Introduction
Intervertebral disc degeneration (IDD) is among the
causes of lower back pain, which leads to disability and societal
burden, and has a negative socioeconomic impact worldwide (1). It is estimated that worldwide, lower
back pain affects ≤84% of people at a certain point in their
lifetime, with 10% becoming chronically disabled (2). Currently, therapies for IDD primarily
aim to relieve pain symptoms, offering only temporary benefits
rather than a permanent cure (3).
Therefore, it is necessary to elucidate the underlying mechanisms
of IDD and lower back pain in order to develop novel effective
treatment methods for these conditions.
The intervertebral disc is a complex structure that
consists of three anatomical sub-structures, including the nucleus
pulposus (NP), annulus fibrosus and cartilaginous endplates, which
are essential for the maintenance of normal function of the disc
(4). Early IDD typically occurs in
the central NP and is characterized by a decrease in the number of
active cells and a loss of extracellular matrix within the NP
region (5). Previous studies have
identified certain molecules associated with IDD (6,7). For
instance, one study reported that early IDD is associated with the
downregulation of caveolin-1 expression and inhibition of canonical
Wnt signaling (6). Furthermore, a
recent study indicated that long non-coding RNAs (lncRNAs), a
subset of non-protein-coding transcripts that are >200
nucleotides in length, are involved in a series of biological
processes (8). Additionally,
competing endogenous RNAs (ceRNAs) participate in microRNA
(miRNA)-dependent crosstalk and contribute to the complexity of
gene regulation, and may aid in the understanding of disease
pathobiology (9). lncRNAs regulate
the expression of neighboring coding genes through various
mechanisms and may function as ceRNAs to modulate the derepression
of miRNA targets (10). In recent
years, the roles of lncRNAs in several diseases and cancers have
been investigated (11,12). However, potential roles of lncRNAs
in IDD are yet to be elucidated.
Wan et al (13) performed microarray data profiling
of NP samples from patients with IDD and identified numerous
differentially expressed lncRNAs and mRNAs in degenerative discs.
Reanalysis of previous datasets using different, updated methods
may lead to the identification of novel molecular markers
associated with the analyzed dataset (14,15).
In the present study, lncRNA and mRNA microarray data from patients
with IDD, including 5 samples with degenerative lumbar NP and 5
normal controls, were downloaded and reanalyzed. Differentially
expressed lncRNAs or differentially expressed genes (DEGs) were
identified and co-expression network analysis was performed,
followed by functional analysis of genes in the network.
Additionally, a ceRNA regulatory network was constructed based on
DEGs and lncRNAs in the co-expression network. Furthermore, a
literature search was performed to identify specific miRNAs
associated with IDD and a specific miRNA-associated ceRNA network
was extracted from the co-expression network. The present study
aimed to identify novel IDD-associated lncRNAs and genes, which may
guide further investigations and contribute to the development of
novel therapeutic strategies for the treatment of IDD.
Materials and methods
Collection of microarray data
The lncRNA and mRNA microarray of IDD was downloaded
from the Gene Expression Omnibus database (16) (http://www.ncbi.nlm.nih.gov/geo/) under the accession
number GSE56081 (13), which is a
subseries of the SuperSeries GSE67567 (13,17,18).
The Arraystar Human lncRNA microarray (version 2.0; Agilent_033010
Probe Name version; Arraystar Inc., Rockville, MD, USA) platform
was used. A total of 5 degenerative lumbar NP samples derived from
patients with IDD and 5 non-degenerative specimens from cadaveric
donors were included in this dataset. As described in the original
study by Wan et al (13),
all specimens were collected within 3 h following disc excision
(for discs from patients with IDD) or mortality (for discs from
cadaveric donors).
Data preprocessing and DEG
screening
Raw data was initially normalized using the robust
multi-array average method (19).
Subsequently, limma package version 3.28.21 (20) of Bioconductor 3.5 (http://www.bioconductor.org/packages/3.5/bioc/html/limma.html)
was adopted to identify DEGs between the IDD samples and normal
controls. False discovery rate (FDR) using Benjamini-Hochberg
method (21) in multi-test package
version 2.2.0 of Bioconductor (22) (http://www.bioconductor.org/packages/2.5/bioc/html/multtest.html)
was used to correct multiple testing to avoid false positives. Only
genes and lncRNAs with |log2fold change (FC)| >1 and
FDR <0.05 were selected as significantly differentially
expressed. Sequences and probe IDs were provided by the platform
but not gene and lncRNA names or symbols, therefore, the sequences
were aligned with hg19 genome annotation (Ensembl) (23) to identify genes corresponding with
each ID. Furthermore, heatmaps were constructed to present
expression profiles of differentially expressed lncRNAs and DEGs
using hierarchical clustering, which was performed using pheatmap
package (http://cran.r-project.org/web/packages/pheatmap/index.html)
in R based on Euclidean distances.
Co-expression network analysis of
differentially expressed lncRNAs and DEGs
To determine the functions of differentially
expressed lncRNAs, Pearson correlation analysis of the DEGs and
differentially expressed lncRNAs was performed using the cor()
function in R. Pearson correlation coefficients between-1 and 0 or
0 and 1 were considered to represent positive and negative
correlation, respectively. In the present study, co-expressed
lncRNA-mRNA pairs were identified when absolute Pearson correlation
coefficient was >0.6. In addition, lncRNA-mRNA co-expression
network was visualized using Cytoscape version 2.8.0 (24).
Functional analysis of genes in the
network
An ontology-based tool, clusterProfiler, offers
three methods for biological term classification and enrichment
analyses of genes, including group Gene Ontology (GO), enrichGO and
enrich Kyoto Encyclopedia of Genes and Genomes (KEGG) (25). In the present study, in order to
elucidate the biological function of genes enriched in the
co-expression network, GO biological process (BP) terms and KEGG
pathway analysis of genes in the network were performed using
enrichGO and enrichKEGG functions in R package clusterProfiler with
P<0.05 as a cutoff value (25).
Construction of ceRNA regulatory
network
To elucidate the roles of lncRNAs and mRNAs in the
co-expression network, data pertaining to lncRNAs and mRNAs was
combined with miRNA data in order to construct a miRNA-lncRNA-mRNA
ceRNA regulatory network. Initially, miRNAs that had regulatory
associations with lncRNAs in the co-expression network were
searched for in two databases; miRcode (26) (http://www.mircode.org/) and starBase (27) (http://starbase.sysu.edu.cn/). Subsequently, the
targets of the identified miRNAs were determined using the
miRTarBase database (http://mirtarbase.mbc.nctu.edu.tw), which includes
experimentally validated microRNA-target interactions (28). The overlapping genes between
targets of the identified miRNAs and mRNAs contained in the
co-expression network were retained for ceRNA construction. ceRNA
and disease-specific ceRNA networks were subsequently constructed.
Based on the identified miRNAs, miRNA targets in the co-expression
network and differentially expressed lncRNAs, a ceRNA regulatory
network was constructed and visualized using Cytoscape version
2.8.0 (24). Furthermore, specific
miRNAs associated with IDD were identified in previously reported
articles by combining search terms ‘intervertebral disc
degeneration’ AND ‘human’ AND ‘miRNA’ in PubMed (https://www.ncbi.nlm.nih.gov/pubmed/),
and a IDD-specific ceRNA network containing specific miRNAs was
extracted from the co-expression ceRNA-regulated network and
visualized using Cytoscape version 2.8.0 (24).
Results
Identification of DEGs and
differentially expressed lncRNAs
Following data normalization, differential
expression analysis of genes and lncRNAs was performed using the
dataset that included 5 IDD samples and 5 controls. A total of 967
genes and 137 lncRNAs were significantly differentially expressed
within the FDR <0.05 and |log2FC| >1 criteria.
Among them, 737 genes were upregulated and 230 genes were
downregulated; 60 lncRNAs were upregulated and 77 lncRNAs were
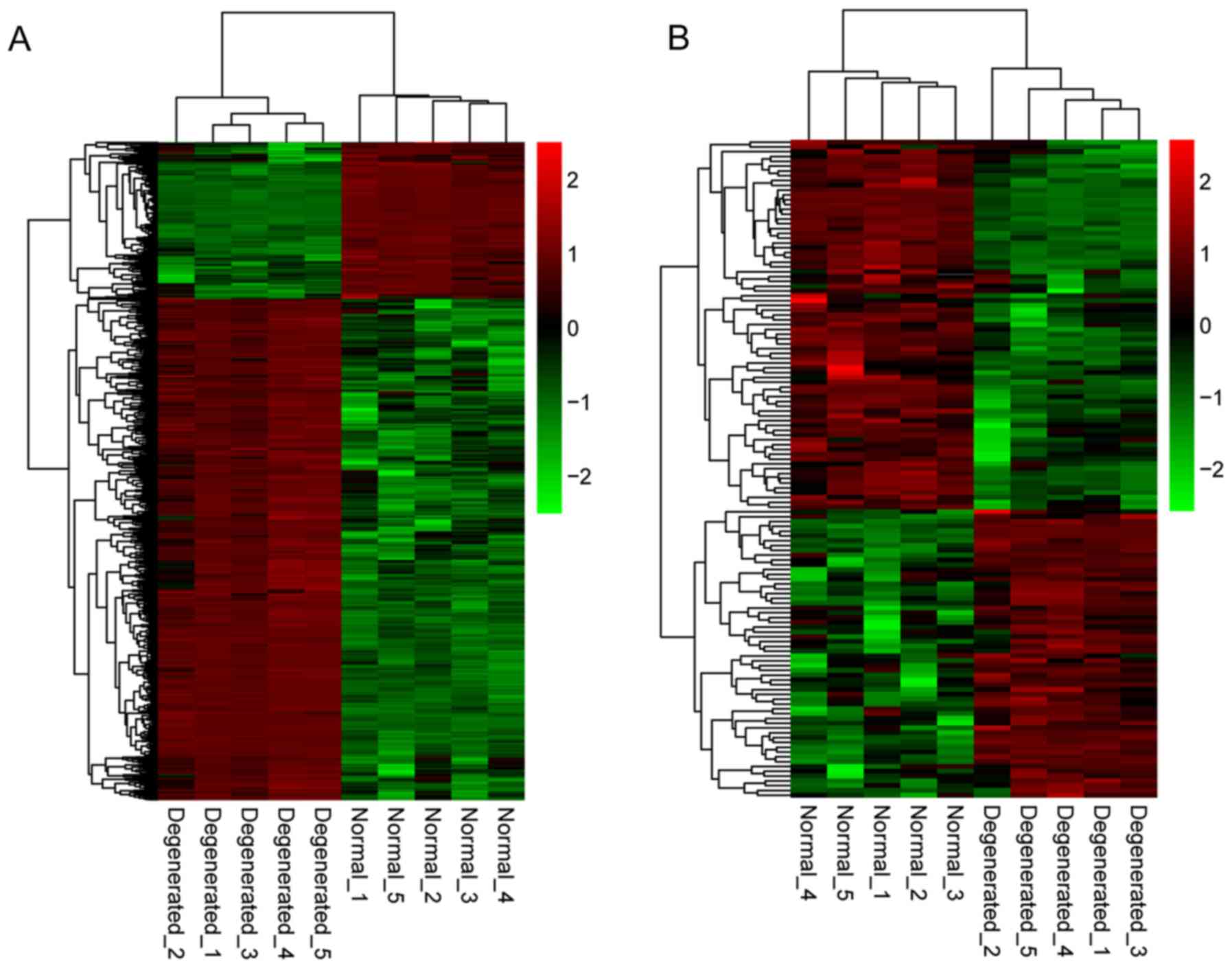
downregulated. Additionally, hierarchical clustering of gene
expression levels and lncRNA expression levels indicated clustering
of normal groups and IDD groups (Fig.
1).
Differential lncRNA-mRNA co-expression
network analysis
Differentially expressed lncRNAs and DEGs with
Pearson correlation coefficients >0.6 were selected. The
lncRNA-mRNA co-expression network, containing 248 nodes and 1,082
edges (448 negative and 634 positive connections), was constructed
(Fig. 2). A total of 39
differentially expressed lncRNAs (27 downregulated and 12
upregulated) and 209 DEGs (75 downregulated and 134 upregulated)
were included in this co-expression network.
Functional analysis of genes and
lncRNAs in the co-expression network
To determine the potential functions of the
identified DEGs and lncRNAs in the co-expression network, GO and
KEGG pathway enrichment analyses were performed in the
co-expression network. The analyses revealed the significant
enrichment of 17 GO BP terms and 10 KEGG pathways (Table I; Fig.
3). BP and KEGG terms that were significantly associated with
members of the co-expression network included ‘skeletal system
development’, ‘response to mechanical stimulus’, ‘bone development’
and ‘focal adhesion’ (Table I;
Fig. 3).
 | Table I.GO and KEGG pathway enrichment
analysis for genes in the lncRNA-mRNA co-expression network. |
Table I.
GO and KEGG pathway enrichment
analysis for genes in the lncRNA-mRNA co-expression network.
| A, GO terms
enriched in the lncRNA-mRNA co-expression network |
|---|
|
|---|
| Description (ID,
name) | Count | FDR | Genes |
|---|
| GO:0001501,
skeletal system development | 14 | 0.000067700 | RBP4, AEBP1,
LGALS3, TUFT1, KLF10, PRELP, MEN1, MEF2D, SBDS, LECT1, TNFSF11,
COL1A2, ACAN, PKD1 |
| GO:0009612,
response to mechanical stimulus | 5 | 0.003660942 | FOS, BTG2, JUN,
PKD1, GFI1 |
| GO:0007179,
transforming growth factor beta receptor signaling pathway | 5 | 0.004694322 | FOS, LTBP2, KLF10,
JUN, COL1A2 |
| GO:0040008,
regulation of growth | 11 | 0.005411270 | MEN1, EPC1, RBP4,
HTRA1, LGMN, BCAR1, NOL8, SIPA1, PEX5, GREM1, GAS6 |
| GO:0001503,
ossification | 6 | 0.009830659 | MEN1, MEF2D, SBDS,
TNFSF11, TUFT1, KLF10 |
| GO:0060348, bone
development | 6 | 0.012887071 | MEN1, MEF2D, SBDS,
TNFSF11, TUFT1, KLF10 |
| GO:0006357,
regulation of transcription from RNA polymerase II promoter | 16 | 0.017700242 | JDP2, EGR2, KLF9,
KLF10, FOXO1, NFYC, MEN1, PLAGL1, FOS, EPC1, SMARCD3, JUN, GFI1,
LCOR, KLF4, NR1H3 |
| GO:0045893,
positive regulation of transcription, DNA-dependent | 12 | 0.019627483 | PLAGL1, MEN1, FOS,
EPC1, EGR2, SMARCD3, JUN, FOXF1, NFYC, FOXO1, KLF4, NR1H3 |
| GO:0051254,
positive regulation of RNA metabolic process | 12 | 0.020731482 | PLAGL1, MEN1, FOS,
EPC1, EGR2, SMARCD3, JUN, FOXF1, NFYC, FOXO1, KLF4, NR1H3 |
| GO:0009628,
response to abiotic stimulus | 10 | 0.024048392 | MEN1, FOS, RBP4,
TNFSF11, BTG2, HMGCR, JUN, PKD1, GFI1, GRK1 |
| GO:0007178,
transmembrane receptor protein serine/threonine kinase signaling
pathway | 5 | 0.029381231 | FOS, LTBP2, KLF10,
JUN, COL1A2 |
| GO:0030198,
extracellular matrix organization | 5 | 0.030296507 | LGALS3, LUM, FOXF1,
COL1A2, ACAN |
| GO:0007611,
learning or memory | 5 | 0.037173540 | FOS, SBDS, EGR2,
HMGCR, JUN |
| GO:0010605,
negative regulation of macromolecule metabolic process | 15 | 0.038521124 | JDP2, KLF10, CDC26,
SIGIRR, TIMP1, MEN1, EPC1, SET, JUN, SERPINA5, BNIP3L, GFI1, LCOR,
KLF4, ADAR |
| GO:0051094,
positive regulation of developmental process | 8 | 0.038680685 | MEN1, TNFSF11,
KLF10, JUN, CLU, GFI1, OTP, ANGPTL4 |
| GO:0042592,
homeostatic process | 15 | 0.045211148 | CLNS1A, RBP4, EGR2,
NCDN, MBP, TIMP1, MEN1, TNFSF11, PRDX6, JUN, SERPINA3, TXNRD1,
XCR1, QSOX1, CLN5 |
| GO:0045597,
positive regulation of cell differentiation | 7 | 0.045820821 | MEN1, TNFSF11,
KLF10, JUN, CLU, GFI1, OTP |
|
| B, KEGG pathways
enriched in the lncRNA-mRNA co-expression network |
|
| Description
(entry, name) | Count | FDR | Genes |
|
| hsa04142,
Lysosome | 4 | 0.014272775 | CLTA, LGMN, CLN5,
CTSF |
| hsa04670, Leukocyte
transendothelial migration | 4 | 0.014534463 | BCAR1, SIPA1, CD99,
ITGAM |
| hsa03320, PPAR
signaling pathway | 3 | 0.018064527 | FADS2, ANGPTL4,
NR1H3 |
| hsa04510, Focal
adhesion | 5 | 0.018720786 | PARVG, PPP1CA, JUN,
BCAR1, COL1A2 |
| hsa04640,
Hematopoietic cell lineage | 3 | 0.025076738 | CD1B, CSF3R,
ITGAM |
| hsa03020, RNA
polymerase | 2 | 0.027129221 | POLR2C,
POLR2J2 |
| hsa00240,
Pyrimidine metabolism | 3 | 0.028840787 | TXNRD1, POLR2C,
POLR2J2 |
| hsa00051, Fructose
and mannose metabolism | 2 | 0.031922905 | PFKFB2, AKR1B1 |
| hsa04810,
Regulation of actin cytoskeleton | 4 | 0.043576076 | PPP1CA, BCAR1,
FGF13, ITGAM |
| hsa04514, Cell
adhesion molecules (CAMs) | 3 | 0.043833628 | CD99, SELPLG,
ITGAM |
Construction of ceRNA regulatory
network and disease-specific ceRNA network
A total of 79 associated miRNAs, which were targeted
by 9 lncRNAs in the co-expression network, were screened by
integrating information from miRcode and starBase databases to
determine regulatory associations between miRNAs and lncRNAs. A
total of 647 pairs of miRNA-gene interactions were obtained by
identifying targets of these miRNAs from genes in the co-expression
network using the miRTarBase database. The ceRNA regulatory network
was therefore constructed based on combining the identified
miRNA-gene and lncRNA-miRNA associations (Fig. 4). The ceRNA network was composed of
236 nodes [79 miRNAs, 9 downregulated lncRNAs, 148 DEGs (including
46 downregulated and 102 upregulated DEGs)] and 1,065 edges (647
miRNA-gene connections, 142 lncRNA-gene connections and 276
lncRNA-miRNAs connections).
Furthermore, IDD-associated miRNAs from previous
studies were searched and the collected data indicated that
hsa-miR-140 may be implicated in human IDD (29,30).
Subsequently, connections between hsa-miR-140 and other DEGs,
lncRNAs and miRNAs were extracted from the co-expression
ceRNA-regulated network and a hsa-miR-140-associated ceRNA network
was constructed (Fig. 5). The
disease-specific ceRNA network included 47 nodes (1 miRNA, 3
lncRNAs, 27 downregulated DEGs, and 16 upregulated DEGs) and 55
edges. hsa-miR-140 was demonstrated to be regulated by three
lncRNAs in the hsa-miR-140-associated ceRNA network, including
KCNQ1 opposite strand/antisense transcript 1 (KCNQ1OT1), OIP5
antisense RNA 1 (OIP5-AS1) and UGDH antisense RNA 1 (UGDH-AS1). The
results of the present study indicated that the aforementioned
lncRNAs may be associated with IDD and the genes that are
co-expressed with these lncRNAs may also be associated with IDD.
KCNQ1OT1 was co-expressed with neurochondrin (NCDN) and lon
peptidase 2, peroxisomal (LONP2). lncRNAs OIP5-AS1 and UGDH-AS1
were co-expressed with several overlapping genes, including
forkhead box F1 (FOXF1) and polycystin 1, transient receptor
potential channel interacting (PKD1).
Discussion
In the present study, 967 genes (737 upregulated and
230 downregulated) and 137 lncRNAs (60 upregulated and 77
downregulated) were differentially expressed between the IDD
samples and controls. A co-expression network was constructed
containing 39 differentially expressed lncRNAs and 209 DEGs that
were primarily involved in ‘skeletal system development’, ‘response
to mechanical stimulus’, ‘bone development’ and ‘focal adhesion’.
Furthermore, a ceRNA network was established, which included 79
miRNAs, 9 downregulated lncRNAs and 148 DEGs. Among the miRNAs,
hsa-miR-140 was a previously reported disease-associated miRNA and
therefore, a hsa-miR-140-associated ceRNA network was extracted. It
was determined that hsa-miR-140 was regulated by three lncRNAs,
including KCNQ1OT1, OIP5-AS1 and UGDH-AS1. KCNQ1OT1 was
co-expressed with NCDN and LONP2. In addition, lncRNAs OIP5-AS1 and
UGDH-AS1 targeted several overlapping co-expressed genes, including
FOXF1 and PKD1.
Aberrant expression of miR-140 was reported to
contribute to osteoarthritis, a degenerative disease characterized
by pathological alterations similar to those identified in patients
with IDD (31,32). miRNA-140 has also been demonstrated
to serve roles in cartilage development and homeostasis by
regulating the expression of matrix metalloprotease-13,
insulin-like growth factor binding protein-5, Sp1 transcription
factor, interleukin-1 and Smad family member 3 (31,33,34).
The results of the present study support the hypothesis that
miR-140 serves a role in the pathogenesis of degenerative disorders
such as IDD. The present study identified three lncRNAs, KCNQ1OT1,
OIP5-AS1 and UGDH-AS1, which were associated with disease-specific
miRNAs, including miR-140.
KCNQ1OT1 has been demonstrated to mediate
transcriptional silencing (35).
In the miR-140-associated ceRNA network, KCNQ1OT1 was co-expressed
with NCDN, which was enriched in the GO term ‘positive regulation
of developmental process’. A recent study reported that NCDN served
as a neuronal target antigen in patients with autoimmune cerebellar
degeneration (36). NCDN, a
cytoplasmic leucine-rich protein that functions in neurite
outgrowth and chondrocyte differentiation, was demonstrated to be
NP-specific and age-associated, indicating its potential functional
implication in the development of NP (37). Therefore, we hypothesize that the
downregulation of KCNQ1OT1 may be associated with the degeneration
of the intervertebral disc through the regulation of NCDN
expression.
Previous studies have demonstrated that lncRNA
OIP5-AS1 acts as a sponge to prevent RNA-binding protein HuR from
binding to its target mRNA and reduce cell proliferation (38). Stimulation of disc cell
proliferation and accumulation of extracellular matrix may slow or
even reverse the degenerative alterations resulting from IDD
(7). To the best of our knowledge,
the role of UGDH-AS1 has not been previously investigated. In the
present study, OIP5-AS1 and UGDH-AS1 shared several overlapping
co-expressed genes, including FOXF1 and PKD1. FOXF1 is a member of
the forkhead family of transcription factors, which are
characterized by a distinct forkhead domain (39). A previous study indicated that
FOXF1 promoted the production of extracellular matrix (40). Importantly, IDD is associated with
an increased breakdown of extracellular matrix (41). Boulter et al (42) reported that PKD1 may serve a role
in skeletal development. Based on the results of the aforementioned
studies and data analyzed in the present study, it may be
hypothesized that the lncRNAs OIP5-AS1 and UGDH-AS1 are involved in
the molecular mechanisms underlying IDD by regulating the
expression of FOXF1 and PKD1. The results of the present study
require further investigation to clarify and confirm the roles of
the identified molecules in IDD.
The present study had several limitations, including
the small sample size of the dataset employed; further analyses
with an increased sample size are necessary to confirm the results.
Furthermore, non-degenerative specimens from cadaveric donors were
used as controls in the dataset that was analyzed in the present
study. Patients with normal discs or with mild disc degeneration
are not typically subjected to surgical intervention. The use of
normal discs from cadaveric donors reflected limited access to less
degenerated tissues from surgical patients. However, cadaveric
tissues were collected within 3 h following mortality to ensure
that the tissues remained intact (13). Additionally, the results of the
analysis in the present study were not experimentally verified. IDD
is a complex and multi-factorial process that is influenced by
genetic predispositions, aging, biomechanical loading and physical
activities, lifestyle and other health-associated factors,
including diabetes (43). Numerous
clinical investigations have indicated that cell-based biological
therapies, including disc cell augmentation, application of growth
factors/cytokines and gene therapy, may be employed for the
treatment of IDD (44,45). The development of biomarkers for
IDD is valuable in the early stages of disc degeneration (45). lncRNAs have been implicated in
multiple pathological processes of IDD (46), and lncRNAs and DEGs in patients
with disc degeneration may have potential as diagnostic or
prognostic biomarkers (47). In
order to determine the potential for the clinical application of
lncRNAs and DEGs identified in the present study, an in vivo
murine IDD model may be used to confirm the roles of these
molecules in IDD in future studies.
In conclusion, the present study identified several
potential genes and lncRNAs, including KCNQ1OT1, OIP5-AS1,
UGDH-AS1, NCDN, FOXF1 and PKD1, that were associated with IDD.
KCNQ1OT1 may be involved in the degeneration of the intervertebral
disc by regulating the expression of NCDN, while the lncRNAs
OIP5-AS1 and UGDH-AS1 may be implicated in the molecular mechanisms
of IDD by affecting the expression of FOXF1 and PKD1. The results
of the present study may contribute to elucidating the molecular
pathogenesis of IDD and aid in the identification of novel
biomarkers associated with IDD.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZGQ, ZQ and YL provided the concept and design of
the research. QZ, ZYW and QS acquired the data. XZ, CF and ZW
analyzed and interpreted the data. FX, YDL and YW performed the
statistical analysis. ZGQ and ZQ drafted the manuscript. YL and ZW
revised the manuscript.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Vadalà G, Russo F, Ambrosio L, Papalia R
and Denaro V: Mesenchymal stem cells for intervertebral disc
regeneration. J Biol Regul Homeost Agents. 30 4 Suppl 1:S173–S179.
2016.
|
|
2
|
Risbud MV and Shapiro IM: Role of
cytokines in intervertebral disc degeneration: Pain and disc
content. Nat Rev Rheumatol. 10:44–56. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gantenbein B, Illien-Jünger S, Chan SC,
Walser J, Haglund L, Ferguson SJ, Iatridis C J and Grad S: Organ
culture bioreactors-platforms to study human intervertebral disc
degeneration and regenerative therapy. Curr Stem Cell Res Ther.
10:339–352. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Colombier P, Clouet J, Hamel O, Lescaudron
L and Guicheux J: The lumbar intervertebral disc: From embryonic
development to degeneration. Joint Bone Spine. 81:125–129. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yang W, Yu XH, Wang C, He WS, Zhang SJ,
Yan YG, Zhang J, Xiang YX and Wang WJ: Interleukin-1β in
intervertebral disk degeneration. Clin Chim Acta. 450:262–272.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Smolders LA, Meij BP, Onis D, Riemers FM,
Bergknut N, Wubbolts R, Grinwis GC, Houweling M, Koerkamp MJ, van
Leenen D, et al: Gene expression profiling of early intervertebral
disc degeneration reveals a down-regulation of canonical Wnt
signaling and caveolin-1 expression: implications for development
of regenerative strategies. Arthritis Res Ther. 15:R232013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang SZ, Rui YF, Tan Q and Wang C:
Enhancing intervertebral disc repair and regeneration through
biology: Platelet-rich plasma as an alternative strategy. Arthritis
Res Ther. 15:2202013. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liz J and Esteller M: lncRNAs and
microRNAs with a role in cancer development. Biochim Biophys Acta.
1859:169–176. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kartha RV and Subramanian S: Competing
endogenous RNAs (ceRNAs): New entrants to the intricacies of gene
regulation. Front Genet. 5:82014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sen K, Sarkar A, Maji RK, Ghosh Z, Gupta S
and Ghosh TC: Deciphering the cross-talking of human competitive
endogenous RNAs in K562 chronic myelogenous leukemia cell line. Mol
Biosyst. 12:3633–3642. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen EG, Zhang JS, Xu S, Zhu XJ and Hu HH:
Long non-coding RNA DGCR5 is involved in the regulation of
proliferation, migration and invasion of lung cancer by targeting
miR-1180. Am J Cancer Res. 7:1463–1475. 2017.PubMed/NCBI
|
|
12
|
Fayda M, Isin M, Tambas M, Guveli M, Meral
R, Altun M, Sahin D, Ozkan G, Sanli Y, Isin H, et al: Do
circulating long non-coding RNAs (lncRNAs) (LincRNA-p21, GAS 5,
HOTAIR) predict the treatment response in patients with head and
neck cancer treated with chemoradiotherapy? Tumour Biol.
37:3969–3978. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wan ZY, Song F, Sun Z, Chen YF, Zhang WL,
Samartzis D, Ma CJ, Che L, Liu X, Ali MA, et al: Aberrantly
expressed long noncoding RNAs in human intervertebral disc
degeneration: A microarray related study. Arthritis Res Ther.
16:4652014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Huang C, Nie F, Qin Z, Li B and Zhao X: A
snapshot of gene expression signatures generated using microarray
datasets associated with excessive scarring. Am J Dermatopathol.
35:64–73. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Minsky N and Roeder RG: Inhibition of
adhesion molecule gene expression and cell adhesion by the
metabolic regulator PGC-1α. PLoS One. 11:e01655982016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Barrett T, Wilhite SE, Ledoux P,
Evangelista C, Kim IF, Tomashevsky M, Marshall KA, Phillippy KH,
Sherman PM, Holko M, et al: NCBI GEO: Archive for functional
genomics data sets-update. Nucleic Acids Res. 41:(Database Issue).
D991–D995. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu X, Che L, Xie YK, Hu QJ, Ma CJ, Pei
YJ, Wu ZG, Liu ZH, Fan LY and Wang HQ: Noncoding RNAs in human
intervertebral disc degeneration: An integrated microarray study.
Genom Data. 5:80–81. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lan PH, Liu ZH, Pei YJ, Wu ZG, Yu Y, Yang
YF, Liu X, Che L, Ma CJ, Xie YK, et al: Landscape of RNAs in human
lumbar disc degeneration. Oncotarget. 7:63166–63176. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Allison DB, Cui X, Page GP and Sabripour
M: Microarray data analysis: From disarray to consolidation and
consensus. Nat Rev Genet. 7:55–65. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ritchie ME, Phipson B, Wu D, Hu Y, Law CW,
Shi W and Smyth GK: Limma powers differential expression analyses
for RNA-sequencing and microarray studies. Nucleic Acids Res.
43:e47. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Benjamini Y, Drai D, Elmer G, Kafkafi N
and Golani I: Controlling the false discovery rate in behavior
genetics research. Behav Brain Res. 125:279–284. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pollard KS, Dudoit S and Van Der Laan MJ:
Multiple testing procedures: The multtest package and applications
to genomics, in Bioinformatics and computational biology solutions
using R and bioconductor. Springer; pp. 249–271. 2005
|
|
23
|
Aken BL, Ayling S, Barrell D, Clarke L,
Curwen V, Fairley S, Banet Fernandez J, Billis K, Giron Garcia C,
Hourlier T, et al: The Ensembl gene annotation system. Database
(Oxford). 2016:pii:baw0932016. View Article : Google Scholar
|
|
24
|
Smoot ME, Ono K, Ruscheinski J, Wang PL
and Ideker T: Cytoscape 2.8: New features for data integration and
network visualization. Bioinformatics. 27:431–432. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yu G, Wang LG, Han Y and He QY:
ClusterProfiler: An R package for comparing biological themes among
gene clusters. OMICS. 16:284–287. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jeggari A, Marks DS and Larsson E:
miRcode: A map of putative microRNA target sites in the long
non-coding transcriptome. Bioinformatics. 28:2062–2063. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
Starbase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42:92–97. 2014. View Article : Google Scholar
|
|
28
|
Chou CH, Chang NW, Sirjana S, Hsu SD, Lin
YL, Lee WH, Yang CD, Hong HC, Wei TY, Tu SJ, et al: miRTarBase
2016: Updates to the experimentally validated miRNA-target
interactions database. Nucleic Acids Res. 44:D239–D247. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Swingler TE, Wheeler G, Carmont V, Elliott
HR, Barter MJ, Abuelmagd M, Donell ST, Boothandford RP,
Hajihosseini MK, Münsterberg A, et al: The expression and function
of microRNAs in chondrogenesis and osteoarthritis. Arthritis Rheum.
64:1909–1919. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tuddenham L, Wheeler G, Ntounia-Fousara S,
Waters J, Hajihosseini MK, Clark I and Dalmay T: The cartilage
specific microRNA-140 targets histone deacetylase 4 in mouse cells.
FEBS Lett. 580:4214–4217. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tardif G, Hum D, Pelletier JP, Duval N and
Martel-Pelletier J: Regulation of the IGFBP-5 and MMP-13 genes by
the microRNAs miR-140 and miR-27a in human osteoarthritic
chondrocytes. BMC Musculoskelet Disord. 10:1482009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tardif G, Pelletier JP, Fahmi H, Hum D,
Zhang Y, Kapoor M and Martel-Pelletier J: NFAT3 and TGF-β/SMAD3
regulate the expression of miR-140 in osteoarthritis. Arthritis Res
Ther. 15:R1972013. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yang J, Qin S, Yi C, Ma G, Zhu H, Zhou W,
Xiong Y, Zhu X, Wang Y, He L and Guo X: MiR-140 is co-expressed
with Wwp2-C transcript and activated by Sox9 to target Sp1 in
maintaining the chondrocyte proliferation. FEBS Lett.
585:2992–2997. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Miyaki S, Nakasa T, Otsuki S, Grogan SP,
Higashiyama R, Inoue A, Kato Y, Sato T, Lotz MK and Asahara H:
MicroRNA-140 is expressed in differentiated human articular
chondrocytes and modulates interleukin-1 responses. Arthritis
Rheum. 60:2723–2730. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mohammad F, Pandey RR, Nagano T, Chakalova
L, Mondal T, Fraser P and Kanduri C: Kcnq1ot1/Lit1 noncoding RNA
mediates transcriptional silencing by targeting to the
perinucleolar region. Mol Cell Biol. 28:3713–3728. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Miske R, Gross CC, Scharf M, Golombeck KS,
Hartwig M, Bhatia U, Schulte-Mecklenbeck A, Bönte K, Strippel C,
Schöls L, et al: Neurochondrin is a neuronal target antigen in
autoimmune cerebellar degeneration. Neurol Neuroimmunol
Neuroinflamm. 4:e3072017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tang X, Jing L and Chen J: Changes in the
molecular phenotype of nucleus pulposus cells with intervertebral
disc aging. PLoS One. 7:e520202012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kim J, Abdelmohsen K, Yang X, De S,
Grammatikakis I, Noh JH and Gorospe M: LncRNA OIP5-AS1/cyrano
sponges RNA-binding protein HuR. Nucleic Acids Res. 44:2378–2392.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mahlapuu M, Enerbäck S and Carlsson P:
Haploinsufficiency of the forkhead gene Foxf1, a target for sonic
hedgehog signaling, causes lung and foregut malformations.
Development. 128:2397–2406. 2001.PubMed/NCBI
|
|
40
|
Ormestad M, Astorga J, Landgren H, Wang T,
Johansson BR, Miura N and Carlsson P: Foxf1 and Foxf2 control
murine gut development by limiting mesenchymal Wnt signaling and
promoting extracellular matrix production. Development.
133:833–843. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Mccann MR and Séguin CA: Notochord cells
in intervertebral disc development and degeneration. J Dev Biol.
4:1–18. 2016. View Article : Google Scholar
|
|
42
|
Boulter C, Mulroy S, Webb S, Fleming S,
Brindle K and Sandford R: Cardiovascular, skeletal, and renal
defects in mice with a targeted disruption of the Pkd1 gene. Proc
Natl Acad Sci USA. 98:12174–12179. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kepler CK, Ponnappan RK, Tannoury CA,
Risbud MV and Anderson DG: The molecular basis of intervertebral
disc degeneration. Spine J. 13:318–330. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Gruber HE, Leslie K, Ingram J, Norton HJ
and Hanley EN: Cell-based tissue engineering for the intervertebral
disc: In vitro studies of human disc cell gene expression and
matrix production within selected cell carriers. Spine J. 4:44–55.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gruber HE and Hanley EN: Do we need
biomarkers for disc degeneration? Biomark Insights. 1:131–133.
2007.PubMed/NCBI
|
|
46
|
Chen WK, Yu XH, Yang W, Wang C, He WS, Yan
YG, Zhang J and Wang WJ: lncRNAs: Novel players in intervertebral
disc degeneration and osteoarthritis. Cell Prolif. 50:2017.
View Article : Google Scholar
|
|
47
|
Du Z, Fei T, Verhaak RG, Su Z, Zhang Y,
Brown M, Chen Y and Liu XS: Integrative genomic analyses reveal
clinically relevant long noncoding RNAs in human cancer. Nat Struct
Mol Biol. 20:908–913. 2013. View Article : Google Scholar : PubMed/NCBI
|