Introduction
Endometriosis is a chronic gynecological disorder
defined as the presence of endometrial tissue within extra-uterine
sites (1,2). It affects 10–15% of women of
reproductive age and results in a markedly reduced quality of life
(3–5). The primary symptoms are infertility
and chronic pain (6,7). It is a hormone-dependent and chronic
inflammatory disease (1,2,8),
indicating that the endometrium and the peritoneal environment are
directly associated with its pathogenesis.
The progression of endometriosis is dependent on
genetic, endocrine, immunological and environmental factors
(9,10). The endometrium, serum and
peritoneal fluid of women with endometriosis have abnormal levels
of inflammatory cytokines, angiogenic, growth and adhesion factors,
and cancer-like molecules (6,11,12).
These soluble factors are thought to have predominant involvement
in disease initiation and progression. Interleukin (IL)-1β has a
proliferative effect on endometriotic cells that does not occur in
healthy endometrial cells (13).
IL-1β stimulates the production of IL-6 and IL-8 in endometriotic
cell cultures, which further induce proliferation (14,15)
and reduce the apoptotic rate (16–19).
Additionally, IL-1β increases the shedding of intercellular
adhesion molecule (ICAM)-1 from peritoneal mesothelial cells,
indicating a role in the neovascularization mediated by IL-6 and
vascular endothelial growth factor (VEGF) (20). Furthermore, higher levels of IL-1β
may indicate the conversion of inflammation from an acute to a
chronic form (21).
Tumor necrosis factor (TNF)-α, IL-6, IL-8, IL-10,
VEGF and C-C motif chemokine 2 (MCP-1) expression is increased in
the peritoneal fluid of women with endometriosis (11,18,21–24).
TNF-α is associated with pluripotency mediation and inflammatory
cytokine production, particularly IL-8, in endometriotic tissues
(25). IL-8 increases the adhesion
of endometrial stroma to extracellular matrix proteins, in addition
to increasing metalloproteinase expression and proliferation
(26–28). IL-8 may be a key cytokine in the
progression of endometriotic lesions, by stimulating growth and
indirectly protecting implants against apoptosis (18). IL-6 is secreted by endometrial
cells and may have an important role in the pathology of
endometriosis with interferon-γ. IL-6 increases macrophage
expression of ICAM-1 in patients with endometriosis (29,30).
Increased peritoneal concentrations of IL-6 and IL-8 are associated
with different stages of the disease (23,31).
In addition, the resistance to progesterone observed in
endometriosis (32–34) is linked to the aberrant expression
of cytokines (35,36).
The aberrant peritoneal environment, in addition to
discrepancies in apoptosis and proliferation, result in abnormal
immune cell clearance in women with endometriosis, providing a
longer survival of the endometrial cells regurgitated by uterine
tubes and the establishment of endometrial implants (5,24,31,37).
Recently, the evaluation of peritoneal cytokines in patients with
endometriosis indicated that the inflammatory environment observed
in these patients is triggered by the establishment and growth of
endometrial implants (38).
Therefore, the present study investigated inflammatory cytokine
secretion in the culture media of healthy and endometriotic primary
cells. It was hypothesized that co-culture with endometriotic cells
may lead to inflammatory cytokine secretion and phenotypic
modifications in healthy endometrial cells associated with
endometriosis implant establishment.
Materials and methods
Human tissues and cell culture
The protocol of the present study was approved by
the Federal University of São Paulo ethical committee (registration
no. 1044/11, São Paulo, Brazil). Written informed consent was
obtained from all patients. Endometrial cells (ECs) from healthy
individuals and patients with endometriosis were obtained from the
Endometrium and Endometriosis Cell Bank of the Pelvic Pain and
Endometriosis Unit of the Federal University of São Paulo. The
selected samples were collected from August 2013 to April 2014.
Human endometrium samples were collected from fertile cycling women
aged 18–45 undergoing laparoscopic surgery for endometriosis stage
IV (endometriosis group; n=3) and tubal ligation (control group;
n=3). Patients had not taken exogenous hormones, given birth or
breastfed for 3 months prior to surgery; patients with
co-morbidities including teratoma, endometrial polyps or any other
proliferative disease were excluded. The collected endometrial
tissue was separated into two for either histological analysis
(39,40) (data not shown) or cell culture.
The tissue for cell culture was immediately placed
in Dulbecco's modified Eagle's medium with nutrient mixture F12
(DMEM/F12; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
containing 400 U/ml penicillin and 400 µg/ml streptomycin (Gibco;
Thermo Fisher Scientific, Inc.), stored at 4°C and processed within
2–24 h. The endometrial tissue was dissociated with 255 units of
collagenase type IA (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany)
and 3 units of dispase (Gibco; Thermo Fisher Scientific, Inc.) in
DMEM/F12 and was subsequently incubated for 40 min in a 37°C water
bath under constant agitation. The cell suspension obtained was
centrifuged at 500 × g for 5 min at room temperature and the pellet
was resuspended in 5 ml EC medium containing DMEM/F12 at pH 7.4, 1%
Minimum Essential Medium non-essential amino acids (Gibco; Thermo
Fisher Scientific, Inc.), 0.1 mmol/l 2-mercaptoethanol
(Sigma-Aldrich; Merck KGaA), 100 µ/ml penicillin and 100 µ/ml
streptomycin and 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.), and inoculated in a pre-coated 25 cm2
cell culture flask. The cells were grown in EC medium supplemented
with 10% FBS at 37°C until approximately 70% confluence was
reached. At this stage, cells were subcultured. The cells were
stored in liquid nitrogen.
Healthy and endometriotic cell
co-culture
Co-cultures were performed in 12-well culture
plates. Healthy ECs were pre-stained with 2 µg/ml Calcein-AM
(Thermo Fisher Scientific, Inc., Waltham, MA, USA) for 30 min at
37°C with 5% CO2. Cell number and viability were
assessed with a Countess™ Automated Cell Counter (Invitrogen;
Thermo Fisher Scientific Inc.); 10 µl cell suspension was
thoroughly mixed with 10 µl 0.4% Trypan Blue (Invitrogen; Thermo
Fisher Scientific Inc.), then, 10 µl was added to the chamber
slide, the cells were settled for 30 sec at room temperature prior
to counting; according to the manufacturer's protocols. Healthy and
endometriotic cells were mixed at a 1:1 ratio and seeded onto
12-well plates at a density of 5,000 cells/cm2 and
cultured in EC medium supplemented with 10% FBS for 10 days. The
adhesion of green (healthy) and unstained (endometriotic) cells was
reported by light microscopy (magnification, ×50, Axio Observer,
Zeiss GmbH, Jena, Germany) 24 h after the initial cell seeding, the
stained and non-stained cells were visually counted in four
different fields for each triplicate. The culture medium was
changed every 2 days and in the day prior to each control point
(1st, 3rd, 7th and 10th days of culture). Cells were harvested
during the 1st, 3rd, 7th and 10th days of culture, suspended in FBS
with 10% dimethyl sulfoxide and stored in liquid nitrogen. The
culture medium was collected and stored at −80°C on days 1, 3, 7
and 10 of culture. As a control, healthy and endometriotic cell
groups were cultured separately as described above.
Culture medium cytokines
detection
IL-1β, IL-6, IL-8, IL-10, IL-12p70 and TNF-α
expression levels were detected in the supernatant culture medium
samples of single cell or co-cultured cells at days 1, 3, 7 and 10
by flow cytometry using the phycoerythrin (PE)-conjugated beads
from the Human Inflammatory Cytokine cytometric bead array kit (BD
Biosciences, Franklin Lakes, NJ, USA). Events acquisition was
performed in a CANTO II 6-colour flow cytometer (BD Biosciences)
and data were analyzed with FlowJo software (version 10.0.7; FlowJo
LLC, Ashland, OR, USA).
Flow cytometry of membrane
markers
Healthy and endometriotic cells prior to the
co-culture seeding and co-cultured cells harvested in the 1st, 3rd,
7th and 10th days of culture were thawed on ice, centrifuged at 500
× g for 5 min 4°C, ressuspended in pH 7.4 1X PBS (Gibco; Thermo
Fisher Scientific, Inc.) with 1% FBS (Gibco; Thermo Fisher
Scientific, Inc.) for blocking and incubated with directly
conjugated antibodies for the surface markers aminopeptidase N
[CD13-allophycocyanin (APC); cat no. 555394; BD Biosciences] at 1:5
dilution ratio and cell surface glycoprotein MUC18 [CD146-PE; cat
no. 561013; BD Biosciences] at 1:20 dilution ratio for 1 h at 4°C.
Cells were washed with PBS 1X pH 7.4 and centrifuged at 500 × g for
5 min 4°C, ressuspended in PBS 1X pH 7,4 and subsequently analyzed
with a BC FACSCanto II flow cytometer (BD Biosciences). The
experiment controls were performed using imunoglubulin G1 isotype
controls conjugated with PE (cat no. 550617) and APC (cat no.
550854) both at 1:5 working dilution ratio (BD Biosciences). Data
were analyzed in FlowJo software (version 10.0.7; FlowJo LLC).
Statistical analysis
The data were analyzed with PASW Statistics 18.0.0
(IBM Corp., Armonk, NY, USA). All cell cultures were performed in
experimental triplicates. Data are presented as the mean ± standard
error (SE). Cytokine data were analyzed with repeated measures
analysis followed by Dunnett's T3 pairwise comparisons post-hoc
test between the groups that presented unequal variances. Cell
marker data were analyzed with repeated measures analysis followed
by a Bonferroni corrected pairwise comparisons test within-subjects
factors. The curves obtained were tested for sphericity using
Mauchly's test and within-subject effects were corrected using the
Greenhouse-Geisser test when sphericity assumption was violated.
Simple and multiple linear regression were performed to identify
the correlation between the variables obtained, where 0< r
<0.25 was null; 0.25 < r <0.50 was weakly correlated;
0.50< r <0.75 was moderately correlated; and 0.75 < r
<1 was strongly correlated (41). P<0.05 was considered to indicate
a statistically significant difference.
Results
Competitive relationship between
healthy and endometriotic cells in co-culture systems
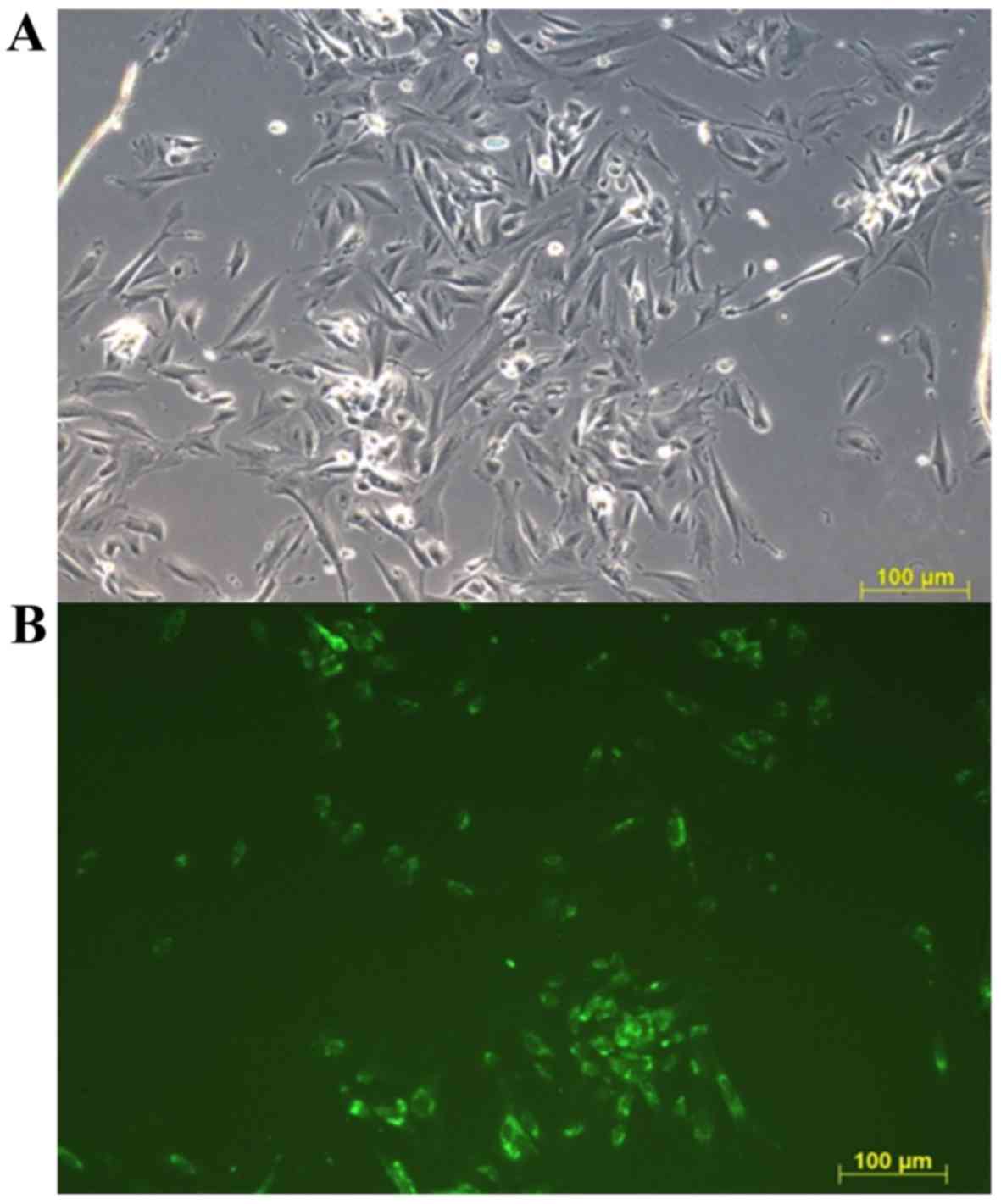
The adhesion of endometriotic (unstained) and
healthy (Calcein-AM stained) cells in the first day of the
co-culture is presented in Fig. 1.
The amount of stained and not stained cells was visually
semiquantitative evaluated in four microscopy fields in each
triplicate. No observable difference was determined in the amount
of endometriotic or healthy cells attached, indicating an equal
number of each type of cell at the beginning of co-cultivation.
Inflammatory cytokine profile of
healthy, endometriotic and co-cultured cells
The levels of inflammatory cytokines (IL-1β, IL-6,
IL-8, IL-10, IL-12p70 and TNF-α) secreted into the culture medium
were determined over time in endometriotic and healthy cells
cultured alone, in addition to co-cultured cells. Insignificant
levels of IL-10, IL12p70 and TNF-α were detected in the culture
medium of all tested combinations of cells (data not shown).
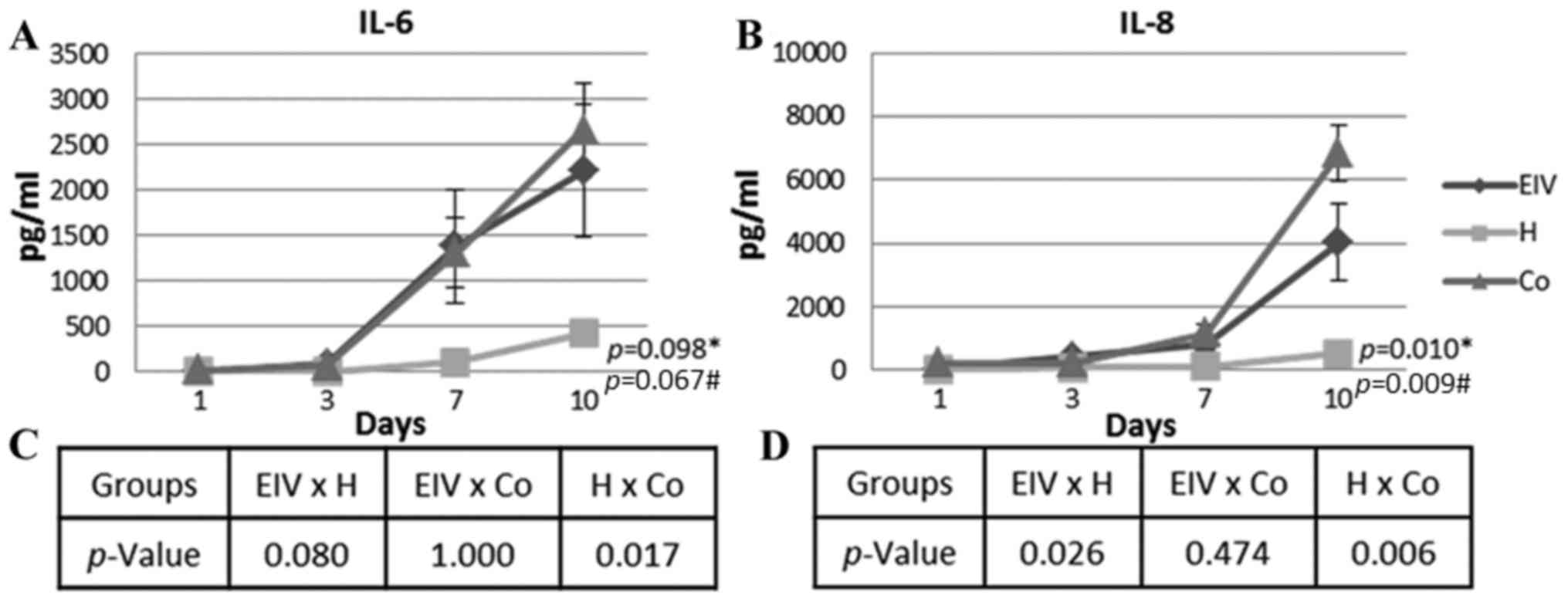
However, the IL-6 concentration increased over time in all groups
(Fig. 2A). The variation observed
for the endometriotic cells was similar to that observed in the
co-cultured ECs (P=1.000, across all time points). The healthy ECs
had a significantly different profile of IL-6 secretion, compared
with co-cultured ECs (P=0.017, across all time points). As
presented in Fig. 2, IL-6
secretion was higher in the endometriosis and co-culture groups. A
strong correlation was detected between the number of cells and the
concentration of IL-6 in the medium of healthy and endometriotic
cells (Table I).
 | Table I.Multiple correlation coefficient
between inflammatory cytokine expression and cell number in
endometriotic and healthy endometrial cells individually and
co-cultured over time. |
Table I.
Multiple correlation coefficient
between inflammatory cytokine expression and cell number in
endometriotic and healthy endometrial cells individually and
co-cultured over time.
|
| Endometriosis | Healthy | Co-cultured
cells |
|---|
|
|
|
|
|
|---|
| Pairs | r | P-value | r | P-value | r | P-value |
|---|
| No. of cells and
IL-6 | 0.87114 | 0 | 0.92528 | 0 |
|
|
| No. of cells and
IL-8 | 0.60255 | 0.0001 | 0.53407 | 0.0087 |
|
|
| IL-6 and IL-8 (all
time points) | 0.84211 | 0 | 0.50648 | 0.0137 | 0.6947 | 0 |
| IL-1β and IL-6 (day
10) |
|
|
|
| 0.5565 | 0.1197 |
| IL-1β and IL-8 (day
10) |
|
|
|
| 0.2158 | 0.5771 |
Pro-angiogenic IL-8 was differentially secreted in
healthy, endometriotic and co-cultured ECs (Fig. 2B). The most significant variation
occurred from day 7, when the concentration increased in all
groups. The differences between the curves of IL-8 concentration
were statistically significant (P=0.010). The secreted IL-8 level
in the healthy EC culture was significantly lower compared with the
endometriosis (P=0.026, across all time points) and co-cultured
(P=0.006, across all time points) groups. Similar to the trend
observed for IL-6, the secretion of IL-8 was similar in the
endometriotic and co-cultured cells (P=0.474, across all time
points). The number of cells and IL-8 concentration were moderately
correlated in healthy and endometriotic cells (Table I). Additionally, there was a
moderate correlation between IL-6 and IL-8 secretion in the healthy
and co-cultured cells. A strong correlation between the secretion
of these cytokines was observed in the endometriotic cells
(Table I).
The co-cultured cells had 48.30 pg/ml (SE, 9.68)
IL-1β at day 10 of culture (data not shown); IL-1β levels at days
1, 3 and 7 were undetectable. The secretion of IL-1β appears to be
associated with the effects of long-term co-culturing of the cells,
although not directly to the secretion of IL-6 or IL-8 (Table I).
Population profile of co-cultured
ECs
Flow cytometry analysis of the cells cultured alone
and co-cultured cells using the size (forward scatter) and
complexity (side scatter) parameters demonstrated the presence of
two main populations in the samples: One and two. Population one
represented <5% of the total events collected in the healthy
(Fig. 3A) and endometriosis
(Fig. 3B) samples alone (2.95%;
SE, 0.44). The percentage of this population increased by 15-fold
in the first day of co-culture (Day 1, 45.93%; SE, 3.58; P=0.000179
vs. cells cultured alone; Fig.
3C). No population differences were observed in the ECs
individually cultured over time (data not shown).
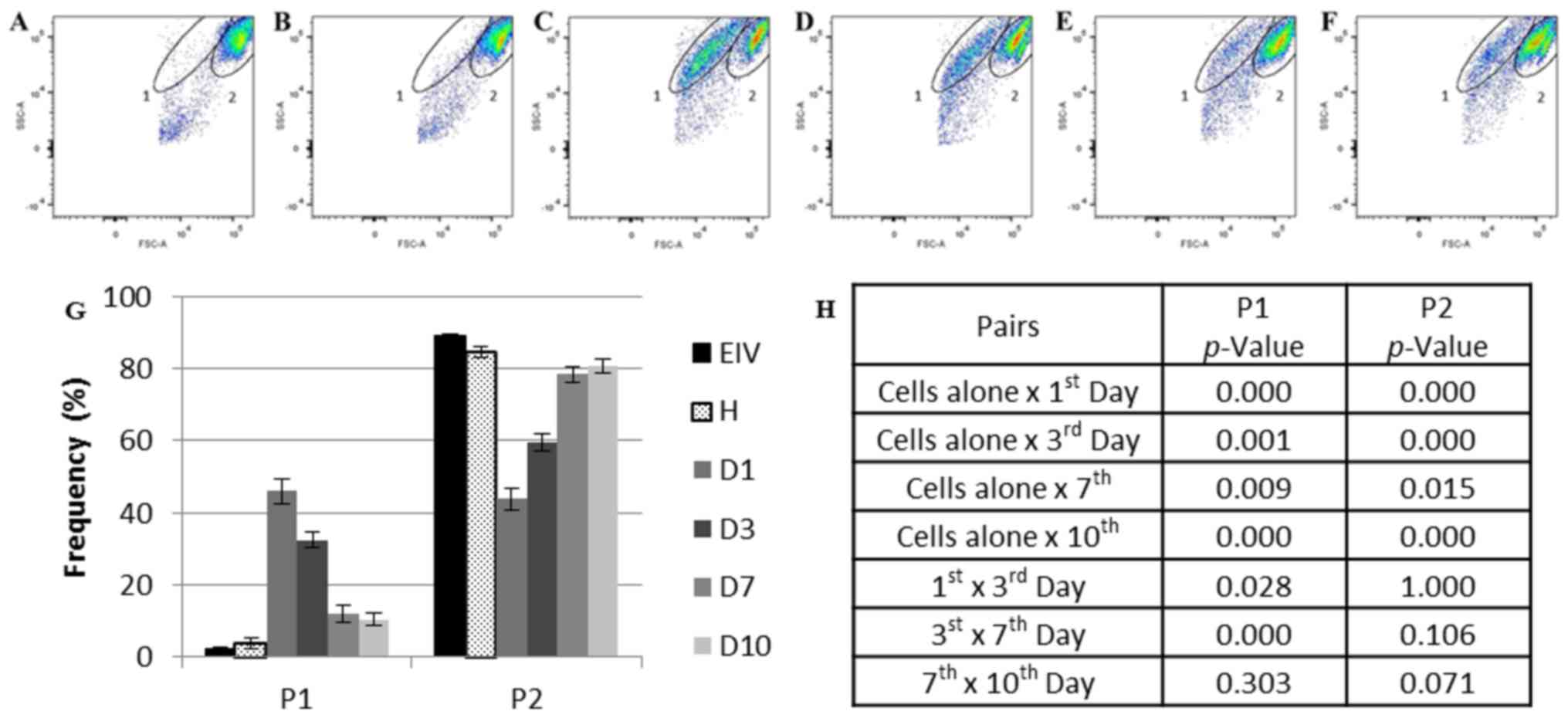 | Figure 3.Flow cytometry analysis of the
healthy, endometriotic and co-cultured ECs. (A) Endometriotic ECs
cultivated alone prior to co-culture and (B) healthy ECs cultured
alone prior to co-culture. (C) Co-cultured ECs at day 1, (D) day 3,
(E) day 7 and (F) day 10. (G) Frequency of P1 and P2 during
co-culture. (H) Statistical analysis of each population frequency
between each day of co-culture and cells cultured alone. The
elliptical delimitations 1 and 2 indicate the two populations
delimited by differences in size and complexity in each sample. The
images are representative for all samples. Data are presented as
the mean of each sample group ± standard error. ECs, endometrial
cells; EIV, endometriotic ECs; D1, day 1 of co-culture; D3, day 3
of co-culture, D7, day 7 of co-culture; D10, day 10 of co-culture;
FSC, forward scatter; H, healthy ECs; P1, population one; P2,
population two; SSC, side scatter. |
Population one diminished at day 3 (32.35%; SE,
2.16; P=0.028412 vs. day 1 of co-culture; Fig. 3D) and day 7 (11.89%; SE, 2.33;
P=0.000076 vs. day 3 of co-culture; Fig. 3E) of co-culture and appeared to
stabilize between days 7 and 10 (Fig.
3F) of culture (10.40%; SE, 1.79; P=0.302732, day 7 vs. day 10
of co-culture).
Population two was more prevalent in the EC samples
cultured alone (87.36%; SE, 1.52; Fig.
3A and B). This percentage reduced by one-half on day 1 of
co-culture (43.78%; SE, 3.03; P=0.000264 vs. cells cultured alone;
Fig. 3C). Despite this population
size increasing over time (day 3, 59.50%; SE, 2.41; P=0.000449 vs.
cells cultured alone; day 7, 78.36%; SE, 2.19; P=0.015462 vs. cells
cultured alone; Fig. 3D and E,
respectively), the original population size detected in ECs
cultured alone was not completely reestablished at day 10 of
co-culture (80.73%; SE, 2.06; P=0.000622 vs. cells cultured alone;
Fig. 3F).
Phenotypic profile of co-cultured ECs
over time
According to the variations detected within the
populations of ECs cultured alone or co-cultured, the stromal
endometrial cell marker CD13 (42,43)
and the endometrial mesenchymal stem cell (eMSC) marker CD146
(44–46) were selected to further characterize
these populations. CD13 expression was not significantly different
within co-cultured or individually cultured populations (Fig. 4).
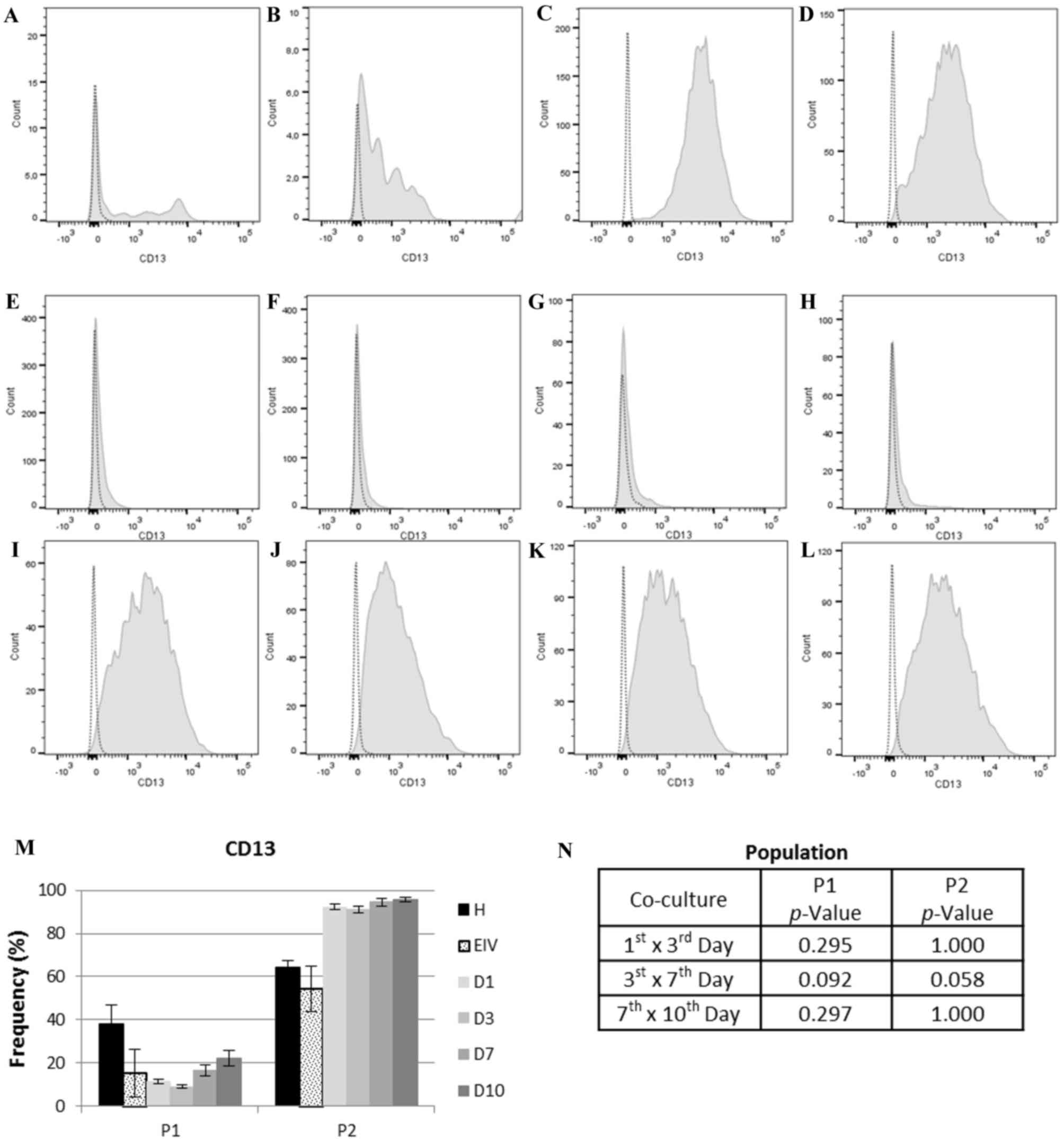 | Figure 4.CD13 expression in each cell
population. In the histograms, light gray dotted lines indicate
background fluorescence obtained with the isotype control
immunoglobulin G1. CD13 expression is represented by the light gray
filled areas. The × axis represents fluorescence intensity and the
y axis represents cell count. (A) Histogram for CD13 expression in
P1 of healthy ECs and (B) EIV. (C) Histogram for CD13 expression in
P2 of healthy ECs and (D) EIV. (E) Histogram for CD13 expression on
P1 at day 1, (F) day 3, (G) day 7 and (H) day 10 of EC co-culture.
(I) Histogram for CD13 expression on P2 at day 1, (J) day 3, (K)
day 7 and (L) day 10 of EC co-culture. (M) CD13 expression in each
population. (N) Statistical analysis of CD13 expression between
each day of co-culture for P1 and P2. Data are presented as the
mean of each sample group ± standard error. ECs, endometrial cells;
CD13, aminopeptidase N; P1, population one; P2, population two; H,
healthy ECs; EIV, endometriotic ECs; D1, day 1 of co-culture; D3,
day 3 of co-culture; D7, day 7 of co-culture; D10, day 10 of
co-culture. |
The expression of eMSC marker CD146 varied
significantly over time (Fig. 5).
Population one had negligible expression of CD146, with the mean
expression at all time points in the co-cultured group of 1.21%
(SE, 0.25). In population two, an increase in CD146 expression was
observed between days 1 and 7 of co-culture (1st vs. 3rd days,
P=0.037; 3rd vs. 7th days, P=0.017), with a slight decrease in
expression between days 7 and 10 of co-culture (P=0.071). The
expression of CD146 in population 2 at 1st day of co-culture was
more similar to endometriotic cells (P=0.102010) compared with
healthy ECs (P=0.051579) (data not shown). These results
demonstrated that population two was predominantly composed of
eMSCs, and that there was an increase in these cells when healthy
and endometriotic cells were co-cultured.
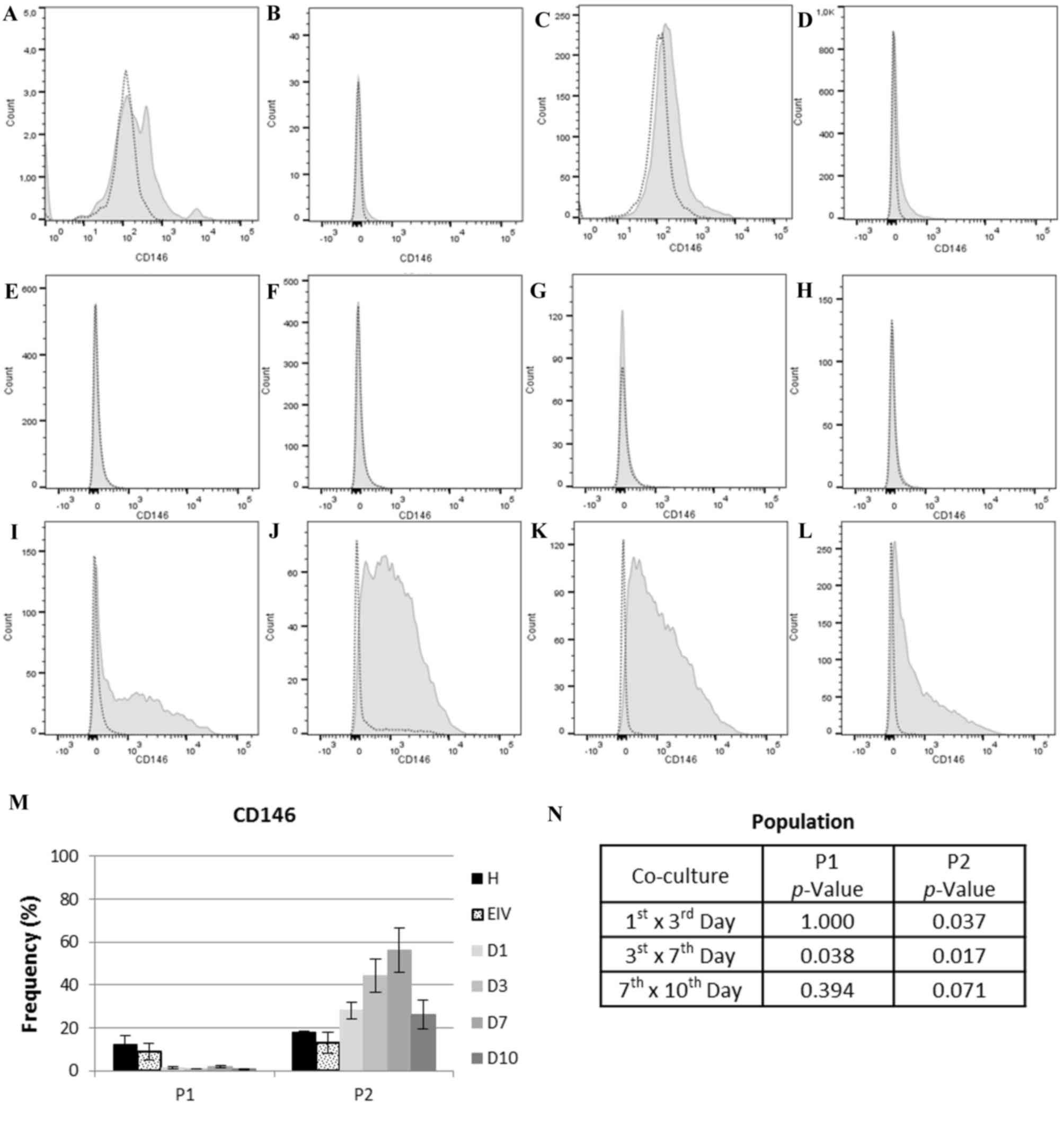 | Figure 5.CD146 expression in cell each
population. In the histograms, light gray dotted lines indicate
background fluorescence obtained with the isotype control
immunoglobulin G1. CD146 expression is represented by the light
gray filled areas. The × axis represents fluorescence intensity and
the y axis represents cell count. (A) Histogram for CD146
expression in P1 of healthy ECs and (B) EIV. (C) Histogram for
CD146 expression in P2 of healthy ECs and (D) EIV. (E) Histogram
for CD146 expression on P1 at day 1, (F) day 3, (G) day 7 and (H)
day 10 of EC co-culture. (I) Histogram for CD146 expression on P2
at day 1, (J) day 3, (K) day 7 and (L) day 10 of EC co-culture. (M)
CD146 expression in each population. (N) Statistical analysis of
CD146 expression between each day of co-culture for P1 and P2. Data
are presented as the mean of each sample group ± standard error.
ECs, endometrial cells; CD146, cell surface glycoprotein MUC18; P1,
population one; P2, population two; H, healthy; ECs, EIV,
endometriosis ECs; D1, day 1 of co-culture; D3, day 3 of
co-culture; D7, day 7 of co-culture; D10, day 10 of co-culture. |
Discussion
The first and most acceptable theory for
endometriosis pathogenesis was proposed almost a century ago
(47), yet the pathophysiology of
this disease remains unclear. As a hormone-dependent and chronic
inflammatory disease (1,2,8),
cytokines and soluble factors appear to serve an important role in
its pathophysiology. The inflammatory content of the peritoneal
fluid and serum, in addition to the eutopic and ectopic endometrium
has been extensively studied (6,11,12,6). However, there are a number of
questions which remain unanswered.
The increased levels of inflammatory cytokines in
the peritoneal fluid, serum and endometrium of women with end
ometriosis has been reported in numerous studies (18,23,28,18). The principal identified factors
include TNF-α, IL-6, IL-8, IL-10, VEGF and MCP-1 (11,18,21–24,49–51).
In the present study, increased levels of IL-6 and IL-8 were
observed in endometriotic cells cultured alone and in co-culture
with healthy ECs, compared with individually cultured healthy ECs.
The levels of IL-1β were additionally increased in the co-culture
at day 10.
Sikora et al (18) proposed that IL-8 is important for
progression of endometriosis. IL-8 increases the adhesion of
endometrial stroma to extracellular matrix proteins, in addition to
metalloproteinase expression and cell proliferation (26–28).
The surgical excision of endometrial lesions leads to a decrease in
the peritoneal levels of IL-8 (38). In the present study, increased
secretion of IL-8 by endometriotic and co-cultured cells was
detected. A previous study indicated that peritoneal immune cells,
particularly macrophages, are the main source of IL-8 (18). The results of the present study
indicated that ECs additionally contribute to IL-8 secretion.
Furthermore, IL-8 expression was moderately correlated with cell
number and IL-6 expression, suggesting that IL-8 secretion may be
independent of cell proliferation and IL-6 levels.
IL-6 is secreted by the endometrium and implants,
and is involved in diverse aspects of reproductive physiology,
including ovarian steroid production, foliculogenesis and early
embryonic implantation (28,49,52,53).
IL-6 is associated with the increased expression of ICAM-1 by
macrophages in patients with endometriosis (29,30).
The present study demonstrated that the secretion of IL-6 by
endometriotic cells was higher compared with healthy or co-cultured
ECs. The increased levels of IL-6 were strongly correlated with
cell numbers, indicating that the increased proliferative rate may
be due to IL-6 signaling. The increased cell number also increases
the IL-6 secretion. It has been well established that endometriotic
stromal cells have a higher proliferative rate compared with
healthy endometrial stromal cells (54,55).
This increase in proliferation rate may be mediated by IL-6.
The levels of IL-6 may be associated with IL-1β
secretion (20). The results of
the present study revealed the presence of IL-1β at day 10 of
co-culture only, with no significant alterations in the expression
of IL-1β in individually cultured healthy or endometriotic cells.
Furthermore, there was no statistically significant correlation
between IL-6 and IL-1β expression. The secretion of IL-1β by the
primary co-cultured ECs may be associated with long-term
co-culture, which may indicate a switch from an acute to chronic
inflammatory response in the endometrial cells (21). The proliferative effects of IL-1β
have been reported in endometriotic cells (13). Furthermore, an increase in IL-1B
expression occurs in the co-culture of endometrial primary cells
and MSCs (56).
The endometrium contains MSCs that are involved in
the cyclical regeneration of this tissue (57–59).
The role of stem cells in the pathogenesis of endometriosis has
been reported (37,60,61).
The most widely accepted theory for endometriosis pathogenesis was
proposed by Sampson (47) and
indicates that the implants establish from cells undergoing
retrograde menstruation; this theory alone is unable to explain all
clinical presentations of the disease. The combined theories of
Sampson (47) and the involvement
of stem cells provide a better explanation for endometriosis
physiopathology. The dysregulation of eMSCs has been proposed as a
key mechanism of endometriosis, in concordance with the retrograde
menstruation theory (62).
The mesenchymal stem cell marker CD146 (46) was used in the present study in
order to identify the presence of MSCs in the pool of primary
cells, and to report the variations in the MSC population during
co-culture. An increase in the number of eMSCs detected by CD146
expression was observed in the co-culture system. These alterations
in CD146 expression over time, together with the alterations in
cytokine secretion, may indicate the involvement of MSCs in
cytokine secretion. A pool of primary cells at a low passage number
were selected, as this contained various cell populations and was
more similar to the composition of the eutopic endometrium. In
previous work (56), the
co-culture of umbilical cord blood MSCs with primary endometrial
cells resulted in increased IL-1β expression. Furthermore,
co-culture eliminated the effect of p27 gene therapy on
endometriotic stromal cells. These observations, together with the
identification and characterization of endometrial MSCs (37,45,58,37), have provided an insight into the
involvement of MSCs in endometriosis pathogenesis. In the present
study, the co-cultivation of a pool of healthy and diseased primary
cells, containing endometrial MSCs, led to the secretion of
cytokines that may be directly involved in the pathogenesis of
endometriosis. This supports the theory that eMSCs have an
important role in the secretion of cytokines and endometriosis.
MSCs express CD13 (44,46),
which is additionally expressed by stromal endometrial cells
(42,43). CD13 is an N-aminopeptidase involved
in the inactivation of IL-8 (64).
Conflictingly, increased levels of CD13 expression due to
cell-to-cell contact may be associated with IL-8 inactivation
resistance (65) and the
inhibition of apoptosis mediated by IL-8 (66). In the present study, CD13
expression was not significantly different between the co-cultured
and individually cultured cells, indicating no physiological
implications. Therefore, CD13 expression in the present study may
be indicative of fibroblastoid cells.
Furthermore, the present study observed that a mixed
cell population derived from the endometrium exhibited a secretory
profile similar to that of endometriotic cells, even in the
presence of healthy ECs. The results obtained demonstrated that the
communication between endometriotic and healthy cells results in a
secretory profile similar to what is reported in the peritoneal
fluid of women with endometriosis (24,28,31,52).
The increased secretion of IL-6 and IL-8 is additionally associated
with the progesterone resistance observed in endometriosis
(32,34). The results of the present study
support this finding, as the endometriotic cells secreted higher
levels of IL-6 and IL-8 compared with the healthy cells.
Additionally, the findings of the present study were consistent
with the retrograde menstruation theory and clarified certain
aspects of the disrupted peritoneal environment of women with
endometriosis. For example, despite the presence of healthy
endometrial cells in retrograde menstruation, the presence of
endometriotic cells may be decisive in the secretory profile of the
cell pool (18,21,37).
The endometrial cells regurgitated into peritoneal cavity may be
responsible for the induction of the aberrant cytokine profile
observed in the peritoneal fluid and eutopic endometrium of women
with endometriosis (18,19,21,18). The variations observed in the
CD146+ population support the putative role of eMSCs in
the pathogenesis of endometriosis.
The secretion alterations observed in the
individually cultured ECs and the co-culture indicate a critical
role for ectopic endometrial cells in the initiation of peritoneal
environment disruption, which is in accordance with a recently
published report (38). Thus, the
cytokine profile observed may not only be generated by molecules
secreted from immune cells. The main limitation of the present
study is that, as an in vitro study, the effects of
EC-secreted cytokines on immune cells present in the peritoneum of
women with endometriosis was not demonstrated. Studies which
isolate each population of cells observed in the present study may
clarify the cell type responsible for the secretion of the
cytokines or reveal the association between healthy and disease
cell contact.
In accordance to previous research (18,19,21,18), the findings of the present study
suggest that the soluble factors secreted by endometriotic cells
may have an important role in the disruption of the cell cycle and
in the establishment of the peritoneal environment of women with
endometriosis. The endometriotic cell pool may be responsible for
the establishment of an inflammatory peritoneal environment
favorable to the initiation and progression of endometrial cell
adhesion and clustering.
Acknowledgements
We thank Colsan Blood Bank (São Paulo, Brazil) for
all the technical support.
Funding
The present study was supported by the São Paulo
Research Foundation (grant no. 2011/14683-7) and the Brazilian
National Council of Technological and Scientific Development (grant
no. 480303/2013-4).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ALI, GAG, MJBCG and ES provided major contributions
to the conception and design of the present study. ALI, RMP, AK and
GK conducted the collection of samples, experimentation and
acquisition of data. ALI and ES were involved in analysis and
interpretation of data. All authors were involved in manuscript
drafting and critical discussion, as well as in the final approval
of the version to be published.
Ethics approval and consent to
participate
The protocol of the present study was approved by
the Federal University of São Paulo ethical committee (São Paulo,
Brazil). Written informed consent was obtained from all
patients.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ulukus M, Cakmak H and Arici A: The Role
of Endometrium in Endometriosis. J Soc Gynecol Investig.
13:467–476. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Falconer H, D'Hooghe T and Fried G:
Endometriosis and genetic polymorphisms. Obstet Gynecol Surv.
62:616–628. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Eskenazi B and Warner ML: Epidemiology of
endometriosis. Obstet Gynecol Clin North Am. 24:235–258. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Vinatier D, Cosson M and Dufour P: Is
endometriosis an endometrial disease? Eur J Obstet Gynecol Reprod
Biol. 91:113–125. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bulun SE: Endometriosis. N Engl J Med.
360:268–279. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Burney RO and Giudice LC: Pathogenesis and
pathophysiology of endometriosis. Fertil Steril. 98:511–519. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Giudice LC and Kao LC: Endometriosis.
Lancet. 364:1789–1799. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Berkley KJ, Rapkin AJ and Papka RE: The
Pains of Endometriosis. Science. 308:1587–1589. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
de Marqui Trovó AB: Genetic polymorphisms
and endometriosis: Contribution of genes that regulate vascular
function and tissue remodeling. Rev Assoc Med Bras. 58:620–32.
2012.(In English, Portuguese). PubMed/NCBI
|
|
10
|
Kobayashi H, Yamada Y, Morioka S, Niiro E,
Shigemitsu A and Ito F: Mechanism of pain generation for
endometriosis-associated pelvic pain. Arch Gynecol Obstet.
289:13–21. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ilie I and Ilie R: Cytokines and
endometriosis-the role of immunological alterations. Biotechnol Mol
Biol Nanomedicine. 1:8–19. 2013.
|
|
12
|
Gazvani R and Templeton A: Peritoneal
environment, cytokines and angiogenesis in the pathophysiology of
endometriosis. Reproduction. 123:217–26. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lebovic DI, Baldocchi RA, Mueller MD and
Taylor RN: Altered expression of a cell-cycle suppressor gene,
Tob-1, in endometriotic cells by cDNA array analyses. Fertil
Steril. 78:849–854. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bersinger NA, Günthert AR, McKinnon B,
Johann S and Mueller MD: Dose-response effect of interleukin
(IL)-1β, tumour necrosis factor (TNF)-α, and interferon-γ on the in
vitro production of epithelial neutrophil activating peptide-78
(ENA-78), IL-8, and IL-6 by human endometrial stromal cells. Arch
Gynecol Obstet. 283:1291–1296. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yoshino O, Izumi G, Shi J, Osuga Y, Hirota
Y, Hirata T, Harada M, Nishii O, Koga K and Taketani Y: Activin-A
is induced by interleukin-1β and tumor necrosis factor-α and
enhances the mRNA expression of interleukin-6 and
protease-activated receptor-2 and proliferation of stromal cells
from endometrioma. Fertil Steril. 96:118–121. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bilotas M, Meresman G, Buquet R, Sueldo C
and Barañao RI: Effect of vascular endothelial growth factor and
interleukin-1beta on apoptosis in endometrial cell cultures from
patients with endometriosis and controls. J Reprod Immunol.
84:193–198. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Vetvicka V, Laganà AS, Salmeri FM, Triolo
O, Palmara VI, Vitale SG, Sofo V and Králíčková M: Regulation of
apoptotic pathways during endometriosis: From the molecular basis
to the future perspectives. Arch Gynecol Obstet. 294:897–904. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sikora J, Smycz-Kubańska M,
Mielczarek-Palacz A and Kondera-Anasz Z: Abnormal peritoneal
regulation of chemokine activation-The role of IL-8 in pathogenesis
of endometriosis. Am J Reprod Immunol. 77:2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gonçalves GA, Camargo-Kosugi CM, Bonetti
TC, Invitti AL, Girão MJ, Silva ID and Schor E: p27kip1
overexpression regulates VEGF expression, cell proliferation and
apoptosis in cell culture from eutopic endometrium of women with
endometriosis. Apoptosis. 20:327–335. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kyama CM, Overbergh L, Mihalyi A, Meuleman
C, Mwenda JM, Mathieu C and D'Hooghe TM: Endometrial and peritoneal
expression of aromatase, cytokines, and adhesion factors in women
with endometriosis. Fertil Steril. 89:301–310. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sikora J, Mielczarek-Palacz A and
Kondera-Anasz Z: Association of the precursor of interleukin-1β and
peritoneal inflammation-role in pathogenesis of endometriosis. J
Clin Lab Anal. 30:831–837. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
May CD, Sphyris N, Evans KW, Werden SJ,
Guo W and Mani SA: Epithelial-mesenchymal transition and cancer
stem cells: A dangerously dynamic duo in breast cancer progression.
Breast Cancer Res. 13:2022011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fan YY, Chen HY, Chen W, Liu YN, Fu Y and
Wang LN: Expression of inflammatory cytokines in serum and
peritoneal fluid from patients with different stages of
endometriosis. Gynecol Endocrinol. 1–6. 2018.(Epub ahead of
Print).
|
|
24
|
Jørgensen H, Hill AS, Beste MT, Kumar MP,
Chiswick E, Fedorcsak P, Isaacson KB, Lauffenburger DA, Griffith LG
and Qvigstad E: Peritoneal fluid cytokines related to endometriosis
in patients evaluated for infertility. Fertil Steril.
107:1191–1199.e2. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sakamoto Y, Harada T, Horie S, Iba Y,
Taniguchi F, Yoshida S, Iwabe T and Terakawa N: Tumor necrosis
factor-alpha-induced interleukin-8 (IL-8) expression in
endometriotic stromal cells, probably through nuclear factor-kappa
B activation: Gonadotropin-releasing hormone agonist treatment
reduced IL-8 expression. J Clin Endocrinol Metab. 88:730–735. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Garcia-Velasco JA and Arici A:
Interleukin-8 stimulates the adhesion of endometrial stromal cells
to fibronectin. Fertil Steril. 72:336–340. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Arici A, Seli E, Zeyneloglu HB, Senturk
LM, Oral E and Olive DL: Interleukin-8 induces proliferation of
endometrial stromal cells: A potential autocrine growth factor. J
Clin Endocrinol Metab. 83:1201–1205. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kalu E, Sumar N, Giannopoulos T, Patel P,
Croucher C, Sherriff E and Bansal A: Cytokine profiles in serum and
peritoneal fluid from infertile women with and without
endometriosis. J Obstet Gynaecol Res. 33:490–495. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Herington JL, Bruner-Tran KL, Lucas JA and
Osteen KG: Immune interactions in endometriosis. Expert Rev Clin
Immunol. 7:611–626. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Young VJ, Brown JK, Saunders PT and Horne
AW: The role of the peritoneum in the pathogenesis of
endometriosis. Hum Reprod Update. 19:558–569. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Barcz E, Milewski Ł, Dziunycz P, Kamiński
P, Płoski R and Malejczyk J: Peritoneal cytokines and adhesion
formation in endometriosis: An inverse association with vascular
endothelial growth factor concentration. Fertil Steril.
97:1380–1386.e1. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Young SL and Lessey BA: Progesterone
function in human endometrium: Clinical perspectives. Semin Reprod
Med. 28:5–16. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Barragan F, Irwin JC, Balayan S, Erikson
DW, Chen JC, Houshdaran S, Piltonen TT, Spitzer TL, George A,
Rabban JT, et al: Human endometrial fibroblasts derived from
mesenchymal progenitors inherit progesterone resistance and acquire
an inflammatory phenotype in the endometrial niche in
endometriosis. Biol Reprod. 94:1182016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lessey BA and Young SL: Homeostasis
imbalance in the endometrium of women with implantation defects:
The role of estrogen and progesterone. Semin Reprod Med.
32:365–375. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wing LY, Chuang PC, Wu MH, Chen HM and
Tsai SJ: Expression and mitogenic effect of fibroblast growth
factor-9 in human endometriotic implant is regulated by aberrant
production of estrogen. J Clin Endocrinol Metab. 88:5547–5554.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wu MH, Lu CW, Chuang PC and Tsai SJ:
Prostaglandin E2: The master of endometriosis? Exp Biol Med
(Maywood). 235:668–677. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gargett CE, Schwab KE, Brosens JJ,
Puttemans P, Benagiano G and Brosens I: Potential role of
endometrial stem/progenitor cells in the pathogenesis of
early-onset endometriosis. Mol Hum Reprod. 20:591–598. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Monsanto SP, Edwards AK, Zhou J,
Nagarkatti P, Nagarkatti M, Young SL, Lessey BA and Tayade C:
Surgical removal of endometriotic lesions alters local and systemic
proinflammatory cytokines in endometriosis patients. Fertil Steril.
105:968–977.e5. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Moen MH and Halvorsen TB: Histologic
confirmation of endometriosis in different peritoneal lesions. Acta
Obstet Gynecol Scand. 71:337–342. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Revised American Society for Reproductive
Medicine classification of endometriosis: 1996. Fertil Steril.
67:817–821. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Vieira S: Introdução à Bioestatística. 4th
edition. Elsevier Ltd; São Paulo: 2008
|
|
42
|
Yoshimoto Kato M, Kato K, Adachi S,
Yamayoshi A, Arima T, Asanoma K, Kyo S, Nakahata T and Wake NK:
Characterisation of side population cells in human normal
endometrium. Hum Reprod. 22:1212–1223. 2007.
|
|
43
|
Seli E, Senturk LM, Bahtiyar OM, Kayisli
UA and Arici A: Expression of aminopeptidase N in human endometrium
and regulation of its activity by estrogen. Fertil Steril.
75:1172–1176. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Schwab KE and Gargett CE: Co-expression of
two perivascular cell markers isolates mesenchymal stem-like cells
from human endometrium. Hum Reprod. 22:2903–2911. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Schwab KE, Hutchinson P and Gargett CE:
Identification of surface markers for prospective isolation of
human endometrial stromal colony-forming cells. Hum Reprod.
23:934–943. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Gargett CE, Schwab KE, Zillwood RM, Nguyen
HP and Wu D: Isolation and culture of epithelial progenitors and
mesenchymal stem cells from human endometrium. Biol Reprod.
80:1136–1145. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Sampson JA: Peritoneal endometriosis due
to the menstrual dissemination of endometrial tissue into the
peritoneal cavity. Am J Obstet Gynecol. 14:422–469. 1927.
View Article : Google Scholar
|
|
48
|
Milewski Ł, Dziunycz P, Barcz E, Radomski
D, Roszkowski PI, Korczak-Kowalska G, Kamiński P and Malejczyk J:
Increased levels of human neutrophil peptides 1, 2, and 3 in
peritoneal fluid of patients with endometriosis: Association with
neutrophils, T cells and IL-8. J Reprod Immunol. 91:64–70. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Jacobs AL, Sehgal PB, Julian J and Carson
DD: Secretion and hormonal regulation of interleukin-6 production
by mouse uterine stromal and polarized epithelial cells cultured in
vitro. Endocrinology. 131:1037–1346. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Szyllo K, Tchorzewski H, Banasik M,
Glowacka E, Lewkowicz P and Kamer-Bartosinska A: The involvement of
T lymphocytes in the pathogenesis of endometriotic tissues
overgrowth in women with endometriosis. Mediators Inflamm.
12:131–138. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kang YJ, Jeung IC, Park A, Park YJ, Jung
H, Kim TD, Lee HG, Choi I and Yoon SR: An increased level of IL-6
suppresses NK cell activity in peritoneal fluid of patients with
endometriosis via regulation of SHP-2 expression. Hum Reprod.
29:2176–2189. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Harada T, Yoshioka H, Yoshida S, Iwabe T,
Onohara Y, Tanikawa M and Terakawa N: Increased interleukin-6
levels in peritoneal fluid of infertile patients with active
endometriosis. Am J Obstet Gynecol. 176:593–597. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Bedaiwy MA, Falcone T, Sharma RK, Goldberg
JM, Attaran M, Nelson DR and Agarwal A: Prediction of endometriosis
with serum and peritoneal fluid markers: A prospective controlled
trial. Hum Reprod. 17:426–431. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
D'Amora P, Maciel TT, Tambellini R, Mori
MA, Pesquero JB, Sato H, Girão MJ, da Silva Guerreiro ID and Schor
E: Disrupted cell cycle control in cultured endometrial cells from
patients with endometriosis harboring the progesterone receptor
polymorphism PROGINS. Am J Pathol. 175:215–224. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Li MQ, Luo XZ, Meng YH, Mei J, Zhu XY, Jin
LP and Li DJ: CXCL8 enhances proliferation and growth and reduces
apoptosis in endometrial stromal cells in an autocrine manner via a
CXCR1-triggered PTEN/AKT signal pathway. Hum Reprod. 27:2107–2116.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Gonçalves GA, Invitti AL, Parreira RM,
Kopelman A, Schor E and Girão MJ: p27kip1 overexpression regulates
IL-1β in the microenvironment of stem cells and eutopic
endometriosis co-cultures. Cytokine. 89:229–234. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Prianishnikov VA: On the concept of stem
cell and a model of functional-morphological structure of the
endometrium. Contraception. 18:213–223. 1978. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Gargett CE, Chan RW and Schwab KE:
Endometrial stem cells. Curr Opin Obstet Gynecol. 19:377–383. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Sasson IE and Taylor HS: Stem cells and
the pathogenesis of endometriosis. Ann N Y Acad Sci. 1127:106–115.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Gargett BE and Chan RW: Endometrial
stem/progenitor cells and proliferative disorders of the
endometrium. Minerva Ginecol. 58:511–526. 2006.PubMed/NCBI
|
|
61
|
Gargett CE and Gurung S: Endometrial
mesenchymal stem/stromal cells, their fibroblast progeny in
endometriosis, and more1. Biol Reprod. 94:1292016. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Hufnagel D, Li F, Cosar E, Krikun G and
Taylor H: The role of stem cells in the etiology and
pathophysiology of endometriosis. Semin Reprod Med. 33:333–340.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Gargett CE and Masuda H: Adult stem cells
in the endometrium. Mol Hum Reprod. 16:818–834. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Kanayama N, Kajiwara Y, Goto J, el Maradny
E, Maehara K, Andou K and Terao T: Inactivation of interleukin-8 by
aminopeptidase N (CD13). J Leukoc Biol. 57:129–134. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Kehlen A, Egbert I, Thiele K, Fischer K,
Riemann D and Langner J: Increased expression of interleukin-8 and
aminopeptidase N by cell-cell contact: Interleukin-8 is resistant
to degradation by aminopeptidase N/CD13. Eur Cytokine Netw.
12:316–324. 2001.PubMed/NCBI
|
|
66
|
Mishima Y, Matsumoto-Mishima Y, Terui Y,
Katsuyama M, Yamada M, Mori M, Ishizaka Y, Ikeda K, Watanabe J,
Mizunuma N, et al: Leukemic cell-surface CD13/aminopeptidase N and
resistance to apoptosis mediated by endothelial cells. J Natl
Cancer Inst. 94:1020–1028. 2002. View Article : Google Scholar : PubMed/NCBI
|