Introduction
Thyroid cancer is the most common type of malignancy
of the endocrine system (1);
annually, ~300,000 novel incidences and 40,000 cases of mortality
are caused by thyroid cancer worldwide (2). This disease can be divided into five
subtypes depending on different pathological features: Papillary
thyroid cancer (PTC), follicular thyroid cancer, poorly
differentiated thyroid cancer, anaplastic thyroid cancer and
thyroid squamous cell carcinoma (3). Amongst these types, PTC is the most
prevalent, and accounts for ~80% all thyroid cancer cases (4). The morbidity of PTC continuously
increases due to unhealthy diets and the exacerbation of
environmental pollution, which suppresses the immune system
(5). Thyroidectomy with
radioiodine ablation and thyroid-stimulating hormone-suppressive
therapy are primary therapeutic techniques for patients with PTC
(6). The majority of patients with
PTC exhibit favourable prognosis (7); however, the treatment outcomes of
patients diagnosed in advanced stages remain unsatisfactory
(1). Therefore, an in-depth
understanding of the mechanisms associated with cancer initiation
and progression is crucial for the identification of novel
therapeutic strategies for the treatment of this disease.
MicroRNAs (miRNAs/miRs) are a series of noncoding
and single-stranded RNA molecules existing universally in all
eukaryotic cells (8). miRNAs serve
as important posttranscriptional regulators by directly binding to
3′-untranslated regions (3′-UTRs) in a sequence-specific manner,
thereby inhibiting their translation or inducing mRNAs degradation
(9); >1,000 mature miRNAs have
been validated in the human genome, and these miRNAs can regulate
up to one-third of all protein-coding genes (10). In addition, >50% of miRNAs have
been detected in cancer-associated genomic regions or fragile sites
(11). This observation indicates
that miRNAs may be serve a role in oncogenesis and the progression
of cancer (12,13). Alterations in miRNA expression have
been reported in PTC and are associated with the occurrence and
development of PTC by regulating various cellular behaviours,
including cell proliferation, the cell cycle, apoptosis,
metastasis, tumour formation and epithelial-mesenchymal transition
(14–16). Thus, investigating the expression
pattern and detailed roles of miRNAs in PTC may aid the development
of potential therapeutic targets for patients with this
malignancy.
miR-744 is emerging as a cancer-associated miRNA in
numerous types of human cancer, including colorectal cancer
(17), cervical cancer (18) and hepatocellular carcinoma
(19); however, the expression and
specific functions of miR-744 in PTC are yet to be investigated,
and the mechanism underlying the regulatory roles of miR-744 in PTC
remains unknown. In the present study, we detected the expression
of miR-744 in PTC tissues and cell lines, and examined the effects
of miR-744 on the progression of PTC. In addition, the mechanism
underlying the tumour-suppressive roles of miR-744 in PTC was
investigated.
Materials and methods
Patients and samples
PTC tissues and adjacent non-cancerous tissues were
obtained from 31 patients (12 male, 19 female; age range, 34–67
years) who had undergone thyroidectomy at Weifang People's Hospital
(Weifang, China) between March 2014 and June 2017. No patients
recruited in this study had been treated with chemotherapy,
radiotherapy, radioiodine ablation or thyroid-stimulating
hormone-suppressive therapy prior to surgical resection. All
tissues were quickly snap-frozen in liquid nitrogen and then stored
at −80°C until RNA isolation. The present study was approved by the
Ethics Committee of Weifang People's Hospital. In addition, written
informed consent was obtained from all participants.
Cell culture and transfection
A total of three human PTC cell lines (TPC-1, BCPAP
and HTH83) and one normal human thyroid cell line (HT-ori3) were
purchased from the American Type Culture Collection (Manassas, VA,
USA). All cell lines were cultured in Dulbecco's Modified Eagle's
medium (DMEM) supplemented with 10% fetal bovine serum (FBS; both
from Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA), 100
U/ml penicillin and 100 µg/ml streptomycin (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany), and cultured at 37°C in a 5%
CO2 humidified incubator. TPC-1 and HTH83 cell lines
exhibited the lowest miR-744 expression levels among the PTC cell
lines. Therefore, these two cell lines were selected for functional
experiments.
The miR-744 mimics and miRNA mimics negative control
(miR-NC) were chemically synthesized by Shanghai GenePharma Co.,
Ltd. (Shanghai, China). The miR-744 mimics sequence was
5′-UGCGGGGCUAGGGCUAACAGCA-3′ and the miR-NC sequence was
5′-UUCUCCGAACGUGUCACGUTT-3′. NIN1 (RPN12) binding protein homolog 1
(NOB1) overexpression plasmid pcDNA3.1-NOB1 and empty pcDNA3.1
plasmid were purchased from GeneRay (Shanghai, China). For
transfection, cells were grown in DMEM containing 10% FBS without
antibiotics and were plated into 6-well plates with a density of
60–70% confluence. Transient transfection of miRNA mimics (100
pmol) and plasmid (4 µg) was conducted using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) according to the manufacturer's
protocols. Transfection efficiencies of miR-744 mimics and
pcDNA3.1-NOB1 were detected using reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) and
western blot analysis at 48 and 72 h after transfection,
respectively. Cell Counting Kit-8 (CCK-8) assay was performed at 24
h post-transfection, while Transwell invasion assay was conducted
following 48 h incubation.
RNA isolation and RT-qPCR
Total RNA was isolated from tissue samples or cells
using TRIzol® (Thermo Fisher Scientific, Inc.). Then,
total RNA was subjected to the detection of concentration using a
NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies; Thermo
Fisher Scientific, Inc., Wilmington, DE, USA). To quantify miR-744
expression, a TaqMan MicroRNA Reverse Transcription Kit (Applied
Biosystems; Thermo Fisher Scientific, Inc.) was utilized to reverse
transcribe RNA into complementary DNA (cDNA). Subsequently, qPCR
was performed on the ABI 7500 Sequence Detection System (Applied
Biosystems; Thermo Fisher Scientific, Inc.) using a TaqMan MicroRNA
PCR Kit (Applied Biosystems; Thermo Fisher Scientific, Inc.). The
thermocycling conditions for qPCR were: 50°C for 2 min, 95°C for 10
min; 40 cycles of denaturation at 95°C for 15 sec; and
annealing/extension at 60°C for 60 sec. To analyze NOB1 mRNA
expression, cDNA was synthesized using the PrimeScript®
RT reagent kit (Takara Bio, Inc., Otsu, Japan) followed by qPCR
with the SYBR Premix Ex Taq™ II kit (Takara Bio, Inc.). The
thermocycling conditions were as follows: 5 min at 95°C, followed
by 40 cycles of 95°C for 30 sec and 65°C for 45 sec. U6 small
nuclear RNA and GAPDH served as endogenous references for miR-744
and NOB1 mRNA, respectively. Relative gene expression was
calculated using the 2−ΔΔCq method (20). The primers were designed as
follows: miR-744, 5′-ACACTCCAGCTGGGTGCGGGGCTAGGGCTAAC-3′ (forward)
and 5′-CTCAACTGGTGTCGTGGA-3′ (reverse); U6,
5′-TCCAAGTGCCGAAAAAGGAAG-3′ (forward) and
5′-CGAGTTCTGAGCTTTCAAGGT-3′ (reverse); NOB1,
5′-GAAAGAACAACGCCCTGGAG-3′ (forward) and
5′-CAGCCTTGAGATGACCTAAGC-3′ (reverse); and GAPDH,
5′-CGGAGTCAACGGATTTGGTCGTAT-3′ (forward) and
5′-AGCCTTCTCCATGGTGGTGAAGAC-3′ (reverse).
Cell Counting Kit-8 (CCK-8) assay
Transfected cells were seeded into 96-well plates at
a density of 2×103 cells/well and incubated at 37°C in a
5% CO2 humidified incubator. A CCK-8 assay was conducted
to detect cell proliferation at 0, 24, 48 and 72 h after
inoculation. In brief, 10 µl of CCK-8 reagent (Dojindo Molecular
Technologies, Inc., Shanghai, China) was added to each well, and
the transfected cells were incubated at 37°C for another 2 h. The
optical density at 490 nm was determined using an ELISA plate
reader (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Each
experiment was performed in quadruplicate.
Transwell invasion assay
Matrigel-coated Transwell chambers (pore size, 8 µm;
BD Biosciences, San Jose, CA, USA) were used to evaluate cell
invasion. After 24 h following incubation, transfected cells were
cultured in FBS-free DMEM at 37°C with 5% CO2 for
additional 24 h. Then, the transfected cells were then harvested
and resuspended in FBS-free DMEM. A total of 5×104
transfected cells were placed into the upper chambers. The lower
chambers were covered with DMEM supplemented with 20% FBS as a
chemoattractant.
After 24 h of incubation, non-invasive cells
remaining on the upper chambers were carefully removed using a
cotton swab. The invasive cells attached to the lower surface of
the membranes were fixed with 70% ethanol at room temperature for
20 min and stained with 0.5% crystal violet at room temperature for
20 min. Then, stained cells in five randomly selected
fields/chamber were counted under a light microscope
(magnification, ×200; IX71; Olympus, Tokyo, Japan).
Bioinformatics prediction and
luciferase reporter assay
TargetScan (release 7.2; http://www.targetscan.org) and miRDB (last modified:
November 7, 2018; http://mirdb.org) were applied to
predict the putative targets of miR-744. The 3′-UTR fragments of
NOB1 containing the wild-type (wt) or mutant (mut) miR-744 binding
site were amplified by Shanghai GenePharma Co., Ltd. and cloned
into the pGL3 luciferase reporter plasmid (Promega Corporation,
Madison, WI, USA) to construct pGL3-NOB1-3′-UTR wt and
pGL3-NOB1-3′-UTR mut, respectively. Cells were seeded into 24-well
plates, and were co-transfected with an oligonucleotide (miR-744
mimics or miR-NC) and one of the two reporter plasmids
(pGL3-NOB1-3′-UTR wt or pGL3-NOB1-3′-UTR mut) using Lipofectamine
2000, in accordance with the manufacturer's protocols. Luciferase
activities were measured at 48 h post-transfection using the
Dual-Luciferase Reporter System (Promega Corporation). The activity
of firefly luciferase was normalized to that of Renilla
luciferase.
Western blot analysis
Total protein was isolated from tissue samples or
cultured cells using radioimmunoprecipitation assay buffer (Nanjing
KeyGen Biotech Co., Ltd., Nanjing, China) that was supplemented
with protease inhibitor (Roche Diagnostics, Basel, Switzerland). A
BCA Protein Assay Kit (Nanjing KeyGen Biotech Co., Ltd.) was
adopted to quantify the concentration of total protein. Equal
amounts of protein (30 µg) were resolved by 10% SDS-PAGE and
transferred to polyvinylidene difluoride membranes (EMD Millipore,
Billerica MA, USA). The membranes were then incubated at room
temperature for 2 h with 5% fat-free milk that was dissolved in
Tris-buffered saline with Tween 20 (TBST) and further incubated at
4°C overnight with the following primary antibodies: Rabbit
anti-human polyclonal NOB1 (cat. no. ab224619; 1:1,000 dilution;
Abcam, Cambridge, UK) and rabbit anti-human monoclonal GAPDH (cat.
no. ab201822; 1:1,000 dilution; Abcam). Following three washes with
TBST, the membranes were exposed to goat anti-rabbit horseradish
peroxidase-conjugated secondary antibody (cat. no. ab205718;
1:5,000 dilution; Abcam) at room temperature for 1 h. A
Plus-enhanced chemiluminescence reagent (PerkinElmer, Inc.,
Waltham, MA, USA) was employed to detect the protein signals. GAPDH
was used as an internal control. Protein expression was quantified
using Quantity One software version 4.62 (Bio-Rad Laboratories,
Inc., Hercules, CA, USA). Each assay was repeated three times.
Statistical analysis
Each assay was repeated at least three times. All
data were presented as the mean ± standard deviation. SPSS version
17.0 (IBM Corp., Chicago, IL, USA) was used to perform statistical
analysis. The association between miR-744 and NOB1 mRNA expression
in PTC tissues was determined using Pearson's correlation analysis.
A two-tailed Student's t-test and one-way analysis of variance
followed by a Tukey's post-hoc test were utilized to analyze the
differences between multiple groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
miR-744 expression is downregulated in
PTC tissues and cell lines
To determine the expression profile of miR-744 in
PTC, miR-744 expression in 31 paired PTC and adjacent noncancerous
tissues were analyzed via RT-qPCR. The expression levels of miR-744
were significantly decreased in PTC tissues than in the adjacent
noncancerous tissues (P<0.05; Fig.
1A). miR-744 expression was also determined in three human PTC
cell lines, namely, TPC-1, BCPAP and HTH83, and one normal human
thyroid cell line, HT-ori3. RT-qPCR analysis revealed that miR-744
was significantly downregulated in all of PTC cell lines compared
with in HT-ori3 (P<0.05; Fig.
1B). TPC-1 and HTH83 cell lines exhibited the lowest miR-744
expression levels among the PTC cell lines. Therefore, these two
cell lines were selected for functional experiments. In summary,
these results suggested that miR-744 may serve a crucial role in
the development of PTC.
miR-744 suppresses cell proliferation
and invasion in PTC
Considering the significant downregulation of
miR-744 in PTC, it was proposed that this miRNA may serve
tumour-suppressive roles in PTC cells. Thus, miR-744 mimics or
miR-NC were transfected into TPC-1 and HTH83 cells; RT-qPCR
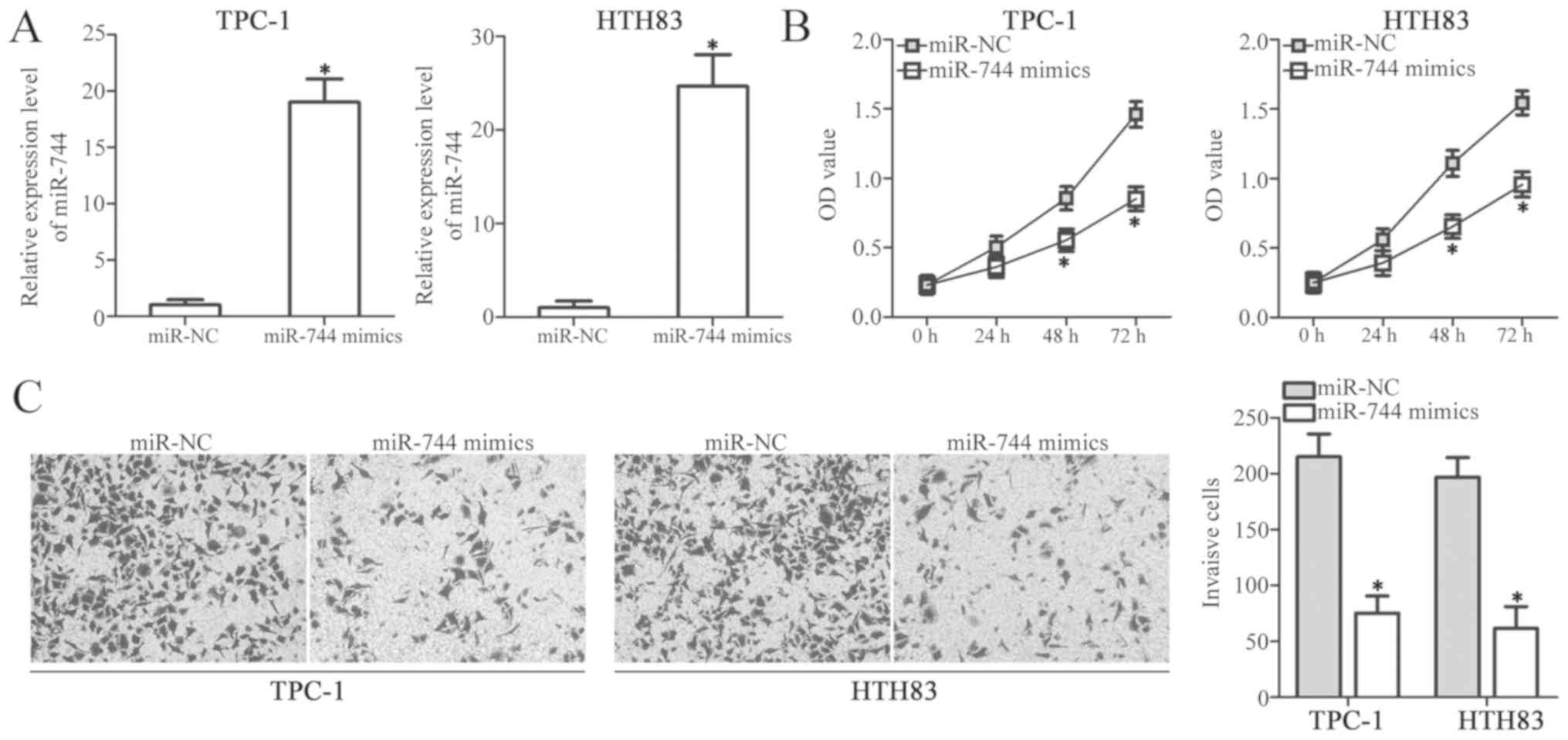
analysis was conducted to determine the transfection efficiency.
miR-744 expression was significantly upregulated in TPC-1 and HTH83
cells transfected with miR-744 mimics compared with the control
(P<0.05; Fig. 2A). A CCK-8
assay was then performed to detect the proliferation of TPC-1 and
HTH83 cells following transfection with miR-744 mimics or miR-NC.
The restoration of miR-744 expression significantly inhibited the
proliferative ability of TPC-1 and HTH83 cells compared with the
miR-NC groups (P<0.05; Fig.
2B). Furthermore, a Transwell invasion assay was performed to
investigate the effect of miR-744 overexpression in PTC cell
invasion. As presented in Fig. 2C,
cell invasion significantly decreased in the miR-744
mimics-transfected TPC-1 and HTH83 cells compared with cells
transfected with miR-NC (P<0.05). These results indicated that
miR-744 overexpression may inhibit the progression of PTC.
NOB1 is a direct target gene of
miR-744 in PTC
To clarify the mechanisms by which miR-744 inhibits
the progression of PTC, bioinformatics analysis was performed to
predict the putative targets of miR-744. NOB1 was reported as
potential target of miR-744 (Fig.
3A) and was selected for further identification as that this
gene is associated with numerous cancer-related processes in PTC
(21–23). To determine whether miR-744 can
target the 3′-UTR of NOB1, we transiently co-transfected TPC-1 and
HTH83 cells with pGL3-NOB1-3′-UTR wt or pGL3-NOB1-3′-UTR mut, and
miR-744 mimics or miR-NC. Following culture for 48 h, a luciferase
reporter assay was utilised to detect the luciferase activity. The
results demonstrated that miR-744 upregulation significantly
reduced the luciferase activity of the plasmid containing a wt
binding site (P<0.05) compared with the corresponding miR-NC
group; however, the luciferase activity of the plasmid harbouring
the mutant miR-744 binding site in TPC-1 and HTH83 cells was
markedly altered (Fig. 3B). NOB1
was also significantly downregulated at the mRNA (P<0.05;
Fig. 3C) and protein (P<0.05;
Fig. 3D and E) levels in TPC-1 and
HTH83 cells transfected with miR-744 mimics compared with in cells
transfected with miR-NC. These results supported our hypothesis
that NOB1 was a direct target of miR-744 in PTC cells.
NOB1 is upregulated in PTC tissues and
negatively correlated with miR-744 expression
In addition, the present study investigated NOB1
expression in PTC tissues and determine the associated with the
miR-744 expression. RT-qPCR revealed that the mRNA expression of
NOB1 was significantly upregulated in the PTC clinical samples
compared with in the adjacent non-cancerous tissues (P<0.05;
Fig. 4A). Spearman's correlation
analysis demonstrated that the mRNA expression of NOB1 was
inversely correlated with that miR-744 expression in PTC tissues
(r=−0.5233, P=0.0025; Fig.
4B).
NOB1 overexpression suppresses
miR-744-induced inhibition of PTC cell proliferation and
invasion
A series of ‘rescue’ experiments were performed to
demonstrate whether NOB1 mediates the functional roles of miR-744
in PTC cells. TPC-1 and HTH83 cells were treated with pcDNA3.1-NOB1
or empty pcDNA3.1 plasmid. Western blot analysis was performed to
evaluate the transfection efficiency; NOB1 protein expression was
significantly upregulated in TPC-1 and HTH83 cells transfected with
pcDNA3.1-NOB1 (P<0.05; Fig.
5A). Subsequently, miR-744 mimics were co-transfected with
pcDNA3.1-NOB1 or pcDNA3.1 into TPC-1 and HTH83 cells. NOB1
downregulation induced by miR-744 overexpression was restored in
the TPC-1 and HTH83 cells co-transfected with pcDNA3.1-NOB1
(P<0.05; Fig. 5B and C). NOB1
upregulation could antagonise the miR-744-induced inhibition of
TPC-1 and HTH83 cell proliferation (P<0.05; Fig. 5D) and invasion (P<0.05; Fig. 5E). In summary, these results
suggested that miR-744 inhibited the proliferation and invasion of
PTC cells, at least in part, by inhibiting NOB1.
Discussion
miRNAs serve important roles in the formation and
progression of PTC by regulating numerous physiological and
pathological behaviours (24,25).
Hence, investigating the functional roles of specific miRNAs in PTC
may contribute in identifying effective therapeutic targets for the
management of patients with PTC. In the present study, miR-744
expression in PTC was determined and the roles of miR-744 in the
progression of PTC and the potential underlying molecular
mechanisms were investigated. miR-744 expression was downregulated
in PTC tissues and cell lines. This observation is consistent with
previous findings on the expression profile of miR-744 in
colorectal cancer (17), cervical
cancer (18) and hepatocellular
carcinoma (19); however, miR-744
expression is upregulated in numerous types of human malignancy.
For instance, miR-744 is overexpressed in the tissues, cell lines
and plasma of pancreatic cancer (26,27).
miR-744 overexpression is significantly correlated with clinical
stage, lymph node metastasis and recurrence (26). miR-744 has also been reported as an
independent poor prognostic factor for patients with pancreatic
cancer (26,27) and has exhibited high expression in
prostate cancer (28), laryngeal
squamous cell carcinoma (29) and
nasopharyngeal carcinoma (30).
These conflicting findings suggest that the expression profile of
miR-744 exhibits tissue specificity in malignant tumors.
Considering that miR-744 is downregulated in PTC, it
was hypothesised that miR-744 may serve a tumour-suppressive role
in the development of PTC. To confirm this hypothesis, CCK-8 and
Transwell invasion assays were conducted using PTC cells
transfected with miR-744 mimics or miR-NC in the present study. The
results revealed that miR-744 restoration inhibited PTC cell
proliferation and invasion in vitro. miR-744 is also
identified as a tumour suppressor in numerous types of human
cancer. For example, miR-744 suppresses cell proliferation and
invasion in colorectal cancer (17) and cervical cancer (18). miR-744 upregulation inhibits cell
proliferation and promotes cell cycle arrest in hepatocellular
carcinoma (31). On the contrary,
miR-744 serves oncogenic roles in prostate cancer (28), laryngeal squamous cell carcinoma
(29), pancreatic cancer (27) and nasopharyngeal carcinoma
(32). These findings indicate
that the biological roles of miR-744 exhibit tissue specificity in
tumorigenesis and tumour development. Therefore, this miRNA may be
a potential therapeutic target for the treatment of patients with
these specific types of cancer.
miRNAs serve crucial roles in the regulation of
their target genes (33). Numerous
human genes, including Notch1 (17), B-cell lymphoma 2 (18), secreted frizzled-related protein 1
precursor (27), glycogen synthase
kinase 3β (27), transducing like
enhancer of split 2 (27), naked
cuticle homolog 1 (28),
programmed cell death 4 (29),
phosphatase and tensin homolog (29), c-myc (31) and Rho GTPase activating protein 5
(32), have been identified as
direct miR-744 targets. In the present study, NOB1 was predicted as
a direct target gene of miR-744 in PTC. NOB1 is an essential
protein encoding the Nin one binding protein by two-hybrid
screening and has been reported in proteasome and ribosome
biogenesis (34,35). NOB1 is highly expressed numerous
types of human cancer, including osteosarcoma (36), lung cancer (37), ovarian cancer (38) and colorectal cancer (39). NOB1 is also overexpressed in PTC,
and its overexpression is closely associated with tumour size and
Union for International Cancer Control stages (21). NOB1 inhibition suppresses the cell
growth and metastasis of PTC, promotes cell apoptosis and arrest in
the G0/G1 stage in vitro and decreases tumour growth in
vivo (22,23). NOB1 downregulation improves the
sensitivity of PTC cells to radiotherapy and chemotherapy (22,23).
These findings suggest that the miR-744-mediated gene silencing of
NOB1 may represent a promising therapeutic strategy for patients
with PTC.
In summary, the present study demonstrated that
miR-744 was downregulated in PTC tissues and cell lines. Functional
experiments revealed that ectopic miR-744 expression attenuated the
proliferation and invasion of PTC cells in vivo.
Mechanistically, NOB1 was identified as a direct target gene of
miR-744 in PTC cells. The results of the present study provided
novel insights into the mechanism underlying the pathogenesis of
PTC and proposed a valuable therapeutic target for the treatment of
patients with this disease; however, the sample size was small.
Therefore, the association between miR-744 and the prognosis of
patients with PTC requires further investigation. In addition, the
regulatory effects of miR-744 on the PTC tumor growth and
metastasis in vivo were not investigated. Furthermore, we
did not conduct phenotypic trials with the agents targeting NOB1.
These limitations of our study are to be resolved these in future
experiments.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
XM made substantial contributions to the design of
the present study and analyzed the data. HL and JG performed the
CCK-8 and Transwell invasion assays. HC conducted RT-qPCR, the
luciferase reporter assay and western blot analysis. All authors
read and approved the final draft of the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Research
Ethics Committee of Weifang People's Hospital (Weifang, China), and
was performed in accordance with the Declaration of Helsinki and
the guidelines of the Ethics Committee of Weifang People's
Hospital. Written informed consent was obtained from all patients
for the use of their clinical tissues.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Liebner DA and Shah MH: Thyroid cancer:
Pathogenesis and targeted therapy. Ther Adv Endocrinol Metab.
2:173–195. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Catalano MG, Poli R, Pugliese M, Fortunati
N and Boccuzzi G: Emerging molecular therapies of advanced thyroid
cancer. Mol Aspects Med. 31:215–226. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu S, Semenciw R, Ugnat AM and Mao Y:
Increasing thyroid cancer incidence in Canada, 1970–1996: Time
trends and age-period-cohort effects. Br J Cancer. 85:1335–1339.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cancer Genome Atlas Research Network:
Integrated genomic characterization of papillary thyroid carcinoma.
Cell. 159:676–690. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Toniato A, Boschin I, Casara D, Mazzarotto
R, Rubello D and Pelizzo M: Papillary thyroid carcinoma: Factors
influencing recurrence and survival. Ann Surg Oncol. 15:1518–1522.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hobert O: Gene regulation by transcription
factors and microRNAs. Science. 319:1785–1786. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Galasso M, Sandhu SK and Volinia S:
MicroRNA expression signatures in solid malignancies. Cancer J.
18:238–243. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhao Y and Srivastava D: A developmental
view of microRNA function. Trends Biochem Sci. 32:189–197. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Anfossi S, Fu X, Nagvekar R and Calin GA:
MicroRNAs, regulatory messengers inside and outside cancer cells.
Adv Exp Med Biol 1056. 87–108. 2018. View Article : Google Scholar
|
|
12
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tie J and Fan D: Big roles of microRNAs in
tumorigenesis and tumor development. Histol Histopathol.
26:1353–1361. 2011.PubMed/NCBI
|
|
14
|
Li R, Dong B, Wang Z, Jiang T and Chen G:
MicroRNA-361-5p inhibits papillary thyroid carcinoma progression by
targeting ROCK1. Biomed Pharmacother. 102:988–995. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang R, Ma Q, Ji L, Yao Y, Ma M and Wen Q:
miR-622 suppresses tumor formation by directly targeting VEGFA in
papillary thyroid carcinoma. Onco Targets Ther. 11:1501–1509. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fang L, Kong D and Xu W: MicroRNA-625-3p
promotes the proliferation, migration and invasion of thyroid
cancer cells by up-regulating astrocyte elevated gene 1. Biomed
Pharmacother. 102:203–211. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shen J and Li M: MicroRNA-744 inhibits
cellular proliferation and invasion of colorectal cancer by
directly targeting oncogene Notch1. Oncol Res. Feb 22–2018.(Epub
ahead of print). View Article : Google Scholar
|
|
18
|
Chen XF and Liu Y: MicroRNA-744 inhibited
cervical cancer growth and progression through apoptosis induction
by regulating Bcl-2. Biomed Pharmacother. 81:379–387. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tan YL, Bai ZG, Zou WL, Ma XM, Wang TT,
Guo W, Liu J, Li JS, Jie-Yin, Zang YJ and Zhang ZT: miR-744 is a
potential prognostic marker in patients with hepatocellular
carcinoma. Clin Res Hepatol Gastroenterol. 39:359–365. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lin S, Meng W, Zhang W, Liu J, Wang P, Xue
S and Chen G: Expression of the NOB1 gene and its clinical
significance in papillary thyroid carcinoma. J Int Med Res.
41:568–572. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu J, Dong BF, Wang PS, Ren PY, Xue S,
Zhang XN, Han Z and Chen G: Silencing NOB1 enhances doxorubicin
antitumor activity of the papillary thyroid carcinoma in vitro and
in vivo. Oncol Rep. 33:1551–1559. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Meng W, Wang PS, Liu J, Xue S, Wang GM,
Meng XY and Chen G: Adenovirus-mediated siRNA targeting NOB1
inhibits tumor growth and enhances radiosensitivity of human
papillary thyroid carcinoma in vitro and in vivo. Oncol Rep.
32:2411–2420. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lee JC, Gundara JS, Glover A, Serpell J
and Sidhu SB: MicroRNA expression profiles in the management of
papillary thyroid cancer. Oncologist. 19:1141–1147. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chruscik A and Lam AK: Clinical
pathological impacts of microRNAs in papillary thyroid carcinoma: A
crucial review. Exp Mol Pathol. 99:393–398. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Miyamae M, Komatsu S, Ichikawa D,
Kawaguchi T, Hirajima S, Okajima W, Ohashi T, Imamura T, Konishi H,
Shiozaki A, et al: Plasma microRNA profiles: Identification of
miR-744 as a novel diagnostic and prognostic biomarker in
pancreatic cancer. Br J Cancer. 113:1467–1476. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhou W, Li Y, Gou S, Xiong J, Wu H, Wang
C, Yan H and Liu T: MiR-744 increases tumorigenicity of pancreatic
cancer by activating Wnt/β-catenin pathway. Oncotarget.
6:37557–37569. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Guan H, Liu C, Fang F, Huang Y, Tao T,
Ling Z, You Z, Han X, Chen S, Xu B and Chen M: MicroRNA-744
promotes prostate cancer progression through aberrantly activating
Wnt/β-catenin signaling. Oncotarget. 8:14693–14707. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li JZ, Gao W, Lei WB, Zhao J, Chan JY, Wei
WI, Ho WK and Wong TS: MicroRNA 744-3p promotes MMP-9-mediated
metastasis by simultaneously suppressing PDCD4 and PTEN in
laryngeal squamous cell carcinoma. Oncotarget. 7:58218–58233.
2016.PubMed/NCBI
|
|
30
|
Yu Q, Zhang F, Du Z and Xiang Y:
Up-regulation of serum miR-744 predicts poor prognosis in patients
with nasopharyngeal carcinoma. Int J Clin Exp Med. 8:13296–13302.
2015.PubMed/NCBI
|
|
31
|
Lin F, Ding R, Zheng S, Xing D, Hong W,
Zhou Z and Shen J: Decrease expression of microRNA-744 promotes
cell proliferation by targeting c-Myc in human hepatocellular
carcinoma. Cancer Cell Int. 14:582014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fang Y, Zhu X, Wang J, Li N, Li D, Sakib
N, Sha Z and Song W: MiR-744 functions as a proto-oncogene in
nasopharyngeal carcinoma progression and metastasis via
transcriptional control of ARHGAP5. Oncotarget. 6:13164–13175.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lim LP, Lau NC, Garrett-Engele P, Grimson
A, Schelter JM, Castle J, Bartel DP, Linsley PS and Johnson JM:
Microarray analysis shows that some microRNAs downregulate large
numbers of target mRNAs. Nature. 433:769–773. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tone Y, Tanahashi N, Tanaka K, Fujimuro M,
Yokosawa H and Toh e-A: Nob1p, a new essential protein, associates
with the 26S proteasome of growing saccharomyces cerevisiae cells.
Gene. 243:37–45. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wyler E, Zimmermann M, Widmann B, Gstaiger
M, Pfannstiel J, Kutay U and Zemp I: Tandem affinity purification
combined with inducible shRNA expression as a tool to study the
maturation of macromolecular assemblies. RNA. 17:189–200. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Luo L, Wang Y, Yin Y, Ge J and Lu X:
Effects of NOB1 on the pathogenesis of osteosarcoma and its
expression on the chemosensitivity to cisplatin. Oncol Lett.
15:3548–3551. 2018.PubMed/NCBI
|
|
37
|
Kong R, Liu W, Guo Y, Feng J, Cheng C,
Zhang X, Ma Y, Li S, Jiang J, Zhang J, et al: Inhibition of NOB1 by
microRNA-330-5p overexpression represses cell growth of non-small
cell lung cancer. Oncol Rep. 38:2572–2580. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lin Y, Xu T, Teng H and Cui M: Anticancer
activity of NOB1-targeted shRNA combination with TRAIL in
epithelial ovarian cancer cells. Int J Clin Exp Pathol.
8:10061–10071. 2015.PubMed/NCBI
|
|
39
|
He XW, Feng T, Yin QL, Jian YW and Liu T:
NOB1 is essential for the survival of RKO colorectal cancer cells.
World J Gastroenterol. 21:868–877. 2015. View Article : Google Scholar : PubMed/NCBI
|