Introduction
Recruitment of inflammatory cells to the vessel wall
from the circulation and their transendothelial migration
constitutes one of the earliest cellular events in the development
of atherosclerotic lesions (1).
Oxidized low-density lipoprotein (oxLDL), a factor known to affect
vessel wall integrity (2),
contributes to the development of atherosclerosis through a number
of mechanisms. One mechanism is the stimulation of endothelial cell
activation and the subsequent induction of expression of a series
of adhesion molecules, including intercellular adhesion molecule-1
(ICAM-1) and vascular cell adhesion molecule-1 (VCAM-1) (3–5).
These adhesion molecules predominantly mediate adhesion of
circulating mononuclear cells to the endothelium and facilitate
their migration into the subendothelial space, thus contributing to
the extravasation of mononuclear cells in the vascular wall, an
initial step in the development of atherosclerosis (6,7).
Moreover, cell-cell interactions between monocytes and endothelial
cells further promote the development of atherosclerosis by
increasing the production of specific cytokines, including tumor
necrosis factor α (TNF-α) and monocyte chemotactic protein-1
(7,8). Adhesion molecules are important in
the initiation and progression of atherosclerotic plaques and the
suppression of secretion or expression of these adhesion molecules
may be of clinical significance. Moreover, immunopharmacological
inhibition of ICAM-1 and VCAM-1 has been demonstrated to inhibit
the development of fatty streaks (9).
Berberine is a botanical alkaloid isolated from
medicinal herbs, including Rhizoma coptidis (Huanglian) and
Cortex phellodendri (Huangbai) (10). It has been extensively used in
traditional Chinese medicine to treat infectious diarrhea (11). Previous studies have reported that
berberine exhibits protective effects against cardiovascular
diseases. For instance, berberine reduces plasma cholesterol
(12) and glucose levels (13) and inhibits the expression of
pro-inflammatory factors, including TNF-α (14). Berberine has been identified to
exert antiatherogenic effects by inhibiting the expression of
matrix metallopeptidase-9 (MMP-9) and extracellular matrix
metalloproteinase inducer (EMMPRIN) (15). In addition, berberine has been
found to inhibit generation of reactive oxygen species and prevent
oxLDL-induced endothelial cellular apoptosis (16), demonstrating a protective effect on
oxLDL-stimulated endothelial cells. However, the effect of
berberine on the extent of monocyte adhesion to endothelial cells
and the potential mechanism involved in this process remains
unclear.
In the present study, the effect of berberine on the
oxLDL-induced adhesion of monocytes to human umbilical vein
endothelial cells (HUVECs) was investigated. In addition, the
expression of adhesion molecules in oxLDL-stimulated HUVECs was
analyzed to clarify the mechanism by which berberine mediates this
process.
Materials and methods
Reagents
RPMI-1640 medium, fetal bovine serum (FBS) and
penicillin/streptomycin (pen/strep, 10,000 U/ml each) were obtained
from Invitrogen Life Technologies (Carlsbad, CA, USA). Phorbol
12-myristate 13-acetate was purchased from Calbiochem (San Diego,
CA, USA). Methyl sulfoxide (DMSO) and berberine were acquired from
Sigma-Aldrich (St. Louis, MO, USA). oxLDL was purchased from
Beijing Union Medical Biochemistry Room (Beijing, China).
Endothelial cell medium (ECM) and human vascular endothelial growth
factor (VEGF) were purchased from ScienCell (Carlsbad, CA, USA).
The 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide)
(MTT) assay was obtained from Roche Diagnostics (Mannheim,
Germany). TRIzol reagent for RNA isolation was purchased from
Invitrogen Life Technologies. Omniscript reverse transcriptase for
first-strand cDNA synthesis was obtained from Qiagen (Shanghai,
China). Taq DNA polymerase was from New England Biolabs
(Ipswich, MA, USA). Green fluorescent protein (GFP) adenovirus
(Ad5/F35-GFP) was from Vector Gene Technology Company Ltd.
(Beijing, China). VCAM-1 and ICAM-1 ELISA Kits were obtained from
R&D Systems (Minneapolis, MN, USA).
Cell culture and treatments
The human acute monocytic leukemia cell line, THP-1,
was purchased from American Type Culture Collection (Rockville, MD,
USA) and cultured in RPMI-1640 medium containing 10% FBS, 10 mM
4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES;
Sigma-Aldrich) and 1% pen/strep solution at a density of
5×105 cells/ml in a 5% CO2 incubator. GFP-Ad
was added to the THP-1 cells for 24 h to obtain GFP expression.
HUVECs were purchased from ScienCell (no. 8000) and cultured in ECM
containing VEGF (3 ng/ml), 10% FBS, 10 mM HEPES and 1% pen/strep
solution at a density of 1×104 cells/ml in a 5%
CO2 incubator. The culture was then passaged 2–6 times.
Growth medium was changed every other day until 80% confluence was
achieved. Following this, the culture medium was replaced with ECM
containing 0.4% FBS for cell starvation for 6 h. Cells were
pretreated with berberine (5, 25 and 50 μM) for 2 h, followed by
oxLDL stimulation (100 μg/ml) for 6 h. Cells were then cocultured
with GFP-Ad-infected-THP-1 cells for 20 min to determine the
adhesive capacity of the monocytic to endothelial cells. Unattached
cells were removed and collected to calculate the percentage of
cells expressing GFP.
Cell viability assay using MTT
Following the indicated treatments, cells were
incubated with 0.5 mg/ml MTT in culture medium for 4 h. The blue
formazan crystals of viable cells were then dissolved in DMSO and
spectrophotometrically analyzed at 570 nm.
Flow cytometry
Flow cytometry was performed according to the
manufacturer’s instructions. Mean fluorescence intensity of GFP
expression in THP-1 cells cocultured with HUVECs was analyzed on a
BD FACSCalibur flow cytometer. The mean intensity of untreated
cells was considered as 100%. Changes in GFP levels in the cells
following berberine treatment were evaluated and standardized
against the untreated cells.
GFP-Ad-infected THP-1 cells and
endothelial cell-monocytic cell adhesion assay
Cells were cultured in 24-well plates at a density
of 2×105 cells/ml in a 5% CO2 incubator and
added to a 106 GFP-Ad (2, 4.5 or 6 μl) titer for 24 h to
determine the appropriate amount of GFP-Ad-infected THP-1 cell
virus (multiplicity of infection, MOI). The percentage of
GFP-expressing cells was then determined using flow cytometry.
GFP-labeled THP-1 cells (1×106) were then added to each
HUVEC-containing well and the incubation was continued for 20 min.
Non-adherent cells were removed using two gentle washes with
phosphate-buffered saline (PBS) and the percentage of GFP mean
fluorescence intensity of the cells, which indicates the number of
bound THP-1 cells, was determined using flow cytometry.
RNA isolation and reverse
transcription-polymerase chain reaction (RT-PCR)
Total RNA was extracted from oxLDL-stimulated HUVECs
using TRIzol reagent according to the manufacturer’s instructions.
Total RNA (2 μg) was reverse transcribed into cDNA using a random
primer and the resultant cDNA was amplified through PCR using the
following primers: human VCAM-1 (575 bp), 5′-ATGCCT
GGGAAGATGGTCGTGA-3′ (sense) and 5′-TGGAGCTGG TAGACCCTCGCTG-3′
(antisense); human ICAM-1 (370 bp), 5′-TGCCACCAATATGGGAAGGC-3′
(sense) and 5′-CCG AGCTCAAGTGTCTAAAG-3′ (antisense); and GAPDH,
(519 bp), 5′-GGTGAAGGTCGGAGTCAACGG-3′ (sense) and
5′-GTCATGAGTCCTTCCACGAT-3′ (antisense). All gels were detected
using the Tanon-4100 digital Gis image system (Beijing, China) and
densitometric analysis was performed using Quantity One (Bio-Rad,
Hercules, CA, USA) to scan the signals.
Cell enzyme-linked immunosorbent assay
(ELISA)
Adhesion molecule expression in HUVECs was
determined by cell ELISA. HUVECs in 96-well plates were pretreated
with berberine (5, 25 and 50 μM) for 2 h and subsequently
stimulated for 6 h with oxLDL (100 μg/ml). HUVECs were then washed
twice with PBS and fixed with 0.025% glutaraldehyde for 10 min.
Following this, cells were incubated with 1 μg/ml monoclonal
antibody against VCAM-1 or ICAM-1 or with 1 μg/ml non-specific
mouse immunoglobulin G1 (IgG1; Sigma-Aldrich) in PBS containing 1%
bovine serum albumin (BSA) at room temperature for 1 h. The latter
was selected as the control. Following incubation, wells were
washed with PBS containing 0.05% Tween-20 and incubated with
horseradish peroxidase-conjugated rabbit anti-mouse IgG (1:5,000
IgG:PBS/BSA) for 1 h at room temperature. Cells were again washed
three times with PBS:Tween and the bound antibodies were detected
by incubation with 3% O-phenylenediamine and 0.03%
H2O2 at room temperature for 30 min. Plates
were read on an ELISA microplate reader at 450 nm. The optical
densities of the wells incubated with non-specific mouse IgG1 were
subtracted from those of the wells incubated with monoclonal
antibodies. Each experiment was performed in triplicate.
Statistical analysis
Data are presented as the mean ± SD. Differences
between treatments were determined through one-way ANOVA (LSD,
S-N-K and Dunnet) using SPSS 13.0 software. P<0.05 was
considered to indicate statistically significant results. All
experiments were performed at least 3 times.
Results
Effects of berberine on cell
viability
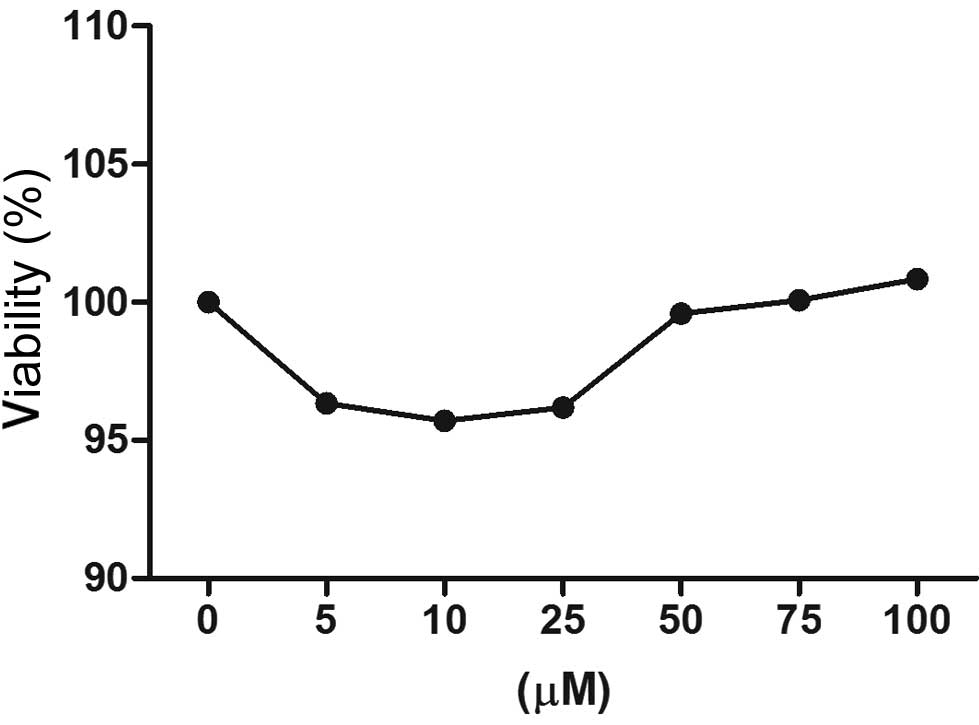
The MTT assay was performed to evaluate the effect
of berberine on the viability of HUVECs. Fig. 1 demonstrates that a concentration
of berberine between 5 and 100 μM led to no significant reduction
(~0–5%) in cell viability. A berberine dose ≤100 μM was therefore
considered noncytotoxic and doses between 5 and 25 μM were used in
subsequent experiments.
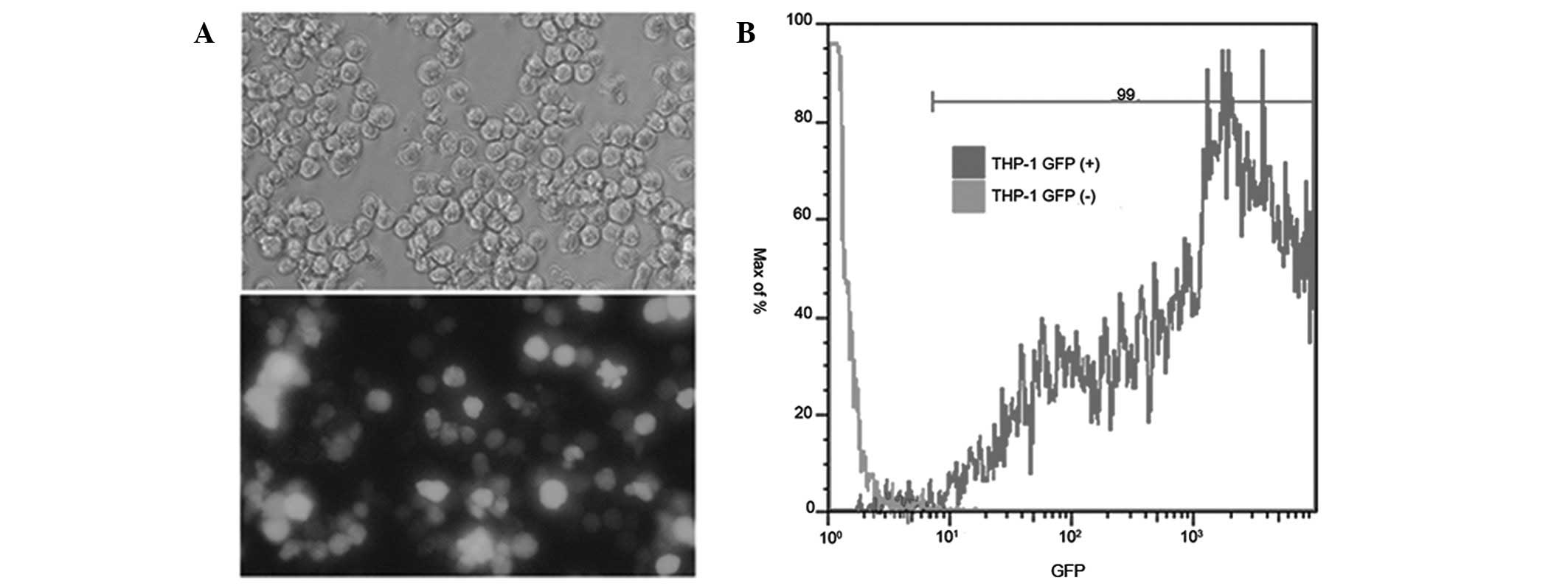
Infection efficiency of GFP-Ad in THP-1
cells
Following incubation of THP-1 cells with GFP-Ad, the
percentage of GFP expression in the cells was determined using flow
cytometry. Fig. 2 shows that the
suitable concentration of GFP-Ad-infected THP-1 cells was 5 MOI and
its infection efficiency was 99%.
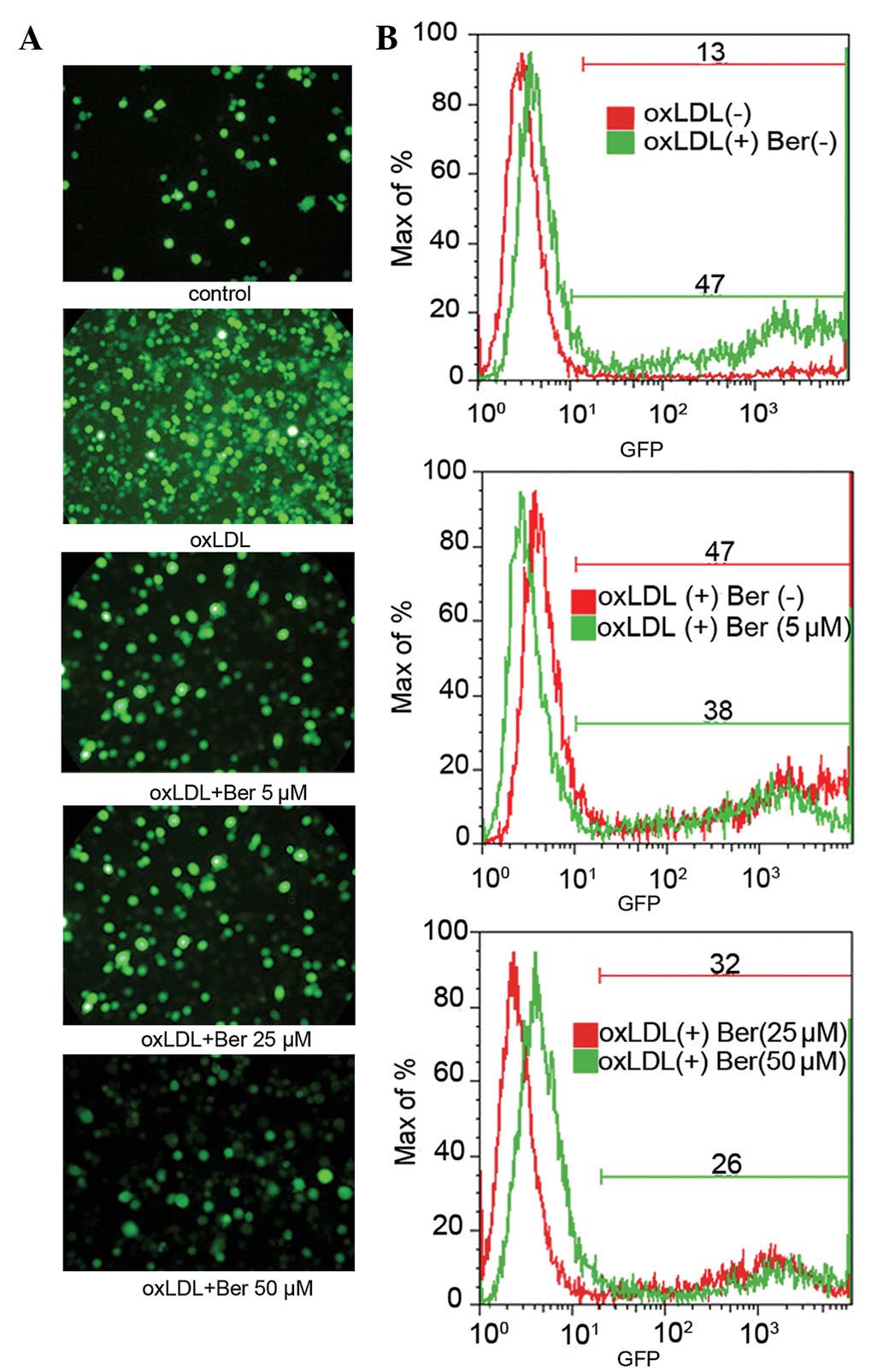
Effects of berberine on the adhesive
capacity of THP-1 cells to HUVECs
To investigate the adhesive capacity of THP-1 cells
to HUVECs, HUVECs were pretreated with berberine for 2 h and then
stimulated with oxLDL (100 μg/ml) for 6 h. The number of adherent
THP-1 cells expressing GFP fluorescence was observed via flow
cytometry and images were captured using fluorescence microscopy.
oxLDL-stimulated HUVECs were identified to exhibit significantly
increased GFP fluorescence intensity compared with the control
group without oxLDL, indicating an enhanced adhesion of monocytes
to HUVECs. However, berberine treatment markedly lowered the
adhesive capacity of monocytes and therefore reduced the GFP
fluorescence intensity of cells (Fig.
3A). Flow cytometry results (Fig.
3B) also demonstrated the same trend, wherein oxLDL treatment
increased the percentage of cells that expressed GFP fluorescence
compared with the control group without oxLDL treatment (13±1.79 vs
47±1.26%, P<0.01). However, berberine pretreatment inhibited
oxLDL-induced cell adhesion in a dose-dependent manner (5–50 μM,
from 47±1.31 to 26±3.08%, respectively).
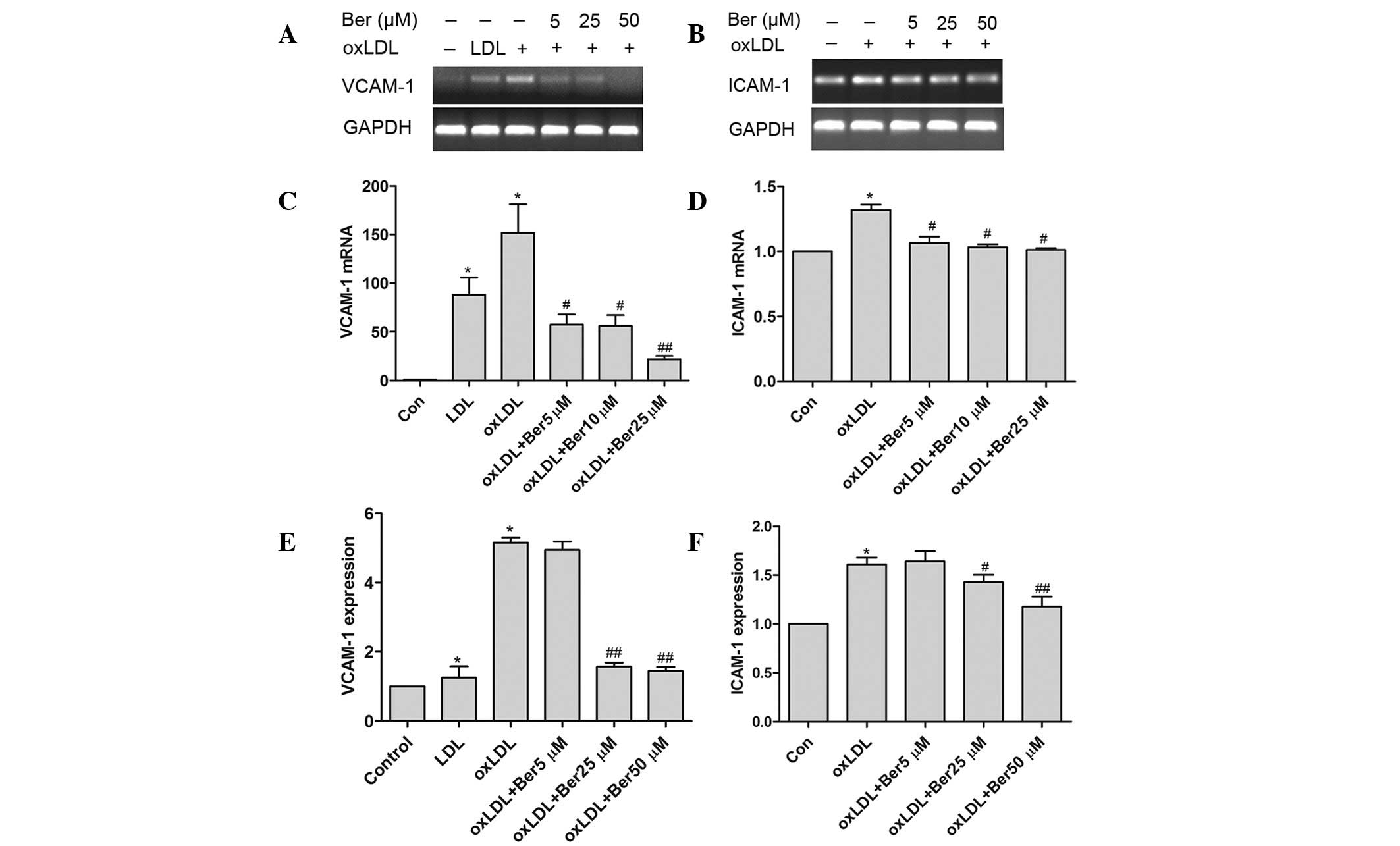
Effects of berberine on the oxLDL-induced
expression of VCAM-1 and ICAM-1 at the surface of HUVECs
oxLDL stimulated HUVECs to express VCAM-1 and
ICAM-1, which led to an increase in monocyte adhesion to HUVECs.
The expression of adhesion molecules was determined to elucidate
the molecular mechanism of berberine inhibition on the adhesive
capacity of monocytes to HUVECs. Fig.
4 demonstrates that berberine significantly suppressed VCAM-1
mRNA expression and inhibited ICAM-1 mRNA expression in the
oxLDL-simulated HUVECs in a dose-dependent manner. Furthermore,
ELISA results (Fig. 4E) indicate
that berberine inhibited expression of VCAM-1 and ICAM-1 in
oxLDL-stimulated HUVECs. The inhibition rate of VCAM-1 following
treatment with 5–50 μM berberine was between −4.6 and −22.3%,
respectively. ICAM-1 expression following treatment with 25–50 μM
berberine was between −12.3 and −27.0%, respectively. However, no
significant effect was observed following treatment with 5 μM
berberine.
Discussion
Recruitment of monocytes to the vessel wall is an
important phenomenon in the early stages of atherosclerosis
(17). In the current study,
berberine was observed to significantly reduce oxLDL-induced
monocyte adhesion to HUVECs in a dose-dependent manner. The
potential mechanisms underlying berberine suppression of the
expression of adhesion molecules, namely, VCAM-1 and ICAM-1, was
also clarified. Results indicate that berberine plays a protective
role in the early stages of the pathogenesis of
atherosclerosis.
oxLDL is a critical factor for the initiation and
acceleration of atherosclerosis and its complications (18–20).
OxLDL leads to endothelial dysfunction, which in turn leads to the
expression of adhesion molecules (21,22)
and the recruitment of monocytes to the subendothelial space
(20). The monocytes then
differentiate into macrophages, resulting in transformation of
macrophages into foam cells, followed by expression of a wide range
of proinflammatory factors that promote the development of
atherosclerosis (17,23). As a positive feedback mechanism,
cell-cell contact between monocytes and endothelial cells further
promotes their interaction through upregulation of the expression
of endothelial adhesion molecules, including VCAM-1 and ICAM-1
(8), resulting in a continual
cycle of self-activation (17).
Therefore, disrupting the processes that contribute to monocyte
adhesion to endothelial cells is an attractive target for treatment
of atherosclerosis. The present study provides direct in
vitro evidence that berberine significantly attenuates VCAM-1
and ICAM-1 expression and correspondingly reduces monocyte adhesion
to oxLDL-stimulated endothelial cells.
Although berberine is commonly used as an
antimicrobial (11) and antitumor
(24) agent in China and Korea,
accumulating studies have indicated that berberine is also
antiatherogenic. For example, berberine effectively improves
glucose metabolism in animal models (25) and clinical studies (13). Berberine reduces plasma cholesterol
(12) and inhibits expression of
proinflammatory factors (14,26).
In addition, berberine suppresses the expression of MMP-9 and
EMMPRIN in macrophages (15),
disturbing the balance of collagen production and degradation and
leading to plaque instability (27). These studies demonstrate that
berberine exerts multiple effects on the development of
atherosclerosis by reducing the major risk factors associated with
atherosclerosis, including diabetes and hyperlipemia and
interfering with microcellular mechanisms, i.e., expression of
EMMPRIN and proinflammatory factors. The present study extends
these observations by demonstrating that berberine decreases
expression of the two adhesion molecules in oxLDL-elicited HUVECs.
These observations indicate that berberine exerts its protective
effect at various stages of atherosclerosis.
In conclusion, berberine disrupts the initiating
process of atherosclerosis by attenuating adhesion of monocytes to
endothelial cells and inhibiting expression of VCAM-1 and ICAM-1.
These results provide new insights into this field of study that
are likely to be used in the identification of novel drugs against
atherosclerosis.
Acknowledgements
The current study was supported by grants from the
National Natural Science Foundation of China (no. 81102837), the
Traditional Chinese Medicine Administration of Zhejiang Province
(no. 2010ZA085) and the Education Department of Zhejiang Province
(no. Y201119884).
References
|
1
|
Price DT and Loscalzo J: Cellular adhesion
molecules and atherogenesis. Am J Med. 107:85–97. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cominacini L, Garbin U, Pasini AF, et al:
Antioxidants inhibit the expression of intercellular cell adhesion
molecule-1 and vascular cell adhesion molecule-1 induced by
oxidized LDL on human umbilical vein endothelial cells. Free Radic
Biol Med. 22:117–127. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jeng JR, Chang CH, Shieh SM and Chiu HC:
Oxidized low-density lipoprotein enhances monocyte-endothelial cell
binding against shear-stress-induced detachment. Biochim Biophys
Acta. 1178:221–227. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Khan BV, Parthasarathy SS, Alexander RW
and Medford RM: Modified low density lipoprotein and its
constituents augment cytokine-activated vascular cell adhesion
molecule-1 gene expression in human vascular endothelial cells. J
Clin Invest. 95:1262–1270. 1995. View Article : Google Scholar
|
|
5
|
Takei A, Huang Y and Lopes-Virella MF:
Expression of adhesion molecules by human endothelial cells exposed
to oxidized low density lipoprotein. Influences of degree of
oxidation and location of oxidized LDL. Atherosclerosis. 154:79–86.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ross R: The pathogenesis of
atherosclerosis: a perspective for the 1990s. Nature. 362:801–809.
1993. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Blankenberg S, Barbaux S and Tiret L:
Adhesion molecules and atherosclerosis. Atherosclerosis.
170:191–203. 2003. View Article : Google Scholar
|
|
8
|
Takahashi M, Ikeda U, Masuyama J, Kitagawa
S, Kasahara T, Shimpo M, Kano S and Shimada K: Monocyte-endothelial
cell interaction induces expression of adhesion molecules on human
umbilical cord endothelial cells. Cardiovasc Res. 32:422–429. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hansson GK: Immune mechanisms in
atherosclerosis. Arterioscler Thromb Vasc Biol. 21:1876–1890. 2001.
View Article : Google Scholar
|
|
10
|
Ikram M: A review on the chemical and
pharmacological aspects of genus Berberis. Planta Med.
28:353–358. 1975. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Stermitz FR, Lorenz P, Tawara JN, Zenewicz
LA and Lewis K: Synergy in a medicinal plant: antimicrobial action
of berberine potentiated by 5′-methoxyhydnocarpin, a multidrug pump
inhibitor. Proc Natl Acad Sci USA. 97:1433–1437. 2000.PubMed/NCBI
|
|
12
|
Kong W, Wei J, Abidi P, et al: Berberine
is a novel cholesterol-lowering drug working through a unique
mechanism distinct from statins. Nat Med. 10:1344–1351. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yin J, Xing H and Ye J: Efficacy of
berberine in patients with type 2 diabetes mellitus. Metabolism.
57:712–717. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee CH, Chen JC, Hsiang CY, Wu SL, Wu HC
and Ho TY: Berberine suppresses inflammatory agents-induced
interleukin-1beta and tumor necrosis factor-alpha productions via
the inhibition of IkappaB degradation in human lung cells.
Pharmacol Res. 56:193–201. 2007. View Article : Google Scholar
|
|
15
|
Huang Z, Wang L, Meng S, Wang Y, Chen T
and Wang C: Berberine reduces both MMP-9 and EMMPRIN expression
through prevention of p38 pathway activation in PMA-induced
macrophages. Int J Cardiol. 146:153–158. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hsieh YS, Kuo WH, Lin TW, Chang HR, Lin
TH, Chen PN and Chu SC: Protective effects of berberine against
low-density lipoprotein (LDL) oxidation and oxidized LDL-induced
cytotoxicity on endothelial cells. J Agric Food Chem.
55:10437–10445. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pamukcu B, Lip GY, Devitt A, Griffiths H
and Shantsila E: The role of monocytes in atherosclerotic coronary
artery disease. Ann Med. 42:394–403. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mitra S, Deshmukh A, Sachdeva R, Lu J and
Mehta JL: Oxidized low-density lipoprotein and atherosclerosis
implications in antioxidant therapy. Am J Med Sci. 342:135–142.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mitra S, Goyal T and Mehta JL: Oxidized
LDL, LOX-1 and atherosclerosis. Cardiovasc Drugs Ther. 25:419–429.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fotis L, Agrogiannis G, Vlachos IS, et al:
Intercellular adhesion molecule (ICAM)-1 and vascular cell adhesion
molecule (VCAM)-1 at the early stages of atherosclerosis in a rat
model. In Vivo. 26:243–250. 2012.PubMed/NCBI
|
|
21
|
Chen H, Li D, Saldeen T and Mehta JL:
Transforming growth factor-beta(1) modulates oxidatively modified
LDL-induced expression of adhesion molecules: role of LOX-1. Circ
Res. 89:1155–1160. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li D, Chen H, Romeo F, Sawamura T, Saldeen
T and Mehta JL: Statins modulate oxidized low-density
lipoprotein-mediated adhesion molecule expression in human coronary
artery endothelial cells: role of LOX-1. J Pharmacol Exp Ther.
302:601–605. 2002. View Article : Google Scholar
|
|
23
|
Steffens S and Mach F: Inflammation and
atherosclerosis. Herz. 29:741–748. 2004. View Article : Google Scholar
|
|
24
|
Anis KV, Rajeshkumar NV and Kuttan R:
Inhibition of chemical carcinogenesis by berberine in rats and
mice. J Pharm Pharmacol. 53:763–768. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yin J, Gao Z, Liu D, Liu Z and Ye J:
Berberine improves glucose metabolism through induction of
glycolysis. Am J Physiol Endocrinol Metab. 294:E148–E156. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen FL, Yang ZH, Liu Y, et al: Berberine
inhibits the expression of TNFalpha, MCP-1 and IL-6 in
AcLDL-stimulated macrophages through PPARgamma pathway. Endocrine.
33:331–337. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Newby AC: Metalloproteinase expression in
monocytes and macrophages and its relationship to atherosclerotic
plaque instability. Arterioscler Thromb Vasc Biol. 28:2108–2114.
2008. View Article : Google Scholar : PubMed/NCBI
|