Introduction
Coronary heart disease (CHD) brings severe health
problems to individuals. CHD prevention is of considerable
importance for patients and medical staff. With the development of
high-throughput sequencing and bioinformatics, it has been found
that circular RNA (circRNA) enriches other types of non-coding RNA
(ncRNA). circRNA is a novel type of ncRNA which could be found
within mammalian cells, and may have been an overlooked feature of
eukaryotic gene expression and regulation (1,2).
circRNA is plentiful and stable in exosomes, and an increasing
number of its functions have been discovered with the development
of modern technology. To date, the functions of circRNA include
roles such as microRNA (miRNA) sponges, splicing or transcriptional
regulators, and agents interacting with RNA binding proteins
(RBPs), influencing the physiological process of aging, insulin
secretion, and tissue development. circRNAs also participate in the
development of atherosclerotic lesions, neurological disorders,
cardiac fibroblasts, cardiac hypertrophy, cancer, and modulating
stress and senescence responses (3–5).
They may be suitable biomarkers for cancer and coronary artery
disease diagnosis, as well as other purposes (6,7).
Collectively, it has been reported that circRNAs
might play crucial roles in fundamental life processes and serve as
novel clinical molecular markers. They might provide new insights
into the prediction, diagnosis, and treatment of diseases and the
rehabilitation of patients. It has been reported that circRNAs are
expressed in various cardiovascular diseases, such as heart failure
and pathological hypertrophy, myocardial infarction, cardiac
senescence and atherosclerosis (8). There are a great many circRNAs highly
expressed in the heart. There are a total of 15,318 cardiac
circRNAs in humans (9). The
specific circRNAs hsa_circ_0124644 and hsa_circ_0098964 are
significantly upregulated in CHD and can potentially be used as
diagnostic markers in CHD (7).
In general, we hypothesized that the development of
CHD has some connection with circRNAs, we studied the circRNA-miRNA
association for CHD, and compared the peripheral blood circRNA
profiles of large independent cohorts of CHD patients and matched
control subjects through retrospective and microarray analysis. The
results revealed that the mechanism of degree of coronary artery
stenosis in patients with CHD may involve mutually regulatory
circRNAs and miRNAs or several circRNAs and miRNAs involved in many
pathways, cell and molecular biological processes, and molecular
features.
Patients and methods
Study population
Study cohorts
In this study, a total of 40 inpatients were
recruited, between November 2016 to February 2017, from the First
Affiliated Hospital of Xinxiang Medical University (Xinxiang,
China). The age range of patients was 35–74 years. The study was
approved by the Ethics Committee of the First Affiliated Hospital
of Xinxiang Medical University. Signed written informed consents
were obtained from the patients and/or guardians. All participants
were examined by coronary angiography (CAG) to verify the presence
of CHD. Inclusion criteria: for the test group (n=20), patients
with CHD (as verified by CAG) and Gensini score (GS)>40 were
included, according to the diagnostic criteria of CHD (I25.105)
based on the International Classification of Diseases 10th edition
(ICD10); for the control group (n=20), patients with CHD and
GS<3 were included.
Exclusion criteria
Subjects were excluded according to the following
criteria: i) with chest pain caused by severe congestive heart
failure, dilated cardiomyopathy, severe neurosis, severe
arrhythmia, menopause syndrome, hyperthyroidism, gallbladder heart
syndrome, stomach and esophageal regurgitation, hiatal hernia,
aortic dissection, cervical spondylosis; ii) with liver and kidney
dysfunction; iii) suffering from any other clinically systemic
acute or chronic inflammatory disease; iv) autoimmune disease; v)
uncontrolled hypertension; vi) malignant arrhythmias or valvular
heart disease; and vii) malignancy.
Data collection
The basic information of patients came from the
hospital medical records. Data were collected concerning age, sex,
heart rate (HR), blood pressure (BP), fasting blood glucose (FBG),
total cholesterol (CHO), triglycerides (TG), apolipoprotein A1
(APOA1), apolipoprotein B (APOB), high-density lipoprotein (HDL),
low-density lipoprotein (LDL), lipoprotein (a) [LP(a)], ejection
fraction (EF), and fractional shortening (FS).
The radial CAGs were read by two experts from the
Cardiovasology based on the GS (Table
I).
 | Table I.Coronary Gensini score. |
Table I.
Coronary Gensini score.
| Degree of coronary
stenosis | Score | Lesion location | Score |
|---|
|
1–25% | 1 | Left main coronary
artery | 5.0 |
| 26–50% | 2 | Left anterior
descending branch or proximal segment of circumflex branch | 2.5 |
| 51–75% | 4 | Middle left anterior
descending branch | 1.5 |
| 76–90% | 8 | Distal segment of
left anterior descending branch | 1.0 |
| 91–99% | 16 | Left circumflex
branch middle and distal segment | 1.0 |
| Full closed | 32 | Right coronary
artery | 1.0 |
|
|
| Small branch | 0.5 |
Microarray analysis of circRNA-miRNA
Fabrication of DNA microarray
High-throughput data integration of circRNA-miRNA
provided insight into the mechanisms underlying CHD. CircRNA Array
v2 (CapitalBio, Corp., Beijing, China) was designed with four
identical arrays per slide (4×180 K format), with each array
containing probes for ~170,340 human circRNAs. Those circRNA target
sequences were all from circBase (http://www.circbase.org/), deepBase (http://rna.sysu.edu.cn/deepBase/browser.php), and the
study of You et al in 2015 (10). Each circRNA was simultaneously
detected using a long and a short probe. The circRNA array also
contained 4,974 Agilent control probes. Samples were analyzed using
Agilent Human miRNA Microarray chips (8×60 K) (release 21.0;
Agilent Technologies, Inc., Santa Clara, CA, USA). The raw data
were normalized by Quantile algorithm using GeneSpring Software
v12.6 (Agilent Technologies, Inc.). The differentially expressed
miRNAs that showed a ≥2-fold change were screened.
RNA extraction and fabrication of DNA microarray
were performed by the CapitalBio Corp.
Test group and collection of whole
blood samples
The patient blood sample collection was performed as
follows: 2 ml of blood were collected from the peripheral blood
samples of all patients after CAG and then stored in anticoagulant
vacutainers treated with EDTA. Then, 750 µl TRI pure LS Reagent
(RP001; BioTeke Corp., Beijing, China) was added to 250 µl of whole
blood samples. Pipettes were shaken repeatedly for mixing of the
whole blood samples. The blood cells were lysed for 5–10 min at
room temperature until fully disrupted and then stored in a
refrigerator at −80°C. Total RNA was extracted as soon as
possible.
RNA extraction
Total RNA containing small RNA was extracted from
blood using the TRI LS pure reagent and was purified with Qiagen
miRNeasy Mini kit (Qiagen GmbH, Hilden, Germany) according to the
manufacturers instructions. The purity and concentration of RNA
were determined by OD260/OD280 readings using
a NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies; Thermo
Fisher Scientific, Inc., Wilmington, DE, USA). RNA integrity was
determined using 1% formaldehyde denaturing gel electrophoresis.
The extracted RNAs were digested, dephosphorylated, denatured,
amplified, and labeled with Cy3-dCTP according to the manufacturers
instructions (11). The purified
RNAs were hybridized to a microarray (CircRNA Array v2; CapitalBio,
Corp.) containing 170,340 human circRNA probes and a microarray
(Agilent Human miRNA Microarray, release 21.0, 8×60 K; Agilent
Technologies, Inc.) containing 2,568 human miRNA probes.
Microarray imaging and data
analysis
The microarray data of the circRNA and miRNA array
were analyzed for data summarization, normalization, and quality
control using GeneSpring software v13.0 (Agilent Technologies,
Inc.). To select the differentially expressed genes, we used
threshold values of ≥2 (miRNA ≥1) fold change and a t-test
P<0.05. The data was Log2 transformed and median centered by
genes using the adjust data function in Cluster 3.0 software
(Cluster Software, Inc., Columbus, OH, USA). Then they were further
analyzed with hierarchical clustering with average linkage
(12). The genes were
differentially expressed in the two groups. The selected circRNAs
were then verified by quantitative polymerase chain reaction
(qPCR), which contained 15 control subjects and 15 patients with
different severities of CHD, and then the microarray tool was used
for pathway and Gene Ontology (GO) term analysis to evaluate the
differentially expressed circRNAs and any corresponding miRNAs.
Finally, we performed tree visualizations using Java TreeView
(Stanford University School of Medicine, Stanford, CA, USA).
Construction of the circRNA-miRNA
network
circRNA-miRNA network was constructed based on
miRanda v3.3 software combined with entropy values <20. These
circRNA-miRNA pairs were chosen to construct the network using the
open source bioinformatics software Cytoscape. In a network
analysis, the degree centrality is defined as the number of links
between one node and the others. A degree is the simplest and most
important measure of a gene centrality within a network determining
the relative importance (13).
Results
Research subjects
The data of the present research were collected from
a total of 40 inpatients admitted to the First Affiliated Hospital
of Xinxiang Medical University, between November 2016 to February
2017, with CHD and GS<3 (control group) or GS >40 (test
group).
General information
General information of patients were collected
including coronary GS (Table I),
age, sex, BP, FBG, CHO, TG, HDL, LDL, EF and FS (Table II). Standard values: FBG, 3.9–6.1
mmol; CHO, 0–5.2 mmol; TG, 0.7–1.7 mmol; APOA1, 1–1.76 g/l; APOB,
0.6–1.14 g/l; HDL, 0.8–1.55 mmol/l; LDL, 1.64–3.62 mmol/l; Lp(a),
0–0.3 g/l.
| Table II.Basic information of inpatients for
the expression profiles of circRNA and miRNA. |
Table II.
Basic information of inpatients for
the expression profiles of circRNA and miRNA.
| Variables | Control group | Test group |
|---|
| Age (years) | 51.6±10.44 | 61.25±9.38 |
| Sex |
|
|
| Male | 5 | 12 |
|
Female | 15 | 8 |
| HR (beats/min) | 68.6±15.8 | 69.95±20.16 |
| BP (mmHg) | 138.30±15.06 | 137.00±17.08 |
| FBG (mmol/l) | 6.17±2.87 | 6.21±2.23 |
| CHO (mmol/l) | 4.18±0.80 | 4.58±1.18 |
| TG (mmol/l) | 1.81±1.30 | 1.79±0.91 |
| APOA1 (g/l) | 1.18±0.18 | 1.09±0.24 |
| APOB (g/l) | 1.02±0.80 | 0.89±0.24 |
| HDL (mmol/l) | 1.16±0.20 | 1.09±0.35 |
| LDL (mmol/l) | 2.53±0.91 | 2.72±0.89 |
| Lp(a) (g/l) | 0.267±0.25 | 0.40±0.39 |
| AST (U/l) | 31.35±34.34 | 27.75±15.04 |
| LDH (U/l) | 67.60±60.32 | 66.80±61.40 |
| HBDH (U/l) | 144.30±30.44 | 177.95±129.54 |
| CK (U/l) | 67.35±22.02 | 103.40±67.54 |
| CK-MB (U/l) | 13.70±6.43 | 14.40±6.18 |
| EF | 63.47±3.68 | 60.42±3.68 |
| FS | 36.23±7.14 | 32.47±5.72 |
Expression profiles of circRNA and
miRNA in the peripheral blood of CHD patients
The mechanism of CHD was investigated with
high-throughput technology in the peripheral blood of CHD
patients.
Expression profiles of circRNAs
There was a significant difference between test and
control group. The results showed clear differences in the
expression profiles of circRNAs between test and control groups
(Fig. 1). Differential expression
was detected in a total of 110 circRNAs, of which 73 were
upregulated and 37 were downregulated in the CHD group (P<0.05,
FC≥2). circRNAs were found to be related to UTY, KDM5D, USP9Y,
ABCA5, SCN9A, CPNE8, and other genes (Tables III and IV). Among the 110 circRNAs, we verified
6 candidate biomarkers by qPCR, and 6 circRNAs with the highest
fold changes (P<0.01), which were taken for further analysis:
hsa_circ_0030769, hsa_circ_0079828, hsa_circ_15486-161,
hsa_circ_0122274, hsa_circ_16316-13 and hsa_circ_0140538. The
results confirmed the findings of the microarray analysis, as the
levels of these hsa_circ_16316-13 were significantly increased in
the CHD patients.
 | Figure 1.Map of cluster analysis for CHD. The
heat map is constructed by hierarchical cluster analysis. Red and
green indicate up- and downregulation during circRNA, respectively.
The heat map includes 110 gene expression levels which were
significantly altered by CHD. The genes in 10 clusters, denoted as
group 1 (a9, a14, H2, H3, H4) and group 2 (a10, a15, Z8, Z10, Z11),
were 73 upregulated and 37 downregulated. CHD, coronary heart
disease; circRNA, circular RNA. |
| Table III.Expression profiles of circRNAs that
were upregulated (P<0.05, FC≥2). |
Table III.
Expression profiles of circRNAs that
were upregulated (P<0.05, FC≥2).
| No. | Probe name | P-value | FC (abs) | Gene symbol | Regulation | Chromosome | Strand | Start | End |
|---|
| 1 |
hsa_circ_0140759 | 0.031 | 45.35329 | UTY | Up | chrY | − | 15466882 | 15481229 |
| 2 |
hsa_circ_16316-13 | 0.046 | 28.26892 | UTY | Up | chrY | − | 15447442 | 15478273 |
| 3 |
hsa_circ_0140760 | 0.045 | 28.18893 | UTY | Up | chrY | − | 15467172 | 15471765 |
| 4 |
hsa_circ_0140758 | 0.047 | 25.23608 | UTY | Up | chrY | − | 15447442 | 15448215 |
| 5 |
hsa_circ_16316-11 | 0.039 | 16.33733 | UTY | Up | chrY | − | 15471646 | 15471866 |
| 6 |
hsa_circ_0140781 | 0.040 | 12.63422 | KDM5D | Up | chrY | − | 21901413 | 21903743 |
| 7 |
hsa_circ_0140736 | 0.043 | 12.46352 | USP9Y | Up | chrY | + | 14821320 | 14885859 |
| 8 |
hsa_circ_16316-9 | 0.026 | 10.90928 | UTY | Up | chrY | − | 15435434 | 15438230 |
| 9 |
hsa_circ_0140746 | 0.033 | 10.40101 | USP9Y | Up | chrY | + | 14870435 | 14885859 |
| 10 |
hsa_circ_0009024 | 0.020 |
9.696754 | − | Up | chrY | + | 21749095 | 21749393 |
| 11 |
hsa_circ_16316-12 | 0.017 |
9.500227 | UTY | Up | chrY | − | 15435434 | 15448215 |
| 12 |
hsa_circ_0007907 | 0.024 |
8.476297 | ZFY | Up | chrY | + | 2829114 | 2829687 |
| 13 |
hsa_circ_0140732 | 0.031 |
7.912563 | USP9Y | Up | chrY | + | 14813938 | 14834120 |
| … | …… |
|
|
|
|
|
|
|
|
| 73 |
hsa_circ_0099619 | 0.003 |
2.00747 | NEDD1 | Up | chr12 | + | 97264281 | 97303673 |
| Table IV.Expression profiles of circRNAs that
were downregulated (P<0.05, FC≥2). |
Table IV.
Expression profiles of circRNAs that
were downregulated (P<0.05, FC≥2).
| No. | Probe name | P-value | FC (abs) | Gene symbol | Regulation | Chromosome | Strand | Start | End |
|---|
| 1 |
hsa_circ_0091074 | 0.039 | 20.66278 | − | Down | chrX | − | 73048902 | 73051109 |
| 2 |
hsa_circ_0140538 | 0.037 | 15.17814 | − | Down | chrX | − | 73045949 | 73051109 |
| 3 |
hsa_circ_0091073 | 0.044 | 14.23409 | − | Down | chrX | − | 73040494 | 73051109 |
| 4 |
hsa_circ_0140539 | 0.046 | 11.15955 | − | Down | chrX | − | 73045949 | 73057338 |
| 5 |
hsa_circ_16166-3 | 0.046 |
8.908921 | − | Down | chrX | − | 73050900 | 73053209 |
| 6 |
hsa_circ_16166-1 | 0.049 |
8.020764 | − | Down | chrX | − | 73050900 | 73057338 |
| 7 |
hsa_circ_0140536 | 0.034 |
5.790904 | ABCA5 | Down | chrX | − | 73044087 | 73044570 |
| 8 |
hsa_circ_0107597 | 0.028 |
4.049471 | − | Down | chr17 | − | 67270099 | 67305564 |
| 9 |
hsa_circ_0140541 | 0.031 |
3.919625 | SCN9A | Down | chrX | − | 73046801 | 73046954 |
| 10 |
hsa_circ_0117953 | 0.002 |
2.986698 | CPNE8 | Down | chr2 | − |
1.67×108 |
1.67×108 |
| 11 |
hsa_circ_0098463 | 0.048 | 2.7141 | DNAH14 | Down | chr12 | − | 39117573 | 39156018 |
| 12 |
hsa_circ_770-66 | 0.042 |
2.692528 | VWA8 | Down | chr1 | + |
2.25×108 |
2.25×108 |
| 13 |
hsa_circ_0030090 | 0.045 |
2.620312 | GPSM2 | Down | chr13 | − | 42259182 | 42335340 |
|
| …… | …… | …… |
| …… |
|
|
|
|
| 37 |
hsa_circ_0079838 | 0.030 |
2.000309 | DPY19L1 | Down | chr7 | − | 34989408 | 35013217 |
Expression profiles of miRNAs
The results showed significant differences in the
expression profiles of miRNAs between test and control group.
Differential expression was detected in a total of 18 miRNAs
(P<0.05, FC≥1), of which 13 were upregulated and 5 were
downregulated in the CHD group (Table
V). We verified 2 candidate miRNAs by qPCR: hsa-let-7c-5p and
hsa-miR-101-5p were significantly expressed in the CHD
patients.
| Table V.Expression profiles of miRNAs
(P<0.05, FC≥1). |
Table V.
Expression profiles of miRNAs
(P<0.05, FC≥1).
| No. | Systematic
name | P-value | FC (abs) | Regulation |
|---|
| 1 |
hsa-miR-148a-3p | 0.001001 | 1.411098 | Up |
| 2 | hsa-miR-1260a | 0.007008 | 1.454044 | Up |
| 3 | hsa-miR-29c-3p | 0.008573 | 1.333568 | Up |
| 4 | hsa-miR-101-5p | 0.015351 | 1.268211 | Up |
| 5 | hsa-miR-545-3p | 0.020073 | 1.175692 | Up |
| 6 | hsa-miR-5100 | 0.020456 | 2.086728 | Up |
| 7 | hsa-miR-194-5p | 0.023195 | 1.715863 | Up |
| 8 | hsa-miR-215-5p | 0.02325 | 1.404127 | Up |
| 9 | hsa-miR-4443 | 0.035173 | 1.384234 | Up |
| 10 | hsa-miR-19a-3p | 0.039729 | 1.331585 | Up |
| 11 | hsa-miR-32-5p | 0.042593 | 1.549326 | Up |
| 12 |
hsa-miR-103a-3p | 0.043258 | 1.156287 | Up |
| 13 | hsa-miR-424-5p | 0.044219 | 1.279992 | Up |
| 14 | hsa-miR-3196 | 0.009454 | 1.148938 | Down |
| 15 | hsa-miR-3651 | 0.010955 | 1.485764 | Down |
| 16 |
hsa-miR-1249-3p | 0.037988 | 1.325854 | Down |
| 17 | hsa-miR-4281 | 0.042548 | 1.095865 | Down |
| 18 |
hsa-miR-1228-3p | 0.042683 | 1.518054 | Down |
circRNA cluster analysis
Cluster analysis was performed on the circRNA
between test and control group. Because there were not sufficient
data concerning miRNA, no cluster analysis was performed as shown
in Fig. 1.
Coexpression of circRNA-miRNA
Examination of the coexpression of circRNA-miRNA to
the GS of CHD showed that a large number of circRNAs take part in
the CHD process and may influence a similarly large number of
miRNAs, and circRNAs participate in the regulation of >100
miRNAs, such as upregulated hsa_circ_16316-13, hsa_circ_0140760,
hsa_circ_0140748; and downregulated hsa_circ_0140538,
hsa_circ_0091073, hsa_circ_0140539 (Tables VI–VIII, Fig.
2).
| Table VI.Coexpression of circRNA-miRNA
(P<0.05, FC>2). |
Table VI.
Coexpression of circRNA-miRNA
(P<0.05, FC>2).
| No. | Probe name | miRNA capable of
bonding to ≥1 circRNA | miRNA capable of
bonding to ≥2 circRNA | Regulation | Gene symbol | Strand | Start | End |
|---|
| 1 |
hsa_circ_16316-13 | 100 | 100 | Up | UTY | − | 15447442 | 15478273 |
| 2 |
hsa_circ_0140760 | 100 | 100 | Up | UTY | − | 15467172 | 15471765 |
| 3 |
hsa_circ_0140758 | 100 | 100 | Up | UTY | − | 15447442 | 15448215 |
| 4 |
hsa_circ_0140736 | 100 | 100 | Up | USP9Y | + | 14821320 | 14885859 |
| 5 |
hsa_circ_16316-12 | 100 | 100 | Up | UTY | − | 15435434 | 15448215 |
| 6 |
hsa_circ_0140732 | 100 | 100 | Up | USP9Y | + | 14813938 | 14834120 |
| 7 |
hsa_circ_0140783 | 100 | 100 | Up | − | − | 22669237 | 22683186 |
| 8 |
hsa_circ_0140757 | 100 | 100 | Up | UTY | − | 15435434 | 15472408 |
| 9 |
hsa_circ_16316-14 | 100 | 100 | Up | UTY | − | 15435434 | 15478273 |
| 10 |
hsa_circ_0140756 | 100 | 100 | Up | UTY | − | 15409586 | 15438230 |
| 11 |
hsa_circ_0140733 | 100 | 100 | Up | USP9Y | + | 14813938 | 14870572 |
| 12 |
hsa_circ_0140779 | 100 | 100 | Up | − | + | 21749095 | 21752658 |
| 13 |
hsa_circ_0092240 | 100 | 100 | Up | USP9Y | + | 14832521 | 14905134 |
| 14 |
hsa_circ_0092257 | 100 | 83 | Up | USP9Y | + | 14954988 | 14969586 |
| … | …… | …… | …… |
|
|
|
|
|
| 73 |
hsa_circ_8216-2 |
1 |
0 | Up | ZFY | + | 2821949 | 2822038 |
| Table VIII.Top 8 circRNAs-miRNA. |
Table VIII.
Top 8 circRNAs-miRNA.
| No. | Probe name | P-value | miRNA no. | Regulation |
|---|
| 1 |
hsa_circ_0140759 | 0.031 |
6 | Up |
| 2 |
hsa_circ_16316-13 | 0.046 | 17 | Up |
| 3 |
hsa_circ_0140760 | 0.045 | 66 | Up |
| 4 |
hsa_circ_0140758 | 0.047 |
3 | Up |
| 5 |
hsa_circ_0140736 | 0.043 |
7 | Up |
| 6 |
hsa_circ_0140781 | 0.040 |
1 | Up |
| 7 |
hsa_circ_0140538 | 0.036 | 100 | Down |
| 8 |
hsa_circ_0091073 | 0.044 | 100 | Down |
Enrichment analysis
Enrichment analysis of the data from the expression
profiles of circRNAs (P<0.05, FC≥2) indicated statistically
significant difference between test and control group using NHGRI
GWAS Catalog, GO, and Reactome.
Disease enrichment analysis
The analysis of disease enrichment was performed
using the NHGRI GWAS Catalog. The results showed that circRNA is
involved in a total of 59 diseases and significantly involved in 15
diseases (P<0.05; Fig. 3).
 | Figure 3.Significantly enriched
NHGRI_GWAS_Catalog disease terms for CHD. The significantly
enriched NHGRI_GWAS_Catalog disease terms include: attention
deficit hyperactivity disorder, cardiovascular heart disease in
diabetics, response to cholinesterase inhibitors in Alzheimer's
disease, complement C3 and C4 levels, fasting plasma glucose,
electroencephalographic traits in alcoholism, age-related macular
degeneration, optic nerve measurement (disc area), F-cell
distribution, metabolite levels (HVA/5-HIAA ratio), non-alcoholic
fatty liver disease histology (other), age-related macular
degeneration (GA), gambling, phosphorus levels, Paget's disease.
CHD, coronary heart disease. |
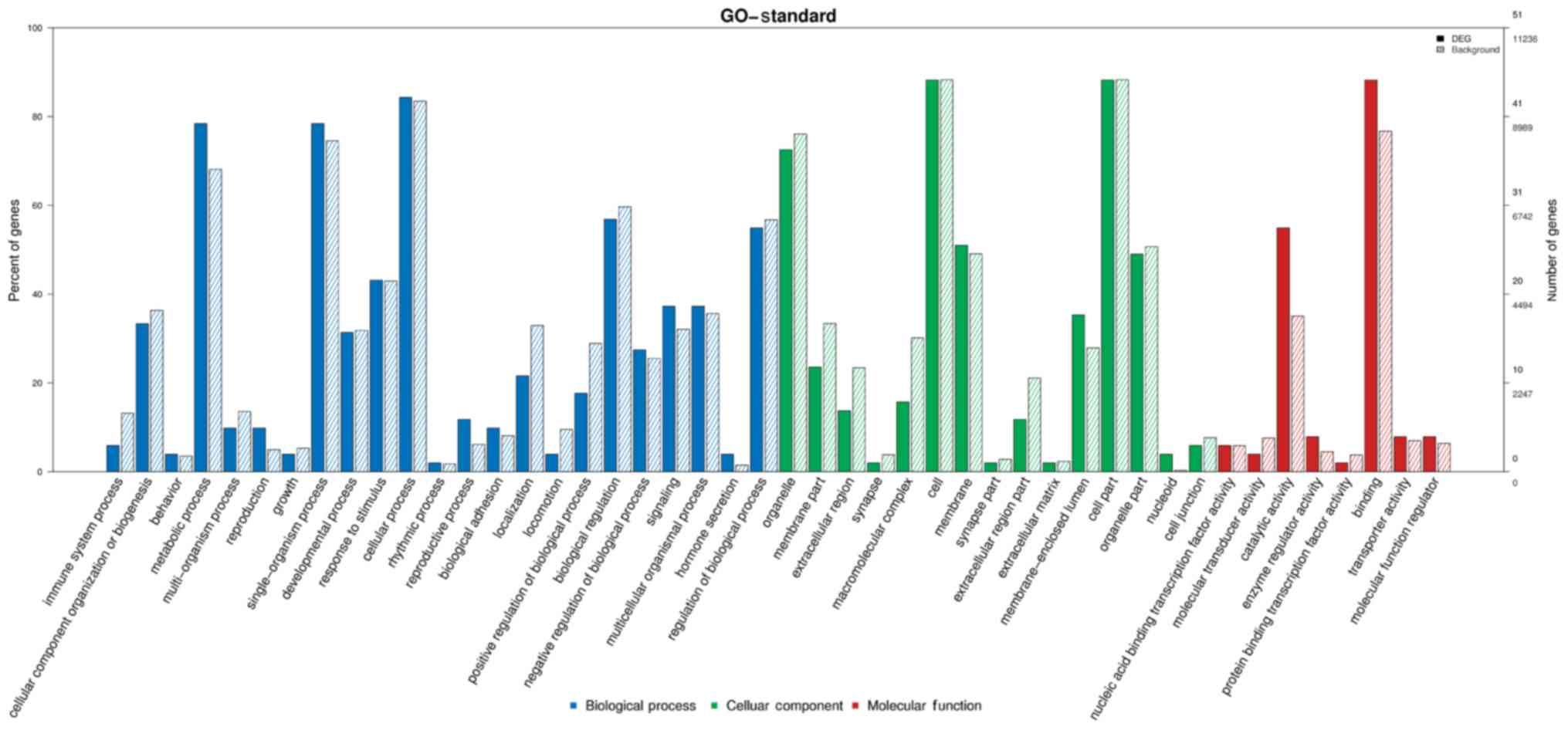
GO enrichment analysis
GO is divided into biological processes,
cellular components, and molecular functions (Figs. 4 and 5). i) The biological processes associated
with circRNA tended to relate more to content, and the results
showed circRNA to be involved in a total of 1,254 terms and
significantly involved in 136 biological processes (P<0.05). ii)
Results showed circRNA to be involved in a total of 269 terms and
significantly involved in 89 molecular functions (P<0.05). iii)
The cellular component category contains a total of 171 terms, 21
of which were found to be significant (P<0.05).
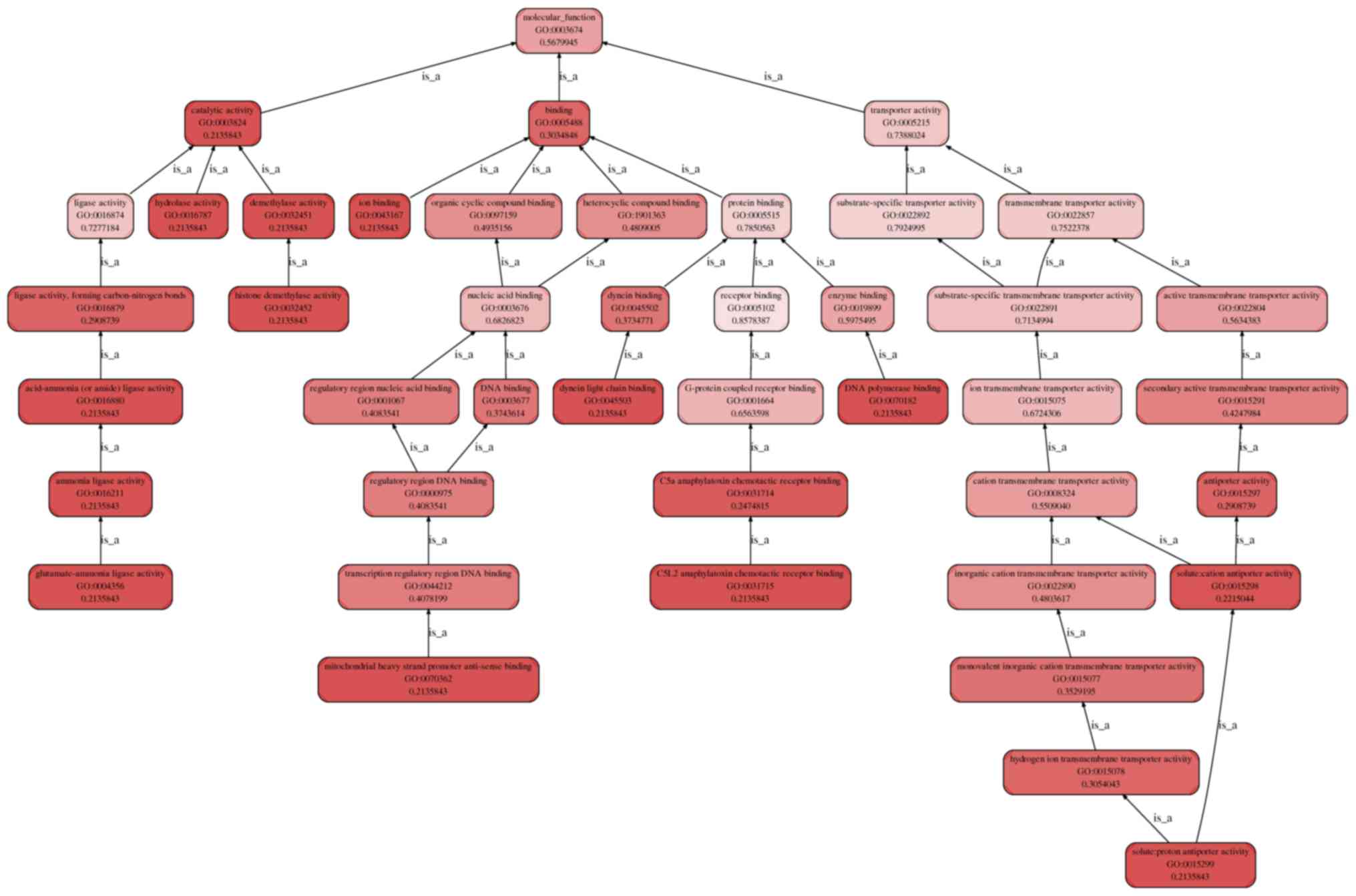 | Figure 5.
Enrichment.Go.hierarchy.molecular_function for CHD. The top 10
molecular functions are: DNA polymerase binding, ion binding,
solute:proton antiporter activity, hydrolase activity, catalytic
activity, histone demethylase activity, demethylase activity,
dynein light chain binding, mitochondrial heavy strand promoter
anti-sense binding, glutamate-ammonia ligase activity. The
molecular function category includes four fields: the
glutamate-ammonia ligase activity, mitochondrial heavy strand
promoter anti-sense binding, C5L2 anaphylatoxin chemotactic
receptor binding, and solute:proton antiporter activity. These
influence the overall catalytic, binding, and transporter activity,
finally changing the molecular function. CHD, coronary heart
disease. |
Reactome pathway
By analyzing data of 110 statistical significant
differences (P<0.05, FC≥2) on the Reactome pathway analysis of
circRNAs from the experimental and control group, we found that it
has 172 pathways significantly involved in 30 (P<0.05; Fig. 6).
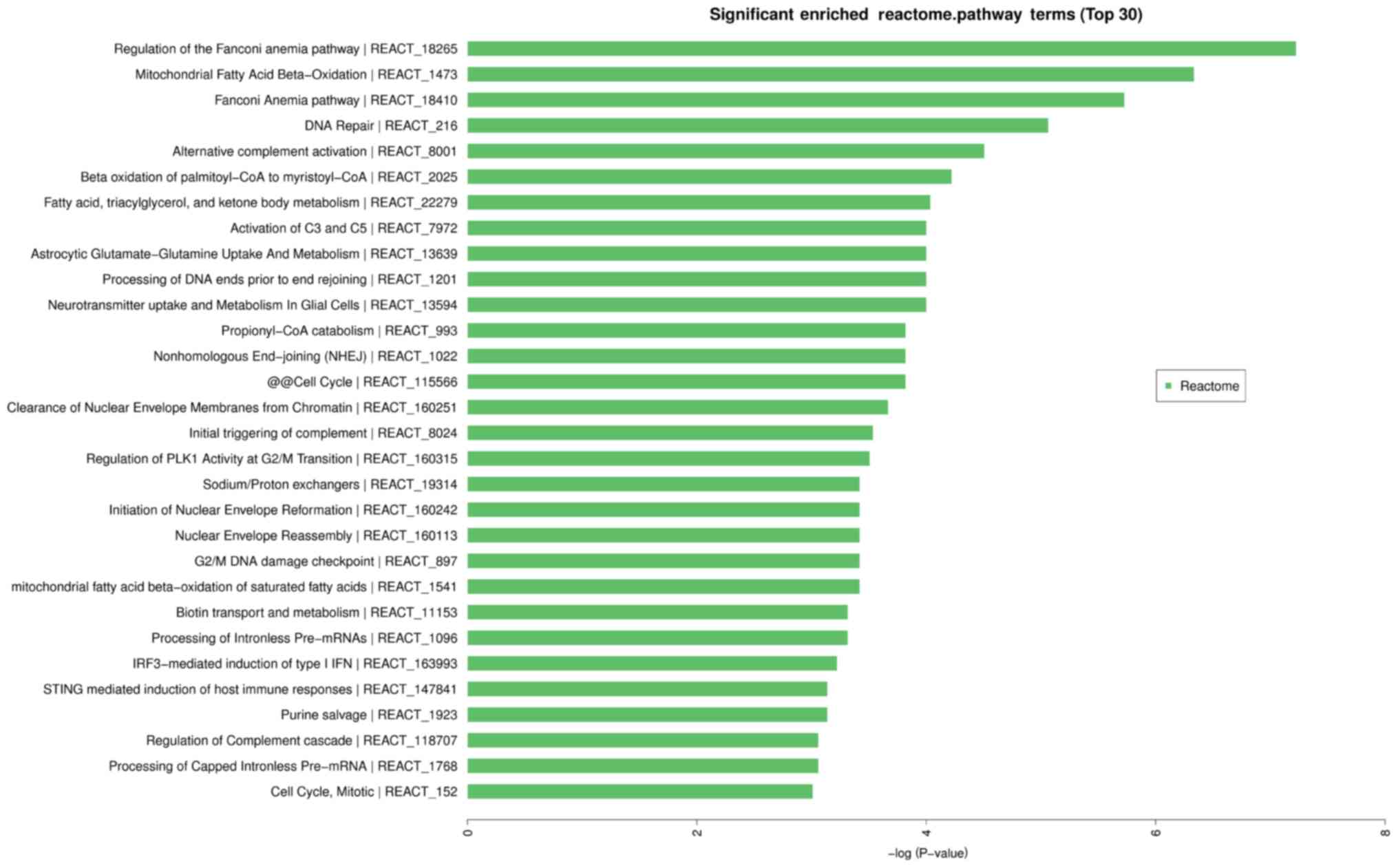 | Figure 6.Enriched Reactome pathway terms for
CHD. The top 10 Reactome pathway terms are: regulation of the
Fanconi anemia pathway, mitochondrial fatty acid β-oxidation,
Fanconi anemia pathway, DNA repair, alternative complement
activation, β oxidation of palmitoyl-CoA to myristoyl-CoA, fatty
acid triacylglycerol and ketone body metabolism, neurotransmitter
uptake and metabolism in glial cells, processing of DNA ends prior
to end rejoining, astrocytic glutamate-glutamine uptake and
metabolism. CHD, coronary heart disease. |
Discussion
CHD still represents the leading cause of mortality
and morbidity worldwide, and the underlying disease,
atherosclerosis, as a common feature of the CHD, is initiated and
propagated by continuous damage, as a result of a variety of
factors. In the regulation of health and disease, circRNAs act as
urgent effectors by miRNA sponges, splicing or transcriptional
regulators, and agents interacting with RBPs.
Long non-coding RNA (lncRNA) works and controls the
atherosclerosis process (14).
There is growing evidence that non-coding RNA (circRNA and miRNA)
is involved in the development and progression of CHD. The
association of circRNA with CHD began with the study of antisense
non-coding RNA in the INK4 locus, ANRIL. In 2010, Burd et al
found that circANRIL expression by human INK4/ARF transcriptional
regulation is associated the atherosclerotic risk (15). The same year, it was confirmed that
ANRIL expression is associated with atherosclerosis risk at
chromosome 9p21 (16). In 2016,
Holdt et al found circANRIL, a prototype of a circRNA
regulating ribosome biogenesis and conferring atheroprotection, and
suggested that circANRIL remains a potential therapeutic target for
the treatment of atherosclerosis (17). Studies have illustrated that
circRNA can contribute to atherosclerosis development and
progression. In addition, circRNA hsa_circ_0124644 can be used for
the diagnosis of coronary artery disease (7). A circRNA, termed heart-related
circRNA (HRCR), has been shown to act as an endogenous miR-223
sponge to inhibit cardiac hypertrophy and heart failure (18). circRNAs are dynamically expressed
in a human induced pluripotent stem cell-derived cardiomyocytes
model of cardiac development and stress response. circRNAs such as
circSLC8A1, circCACNA1D, circSPHKAP and circALPK2 may serve as
biomarkers of cardiomyocytes (CMs), and circSLC8A1 is increased
abnormally in heart tissues of patients with dilated cardiomyopathy
(19,20). Through microarray analysis, the
results of this study revealed that the mechanism of CHD may be
related to the expression of circRNA and miRNA and the
co-expression of circRNA-miRNA, as well as circ_0091074,
hsa_circ_0140538, and hsa_circ_0140539 may act on hsa-miR-101-5p,
hsa-miR-148a-3p, and hsa-miR-1260a and may be involved in CHD.
The results of enrichment analysis of the data from
the expression profiles of circRNAs indicated statistically
significant difference between test and control group using NHGRI
GWAS Catalog, GO, and Reactome. For example, cardiovascular heart
disease in diabetics is included in the result analysis of disease
enrichment. The result of enrichment analysis of GO showed that
circRNA may be additionally involved in other than the biological
process, molecular function and cellular component. It is important
that the expression profile changes of circRNA are involved in
lipid metabolism pathway, for example, mitochondrial fatty acid
β-oxidation, β oxidation of palmitoyl-CoA to myristoyl-CoA, fatty
acid-triacylglycerol-and ketone body metabolism, mitochondrial
fatty acid β-oxidation of saturated fatty acids.
The mechanisms involved in cardiovascular aging, and
the potential for targeting novel pathways implicated in
endothelial dysfunction, mitochondrial oxidative stress, chromatin
remodeling and genomic instability have been reported (19). miRNAs can regulate protein
expression, and thus constitute potential miRNAs as therapeutic
targets in cardiac and vascular disease, and can be used as novel
biomarkers (20,21). Likewise, it has been reported that
cardiovascular diseases are closely related to the expression of
circRNA, and they point to a high abundance of specific
cardiac-expressed circRNA (9). The
key cardiac genes include TTN, RYR2 and DMD in this study. In
addition, it has been reported that lncRNA MALAT1, a highly
abundant and conserved imprinted gene, has been implicated in many
cardiovascular diseases; MALAT1 positively regulates the expression
of Smad4 through sponging miR-204, and promotes osteogenic
differentiation of human aortic valve interstitial cells (22).
We observed significant positive associations
between circRNA and miRNA and the co-expression of circRNA-miRNA,
but the causal relevance of these associations remains uncertain.
In the study of CHD, little research has been conducted on
circRNAs, and especially the studies on the circRNA of coronary
artery atherosclerosis are very few. Atherosclerosis is the main
pathological basis of acute coronary syndrome. Thus, we aim to
determine the positive associations between circRNA and CHD in a
larger sample, and the positive co-expression between circRNA and
miRNA in future investigations. Specifically, cell lines should be
used to show that circRNA-miRNA mutually regulate the mechanism of
mRNA, and a model animal should be established to confirm that
circRNA/miRNA regulate mRNA and affect the formation and
development of atherosclerosis.
In conclusion, the mechanism underlying CHD may be
closely related to up- or downregulated circRNA and miRNA and
co-expression of circRNA-miRNA specifically involved regulate
multiple pathways and multiple cellular and molecular biological
processes.
Acknowledgements
Not applicable.
Funding
This study was supported by the Foundation of Henan
Educational Committee (grant nos. 18A320005 and 19A360032), and
Beijing Medical and Health Foundation (grant no. B17246-045).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors contributions
FLi was a major contributor in writing the
manuscript, and responsible for conducting the experiments. GZhao
contributed greatly in the designing of the study and was in main
charge of the reviewing. ZC and XW acquired the data. FLv and YZ
participated in the analysis and interpretation of the data. XY and
WL designed the methods of this study. RC and JL were responsible
for the follow-up of the patients. ML and GZhang participated in
the design of the study. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
the First Affiliated Hospital of Xinxiang Medical University
(Xinxiang, China). Signed written informed consents were obtained
from the patients and/or guardians.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jeck WR and Sharpless NE: Detecting and
characterizing circular RNAs. Nat Biotechnol. 32:453–461. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Salzman J: Circular RNA Expression: Its
potential regulation and function. Trends Genet. 32:309–316. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Qu S, Zhong Y, Shang R, Zhang X, Song W,
Kjems J and Li H: The emerging landscape of circular RNA in life
processes. RNA Biol. 14:992–999. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tang CM, Zhang M, Huang L, Hu ZQ, Zhu JN,
Xiao Z, Zhang Z, Lin QX, Zheng XL, Yang M, et al: CircRNA_000203
enhances the expression of fibrosis-associated genes by
derepressing targets of miR-26b-5p, Col1a2 and CTGF, in cardiac
fibroblasts. Sci Rep. 7:403422017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Du WW, Yang W, Chen Y, Wu ZK, Foster FS,
Yang Z, Li X and Yang BB: Foxo3 circular RNA promotes cardiac
senescence by modulating multiple factors associated with stress
and senescence responses. Eur Heart J. 38:1402–1412.
2017.PubMed/NCBI
|
|
6
|
Li Y, Zheng Q, Bao C, Li S, Guo W, Zhao J,
Chen D, Gu J, He X and Huang S: Circular RNA is enriched and stable
in exosomes: A promising biomarker for cancer diagnosis. Cell Res.
25:981–984. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhao Z, Li X, Gao C, Jian D, Hao P, Rao L
and Li M: Peripheral blood circular RNA hsa_circ_0124644 can be
used as a diagnostic biomarker of coronary artery disease. Sci Rep.
7:399182017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fan X, Weng X, Zhao Y, Chen W, Gan T and
Xu D: Circular RNAs in cardiovascular disease: An overview. BioMed
Res Int. 2017:51357812017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tan WL, Lim BT, Anene-Nzelu CG,
Ackers-Johnson M, Dashi A, See K, Tiang Z, Lee DP, Chua WW, Luu TD,
et al: A landscape of circular RNA expression in the human heart.
Cardiovasc Res. 113:298–309. 2017.PubMed/NCBI
|
|
10
|
You X, Vlatkovic I, Babic A, Will T,
Epstein I, Tushev G, Akbalik G, Wang M, Glock C, Quedenau C, et al:
Neural circular RNAs are derived from synaptic genes and regulated
by development and plasticity. Nat Neurosci. 18:603–610. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Patterson TA, Lobenhofer EK,
Fulmer-Smentek SB, Collins PJ, Chu TM, Bao W, Fang H, Kawasaki ES,
Hager J, Tikhonova IR, et al: Performance comparison of one-color
and two-color platforms within the MicroArray Quality Control
(MAQC) project. Nat Biotechnol. 24:1140–1150. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Eisen MB, Spellman PT, Brown PO and
Botstein D: Cluster analysis and display of genome-wide expression
patterns. Proc Natl Acad Sci USA. 95:14863–14868. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Barabási AL and Oltvai ZN: Network
biology: Understanding the cell's functional organization. Nat Rev
Genet. 5:101–113. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu Y, Zheng L, Wang Q and Hu YW: Emerging
roles and mechanisms of long noncoding RNAs in atherosclerosis. Int
J Cardiol. 228:570–582. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Burd CE, Jeck WR, Liu Y, Sanoff HK, Wang Z
and Sharpless NE: Expression of linear and novel circular forms of
an INK4/ARF-associated non-coding RNA correlates with
atherosclerosis risk. PLoS Genet. 6:e10012332010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Holdt LM, Beutner F, Scholz M, Gielen S,
Gäbel G, Bergert H, Schuler G, Thiery J and Teupser D: ANRIL
expression is associated with atherosclerosis risk at chromosome
9p21. Arterioscler Thromb Vasc Biol. 30:620–627. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Holdt LM, Stahringer A, Sass K, Pichler G,
Kulak NA, Wilfert W, Kohlmaier A, Herbst A, Northoff BH, Nicolaou
A, et al: Circular non-coding RNA ANRIL modulates ribosomal RNA
maturation and atherosclerosis in humans. Nat Commun. 7:124292016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang K, Long B, Liu F, Wang JX, Liu CY,
Zhao B, Zhou LY, Sun T, Wang M, Yu T, et al: A circular RNA
protects the heart from pathological hypertrophy and heart failure
by targeting miR-223. Eur Heart J. 37:2602–2611. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Siede D, Rapti K, Gorska AA, Katus HA,
Altmüller J, Boeckel JN, Meder B, Maack C, Völkers M, Müller OJ, et
al: Identification of circular RNAs with host gene-independent
expression in human model systems for cardiac differentiation and
disease. J Mol Cell Cardiol. 109:48–56. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lei W, Feng T, Fang X, Yu Y, Yang J, Zhao
ZA, Liu J, Shen Z, Deng W and Hu S: Signature of circular RNAs in
human induced pluripotent stem cells and derived cardiomyocytes.
Stem Cell Res Ther. 9:562018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Vegter EL, Schmitter D, Hagemeijer Y,
Ovchinnikova ES, van der Harst P, Teerlink JR, O'Connor CM, Metra
M, Davison BA, Bloomfield D, et al: Use of biomarkers to establish
potential role and function of circulating microRNAs in acute heart
failure. Int J Cardiol. 224:231–239. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xiao X, Zhou T, Guo S, Guo C, Zhang Q,
Dong N and Wang Y: LncRNA MALAT1 sponges miR-204 to promote
osteoblast differentiation of human aortic valve interstitial cells
through up-regulating Smad4. Int J Cardiol. 243:404–412. 2017.
View Article : Google Scholar : PubMed/NCBI
|