Introduction
Lung cancer is an important public health concern
and the leading cause of cancer-associated mortality worldwide
(1). Pathological findings have
indicated that lung cancer is a respiratory disease that may be
associated with air contamination caused by industrial pollution
(2). Non-small cell lung carcinoma
(NSCLC) and small cell lung carcinoma are the two types of lung
cancer, with incidence rates of ~85% and ~15%, respectively,
according to a clinical statistical analysis of human cancers
(3). NSCLCs include large-cell
carcinoma, squamous cell carcinoma and adenocarcinoma, the
incidences of which are increasing (4–6).
NSCLC represents ~85% of all cases of lung cancer and results in a
subsequently high mortality rate (7). NSCLC is initiated from the non-small
cells in the lung, and is the most common type of cancer worldwide
(8). According to clinical
statistical investigations into lung cancer, >80% of newly
diagnosed patients are in the middle or severe stages of the
disease (9–11).
Although continuing investigations have sought to
improve the efficacy of treatment for patients with NSCLC, the
5-year survival rate remains poor at <15% (5,12,13).
In addition, the majority of newly diagnosed patients with NSCLC
are at an advanced stage, contributing to the high mortality rate
(14,15). Previous studies have reported that
the migration and invasion of NSCLC cells are the primary causes of
the poor survival rate during treatment and the rate of recurrence
for patients with NSCLC (16,17).
As a result, the development of effective agents that inhibit
migration and invasion has become a necessity for the treatment of
patients with cancer (18,19).
Radiotherapy, chemotherapy and other treatments used
in patients with locally advanced NSCLC have been demonstrated to
improve outcomes more effectively than up-front surgical resection
(20–22). These conventional therapies have
been demonstrated to be effective at temporarily controlling the
disease (23,24); however, previous studies on
nasopharyngeal carcinoma have demonstrated that high-dose
irradiation and/or chemical drugs may be associated with an
increased probability of toxicity and damage to the immune system,
which led to more rapid migration and invasion (25–27).
A number of previous studies have proposed more advanced
oncotherapeutic techniques to improve patient outcomes, including
virotherapy, immunotherapy, targeted therapy, stem cell therapy,
gene therapy and comprehensive care (28–31).
According to previous analyses of therapeutic efficacy, targeted
therapy exhibits the potential for human cancer treatment and the
possibility of eradicating tumor cells; for example, a tumor-free
condition was attained by targeting the digoxin-specific antigen of
tumor cells (32,33).
Metastasis-associated protein 2 (MTA2) is a member
of the MTA family of chromatin remodeling proteins, which have been
reported to serve roles in tumor progression and metastasis
(34,35). Previous studies have demonstrated
that MTA2 is overexpressed in a number of cancers, which indicated
that the inhibition of MTA2 activity may be beneficial for the
treatment of human cancer (36,37).
Additional studies have indicated that MTA2 may be required for the
invasion and metastasis of human pituitary adenomas through the
epithelial-mesenchymal transition pathway, and overexpression of
MTA2 may contribute to breast cancer cell growth, resulting in an
enhanced anchorage-independent growth and metastasis (38,39).
In addition, MTA2 has been associated with bone invasion and tumor
stage in human pituitary adenomas (40). These previous reports suggested
that MTA2 may be a potential target for inhibiting tumor growth and
aggressiveness in the treatment of cancer.
The present study investigated the expression and
function of MTA2 and MTA2-mediated signaling in NSCLC cells.
Although our previous study suggested that MTA2 may promote the
metastasis of NSCLC by inhibiting the expression of epithelial cell
adhesion molecule (Ep-CAM) and E-cadherin, the mechanisms of
MTA2-mediated signaling pathway have not been elucidated in NSCLC
cells (41). The results of the
present study demonstrated that the extracellular signal-regulated
kinase (ERK) and RAC-α serine/threonine protein kinase (AKT)
signaling pathways may be involved in the progression of
MTA2-mediated migration and invasion of NSCLC cells. Additionally,
the inhibitory effects of MTA2-mediated tumor growth were analyzed
in vitro and in vivo.
Materials and methods
Ethics statement
The present study was performed in accordance with
the recommendations in The Guide for the Care and Use of Laboratory
Animals of Tianjin Medical University (Tianjin, China).
Experimental protocols were approved by The Chinese Association for
Laboratory Animal Science (Beijing, China). All surgeries and
euthanasia were performed under sodium pentobarbital anesthesia,
and all efforts were made to minimize suffering.
Cell culture and reagents
A549 and H358 human lung carcinoma cells were
purchased from The American Type Culture Collection (ATCC;
Manassas, VA, USA). A549 and H358 cells were cultured in RPMI-1640
medium (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 10% heat-inactivated FBS (Gibco; Thermo Fisher
Scientific, Inc.), 3 mM L-glutamine, 50 µg/ml gentamicin
(BioWhittaker; Lonza Group, Ltd., Basel, Switzerland) and 1%
penicillin/streptomycin. The MRC-5 (no. 55-X™; ATCC) normal lung
cell line were cultured in minimum essential medium (MEM; Gibco;
Thermo Fisher Scientific, Inc.) supplemented with 10%
heat-inactivated FBS. Cells were cultured at 37°C with 5%
CO2. All cells were demonstrated to be free from
mycoplasma contamination.
Transfection of micro (mi)RNA mimics
and small interfering (si)RNA
All siRNAs were synthesized by Invitrogen (Thermo
Fisher Scientific, Inc.), including si-MTA2 and siRNA-control
(si-MTA2 sense strand, 5′-UGAACAAGACAGAGCUCAATT-3′ and antisense
strand, 5′-UUGAGCUCUGUCUUGUUCATT-3′; siRNA-control sense strand,
5′-UGAUGAUCCACCAAGAGCUCUUGCC-3′ and antisense strand,
5′-UUGAGCUCUGUCUUGUUCATT-3′; miMTA2 sense strand,
5′-CACTCGAGAGTCCACCTCCAGTGTAGdTdT-3′ and antisense strand,
3′-dTdTCAGCGGCCGCAGTCAATGGAATGCTTG-5′; miRNA mimics sense strand,
5′-CGUGAUUGCGAGACUCUGAdTdT-3′ and antisense strand,
3′-dTdTGCACUAACGCUCUGAGACU-5′). A549 cells (1×106) were
transfected with 100 pmol plentivirus-si-MTA2 or
plentivirus-siRNA-control at 25°C for 48 h (Ambion; Thermo Fisher
Scientific, Inc.) using the Cell Line Nucleofector kit L and a
Nucleofector I electroporation device according to a prewritten
program (both from Lonza Group, Ltd.). All procedures were
performed according to the manufacturer's instructions. The
efficiency was determined by RT-qPCR (data not shown) as described
below.
Transfection of pMTA2
A549 cells (1×106) were cultured MEM with
5% FBS in six-well plate until 90% confluence. The media was
subsequently removed. The MTA2 gene (GenBank, Y14808.1) was
synthesized and cloned into pCMVp-NEO-X system (Takara
Biotechnology Co., Ltd., Dalian, China). The recombinant vector was
named pCMVp-NEO-MTA2 (pMTA2). Cells were transfected by
pCMVp-NEO-MTA (2 µg) using Lipofectamine 2000 (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany), according to the manufacturer's
instructions. Following 72 h transfection, subsequent
experimentations were performed.
ELISA analysis
The affinity of the antibody against (Ab)MTA2 (cat.
no. ab8106; Abcam, Cambridge, UK) with MTA2 was analyzed using an
MTA2 commercial ELISA kit (cat. no. M7569-200UL; Thermo Fisher
Scientific, Inc.), according to manufacturer's instructions.
Briefly, A549 cells (1×103) were cultured in 96-well
plates (Invitrogen; Thermo Fisher Scientific, Inc.) pre-coated
overnight at 4°C with AbMTA2, blocked with 1% bovine serum albumin
(BSA; Sigma-Aldrich; Merck KGaA) in PBS for 1 h at 4°C, and
incubated with standard MTA2 dilutions for 2 h at 37°C.
Subsequently, AbMTA2 for 30 min at 37°C followed by washes with PBS
three times. The results were measured using an ELISA reader
(Bio-Rad Laboratories, Inc., Hercules, CA, USA) reader at a
wavelength of 450 nm.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from A549 cells
(1×108) pre- or post-treatment with AbMTA2 (5 mg/ml,
1×108), siMTA2 transfection (1×108) or pMTA2
transfection (1×108) using an RNAeasy Mini kit (Qiagen
Sciences, Inc., Gaithersburg, MD, USA) according to the
manufacturer's instructions. Total RNA (1 µg) was reverse
transcribed into cDNA using the Reverse Transcription kit (Qiagen
Sciences, Inc.) and the quality was confirmed using 1.5% agarose
(Sigma-Aldrich; Merck KGaA) electrophoresis. cDNA (10 ng) was
subjected to qPCR analysis with SYBR Green Master Mix system
(Bio-Rad Laboratories, Inc.), according to the manufacturer's
instructions. All the forward and reverse primers were synthesized
by Invitrogen; Thermo Fisher Scientific, Inc. (Table I). Cycling conditions were as
follows: 45 cycles of denaturation at 95°C for 2 min, annealing at
66°C for 30 sec with touchdown to 56°C for 30 sec, and extension at
72°C for 10 min. Relative mRNA expression changes were calculated
using the 2−ΔΔCq method (42). Experiments were repeated three
times and the results are expressed as the n-fold change relative
to the control.
 | Table I.Sequences of primers. |
Table I.
Sequences of primers.
|
| Sequence |
|---|
|
|
|
|---|
| Gene name | Reverse | Forward |
|---|
| VEGFR1 |
5′-TCACTGCCACTCTAATTGTC-3′ |
5′-CCATATGCGGTACAAGTCA-3′ |
| VEGFR2 |
5′-AAGGCGAGACCTGCATTC-3′ |
5′-CTGCCCTCTTCTGAGCTCT-3′ |
| VEGFR3 |
5′-AGCCATTCATCAACAAGCCT-3′ |
5′-GGCAACAGCTGGATGTCATA-3′ |
| VEGF |
5′-TGCATTCACATTGTGCTGCTGTAG-3′ |
5′-GCAGATTATGCGGATCAAACC-3′ |
| EGF |
5′-CATCCAGTGAGACCAATGAG-3′ |
5′-GTAGCCGCCAGTTCACCATT-3′ |
| Ep-CAM |
5′-TGCTGAACTGAAGTACACTGGCATTGGTTTTG-3′ |
5′-CCTGAACTGAAGTACTGGCATTGGTCAGTCA-3′ |
| TAF |
5′-ACTGGCAGTATGTGCACTGC-3′ |
5′-CAGCCTGGGTCAGGGTCAATCCCT-3′ |
| β-actin |
5′-CATCTCTTGCTCGAAGTCCA-3′ |
5′-ATCATGTTTGAGACCTTCAACA-3′ |
MTT assay
A549, pMTA2-, or siMTA2-transfected cells
(1×103 cells/well) were incubated with AbMTA2 in 96-well
plates for 72 h at 37°C in triplicate for each condition. At each
time-point, 20 µl MTT (5 mg/ml) in PBS solution was added to each
well and the plate was further incubated for 4 h at 37°C. The
medium was removed and 100 µl dimethyl sulfoxide was added to the
wells to solubilize the crystals. The optical density was measured
using an ELISA microplate reader (Bio-Rad Laboratories, Inc.) at
450 nm.
Induction of apoptosis in A549
cells
A549, pMTA2-, siMTA2-transfected or AbMTA2-treated
cells (1×106) were cultured in six-well plates until 80%
confluence was reached. Apoptosis was assessed by incubating the
cells with chemotherapeutic agent Taxol® (5 mg/ml) or
PBS for 72 h at 37°C. Following incubation with AbMTA2, the cells
were trypsinized and harvested by centrifuging at 2,000 × g for 10
min at room temperature. The cells were washed in cold PBS,
adjusted to 1×106 cells/ml with PBS, labeled with
Annexin V-fluorescein isothiocyanate (FITC)/propidium iodide
(Annexin V-FITC kit; BD Biosciences, Franklin Lakes, NJ, USA), and
analyzed using a FACScan flow cytometer (BD Biosciences) and FlowJo
10.0.7 software (Tree Star, Inc., Ashland, OR, USA).
Cell invasion and migration
assays
A549, pMTA2-, siMTA2-transfected, AbMTA2-treated or
PBS-treated cells were seeded into the upper chamber of each
insert. Subsequently, 500 µl DMEM containing 10% FBS was added to a
24-well plate for 24 h. Migration and invasion analysis of cells
was conducted in a 24-well culture plate with chamber inserts (BD
Biosciences). For the migration assays, 1×104 cells/well
were placed into the upper chamber with a non-coated membrane. For
the invasion assays, cells (5×104 cells/well) were
placed into the upper chamber with a Matrigel-coated membrane. All
procedures were performed according to the manufacturer's
instructions. The cells were fixed and stained for 30 min in a 0.1%
crystal violet solution in PBS. The tumor cell invasion and
migration was counted in at least three random fields/membrane, by
light microscopy (Olympus Corporation, Tokyo, Japan) at ×40
magnification.
Western blot analysis
A549 cells (1×107) were treated with
AbMTA2 (40 ng/ml) for 72 h. The A549 cells were harvested by
scraping and lysed in radioimmunoprecipitation assay buffer
(Invitrogen; Thermo Fisher Scientific, Inc.) followed by
homogenization at 4°C for 10 min. Protein concentration was
measured with a bicinchoninic acid protein assay kit (Thermo Fisher
Scientific, Inc.). Proteins (10 µg) were analyzed by 15% SDS-PAGE
and transferred onto polyvinylidene fluoride membranes (EMD
Millipore, Billerica, MA, USA). Membranes were blocked with 5% BSA
(Sigma-Aldrich; Merck KGaA) for 2 h at room temperature and probed
with antibodies (Abcam) against the following proteins: MTA2
(1:1,000; cat. no. ab5392), ERK (1:1,000; cat. no. ab17942), AKT
(1:1,000; cat. no. ab8805), matrix metalloproteinase 2 (1:5,000;
MMP2; cat. no. ab7033), MMP-9 (cat. no. ab38898), cardiotrophin-1
(CT-1; cat. no. ab80527), rRNA 2′-O-methyltransferase fibronectin
(FIB; 1:1,000; cat. no. ab2413), E-cadherin (1:1,000; cat. no.
ab76055), Snail (1:1,000; cat. no. ab53519), caspase-3 (1:2,000;
cat. no. ab13585), Bcl2-associated agonist of cell death (Bad;
1:1,000; cat. no. ab32060), N-cadherin (1:2,000; cat. no. ab18203),
p-ERK (1:2,000; cat. no. ab192591), pAKT (1:2,000; cat. no.
ab81283), vimentin (1:1,000; cat. no. ab92547) and β-actin
(1:2,000; cat. no. ab8827) for 12 h at 4°C. The horseradish
peroxidase-conjugated anti-rabbit IgG secondary antibody (cat. no.
VPA00764; Bio-Rad Laboratories, Inc.) was used at a 1:5,000
dilution for 2 h at 4°C and detected using enhanced
chemiluminescence substrate ECL Select™ (GE Healthcare Life
Sciences, Little Chalfont, UK), according to the manufacturer's
instructions. The density of the bands was analyzed using Quantity
One software version 4.62 (Bio-Rad Laboratories, Inc.).
Immunofluorescence
A549 cells (1×106) were cultured in
six-well plates until a 90% confluent monolayer was achieved. The
cells were subsequently incubated with a mouse anti-human MTA2
primary antibody (1:1,000; cat. no. ab5392; Abcam) for 12 h at 4°C.
The cells were washed with PBS to completely remove the residual
antibody and incubated with horseradish peroxidase-conjugated
anti-rabbit IgG secondary antibody (1:5,000; cat. no. 1721019;
Bio-Rad Laboratories, Inc.) for 2 h at 37°C. Following washing in
PBS, cells were mounted with anti-fade reagent DAPI (Invitrogen;
Thermo Fisher Scientific, Inc.) for 2 h at 37°C and viewed with
fluorescent microscope (Olympus Corporation, Tokyo, Japan) at ×40
magnification.
Immunohistochemistry
Tumors from NSCLC carcinoma xenograph mice treated
with AbMTA2 or PBS were excised on day 30 and fixed using 10%
formaldehyde followed by embedding in paraffin wax, and cut into
serial sections (4 µm). Tissues were washed with PBS-Tween-20
(PBST) three times at room temperature and antigen retrieval was
performed on the tumor sections using a microwave heating AR
(43) subsequent to a series of
ethanols (100, 95 and 80%). Tumor sections were washed with PBST at
room temperature and incubated with primary antibodies: EGF (1:500;
cat. no. ab9695), and VEGF (1:500; cat. no. ab32152) (both from
Abcam) at 37°C for 2 h. Then, sections were blocked with 5% BSA
(Sigma-Aldrich; Merck KGaA) at 37°C for 2 h and horseradish
peroxidase-conjugated anti-rabbit IgG secondary antibody (Bio-Rad
Laboratories, Inc.) was used to incubate primary antibodies at a
1:5,000 dilution at 37°C for 2 h. A Ventana Benchmark Automated
Staining System (Ventana Medical Systems, Inc., Tucson, AZ, USA)
was used for the observation of protein expression.
Animal study
A total of 80 specific pathogen-free female nude
mice (age, 6 weeks; body weight, 30–36 g) were purchased from
Shanghai SLAC Laboratory Animal Co., Ltd. (Shanghai, China). All
animals were given free access to food and water and housed in a
temperature-controlled facility at 23±1°C and relative humidity of
50±5% with a 12-h light/dark cycle. Mice were subcutaneously
implanted with MTA2-overexpression (n=20), MTA2-silenced (n=20) or
A549 tumor cells (1×106 cells) (n=20), AbMTA2 (n=20).
The A549 group was divided into two groups (n=20/group) and
received intravenously injected AbMTA2 (500 ng/kg) or PBS (500
ng/kg). Treatments began on day 7 following tumor cell
implantation, when the tumor diameter reached 5–7 mm. The treatment
was continued for 7 days at a frequency of once per day. The tumor
volumes were calculated according to a previous study (44). Animals (n=15 in each group) were
housed for 120 days to investigate the role of MTA2 in NSCLC.
Statistical methods
All data are presented as the mean ± standard error
of the mean of triplicate experiments, and were analyzed using
GraphPad Prism version 5.0 software (GraphPad Software, Inc., La
Jolla, CA, USA). Unpaired data were analyzed using Student's t-test
and comparisons of data between multiple groups were performed
using one-way analysis of variance followed by Whitney rank test or
Fisher's exact test. The Kaplan-Meier test was used to estimate the
risk of relapse and re-treatment during a 120-day treatment.
P<0.05 was considered to indicate a statistically significant
difference.
Results
MTA2 expression and its function in
NSCLC cells
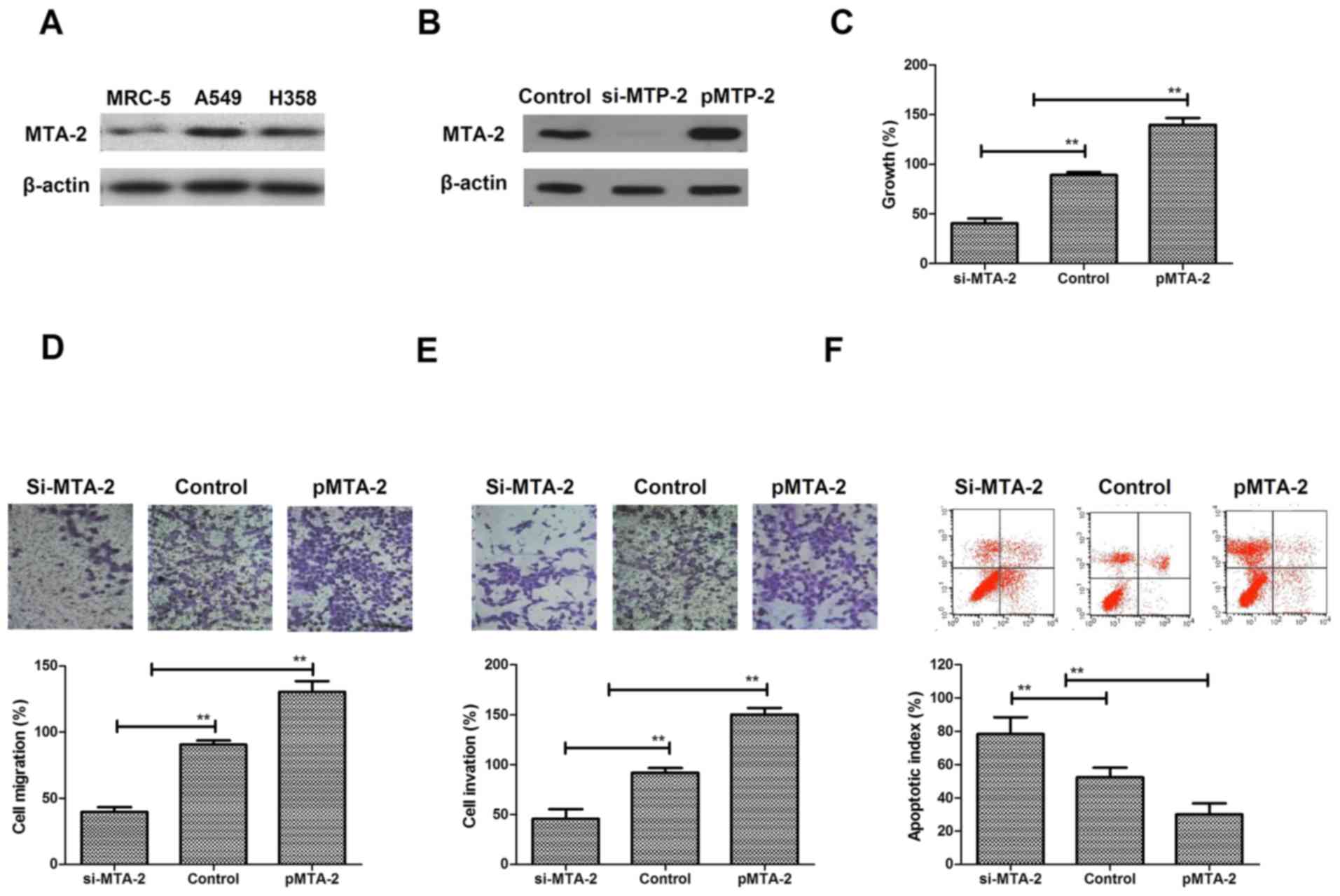
A comparison of the levels of expression revealed
that the MTA2 protein was expressed at notably higher levels in two
NSCLC cell lines, A549 and H358, compared with a normal lung cell
line, MRC-5 (Fig. 1A). To analyze
the function of MTA2 in NSCLC cells, A549 cell cultures were
established by transfection with MTA2 overexpression plasmids
(pMTA2) or lentivirus-mediated siRNA-MTA2. MTA2 expression was
increased following transfection with pMTA2, and decreased
following si-MTA2 transfection (Fig.
1B). It was observed that MTA2 overexpression resulted in a
significant increase in A549 cell growth, whereas si-MTA2
transfection inhibited NSCLC growth (Fig. 1C). Migration and invasion assays
demonstrated that MTA2 is positively associated with aggressiveness
(Fig. 1D and E) MTA2
downregulation inhibited migration and invasion of NSCLC cells,
while MTA2 overexpression promoted migration and invasion of NSCLC
cells. The apoptosis assay demonstrated that pMTA2 decreased the
apoptosis of A549 cells induced by the chemotherapeutic agent
Taxol® compared to control (Fig. 1F). The results of the present study
indicated that MTA2 proteins are more highly expressed in NSCLC
cells and may be positively associated with growth, aggressiveness
and apoptotic resistance of NSCLC cells.
AbMTA2 suppresses MTA2-induced
aggressiveness in NSCLC cells
The efficacy of AbMTA2 was assessed in NSCLC cells,
which indicated that AbMTA2 was able to specifically bind to MTA2,
as determined by ELISA analysis (Fig.
2A). Immunofluorescence and western blotting experiments
demonstrated that AbMTA2 efficiently suppressed MTA2 protein
expression in A549 cells (Fig.
2B). In addition, AbMTA2-suppressed expression cells exhibited
decreased expression of MTA2 target genes, including MMP-2, MMP-9,
CT-I and FIB compared with untransfected control cells (Fig. 2C). In addition, migration and
invasion assays demonstrated that AbMTA2 markedly inhibited the
aggressiveness of A549 cells (Fig. 2D
and E). Furthermore, apoptotic resistance was inhibited in A549
cells following treatment with AbMTA2 for 48 h (Fig. 2F). The results of the present study
indicated that AbMTA2 may be a potential agent for the inhibition
of migration and invasion in NSCLC cells.
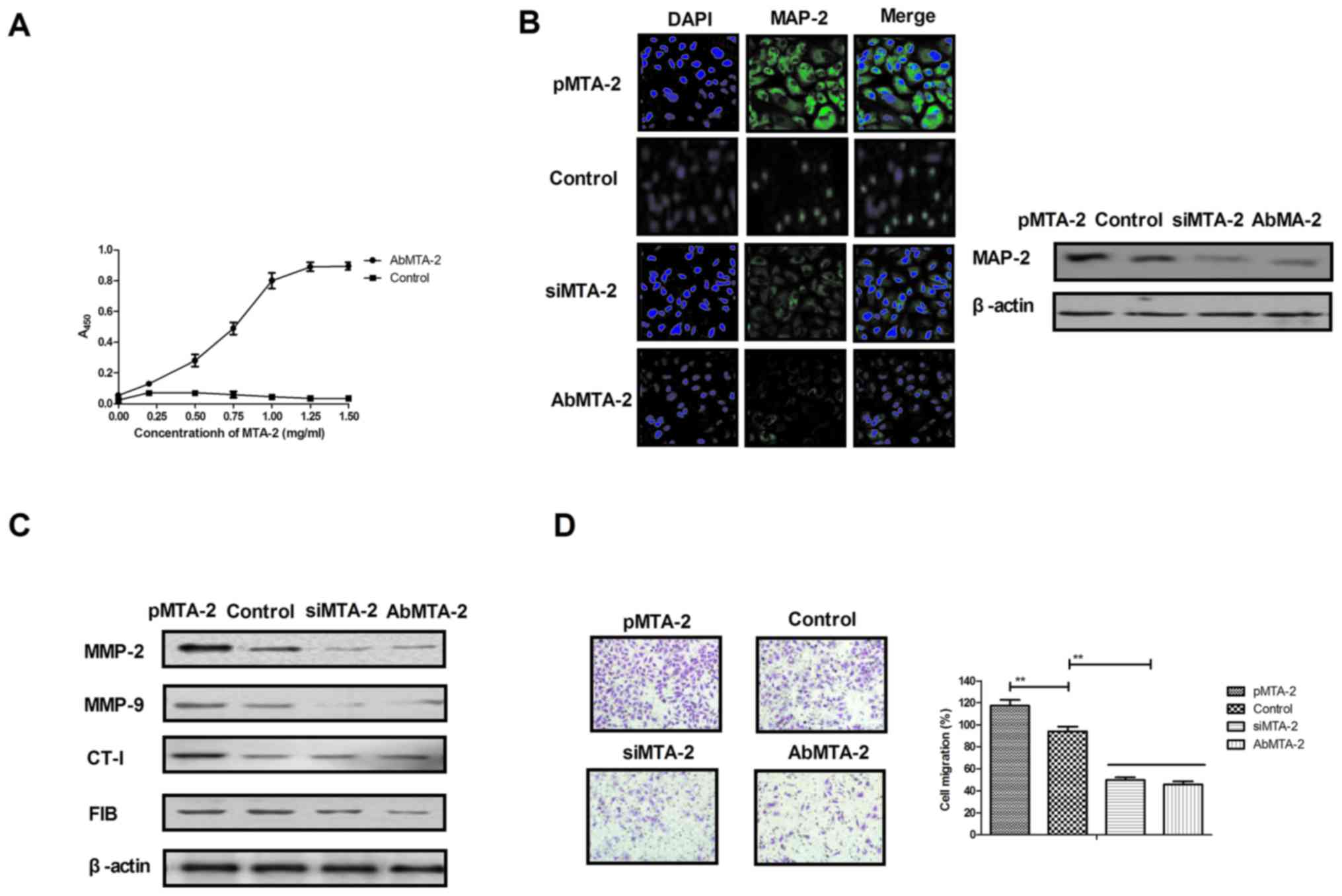 | Figure 2.Targeting MTA2 suppresses the growth
and aggressiveness of non-small cell lung carcinoma cells. (A)
Affinity of AbMTA2 with MTA2 as determined by ELISA analysis. (B)
Immunofluorescence and western blotting experiments analyzed the
efficiency of AbMTA2 targeting for MTA. Magnification, ×200. (C)
Expression of MTA2 target genes in A549 cells. (D) Migration of
A549 cells following treatment with AbMTA2. Magnification, ×400.
(E) Invasion of A549 cells following treatment with AbMTA2.
Magnification, ×400. (F) Apoptotic resistance of A549 cells
following treatment with AbMTA2. The data are presented as the mean
± standard error of the mean of three independent experiments.
**P<0.01. MTA2, metastasis associated protein MTA2; siRNA, small
interfering RNA; MMP-2, 72 kDa type IV collagenase; MMP-9, matrix
metalloproteinase-9; CT-1, cardiotrophin-1; FIB, rRNA
2′-O-methyltransferase fibrillarin; Ab, antibody. |
MTA2 regulates migration and invasion
of NSCLC cells via the ERK/AKT and VEGF signaling pathways
To analyze the mechanisms underlying MTA2-mediated
growth and metastasis in NSCLC cells, the ERK/AKT and VEGF
signaling pathways were analyzed. The protein expression levels of
E-cadherin, Snail, N-cadherin and Vimentin were analyzed, and it
was observed that AbMTA2 markedly inhibited the expression of these
proteins in A549 cells (Fig. 3A).
To confirm that AbMTA2 inhibited the epithelial-mesenchymal
transition process via the ERK/AKT signaling pathway, the protein
expression levels of ERK and AKT were analyzed. The data
demonstrated that pMTA2 increased, while si-MTA2 and AbMTA2
suppressed ERK and AKT expression in A549 cells compared with
untreated control cells (Fig. 3B).
The expression levels of apoptosis-associated proteins Caspase-3
and Bad also exhibited a decreased expression following treatment
with AbMTA2 and si-MTA2 (Fig. 3C).
However, pMTA2 increased the expression of caspase-3 and Bad in
A549 cells.
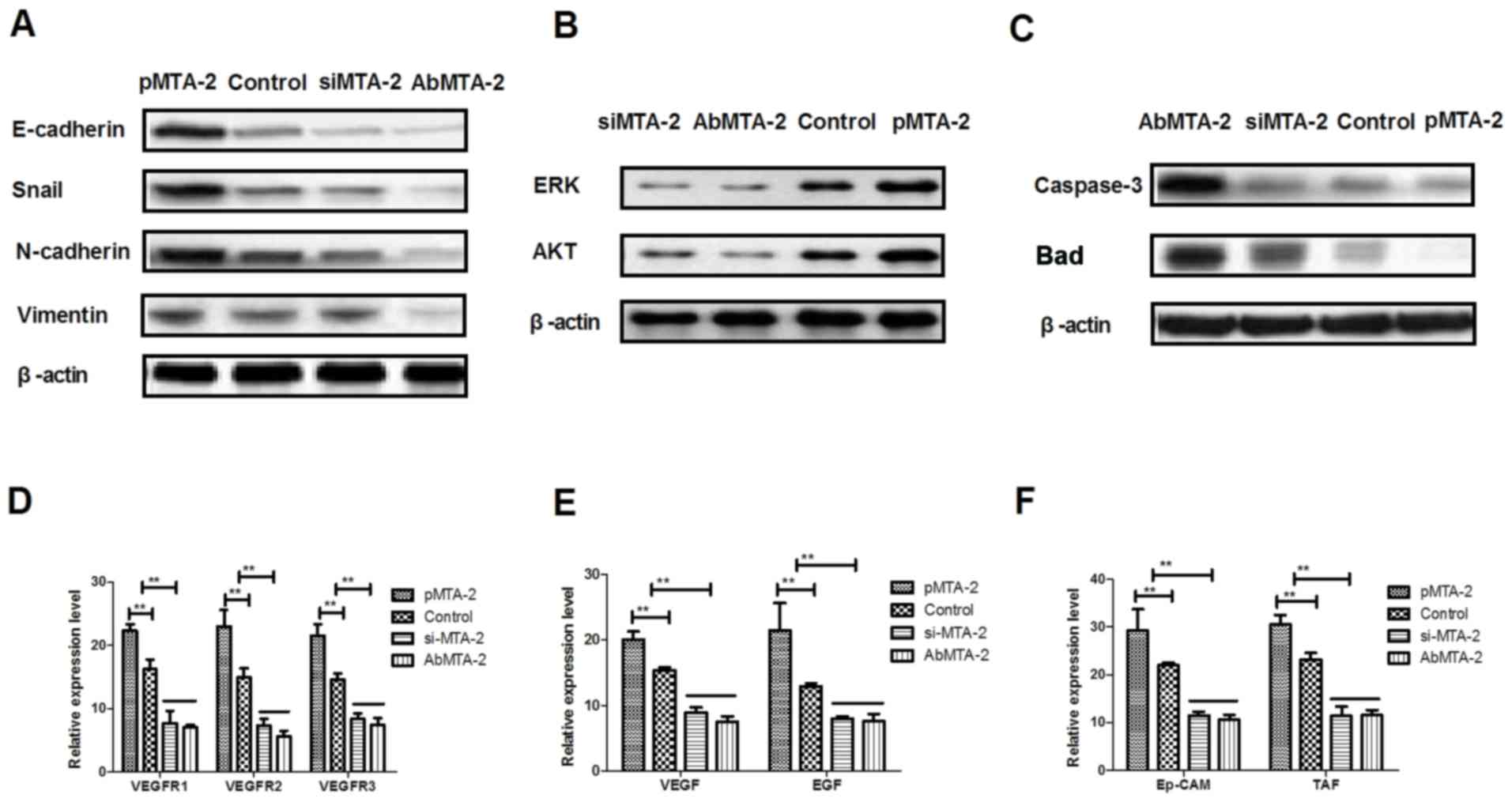 | Figure 3.Mechanism of MTA2-mediated signaling
pathways in non-small cell lung carcinoma cells. (A) E-cadherin,
Snail, N-cadherin and vimentin expression levels in A549 cells
following treatment with the indicated agents. (B) ERK and AKT
expression in A549 cells following treatment with the indicated
agents. (C) Apoptosis-associated proteins caspase-3 and Bad
expression levels in A549 cells. (D) VEGFR1, VEGFR2 and VEGFR3
expression levels in AbMTA2-treated A549 cells. (E) VEGF and EGF
expression levels in AbMTA2-treated A549 cells. (F) Ep-CAM and TAF
expression in A549 cells. The data are presented as the mean ±
standard error of the mean of three independent experiments.
**P<0.01 vs. control. MTA2, metastasis associated protein MTA2;
Snail, zinc-finger protein SNAI1; ERK, extracellular
signal-regulated kinase; AKT, RAC-α serine/threonine protein
kinase; Bad, Bcl2-associated agonist of cell death; VEGFR, vascular
endothelial growth factor receptor; EGF, pro-epidermal growth
factor; Ep-CAM, epithelial cell adhesion molecule; TAF, tumor
angiogenesis factor; si, small interfering; Ab, antibody. |
It was also demonstrated that VEGF and EGF mRNA
expression levels were decreased following treatment with AbMTA2
and si-MTA2 in A549 cells (Fig.
3E). AbMTA2 and si-MTA2 resulted in the decreased mRNA
expression of Ep-CAM and tumor angiogenesis factor in A549 cells,
while pMTA2 exerted the reverse effect (Fig. 3F). The present results demonstrated
that si-MTA2 exerted similar effects on inhibition of ERK/AKT and
VEGF signaling pathways in NSCLC cells, while pMTA2 promoted
ERK/AKT and VEGF signaling pathways in NSCLC cells. The results of
the present study demonstrated that MTA2 regulated the expression
of components of the ERK/AKT and VEGF signaling pathways in A549
cells in vitro.
In vivo inhibitory effects of AbMTA2
on A549-bearing mice
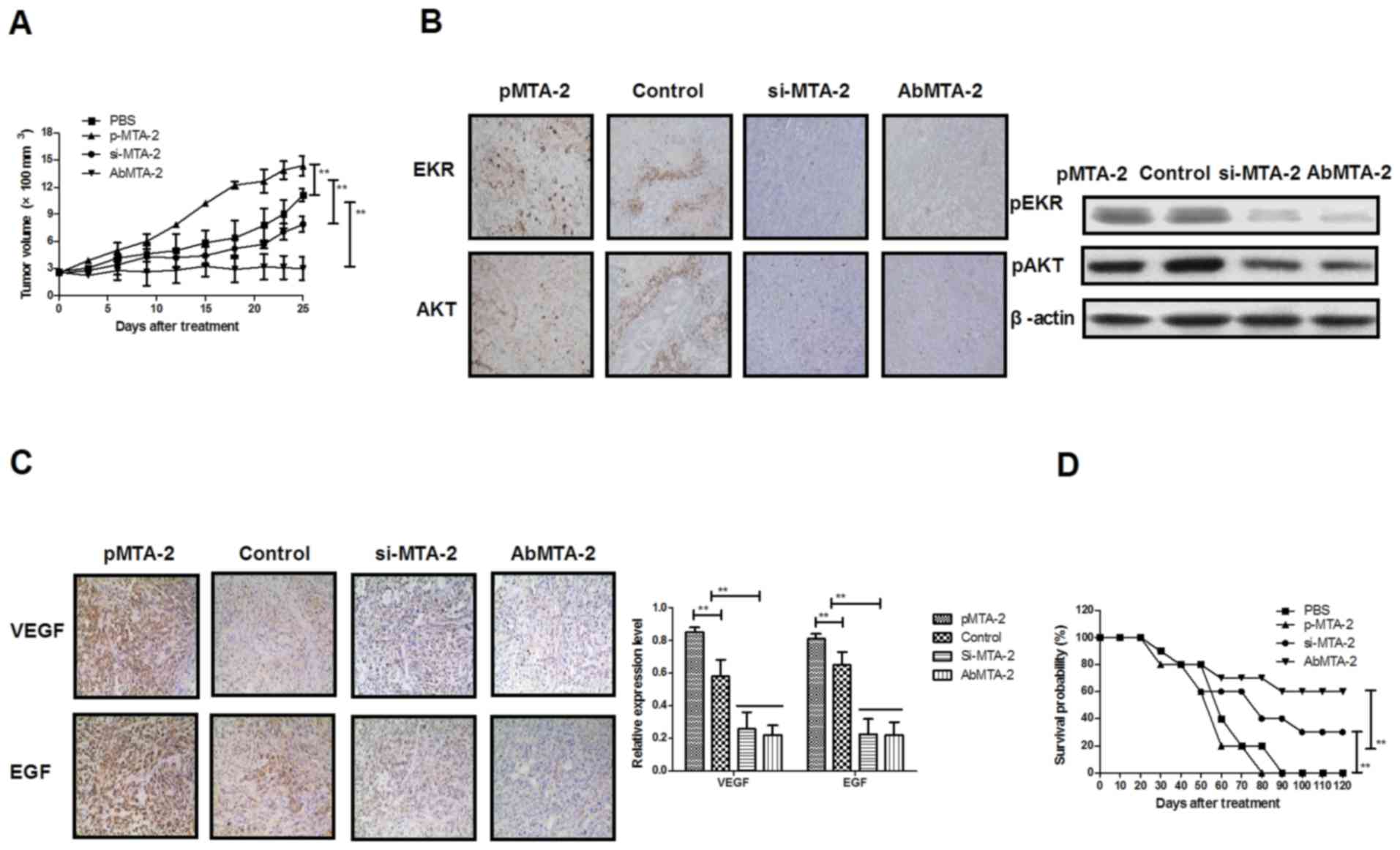
To investigate whether the inhibition of MTA2
protein expression was able to inhibit NSCLC growth in vivo,
NSCLC-bearing mice were established for further analysis. The
antitumor efficacy of AbMTA2 was assessed in the NSCLC mouse model.
The results in Fig. 4A
demonstrated that tumor size was significantly inhibited in
A549-bearing nude mice following AbMTA2 treatment, compared with
mice treated with PBS, si-MTA2 transfection and pMTA2 transfection
(P<0.01), whereas MTA2-overexpressing cells developed a large
tumor mass. We demonstrated that si-MTA2 transfection significantly
inhibited tumor growth compared to pMTA2 transfection and
PBS-treated mice (P<0.01). Immunohistochemistry and western
blotting results of the present study indicated that ERK and AKT
protein expressions were decreased following treatment with AbMTA2
and si-MTA2 transfection compared with pMTA2 transfection and
PBS-treated mice (Fig. 4B). In
addition, it was observed that VEGF protein expression was reduced
in AbMTA2 and si-MTA2 transfection groups compared to pMTA2
transfection and PBS-treated groups (Fig. 4C). Additionally, long-term (120-day
observation) survival analysis demonstrated that AbMTA2 therapy and
si-MTA2 transfection significantly prolonged the survival of
NSCLC-bearing mice compared with pMTA2 transfection and PBS-treated
groups (Fig. 4D; n=12/group). The
results of the present study indicated that treatment with AbMTA2
significantly decreased ERK, AKT and VEGF protein expression
levels, which may translate into long-term survival and tumor-free
living in A549-bearing mice.
Discussion
The occurrence of lung cancer has been associated
with industrial pollution and destruction of the ecological
environment in developed countries (45). Although a number of clinical
treatments have been proposed, these strategies frequently lead to
toxic side effects and are ineffective (46). Molecular-targeted therapies have
the potential to be highly targeted alternatives, with decreased
toxicity to normal human tissues and with the capacity to
completely eradicate the tumor (47–49).
These therapies work by using multi-targeted therapeutic drugs that
are able to directly target antigens on malignant cells. To date, a
number of molecular-targeted therapies have been demonstrated to be
successful in treating human lung cancer (50). A previous study demonstrated that
MTA2 may serve an important role in tumor progression, and may be a
potential target for human cancer therapy (51). In the present study, a targeted
therapy antibody towards MTA2 was constructed for the treatment of
human NSCLC, and its efficacy was investigated in NSCLC cells and
xenograft mice. The results demonstrated that antibody targeting of
MTA2 may be an efficient strategy for the treatment of NSCLC.
NSCLC is the main type of human lung cancer (>80%
of lung cancer cases) and is frequently diagnosed at an advanced
stage. Therefore, the majority of patients with NSCLC exhibit a
limited survival rate post-diagnosis (52). In addition, the relatively higher
rates of morbidity and mortality of NSCLC, compared with other
types of human cancer, have become an issue of increasing
importance (53). A previous study
reported that the number of cases of NSCLC has been increasing in
recent years, which has become a public health focus (54). Therefore, understanding the
pathogenesis of NSCLC and the mechanisms of its progression is
required for the development of treatments for NSCLC.
A number of signaling pathways have been suggested
to be involved in NSCLC progression, and have been targets of
clinical and literature research. It was hypothesized that NSCLC
may become resistant to therapies through feedback mechanisms that
may compensate for targeted inhibition (55). Molecular targets of the VEGF and
ERK/AKT pathways may simultaneously inhibit two important signaling
pathways activated in NSCLC cells and may overcome one potential
aspect of resistance to single-agent therapy. Previous studies have
suggested that treatment options may be available for targeting the
VEGF and ERK/AKT-mediated pathways (56,57).
In the present study, the MTA2-mediated mechanism in NSCLC cells
was analyzed to elucidate the role of signaling pathways following
treatment with AbMTA2. The results indicated that MTA2 may regulate
the growth and aggressiveness of NSCLC via the ERK/AKT and VEGF
signaling pathways. Notably, it was identified that AbMTA2
treatment decreased the apoptotic resistance of NSCLC cells induced
by chemotherapy with Taxol®. Treatment with AbMTA2
inhibited growth through the VEGF signaling pathway and suppressed
migration and invasion via the ERK/AKT signaling pathway.
The present study identified AbMTA2 to be an
efficient anti-NSCLC agent for the inhibition of development and
metastasis, which has been reported to be associated with NSCLC
cell proliferation, migration and invasion (58). The results of the present study
demonstrated that AbMTA2 regulated proliferation, migration and
invasion through the ERK/AKT and VEGF signaling pathways in tumor
cells and xenograft models. Notably, it was additionally observed
that AbMTA2 decreased apoptotic resistance via downregulation of
caspase-3 and Bad expression in NSCLC cells. The results of the
present study suggested that MTA2 may be a potential target in
NSCLC and that AbMTA2 may be able to inhibit NSCLC proliferation
and invasion through inhibition of the ERK/AKT and VEGF signaling
pathways.
References
|
1
|
Magnuson WJ, Yeung JT, Guillod PD,
Gettinger SN, Yu JB and Chiang VL: Impact of deferring radiation
therapy in patients with epidermal growth factor receptor-mutant
non-small cell lung cancer who develop brain metastases. Int J
Radiat Oncol Biol Phys. 95:673–679. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fenton-Ambrose L and Kazerooni EA:
Preventative care: Lung-cancer screens now worth the cost. Nature.
514:352014. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhukovsky M, Varaksin A and Pakholkina O:
Statistical analysis of observational study of the influence of
radon and other risk factors on lung cancer incidence. Radiat Prot
Dosimetry. 160:108–111. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Brody H: Lung cancer. Nature. 513:S12014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Moro-Sibilot D, Smit E, de Castro Carpeño
J, Lesniewski-Kmak K, Aerts JG, Villatoro R, Kraaij K, Nacerddine
K, Dyachkova Y, Smith KT, et al: Non-small cell lung cancer
patients with brain metastases treated with first-line
platinum-doublet chemotherapy: Analysis from the European FRAME
study. Lung Cancer. 90:427–432. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Barnett SA, Downey RJ, Zheng J, Plourde G,
Shen R, Chaft J, Akhurst T, Park BJ and Rusch VW: Utility of
routine pet imaging to predict response and survival after
induction therapy for non-small cell lung cancer. Ann Thorac Surg.
101:1052–1059. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jiang SY, Zhao J, Wang MZ, Huo Z, Zhang J,
Zhong W and Xu Y: Small-cell lung cancer transformation in patients
with pulmonary adenocarcinoma: A case report and review of
literature. Medicine (Baltimore). 95:e27522016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kong R, Feng J, Ma Y, Zhou B, Li S, Zhang
W, Jiang J, Zhang J, Qiao Z, Zhang T, et al: Silencing NACK by
siRNA inhibits tumorigenesis in non-small cell lung cancer via
targeting Notch1 signaling pathway. Oncol Rep. 35:2306–2314. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bablekos GD, Analitis A, Michaelides SA,
Charalabopoulos KA and Tzonou A: Management and postoperative
outcome in primary lung cancer and heart disease co-morbidity: A
systematic review and meta-analysis. Ann Transl Med. 4:2132016.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu Y, Ren Z, Wang J and Zhang S:
Epidermal growth factor receptor-tyrosine kinase inhibitor therapy
is especially beneficial to patients with exon 19 deletion compared
with exon 21 L858R mutation in non-small-cell lung cancer:
Systematic review and meta analysis. Thorac Cancer. 7:406–414.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Abar L, Vieira AR, Aune D, Stevens C,
Vingeliene S, Navarro Rosenblatt DA, Chan D, Greenwood DC and Norat
T: Blood concentrations of carotenoids and retinol and lung cancer
risk: An update of the WCRF-AICR systematic review of published
prospective studies. Cancer Med. 5:2069–2083. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xie FJ, Lu HY, Zheng QQ, Qin J, Gao Y,
Zhang YP, Hu X and Mao WM: The clinical pathological
characteristics and prognosis of FGFR1 gene amplification in
non-small-cell lung cancer: A meta-analysis. Onco Targets Ther.
9:171–181. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lim SH, Sun JM, Lee SH, Ahn JS, Park K and
Ahn MJ: Pembrolizumab for the treatment of non-small cell lung
cancer. Expert Opin Biol Ther. 16:397–406. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kular H, Mudambi L, Lazarus DR, Cornwell
L, Zhu A and Casal RF: Safety and feasibility of prolonged
bronchoscopy involving diagnosis of lung cancer, systematic nodal
staging, and fiducial marker placement in a high-risk population. J
Thorac Dis. 8:1132–1138. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nakagawa K, Asamura H, Tsuta K, Nagai K,
Yamada E, Ishii G, Mitsudomi T, Ito A, Higashiyama M, Tomita Y, et
al: The novel one-step nucleic acid amplification (OSNA) assay for
the diagnosis of lymph node metastasis in patients with non-small
cell lung cancer (NSCLC): Results of a multicenter prospective
study. Lung Cancer. 97:1–7. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Müller B, Bovet M, Yin Y, Stichel D, Malz
M, González-Vallinas M, Middleton A, Ehemann V, Schmitt J, Muley T,
et al: Concomitant expression of far upstream element (FUSE)
binding protein (FBP) interacting repressor (FIR) and its splice
variants induce migration and invasion of non-small cell lung
cancer (NSCLC) cells. J Pathol. 237:390–401. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhao Q, Yue J, Zhang C, Gu X, Chen H and
Xu L: Inactivation of M2 AChR/NF-κB signaling axis reverses
epithelial-mesenchymal transition (EMT) and suppresses migration
and invasion in non-small cell lung cancer (NSCLC). Oncotarget.
6:29335–29346. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang H, Zhu X, Li N, Li D, Sha Z, Zheng X
and Wang H: miR-125a-3p targets MTA1 to suppress NSCLC cell
proliferation, migration, and invasion. Acta Biochim Biophys Sin
(Shanghai). 47:496–503. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Roth MT, Ivey JL, Esserman DA, Crisp G,
Kurz J and Weinberger M: Individualized medication assessment and
planning: Optimizing medication use in older adults in the primary
care setting. Pharmacotherapy. 33:787–797. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ibrahim M, Parry S, Wilkinson D, Bilbe N,
Allen D, Forrest S, Maxwell P, O'Grady A, Starczynski J, Tanier P,
et al: ALK immunohistochemistry in non-small cell lung carcinoma
(NSCLC): Discordant staining can impact patient treatment regimen.
J Thorac Oncol. 11:2241–2247. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cadranel J, Park K, Arrieta O, Pless M,
Bendaly E, Patel D, Sasane M, Nosal A, Swallow E, Galebach P, et
al: Characteristics, treatment patterns, and survival among ALK+
non-small cell lung cancer (NSCLC) patients treated with
crizotinib: A chart review study. Lung Cancer. 98:9–14. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Passaro A, Spitaleri G and de Marinis F:
First-line treatment in NSCLC harboring EGFR common mutations: EGFR
TKI in monotherapy or in combination with anti-VEGF? Expert Rev
Anticancer Ther. 16:799–801. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Izuishi K and Mori H: Recent strategies
for treating stage iv gastric cancer: Roles of palliative
gastrectomy, chemotherapy, and radiotherapy. J Gastrointestin Liver
Dis. 25:87–94. 2016.PubMed/NCBI
|
|
24
|
Hsieh CE, Lin CY, Lee LY, Yang LY, Wang
CC, Wang HM, Chang JT, Fan KH, Liao CT, Yen TC, et al: Adding
concurrent chemotherapy to postoperative radiotherapy improves
locoregional control but not overall survival in patients with
salivary gland adenoid cystic carcinoma-a propensity score matched
study. Radiat Oncol. 11:472016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Khalil EM and Anwar MM: Treatment results
of pediatric nasopharyngeal carcinoma, NCI, Cairo University
experience. J Egypt Natl Canc Inst. 27:119–128. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang Y, Lan G, Si Y, Deng Z, Sun J, Yang
Y, Han X, Weng J and Zhou F: Treatment and outcome of recurrent
cervical lymph nodes in patients with nasopharyngeal carcinoma
after radiotherapy. Zhonghua Er Bi Yan Hou Tou Jing Wai Ke Za Zhi.
51:183–188. 2016.(In Chinese). PubMed/NCBI
|
|
27
|
Casanova M, Özyar E, Patte C, Orbach D,
Ferrari A, Veyrat-Follet C, Errihani H, Pan J, Zhang L, Shen L, et
al: International randomized phase 2 study on the addition of
docetaxel to the combination of cisplatin and 5-fluorouracil in the
induction treatment for nasopharyngeal carcinoma in children and
adolescents. Cancer Chemother Pharmacol. 77:289–298. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Co J, Mejia MB and Dizon JM: Evidence on
effectiveness of intensity-modulated radiotherapy versus
2-dimensional radiotherapy in the treatment of nasopharyngeal
carcinoma: Meta-analysis and a systematic review of the literature.
Head Neck. 38 Suppl 1:E2130–E2142. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Colaco RJ, Betts G, Donne A, Swindell R,
Yap BK, Sykes AJ, Slevin NJ, Homer JJ and Lee LW: Nasopharyngeal
carcinoma: A retrospective review of demographics, treatment and
patient outcome in a single centre. Clin Oncol (R Coll Radiol).
25:171–177. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Setton J, Wolden S, Caria N and Lee N:
Definitive treatment of metastatic nasopharyngeal carcinoma: Report
of 5 cases with review of literature. Head Neck. 34:753–757. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Caponigro F, Longo F, Ionna F and Perri F:
Treatment approaches to nasopharyngeal carcinoma: A review.
Anticancer Drugs. 21:471–477. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Marcy PY, Zhu Y and Bensadoun RJ: Target
volumes in radiotherapy-head and neck tumors intensity-modulated
radiation therapy (IMRT) of nasopharyngeal carcinoma: Practical
aspects in the delineation of target volumes and organs at risk.
Cancer Radiother. 9:240–250. 2005.(In French). View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li Y, Xie G and Pan Z: Advancement
biological target therapy of nasopharyngeal carcinoma. Lin Chung Er
Bi Yan Hou Tou Jing Wai Ke Za Zhi. 29:671–673. 2015.(In Chinese).
PubMed/NCBI
|
|
34
|
Miyashita T, Tajima H, Munemoto M, Shah
FA, Harmon JW, Watanabe T, Shoji M, Okamoto K, Nakanuma S, Sakai S,
et al: Impact of histone deacetylase 1 and metastasis-associated
gene 1 expression in esophageal carcinogenesis. Oncol Lett.
8:758–764. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lee MH, Na H, Na TY, Shin YK, Seong JK and
Lee MO: Epigenetic control of metastasis-associated protein 1 gene
expression by hepatitis B virus X protein during
hepatocarcinogenesis. Oncogenesis. 3:e882014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang Y, Jia H, Lin H, Tan X, Du Z, Chen H,
Xu Y, Han X, Zhang J, Zhao S, et al: Metastasis-associated gene,
mag-1 improves tumour microenvironmental adaptation and potentiates
tumour metastasis. J Cell Mol Med. 16:3037–3051. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Reddy SD, Pakala SB, Molli PR, Sahni N,
Karanam NK, Mudvari P and Kumar R: Metastasis-associated protein
1/histone deacetylase 4-nucleosome remodeling and deacetylase
complex regulates phosphatase and tensin homolog gene expression
and function. J Biol Chem. 287:27843–27850. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Fu J, Qin L, He T, Qin J, Hong J, Wong J,
Liao L and Xu J: The TWIST/Mi2/NuRD protein complex and its
essential role in cancer metastasis. Cell Res. 21:275–289. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gururaj AE, Holm C, Landberg G and Kumar
R: Breast cancer-amplified sequence 3, a target of
metastasis-associated protein 1, contributes to tamoxifen
resistance in premenopausal patients with breast cancer. Cell
Cycle. 5:1407–1410. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Jia W, Zhu J, Martin TA, Sanders AJ, Yang
X, Cheng S, Yu H, Jia G, Liu X, Lu R and Jiang WG: Expression of
metastasis-associated gene-1 is associated with bone invasion and
tumor stage in human pituitary adenomas. Cancer Genomics
Proteomics. 12:113–118. 2015.PubMed/NCBI
|
|
41
|
Zhang B, Zhang H and Shen G:
Metastasis-associated protein 2 (MAP2) promotes the metastasis of
non-small-cell lung cancer through the inhibition of the cell
adhesion molecule Ep-CAM and E-cadherin. Jpn J Clin Oncol.
46:3932016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Shi S, Zhang P, Cheng Q, Wu J, Cui J,
Zheng Y, Bai XY and Chen X: Immunohistochemistry of deparaffinised
sections using antigen retrieval with microwave combined pressure
cooking versus immunofluorescence in the assessment of human renal
biopsies. J Clin Pathol. 66:374–380. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhuang T, Djemil T, Qi P, Magnelli A,
Stephans K, Videtic G and Xia P: Dose calculation differences
between Monte Carlo and pencil beam depend on the tumor locations
and volumes for lung stereotactic body radiation therapy. J Appl
Clin Med Phys. 14:40112013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lee YT, Liu CJ, Hu YW, Teng CJ, Tzeng CH,
Yeh CM, Chen TJ, Lin JK, Lin CC, Lan YT, et al: Incidence of second
primary malignancies following colorectal cancer: A distinct
pattern of occurrence between colon and rectal cancers and
association of co-morbidity with second primary malignancies in a
population-based Cohort of 98,876 patients in Taiwan. Medicine
(Baltimore). 94:e10792015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hui D, Elsayem A, Li Z, De La Cruz M,
Palmer JL and Bruera E: Antineoplastic therapy use in patients with
advanced cancer admitted to an acute palliative care unit at a
comprehensive cancer center: A simultaneous care model. Cancer.
116:2036–2043. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Beck JT: Potential role for mammalian
target of rapamycin inhibitors as first-line therapy in hormone
receptor-positive advanced breast cancer. Onco Targets Ther.
8:3629–3638. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Uhara H: Molecular target therapies for
skin cancers. Nihon Rinsho. 73:1391–1397. 2015.PubMed/NCBI
|
|
49
|
Oda K: Recent process of molecular target
therapies in ovarian serous and endometrioid carcinomas on basis of
intergrated genomic characterization. Gan To Kagaku Ryoho.
42:169–173. 2015.(In Japanese). PubMed/NCBI
|
|
50
|
Chen C, Fei Z, Chen L, Bai P, Lin X and
Pan J: Will weight loss cause significant dosimetric changes of
target volumes and organs at risk in nasopharyngeal carcinoma
treated with intensity-modulated radiation therapy? Med Dosim.
39:34–37. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Liu SL, Han Y, Zhang Y, Xie CY, Wang EH,
Miao Y, Li HY, Xu HT and Dai SD: Expression of
metastasis-associated protein 2 (MAP2) might predict proliferation
in non-small cell lung cancer. Target Oncol. 7:135–143. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Kim DS, Park KM, Won YS, Kim JY, Lee JK,
Kim JG, Oh ST, Jung SS and Kang WK: Occurrence and prognosis of
symptomatic venous thromboembolism in colorectal cancer surgery
patients. Vasc Specialist Int. 30:49–55. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Wink KC, Belderbos JS, Dieleman EM, Rossi
M, Rasch CR, Damhuis RA, Houben RM and Troost EG: Improved
progression free survival for patients with diabetes and locally
advanced non-small cell lung cancer (NSCLC) using metformin during
concurrent chemoradiotherapy. Radiother Oncol. 118:453–459. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Charvat H, Sasazuki S, Inoue M, Iwasaki M,
Sawada N, Shimazu T, Yamaji T and Tsugane S; JPHC Study Group, :
Prediction of the 10-year probability of gastric cancer occurrence
in the Japanese population: The JPHC study cohort II. Int J Cancer.
138:320–331. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Shameem R, Hamid MS, Xu KY and Wu S:
Comparative analysis of the effectiveness of abiraterone before and
after docetaxel in patients with metastatic castration-resistant
prostate cancer. World J Clin Oncol. 6:64–72. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Trinh XB, Tjalma WA, Vermeulen PB, Van den
Eynden G, van der Auwera I, Van Laere SJ, Helleman J, Berns EM,
Dirix LY and van Dam PA: The VEGF pathway and the AKT/mTOR/p70S6K1
signalling pathway in human epithelial ovarian cancer. Br J Cancer.
100:971–978. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhu X, Song Y, Huo R, Zhang J, Sun S, He
Y, Gao H, Zhang M, Sun X, Zhai T, et al: Cyr61 participates in the
pathogenesis of rheumatoid arthritis by promoting proIL-1β
production by fibroblast-like synoviocytes through an AKT-dependent
NF-κB signaling pathway. Clin Immunol. 157:187–197. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Nakamura K, Nozawa K, Aoyagi Y, Ishihara
S, Matsuda K, Fukushima J and Watanabe T: A case report of thyroid
gland metastasis associated with lung metastasis from colon cancer.
Tumori. 97:229–232. 2011. View Article : Google Scholar : PubMed/NCBI
|