA) to induce DPC odontoblast
differentiation without cell proliferation reducing, cells were
treated with different concentrations (0, 50, 100, 250, 500 and
1,000 nM) of LMK-235 for 3 days, and the 0 nM group was set as the
control group. The gene expression levels of odontoblast markers,
such as DSPP, ALP and Runx2, were detected by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR).
After the concentration was confirmed, the effect of
LMK-235 was tested onodontoblast differentiationin normal medium
[Dulbecco's modified Eagle's medium supplemented with 10% fetal
bovine serum (Hyclone)] and mineralizing medium. Cells
(1×106) were seeded in a 60-mm culture dish, and the
cells were divided into the following four groups: Control
(incubated in normal growth medium and Dulbecco's modified Eagle's
medium supplemented with 10% fetal bovine serum), LMK-235 (in
normal growth medium supplemented with 100 nM LMK-235), mineralized
inductive (MI; in mineralizing inductive medium), and LMK-235
+mineralized inductive (MI+LMK-235; in mineralizing inductive
mediumsupplemented with 100 nM LMK-235). The mineralizing inductive
medium contained 10% fetal bovine serum, 50 mg/ml ascorbic acid, 10
mM β-glycerophosphate and 100 nM dexamethasone (all from
Sigma-Aldrich; Merck KGaA). The medium was changed every 3 days.
ALP activity was tested 1week after treatment, and Alizarin Red S
staining (room temperature, 30 min) was completed after a 21-day
treatment. Furthermore, the mRNA and protein expression levels of
DSPP, ALP, Runx2 and OCN were detected by RT-qPCR and western
blotting after 7, 14 and 21 days of culture. To explore the
potential mechanism behind odontoblast differentiation activated by
LMK-235, the mRNA expression levels of vascular endothelial growth
factor (VEGF), RAC-gamma serine/threonine-protein kinase (AKT3) and
mechanistic target of rapamycin (mTOR) were detected soon
afterwards.
Cell proliferation assay
To evaluate the effects of LMK-235 on cell viability
at different concentrations, cell proliferation and viability were
measured using MTT assay. Briefly, DPCs (4.0×103
cells/well) were seeded into 96-well plates and incubated for 24 h.
Various concentrations of LMK-235 were added (0, 50, 100, 250, 500
and 1,000 nM) in a culture medium volume of 100 ml in each well.
MTT (~15 µl; 5%; Sigma-Aldrich; Merck KGaA) was added at the1, 3, 5
and 7-day time points. The DPCs were further incubated for 4 h in
5% CO2 at 37°C, and then 100 ml dimethyl sulfoxide
(Sigma-Aldrich; Merck KGaA) was added to dissolve the formazan
product. Each condition was prepared 5 times, and the optical
densities were read using an enzyme-linked immunosorbent assay
reader at 562 nm.
RNA preparation and RT-qPCR
Total RNA extracted from DPCs was prepared using
RNAiso Plus (Takara Biotechnology Co., Ltd., Dalian, China)
according to the manufacturer's protocol. cDNA was synthesized from
0.5 mg total RNA using the PrimeScript RT Reagent kit (Takara
Biotechnology Co., Ltd.). qPCR was completed using SYBR Green PCR
Master Mix (Takara Biotechnology Co., Ltd.) on an ABI Prism 7500
sequence detection system. The reaction conditions were according
to the manufacturer's protocol as follows: 40 cycles of
denaturation at 95°C for 15 sec and amplification at 60°C for 60
sec. All reactions of each sample were tested in triplicate and
normalized to β-actin Mrna (19).
All of the specific primers are listed in Table I.
 | Table I.Primers used for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primers used for reverse
transcription-quantitative polymerase chain reaction.
| Gene | Sequence |
|---|
| β-actin | F:
5′-GAGCTACGAGCTGCCTGACG −3′ |
|
| R:
5′-CCTAGAAGCATTTGCGGTGG −3′ |
| DSPP | F:
5′-TGGCGATGCAGGTCACAAT −3′ |
|
| R:
5′-CCATTCCCACTAGGACTCCCA −3′ |
| Runx2 | F:
5′-TCAACGATCTGAGATTTGTGGG −3′ |
|
| R:
5′-GGGGAGGATTTGTGAAGACGG −3′ |
| ALP | F:
5′-CCAAAGGCTTCTTCTTGCTG-3′ |
|
| R:
5′-CCACCAAATGTGAAGACGTG-3′ |
| OCN | F:
5′-GGCGCTACCTGTATCAATGG −3′ |
|
| R:
5′-GTGGTCAGCCAACTCGTCA −3′ |
| VEGF | F: 5′-
CTACCTCCACCATGCCAAGT-3′ |
|
| R: 5′-
CACACAGGATGGCTTGAAGA-3′ |
| AKT3 |
|
| mTOR | F:
5′-AATGGACAGAAGCTATCCAGGC −3′ |
|
| R:
5′-TGATGGGTTGTAGAGGCATCC −3′ |
|
| F:
5′-TCCGAGAGATGAGTCAAGAGG −3′ |
|
| R:
5′-CACCTTCCACTCCTATGAGGC −3′ |
Western blot analysis
DPCs were solubilized in lysis buffer (Beyotime
Institute of Biotechnology, Haimen, China) for 20 min, followed by
centrifugation at 4°C at 14,000 × g for 20 min. The protein
concentrations of the cell lysates were detected using a Protein
Assay kit (Beyotime Institute of Biotechnology). Bromophenol blue
(10%) was added to 20 µg protein in each group, the mixture was
boiled for 5 min and 10% SDS-PAGE was used to separate the total
protein. After electrophoresis, proteins in the gels were
transferred to polyvinylidene fluoride membranes (EMD Millipore,
Billerica, MA, USA). The membranes were blocked afterwards in 5%
low-fat milk solubilized in Tris-buffered saline with 0.1 Tween-20
(TBST) at room temperature for 1 h, and rinsedwithTBST three times
(10 min each). The membranes then were incubated with primary
rabbit against human monoclonal antibodies: GAPDH (1:10,000; cat
no. ab128915) and Runx2 (1:1,000; cat no. ab192256) (both from
Abcam, Cambridge, MA, USA), and mouse anti-human DSPP (1:1,000; cat
no. sc-73632; Santa Cruz Biotechnology, Inc., Dallas, TX, USA),
overnight at 4°C. After three washes with TBST (10 min each), the
membranes were incubated again with goat anti-mouse IgG (1:10,000;
cat no. 926–32210) or goat anti-rabbit IgG (1:10,000; cat no.
926-68021) (both from LI-COR Biosciences, Lincoln, NE, USA)
secondary antibodies for 1 h at room temperature. Membranes were
then washed three times with TBST, and then visualized using an
Odyssey infrared imaging system (LI-COR Biosciences). Integrated
optical density was detected using ImageJ version 1.47 (National
Institutes of Health, Bethesda, MD, USA).
ALP activity test
For the ALP activity test, cells cultured for 1 week
were fixed with 10% formalin, incubated with 0.1% Triton X-100 for
5 min at room temperature, and then the ALP activity was determined
using the Leukocyte Alkaline Phosphatase kit (Sigma-Aldrich; Merck
KGaA) according to the manufacturer's protocol. The absorbance was
detected using a microplate reader at a wavelength of 520 nm. A
Bicinchoninic Acid protein assay kit (Beyotime Institute of
Biotechnology) was used to quantify the protein content. The ALP
activities in the DPCs of the 4 groups were normalized against the
total protein content.
Alizarin Red S staining
Cells were cultured in 6-well plates for 21 days,
washed using phosphate-buffered saline (PBS) and then fixed with 4%
formalin for 10 min. The cells were stained with 2% Alizarin Red S
(Sigma-Aldrich; Merck KGaA) dissolved in distilled water, pH 4.4,
at room temperature for 30 min. The cells were gently washed with
distilled water three times to remove the unbound Alizarin Red S.
The stained cells were imaged using an inverted microscope.
Statistical analysis
Statistical analysis were performed using SPSS 19.0
software (IBM Corp., Armonk, NY, USA). Data are expressed as the
mean ± standard deviation. Differences were analysed using a
one-way analysis of variance followed by a Bonferroni test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Effect of LMK-235 on cell
proliferation
HDACi might induce growth arrest and apoptosis in
tumor cells and non-tumor cells (20,21).
The results of the present study indicated that cell growth was
reduced in the 250 and 500 nM groups compared with the 0 nM group
at days 3 and 5. In addition, the proliferation of the 1,000 nM
group was reduced at days 1, 5 and 7 compared with the 0 nM group.
However, low concentrations of LMK-235 (50 and 100 nM) barely
affected the proliferation of DPCs (Fig. 1A).
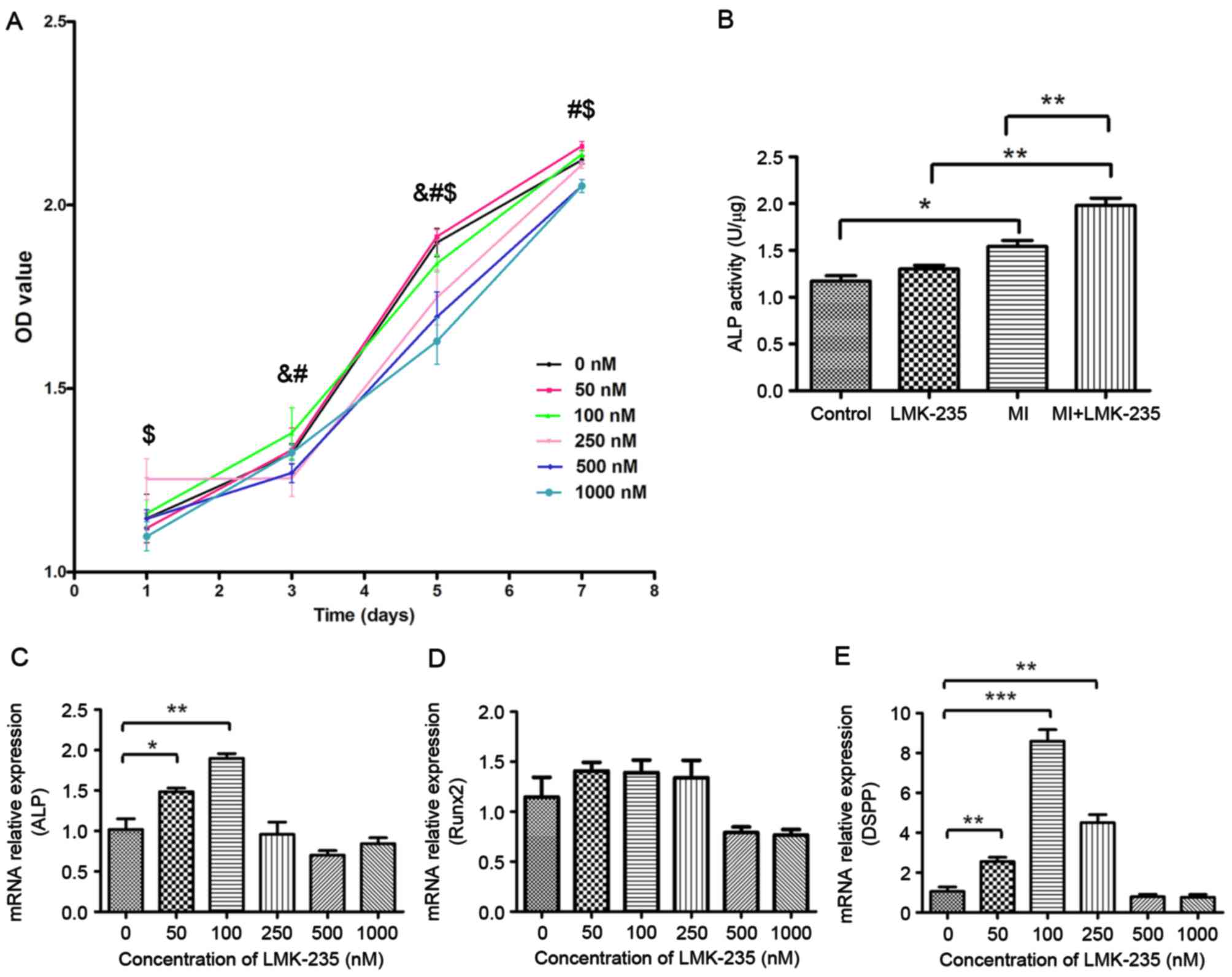 | Figure 1.Effect of LMK-235 on DPCs. (A) Cell
viability, OD value for 0 nM group was compared with the remaining
groups. &P<0.05 vs. 250 nM; #P<0.05
vs. 500 nM; $P<0.05 vs. 1,000 nM. (B) ALP activity,
and (C) ALP, (D) Runx2 and (E) DSPP mRNA expression levels in DPCs
following treatment with LMK-235. *P<0.05, **P<0.01,
***P<0.001. Data are presented as the mean ± standard deviation.
OD, optical density; DPCs, dental pulp cells; ALP, alkaline
phosphatase; DSPP, dentin sialophosphoprotein; Runx2, runt-related
transcription factor 2; MI, mineralized inductive. |
Effect of LMK-235 on ALP activity in
DPCs
The LMK-235 treated group demonstrated no
significant differences in ALP activity compared with the control
group. However, ALP activity of the MI+LMK-235 group was
significantly increased when compared with the MI group (Fig. 1B).
Determination of LMK-235 concentration
to promote odontoblast differentiation
To find the proper concentration of LMK-235 to
promote DPCs odontoblast differentiation, the expression levels of
odontoblast-specific genes were detected in DPCs incubated with
different concentrations of LMK-235 for 3 days by RT-qPCR. The
expression of ALP mRNA was significantly upregulated at low
concentrations of LMK-235 (50 and 100 nM) when compared with the
control group (Fig. 1C). In
addition, the expression of Runx2 was higher in the 50,100 and 250
nM groups than in the control group, although these differences
were not statistically significant (Fig. 1D). Furthermore, the expression of
DSPP mRNA was also increased in the DPCs of the 50,100 and 250 nM
groups compared with the control group (Fig. 1E). The expression level of the 100
nM group was 8.22-times that of the control group (P<0.001).
Conversely, the expression levels of all three of these factors
were downregulated in the 500 and 1,000 nM groups.
Combined with the effect of LMK-235 on cell
proliferation, 100 nM was chosen as the experimental concentration
at which cell proliferation was not affected, and the expressions
of odontoblast genes were upregulated in DPCs.
LMK-235 combined with mineralizing
medium increases the mRNA expression of odontoblast-specific
factors in DPCs
As presented in Fig.
2, the effect of LMK-235 treatment on DPCs during mineralized
induction was evaluated. DSPP and ALP mRNA expression levels were
increased in the MI+LMK-235 group and the MI group compared with
the LMK-235 treated group and the control group at days 7 and 14
respectively. Furthermore, them RNAexpression levels in the
MI+LMK-235 group were significantly higher than that in the MI
group at those times. At day 21, there was no significant
differences in DSPP and ALP mRNA expression between the MI+LMK-235
and the MI groups (Fig. 2A and B,
respectively).
 | Figure 2.LMK-235 combined with mineralizing
medium increases the mRNA expression of odontoblast-specific
factors in DPCs. mRNA expression levels of (A) DSPP, (B) ALP, (C)
Runx2 and (D) OCN in DPCs following treatment with LMK-235 for 7,
14 and 21 days. Data are presented as the mean ± standard
deviation. *P<0.05, **P<0.01, ***P<0.001. DPCs, dental
pulp cells; ALP, alkaline phosphatase; DSPP, dentin
sialophosphoprotein; Runx2, runt-related transcription factor 2;
MI, mineralized inductive; OCN, osteocalcin. |
Runx2 mRNA expression was significantly increased in
the MI+LMK-235 and MI groups when compared with the LMK-235 treated
group and the control group at days 7 and 14. The expression levels
in the MI+LMK-235 group and MI group were 1.45- and 1.47-times
those in the LMK-235 group and the control group, respectively,
with a significant difference at day 21. In addition, the Runx2
mRNA expression level in the MI+LMK-235 group was notably increased
relative to that of the MI group at day 21 (Fig. 2C).
OCN mRNA expression was not altered significantly
among the 4 groups at days 7 and 14. However, OCN mRNA expression
in the MI+LMK-235 group and the MI group was 1.51- and
1.45-timesthat of the LMK-235 treated group and the control group,
respectively, at day 21, whereas there was no significant
difference between the expression levels in the MI+LMK-235 and MI
groups (Fig. 2D).
LMK-235 combined with mineralizing
medium increases the protein expression of odontoblast specific
factors in DPCs
As presented in Fig.
3, DSPP protein expression was markedly higher in the
MI+LMK-235 group compared with the MI group at days 7 and 14. At
day 21, DSPP protein expression was slightly higher in the
MI+LMK-235 group compared with the MI group without significant
difference. Runx2 protein expression was upregulated by LMK-235
treatment in normal culture medium and mineralizing medium, and the
expression level in the MI+LMK-235 group was 1.22-, 1.29- and
1.17-times that in the MI group at days 7,14 and 21, respectively
(Fig. 3).
 | Figure 3.LMK-235 combined with mineralizing
medium increases the protein expression of odontoblast specific
factors in DPCs. Representative western blot images of protein
expression levels of DSPP and Runx2 at (A) 7, (B) 14 and (C) 21
days. C, control group; L, LMK-235 group; M, MI group; ML,
MI+LMK-235 group. Quantification of (D) DSPP and (E) Runx2 protein
expression levels. Data are presented as the mean ± standard
deviation. *P<0.05, **P<0.01, ***P<0.001. DPCs, dental
pulp cells; DSPP, dentin sialophosphoprotein; Runx2, runt-related
transcription factor 2; MI, mineralized inductive. |
Effect of LMK-235 on Alizarin Red S
staining of DPCs
The control group and the LMK-235 treated group
exhibited limited mineralized nodules stained by Alizarin Red S.
Conversely, the MI+LMK-235 group exhibited more and larger
calcified nodules and stronger staining when compared with the MI
group (Fig. 4A-D).
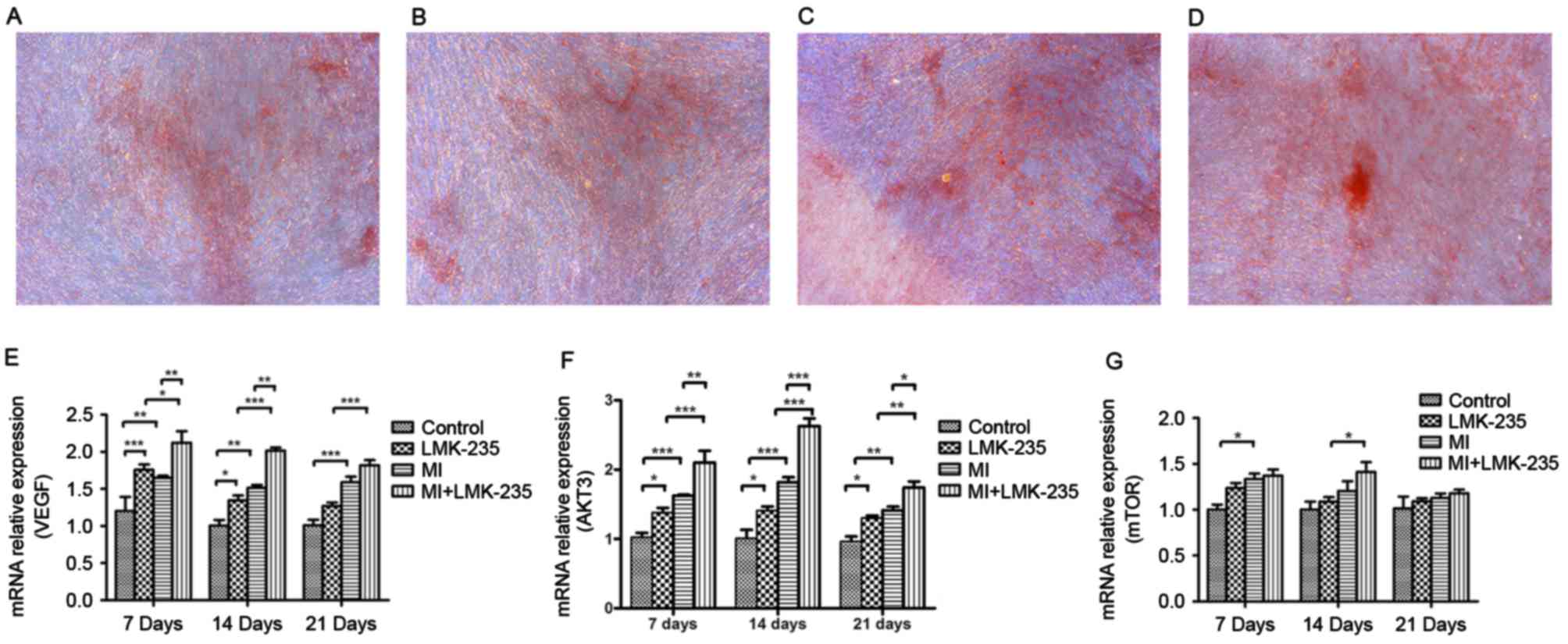 | Figure 4.Alizarin Red S staining and the effect
of the VEGF/AKT/mTOR pathway on odontoblast differentiation induced
by LMK-235. Alizarin Red S staining of the (A) control, (B)
LMK-235, (C) MI and (D) MI+LMK-235 groups (magnification, ×100).
mRNA expression levels of (E) VEGF, (F) AKT and (G) mTOR. Data are
presented as the mean ± standard deviation. *P<0.05,
**P<0.01, ***P<0.001. DSPP, dentin sialophosphoprotein; MI,
mineralized inductive; VEGF, vascular endothelial growth factor;
mTOR, mechanistic target of rapamycin; AKT, RAC-gamma
serine/threonine-protein kinase. |
VEGF/AKT/mTOR pathway may take part in
odontoblast differentiation induced by LMK-235
mRNA expression of VEGF and AKT3 in the MI+LMK-235
group was increased significantly compared with the MI group at
days 7 and 14. At day 21, the mRNA expression of AKT3 was notably
upregulated in the MI+LMK-235 group compared with that of the MI
group, whereas there was no significant difference between the mRNA
levels of VEGF in the MI+LMK-235 and the MI groups (Fig. 4E and F). mTOR mRNA expression
increased slightly with no significant difference in the MI+LMK-235
group compared with the MI group at days 7, 14 and 21 (Fig. 4G).
Discussion
The present study focused on the potential molecular
mechanism underlying the role of LMK-235 in odontoblast
differentiation of DPCs, which possibly would influence dentin
formation and regeneration process. In this experiment, the
proliferative potential of DPCs cultured with different
concentration of LMK-235 was investigated. High concentrations of
LMK-235 decreased the proliferation of DPCs compared with that of
the control group, whereas the proliferation of those DPCs treated
with low concentrations of LMK-235 (50 and 100 nM) was not
noticeably affected. These results were consistent with Paino et
al (12), who demonstrated
that low concentrations of VPA did not reduce cell viability of
either DPSCs or primary osteoblasts after 2 days of culture, where
as treatment with high concentrations of VPA for >48 h decreased
cell proliferation. To confirm the appropriate concentration of
LMK-235 that might promote DPC odontoblast differentiation, the
gene expression levels of specific factors during odontoblast
differentiation were determined by RT-qPCR. The results indicated
that DSPP, ALP and Runx2 mRNA expression levels, which were of
great importance in odontoblast differentiation (13), were increased in DPCs when cultured
with a low concentration of LMK-235, especially at the
concentration of 100 nM. In this sense, the concentration of 100
nMLMK-235 activated the odontoblast differential potency of DPCs
without reducing cell proliferation in vitro. According to
the results mentioned above, LMK-235 at a concentration of 100 nM
was subsequently used to study DPC odontoblast differentiation.
ALP is an important marker in the early phase of
odontoblast differentiation, and its activity has been reported to
markedly increase during this process (22). The ALP activity in this experiment
indicated that LMK-235 might exhibit a more obvious effect on
enhancing ALP activity of DPCs with the synergistic fact of
mineralizing medium. The mRNA expression of ALP was increased
markedly in the MI+LMK-235 group compared with the MI group at days
7 and 14, indicating hat LMK-235 might increase ALP expression
during the early stages of odontoblast differentiation. Changes
between ALP gene expression and activity in the LMK-235 treated
group and the control group were inconsistent, and this might be
caused by delayed protein expression. There was no significant
difference in ALP mRNA expression between the MI+LMK-235 and MI
groups at days 21. This effect might be due to that LMK-235
accelerated DPC odontoblast differentiation and ALP expression
reached a peak in the early stage; thus, there were no significant
differences that could be identified between the MI+LMK-235 and MI
groups at later stage.
DSPP, a member of small integrin-binding ligand
N-linked glycoproteins, is widely regarded as a specific marker of
odontoblast that regulates the progress of dentin formation, and
its protein is nearly 400× more expressed in the dentin than in the
bone (23). DSPP is reported to be
overexpressed at 1–3 days after injury in vivo and
upregulated during the early stages of odontoblast differentiation
in vitro (24). DSPP
expression was proven to be regulated by histone modification. Gu
et al (25) demonstrated
that after DPSC were cultured in osteo differentiation medium, the
increase of DSPP expression was associated with histone H3
acylation. Wang et al (26)
indicated that p300, a HAT, could promote gene expression of
odontoblast markers via enhancing acetylation of H3K9 in the
promoter regions of DSPP gene. In the present study, RT-qPCR
analysis demonstrated that DSPP mRNA expression in the MI+LMK-235
group was significantly upregulated when compared with that of the
MI group at days 7 and 14, and was slightly higher at day 21.
Protein expression examined by western blotting confirmed the
outcomes above. These results indicated that LMK-235 may increase
the expression of DSPP, especially during the early stages of
differentiation, which contributed to DPC odontoblast
differentiation.
Runx2, a crucial factor of odontoblast and
osteoblast differentiation at both the early and late stages, is
reported to be continuously upregulated during the differentiation
of mesenchymal stem cells, especially at the late period (27). The results of Runx2 expression was
not consistent as those by Jin et al (13) that Runx2 expression was not
affected in DPSCs treated with TSA. The reason behind these
differences might be that HDAC4 could modulate Runx2 activity
(28), and Runx2 gene and protein
can be upregulated due to the inhibition of HDAC4 by LMK-235. These
data above suggested that LMK-235 might improve the expression of
Runx2 during odontoblast differentiation. The expression of DSPP is
elevated in 3-day-old transgenic mice overexpressing Runx2, but
decreased in those mice by the age of 1 month (29). The phenomenon mentioned above that
DSPP expression was slightly higher in the MI+LMK-235 group
compared with the MI group may be due to the overexpression of
Runx2 at the late stage of DPSC odontoblast differentiation. The
potential molecular mechanism of DSPP and Runx2 expression
underlying the role of LMK-235 needs to be further studied.
OCN, a gamma-carboxyglutamic acid containing
protein, is always expressed during the late period of odontoblast
and osteoblast differentiation (30). The expression of OCN mRNA
demonstrated that OCN expression was not apparently altered by
LMK-235, indicating that LMK-235 might promote odontoblast
differentiation mainly at the early stage.
Alizarin Red S staining is regarded as a crucial
method to detect odontoblast differentiation (31). More and larger calcified nodules
and stronger staining was observed in the MI+LMK-235 group when
compared with the MI group at day 21, indicating that the formation
of calcified nodules was increased by LMK-235.
The present study also aimed to illustrate the
potential molecular mechanism of LMK-235 in odontoblast
differentiation. In recent years, the phosphoinositide 3-kinase
(PI3K)/AKT pathway has proven to serve a pivotal role in the growth
and osteogenic differentiation of mesenchymal stem cells in
vitro (32). AKT is regarded
as the chief target of PI3K signalling. Other extracellular signals
or growth factors, such as VEGF, might activate the PI3K/Akt
signalling pathway to regulate fundamental cellular processes. VEGF
silencing suppresses the proliferation of human SaOS-2 cells and
reduces angiogenesis by inactivating the VEGF/PI3K/AKT pathway.
Additionally, VEGF expression is regarded to be repressed by HDACs,
especially HDAC4 (33). Moreover,
an mTOR inhibitor attenuates osteoblast differentiation of human
periodontal ligament cells and osteoblasts (34). Therefore, it was hypothesized that
LMK-235 might promote odontoblast differentiation by activating he
VEGF/AKT/mTOR pathway through reducing HDAC4 activity to a certain
extent. The RT-qPCR results indicated that VEGF and AKT mRNA
expression in DPCs were increased markedly after odontoblast
induction, and the promotion effects were enhanced by LMK-235. mTOR
mRNA expression was slightly upregulated in the MI+LMK-235 group
when compared with that of the MI group. This phenomenon might be
caused by the fact that mTOR was partially activated, whereas some
other pathways downstream might participate in the subsequent
reaction of AKT activation. Thus, the VEGF/AKT/mTOR pathway might
take part in odontoblast differentiation induced by LMK-235.
In conclusion, the present study demonstrated that a
certain concentration of LMK-235, a specific HDAC4 and HDAC5
inhibitor, was proven to have the ability to promote DPC
odontoblast differentiation without reducing cell proliferation
in vitro, and the VEGF/AKT/mTOR signalling pathways might
take part in this process. Depending on these results above,
further studies on LMK-235 as a regulator of dental tissue
regeneration are necessary, providing therapeutic application
potential in the future.
Acknowledgements
The present study was supported the Medical Research
Foundation of Guangdong Province (Guangzhou, China; grant no.
A2016541), the President Foundation of Nanfang Hospital, Southern
Medical University (Guangzhou, China; grant no. 2014C020), the
Scientific Research Staring Foundation of Southern Medical
University (Guangzhou, China; grant no. PY2014N051), and the
National Natural Science Foundation of China (Beijing, China; grant
no. 81371137).
References
|
1
|
Jo YY, Lee HJ, Kook SY, Choung HW, Park
JY, Chung JH, Choung YH, Kim ES, Yang HC and Choung PH: Isolation
and characterization of postnatal stem cells from human dental
tissues. Tissue Eng. 13:767–773. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Seo BM, Miura M, Gronthos S, Bartold PM,
Batouli S, Brahim J, Young M, Robey PG, Wang CY and Shi S:
Investigation of multipotent postnatal stem cells from human
periodontal ligament. Lancet. 364:149–155. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ji YM, Jeon SH, Park JY, Chung JH, Choung
YH and Choung PH: Dental stem cell therapy with calcium hydroxide
in dental pulp capping. Tissue Eng Part A. 16:1823–1833. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Arnsdorf EJ, Tummala P, Castillo AB, Zhang
F and Jacobs CR: The epigenetic mechanism of mechanically induced
osteogenic differentiation. J Biomech. 43:2881–2886. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lehrmann H, Pritchard LL and Harel-Bellan
A: Histone acetyltransferases and deacetylases in the control of
cell proliferation and differentiation. Adv Cancer Res. 86:41–65.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Marks PA, Richon VM and Rifkind RA:
Histone deacetylase inhibitors: Inducers of differentiation or
apoptosis of transformed cells. J Natl Cancer Inst. 92:1210–1216.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jeon EJ, Lee KY, Choi NS, Lee MH, Kim HN,
Jin YH, Ryoo HM, Choi JY, Yoshida M, Nishino N, et al: Bone
morphogenetic protein-2 stimulates Runx2 acetylation. J Biol Chem.
281:16502–16511. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fu Y, Zhang P, Ge J, Cheng J, Dong W, Yuan
H, Du Y, Yang M, Sun R and Jiang H: Histone deacetylase 8
suppresses osteogenic differentiation of bone marrow stromal cells
by inhibiting histone H3K9 acetylation and RUNX2 activity. Int J
Biochem Cell Biol. 54:68–77. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Huynh NC, Everts V, Pavasant P and
Ampornaramveth RS: Inhibition of histone deacetylases enhances the
osteogenic differentiation of human periodontal ligament cells. J
Cell Biochem. 117:1384–1395. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee HW, Suh JH, Kim AY, Lee YS, Park SY
and Kim JB: Histone deacetylase 1-mediated histone modification
regulates osteoblast differentiation. Mol Endocrinol. 20:2432–2443.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nissen-Meyer LS, Svalheim S, Taubøll E,
Reppe S, Lekva T, Solberg LB, Melhus G, Reinholt FP, Gjerstad L and
Jemtland R: Levetiracetam, phenytoin, and valproate act differently
on rat bone mass, structure, and metabolism. Epilepsia.
48:1850–1860. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Paino F, La Noce M, Tirino V, Naddeo P,
Desiderio V, Pirozzi G, De Rosa A, Laino L, Altucci L and Papaccio
G: Histone deacetylase inhibition with valproic acid downregulates
osteocalcin gene expression in human dental pulp stem cells and
osteoblasts: Evidence for HDAC2 involvement. Stem Cells.
32:279–289. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jin H, Park JY, Choi H and Choung PH: HDAC
inhibitor trichostatin A promotes proliferation and odontoblast
differentiation of human dental pulp stem cells. Tissue Eng Part A.
19:613–624. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kwon A, Park HJ, Baek K, Lee HL, Park JC,
Woo KM, Ryoo HM and Baek JH: Suberoylanilide hydroxamic acid
enhances odontoblast differentiation. J Dent Res. 91:506–512. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lu J, Qu S, Yao B, Xu Y, Jin Y, Shi K,
Shui Y, Pan S, Chen L and Ma C: Osterix acetylation at K307 and
K312 enhances its transcriptional activity and is required for
osteoblast differentiation. Oncotarget. 7:37471–37486. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xia ZY, Hu Y, Xie PL, Tang SY, Luo XH,
Liao EY, Chen F and Xie H: Runx2/miR-3960/miR-2861 positive
feedback loop is responsible for osteogenic transdifferentiation of
vascular smooth muscle cells. Biomed Res Int. 2015:6240372015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hansen FK, Sumanadasa SD, Stenzel K, Duffy
S, Meister S, Marek L, Schmetter R, Kuna K, Hamacher A, Mordmüller
B, et al: Discovery of HDAC inhibitors with potent activity against
multiple malaria parasite life cycle stages. Eur J Med Chem.
82:204–213. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li A, Liu Z, Li M, Zhou S, Xu Y, Xiao Y
and Yang W: HDAC5, a potential therapeutic target and prognostic
biomarker, promotes proliferation, invasion and migration in human
breast cancer. Oncotarget. 7:37966–37978. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ganai SA: Histone deacetylase inhibitor
sulforaphane: The phytochemical with vibrant activity against
prostate cancer. Biomed Pharmacother. 81:250–257. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fu M, Shi W, Li Z and Liu H: Activation of
mPTP-dependent mitochondrial apoptosis pathway by a novel pan HDAC
inhibitor resminostat in hepatocellular carcinoma cells. Biochem
Biophys Res Commun. 477:527–533. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu G, Xu G, Gao Z, Liu Z, Xu J, Wang J,
Zhang C and Wang S: Demineralized dentin matrix induces
odontoblastic differentiation of dental pulp stem cells. Cells
Tissues Organs. 201:65–76. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Suzuki S, Sreenath T, Haruyama N,
Honeycutt C, Terse A, Cho A, Kohler T, Müller R, Goldberg M and
Kulkarni AB: Dentin sialoprotein and dentin phosphoprotein have
distinct roles in dentin mineralization. Matrix Biol. 28:221–229.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen Y, Zhang Y, Ramachandran A and George
A: DSPP is essential for normal development of the
dental-craniofacial complex. J Dent Res. 95:302–310. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gu S, Liang J, Wang J and Liu B: Histone
acetylation regulates osteodifferentiation of human dental pulp
stem cells via DSPP. Front Biosci (Landmark Ed). 18:1072–1079.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang T, Liu H, Ning Y and Xu Q: The
histone acetyltransferase p300 regulates the expression of
pluripotency factors and odontogenic differentiation of human
dental pulp cells. PLoS One. 9:e1021172014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang X, Yang M, Lin L, Chen P, Ma KT,
Zhou CY and Ao YF: Runx2 overexpression enhances osteoblastic
differentiation and mineralization in adipose-derived stem cells in
vitro and in vivo. Calcif Tissue Int. 79:169–178. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sun X, Wei L, Chen Q and Terek RM: HDAC4
represses vascular endothelial growth factor expression in
chondrosarcoma by modulating RUNX2 activity. J Biol Chem.
284:21881–21890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li S, Kong H, Yao N, Yu Q, Wang P, Lin Y,
Wang J, Kuang R, Zhao X, Xu J, et al: The role of runt-related
transcription factor 2 (Runx2) in the late stage of odontoblast
differentiation and dentin formation. Biochem Biophys Res Commun.
410:698–704. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cutarelli A, Marini M, Tancredi V,
D'Arcangelo G, Murdocca M, Frank C and Tarantino U: Adenosine
Triphosphate stimulates differentiation and mineralization in human
osteoblast-like Saos-2 cells. Dev Growth Differ. 58:400–408. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tsukamoto Y, Fukutani S, Shin-Ike T,
Kubota T, Sato S, Suzuki Y and Mori M: Mineralized nodule formation
by cultures of human dental pulp-derived fibroblasts. Arch Oral
Biol. 37:1045–1055. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tong Y, Feng W, Wu Y, Lv H, Jia Y and
Jiang D: Mechano-growth factor accelerates the proliferation and
osteogenic differentiation of rabbit mesenchymal stem cells through
the PI3K/AKT pathway. BMC Biochem. 16:12015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dung TT, Yi YS, Heo J, Yang WS, Kim JH,
Kim HG, Park JG, Yoo BC, Cho JY and Hong S: Critical role of
protein L-isoaspartyl methyltransferase in basic fibroblast growth
factor-mediated neuronal cell differentiation. BMB Rep. 49:437–442.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bae WJ, Auh QS, Kim GT, Moon JH and Kim
EC: Effects of sodium tri- and hexameta-phosphate in vitro
osteoblastic differentiation in Periodontal Ligament and
Osteoblasts, and in vivo bone regeneration. Differentiation.
92:257–269. 2016. View Article : Google Scholar : PubMed/NCBI
|














