Introduction
Deep vein thrombosis (DVT) and pulmonary embolism
(PE) are collectively known as venous thromboembolism (VTE). PE
includes acute pulmonary embolism (APE) and chronic thromboembolic
pulmonary hypertension (CTEPH). PE is a major health problem, with
a high incidence, mortality, misdiagnosis and low detection rate
(1). There is much debate with
regard to the diagnosis, treatment and prevention of PE, as the
etiology and pathogenesis of this disease have not been well
characterized. It has been reported that patients with APE and
CTEPH possess lowered immune function (2,3). In
this study, we used human cDNA microarray analysis to determine the
mRNA expression levels of cytokines in PBMCs isolated from PE
patients and controls. We designed this in vitro study to
observe changes in the immune function of PE patients.
Patients and methods
Patients
Twenty patients with PE were recruited from the
Tongji Hospital (Tongji University School of Medicine, Shanghai,
China) between 2007 and 2008. A diagnosis of PE required any two of
the following three criteria to be met: i) selective pulmonary
angiography demonstrating pulmonary artery obstruction or filling
defect; ii) lung ventilation/perfusion scan showing single or
multiple blood perfusion defects, normal or; iii) clinical
diagnosis: there are risk factors for PE and other cardiovascular
diseases that are able to be excluded by clinical performance,
electrocardiogram and chest film. Arterial blood gas analysis
suggested that hypoxemia and hypocapnia, D-dimer detection,
echocardiography and chest computed tomography support the
diagnosis of PE. We selected 20 patients admitted to the Department
of Cardiology, Tongji Hospital, at the same time as the controls
used in the study. The patients were divided into two groups: i)
the PE patient group: 20 patients (11 males and 9 females), with a
mean age of 70±14 years (range, 44–89 years), including 3 cases of
CTEPH; ii) the control group: 20 patients (11 males and 9 females)
without PE, DVT, arterial thrombosis and congenital coagulation
abnormality, with a mean age of 72±14 years (range, 44–91 years),
who were matched for gender and age with the PE group. There was no
significant statistical difference between the age of the two
groups (P>0.05). The clinical trial was approved by the Ethics
Committee of the Tongji University and written informed consent was
obtained.
Gene expression profiling
Agilent G4112A Whole Human Genome Oligo Microarrays
were purchased from Agilent Technologies (Palo Alto, CA, USA). The
microarray was composed of 44,290 spots, including 41,675 genes or
transcripts, 314 negative control spots, 1,924 positive control
spots and 359 blank spots. The functions of >70% of the genes in
the microarray were known. All patients were subjected to
microarray analysis.
Total RNA isolation
A 5-ml sample of peripheral blood anticoagulated
with EDTA was drawn from patients suspected with PE immediately
following hospital admittance and from those patients without PE.
Mononuclear cells were obtained through density gradient
centrifugation (3,000 × g) with Ficoll solution and the remaining
red blood cells were destroyed with erythrocyte lysis buffer
(Qiagen, Hilden, Germany). Total mononuclear cell RNA was extracted
with TRIzol (Invitrogen Life Technologies, Carlsbad, USA) and
purified with RNeasy column (Qiagen) according to the
manufacturer’s instructions. Isolated total RNA was evident and
quantified by means of Nanodrop ND-1000 spectrophotometer (Nanodrop
Technology, Cambrige, UK).
Detection of gene expression
Approximately 1 μg of total RNA was reverse
transcribed into double-strand cDNA. Following purification, in
vitro amplification was performed using the Low RNA Input
Linear Amplification kit (Agilent Technologies) and modified UTP
[aaUTP, 5-(3-aminoallyl)-UTP] was used to replace UTP. The
integrated aaUTP is capable of interacting with the Cy3 NHS
ester-forming fluorescent products, which are then used for
hybridization. The integration rate of fluorescence was determined
using a Nanodrop ND-1000 spectrophotometer. Following this, the
hybridization mixture was prepared with the oligonucleotide
microarray in situ Hybridization Plus kit (Agilent
Technologies). Approximately 750 ng of fluorescent products were
fragmented at 60°C and hybridization was conducted in Human
Whole-Genome 60-mer oligo-chips (G4112F, Agilent Technologies) at
60°C for 17 h at 10 rpm. Following hybridization, the chips were
washed with Gene Expression Wash Buffer (Agilent Technologies)
according to the manufacturer’s instructions. Original signals were
obtained using Scanner and Feature Extraction software (Agilent
Technologies). The standardization of original signals was
performed with RMA standardized methods and standardized signal
values were used for the screening of differentially expressed
genes.
RT-PCR
Three differentially expressed genes in the
microarray analysis were selected and their expression was
confirmed by RT-PCR. Among the genes with differential expression,
three genes were randomly selected and subjected to RT-PCR along
with the housekeeping gene (GAPDH). The relative expression was
indicated as the expression of the target genes normalized by that
of GAPDH (2-ΔΔCt). Melting curve and 2-ΔΔCt
methods were used to compare differences in gene expression between
the control and PE groups. Results from RT-PCR were consistent with
those obtained from the microarray analysis.
Statistical analysis
The independent samples t-test was used to compare
mRNA levels in samples from PE patients and controls. Statistical
tests were performed using SPSS 17.0, and P<0.05 was considered
to indicate a statistically significant result. Prior to the
t-test, a test for equality of variances was performed; if
variances were not equal, the t-test result was corrected.
Results
IFN mRNA expression levels
PBMCs obtained from PE patients demonstrated low IFN
mRNA expression levels. Expression of the 12 genes encoding type I
IFNs, as well as the gene encoding IFNγ, were examined (Fig. 1). In PBMCs from PE patients, the
expression levels of genes encoding IFNα5, IFNα6, IFNα8, IFNα14,
IFNκ, IFNω1 and IFNɛ1 were significantly lower than those detected
in PBMCs from controls (P<0.05). IFNγ mRNA expression was
significantly downregulated in PBMCs from PE patients compared with
controls (P<0.01).
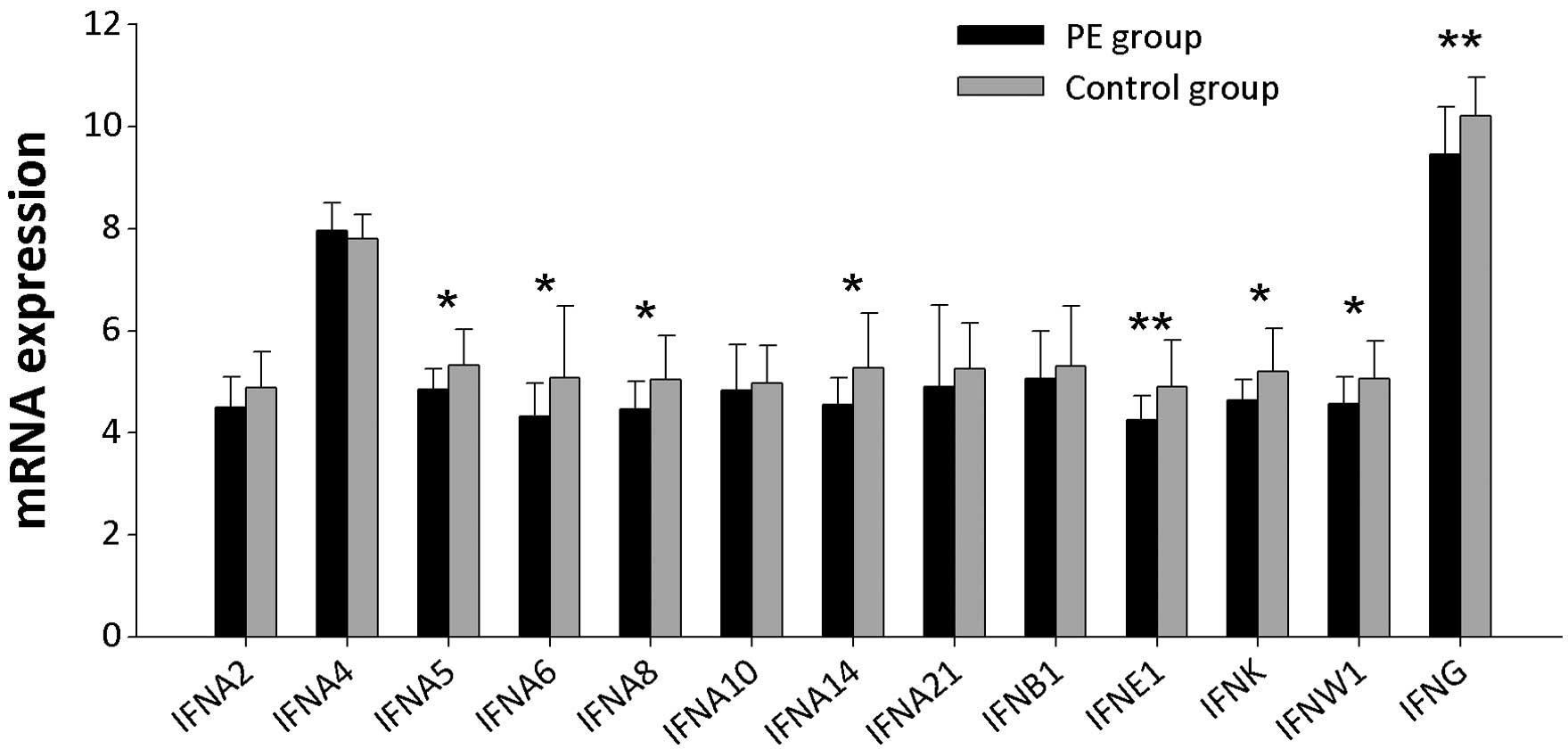 | Figure 1IFN mRNA expression levels in PBMCs
from PE patients and controls. IFNA2, IFNA4, IFNA5, IFNA6, IFNA8,
IFNA10, IFNA14, IFNA21, IFNB1, IFNE1, IFNK, IFNW1 and INFG
represent the gene symbols of IFNα2, IFNα4, IFNα5, IFNα6, IFNα8,
IFNα10, IFNα14, IFNα21, IFNβ1, IFNɛ1, IFNκ, IFNω1 and IFNγ,
respectively. *P<0.05 and **P<0.01.
PBMCs, peripheral blood mononuclear cells; PE, pulmonary
embolism. |
IL mRNA expression levels
The expression levels of 37 genes encoding ILs were
examined (Fig. 2). The expression
levels of genes encoding IL1α, IL2, IL3, IL9, IL13, IL17β, IL19,
IL22, IL23α, IL24, IL25 and IL31 were significantly lower in PBMCs
from PE patients compared with the controls (P<0.05). IL10 and
IL28A gene expression levels were greater in PBMCs from PE patients
than controls (P<0.05).
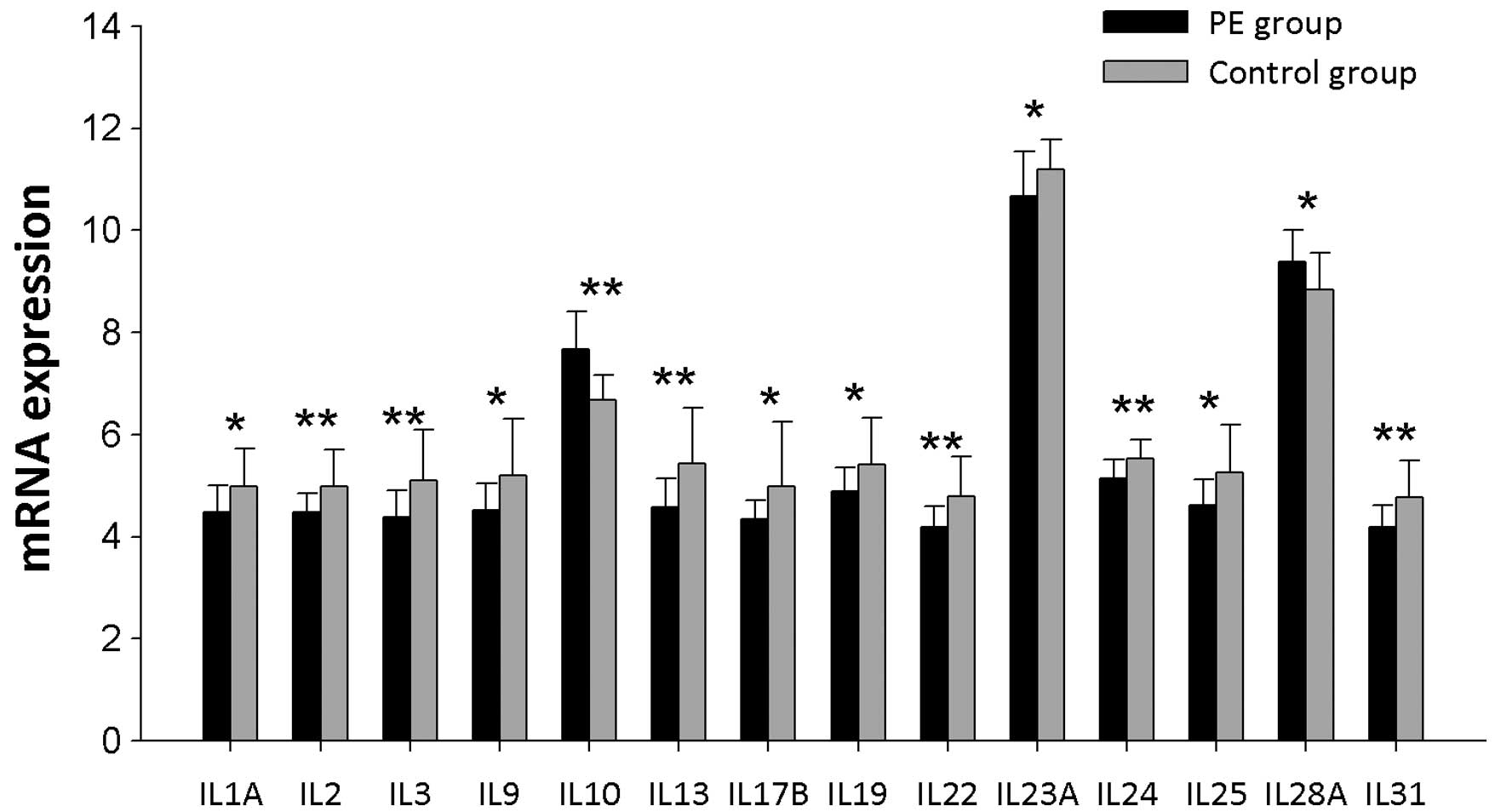 | Figure 2IL mRNA expression levels were
significantly different in PBMCs from PE patients and controls.
IL1A, IL2, IL3, IL9, IL10, IL13, IL17B, IL19, IL22, IL23A, IL24,
IL25, IL28A, and IL31 represent the gene symbols of IL1α, IL2, IL3,
IL9, IL10, IL13, IL17β, IL19, IL22, IL23α, IL24, IL25, IL28α and
IL31, respectively. *P<0.05 and
**P<0.01. PBMCs, peripheral blood mononuclear cells;
PE, pulmonary embolism. |
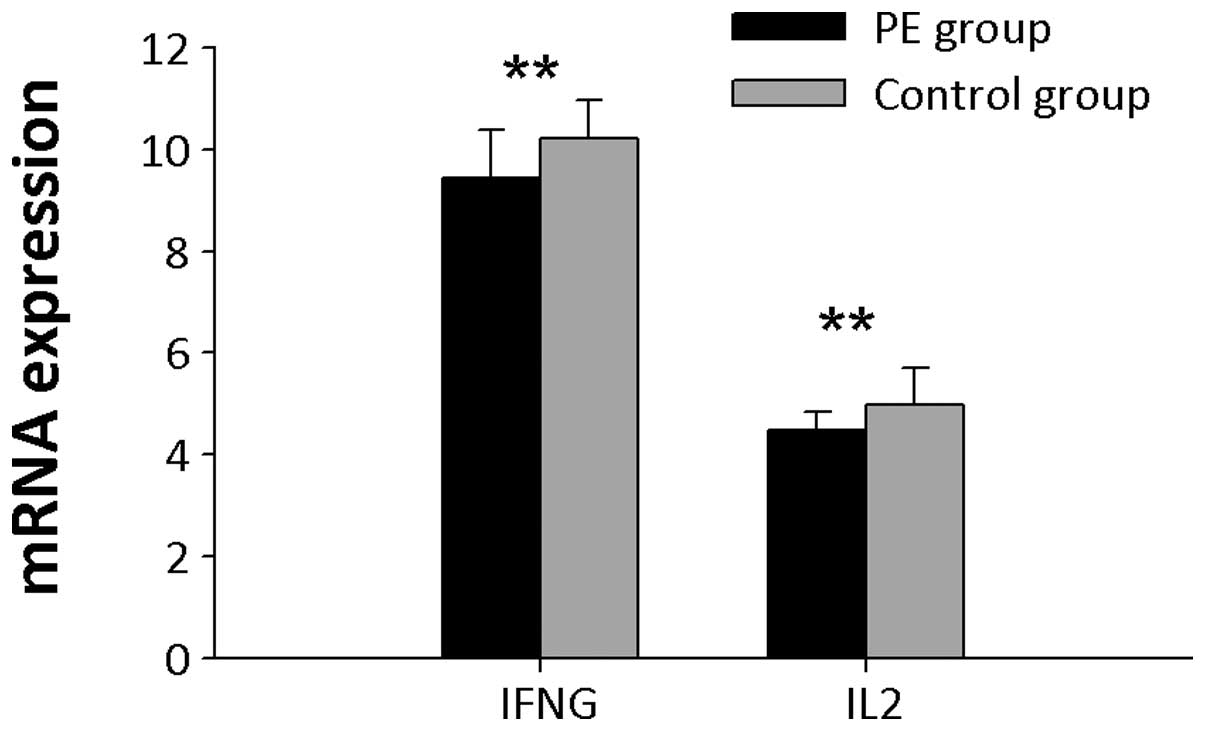
Th1/Th2 mRNA expression levels
Th1 cytokine mRNA expression levels including IFNγ
and IL2 mRNAs were detected (Fig.
3), and in PE patients, both were significantly downregulated
compared with the control group (P<0.01). Th2 cytokine mRNA
expression levels, including IL4, IL6 and IL10 mRNAs were examined
(Fig. 4). IL10 mRNA expression
levels were significantly higher in PBMCs from PE patients than
controls (P<0.01). No significant difference in IL4 and IL6 mRNA
expression levels was observed between the two groups.
Chemokine mRNA expression levels
Twelve genes encoding CXC chemokines were detected
(Fig. 5). In PE patients, mRNA
expression levels of Cxcl1, Cxcl2, Cxcl6, Cxcl13 and Cxcl14 were
significantly upregulated (P<0.05), and Cxcl10 mRNA expression
levels were significantly downregulated compared with controls
(P<0.01). Twenty-three genes encoding CC chemokines were
examined (Figs. 6 and 7) and the mRNA expression levels of CC
chemokines were significantly lower in PE patients than controls
(P<0.01). C chemokine mRNA expression levels including Xcl1 and
Xcl2 mRNAs were detected (Fig. 8),
and Xcl1 mRNA expression was significantly lower in PE patients
(P<0.01). No significant difference in Xcl2 mRNA expression
levels was observed between the two groups. CX3C chemokine Cx3cl1
mRNA expression was detected, and there was no significant
difference between the two groups.
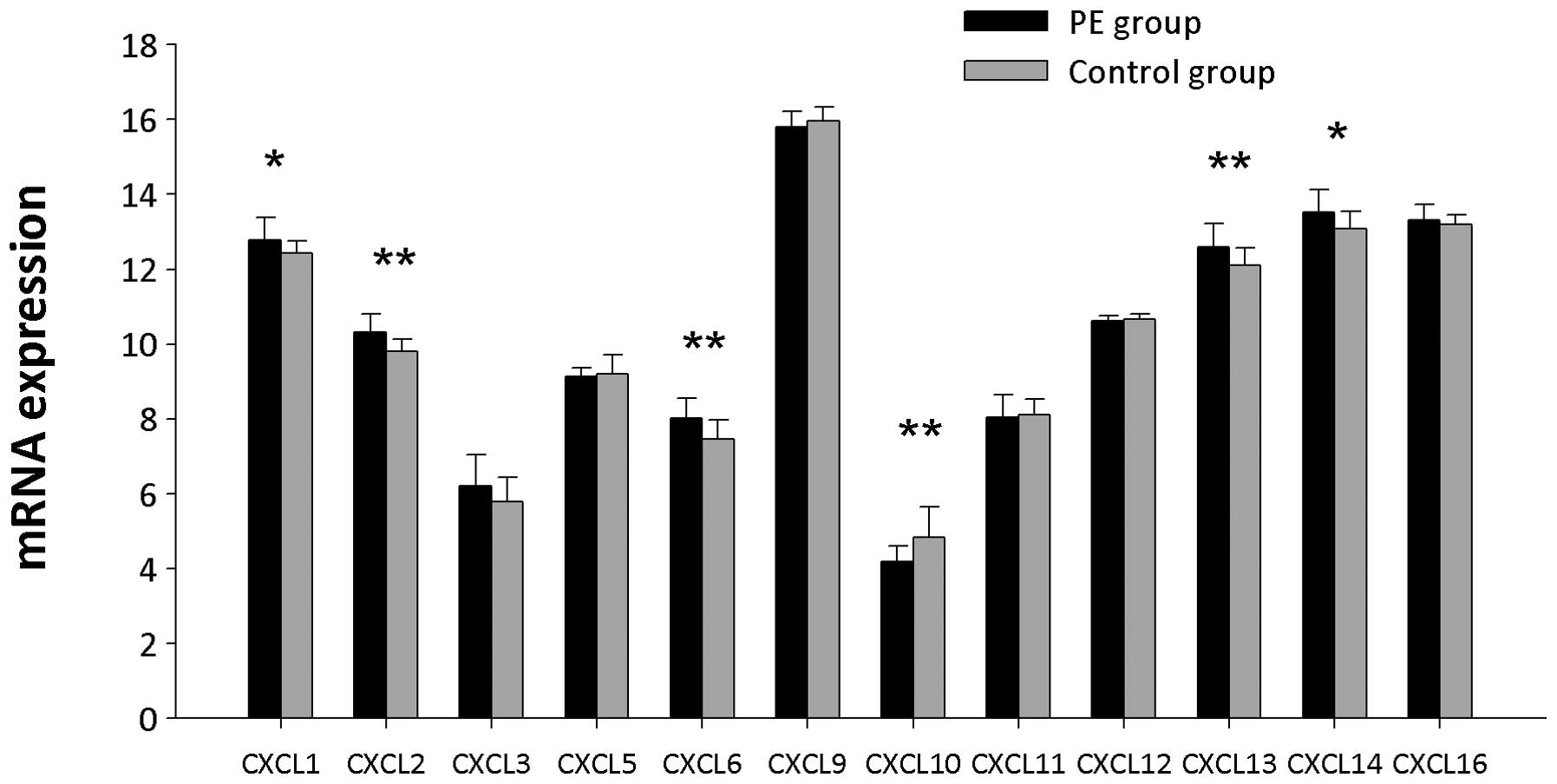 | Figure 5CXC chemokine mRNA expression levels
in PBMCs from PE patients and controls. CXCL1, CXCL2, CXCL3, CXCL5,
CXCL6, CXCL9, CXCL10, CXCL11, CXCL12, CXCL13, CXCL14 and CXCL16
represent the gene symbols of Cxcl1, Cxcl2, Cxcl3, Cxcl5, Cxcl6,
Cxcl9, Cxcl10, Cxcl11, Cxcl12, Cxcl13, Cxcl14 and Cxcl16,
respectively. *P<0.05 and **P<0.01.
PBMCs, peripheral blood mononuclear cells; PE, pulmonary
embolism. |
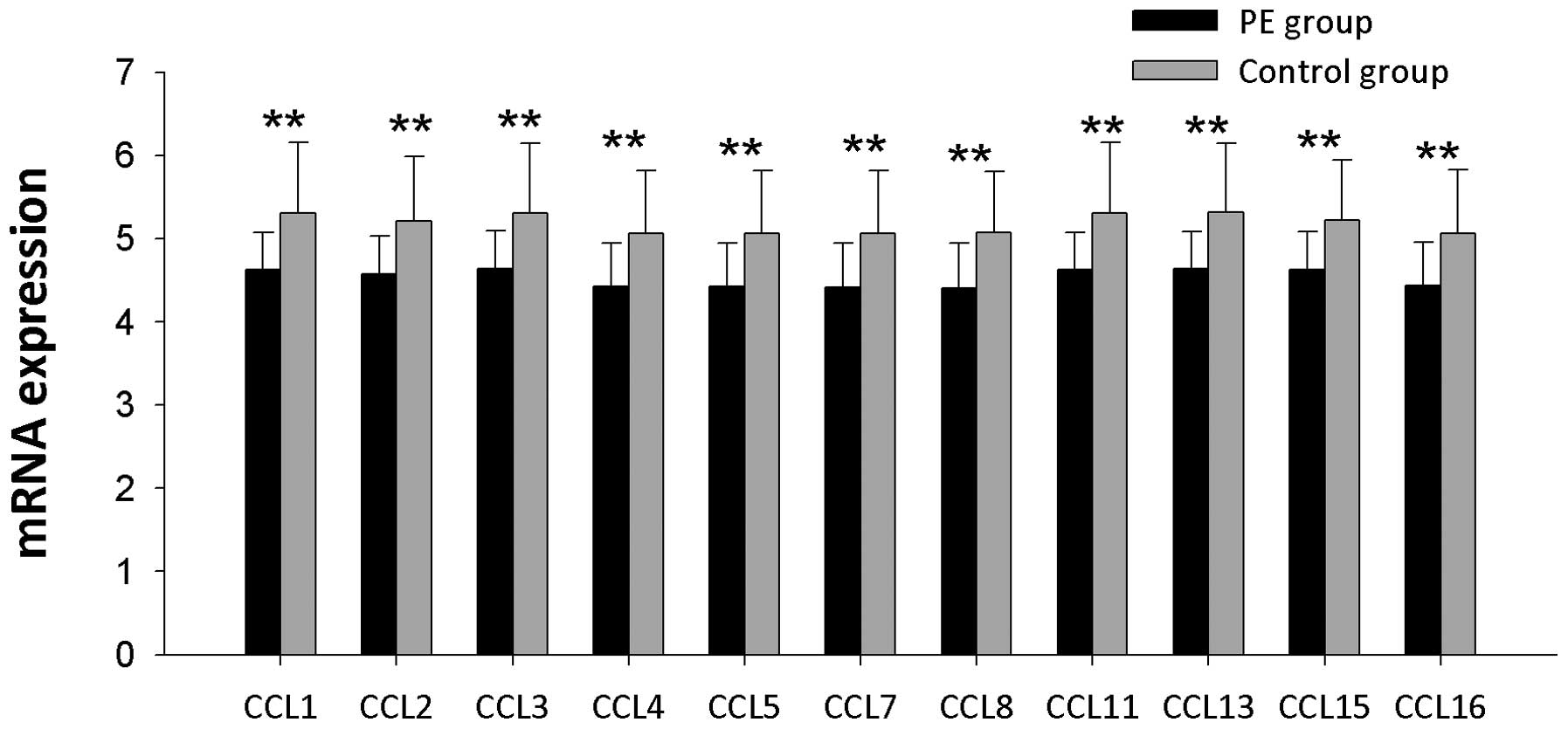 | Figure 6CC chemokine mRNA expression levels in
PBMCs from PE patients and controls. CCL1, CCL2, CCL3, CCL4, CCL5,
CCL7, CCL8, CCL11, CCL13, CCL15 and CCL16 represent the gene
symbols of Ccl1, Ccl2, Ccl3, Ccl4, Ccl5, Ccl7, Ccl8, Ccl11, Ccl13,
Ccl15 and Ccl16, respectively. *P<0.05 and
**P<0.01. PBMCs, peripheral blood mononuclear cells;
PE, pulmonary embolism. |
 | Figure 7CC chemokine mRNA expression levels in
PBMCs from PE patients and controls. CCL17, CCL18, CCL19, CCL20,
CCL21, CCL22, CCL23, CCL24, CCL25, CCL26, CCL27 and CCL28 represent
the gene symbols of Ccl17, Ccl18, Ccl19, Ccl20, Ccl21, Ccl22,
Ccl23, Ccl24, Ccl25, Ccl26, Ccl27 and Ccl28, respectively.
*P<0.05 and **P<0.01. PBMCs, peripheral
blood mononuclear cells; PE, pulmonary embolism. |
TNF superfamily and TNF receptor
superfamily mRNA expression levels
Thirty-eight genes encoding members of the TNF
superfamily and TNF receptor superfamily were examined (Fig. 9). In PE patients, the mRNA
expression levels of TNF superfamily members 1, 9 and 13, and TNF
receptor superfamily members 1A, 1B, 9, 10B, 10C, 10D and 19L, were
significantly upregulated (P<0.05), whereas TNF receptor
superfamily members 11B, 19 and 25, were significantly
downregulated compared with controls (P<0.05).
 | Figure 9TNF superfamily and TNF receptor
superfamily mRNA expression levels which were significantly
different in PBMCs from PE patients and controls. LTA, TNFSF9,
TNFSF13, TNFRSF1A, TNFRSF1B, TNFRSF9, TNFRSF10B, TNFRSF10C,
TNFRSF10D, TNFRSF11B, TNFRSF19, TNFRSF19L, and TNFRSF25 represent
the gene symbols of TNF superfamily members 1, 9 and 13, and TNF
receptor superfamily members 1A, 1B, 9, 10B, 10C, 10D, 11B, 19, 19L
and 25, respectively. *P<0.05 and
**P<0.01. PBMCs, peripheral blood mononuclear cells;
PE, pulmonary embolism. |
Colony stimulating factor mRNA expression
levels
Six genes encoding colony stimulating factors were
detected (Fig. 10) and the mRNA
expression levels of granulocyte-macrophage colony stimulating
factor (GM-CSF), granulocyte colony-stimulating factor (G-CSF),
erythropoietin (EPO), thrombopoietin (THPO) and mast cell growth
factor (KITLG) were significantly lower in PBMCs from PE patients
than controls (P<0.05).
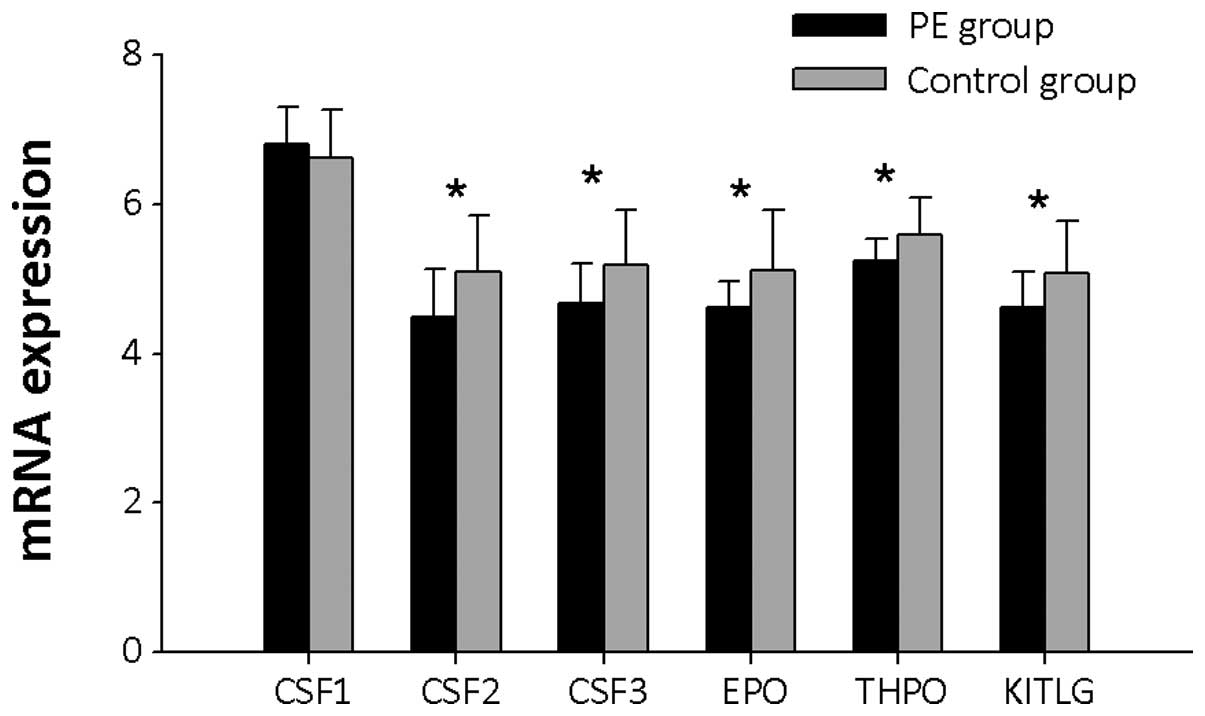 | Figure 10CSF mRNA expression levels in PBMCs
from PE patients and controls. CSF1, CSF2, CSF3, EPO, THPO and
KITLG represent the gene symbols of macrophage colony stimulating
factor, granulocyte-macrophage colony stimulating factor,
granulocyte colony stimulating factor, erythropoietin,
thrombopoietin and mast cell growth factor, respectively.
*P<0.05 and **P<0.01. PBMCs, peripheral
blood mononuclear cells; PE, pulmonary embolism; CSF, colony
stimulating factor; EPO, erythropoietin; THPO, thrombopoietin;
KITLG, mast cell growth factor. |
Other cytokine mRNA expression
levels
We examined eight genes associated with transforming
growth factor (TGF), epidermal growth factor (EGF) and vascular
endothelial growth factor (VEGF) (Fig. 11). The mRNA expression levels of
TGFβ1, TGFβ1-induced transcript 1, EGF and VEGF were significantly
upregulated (P<0.01), whereas TGFβ3 mRNA was significantly
downregulated (P<0.05) in PBMCs from PE patients compared with
controls.
 | Figure 11Other cytokine mRNA expression levels
in PBMCs from PE patients and controls. TGFB1, TGFB1I1, TGFB2,
TGFB3, EGF, VEGF, VEGFB and VEGFC represent the gene symbols of
TGFβ1, TGFβ1-induced transcript 1, TGFβ2, TGFβ3, epidermal growth
factor, VEGF, VEGFB and VEGFC, respectively. *P<0.05
and **P<0.01. PBMCs, peripheral blood mononuclear
cells; PE, pulmonary embolism; VEGF, vascular endothelial growth
factor; TGF, transforming growth factor; EGF, epidermal growth
factor. |
Discussion
IFNs are classified into two major groups: type I
and type II. Type I IFNs include IFNα, IFNβ, IFNɛ, IFNω and IFNκ
(4). We detected type I IFN mRNAs
in the PE patients compared with controls. We demonstrated that
IFNα5, IFNα6, IFNα8, IFNα14, IFNκ, IFNω1 and IFNɛ1 mRNA expression
levels were significantly lower in PE patients compared with
controls. The type I IFNs possess antiviral, antiproliferative and
immunomodulatory effects (5). It
has been reported that IFNα/β receptor knockout mice are
susceptible to viral infection (6). Mice treated with antibodies against
IFNα/β may be lethally infected with filoviruses (7). Thus, our results suggest that PE
patients are susceptible to viral infection. In this study, we also
examined IFNγ mRNA expression levels which were observed to be
significantly downregulated in PE patients compared with controls.
IFNγ is important in the defence against intracellular bacteria
(Mycobacterium, Listeria) and parasites (Leishmania) (8). A caudal vena cava thrombus in a dog
with leishmaniasis has been reported (9). It has also been demonstrated that
three patients with pulmonary tuberculosis suffered from PTE or DVT
(10). Therefore, our results show
that PE patients are susceptible to intracellular bacteria and
parasites.
T-helper lymphocytes can be divided into Th1 and Th2
subsets based on their functional abilities and the profile of the
cytokines they produce (11). Th1
and Th2 cells express different cytokines: Th1 cells produce IL2,
IL12 and IFNγ, whereas Th2 cells produce IL4, IL5, IL6, IL10 and
IL13 (12). Th1 cells promote
cellular immunity, while Th2 cells induce humoral immunity and
upregulate antibody production (13). The shift in Th1/Th2 balance leads
to immune dysfunction. Our results demonstrated the presence of
lower mRNA expression levels of Th1 cytokines (IFNγ and IL2) and
higher mRNA expression levels of Th2 cytokine (IL10) in PE
patients, suggesting a shift towards Th2 dominance in PE patients.
Thus, our results suggest that an immune dysfunction in PE patients
may exist.
Chemokines can be classified into four classes; CXC,
CC, C and CX3C according to their structure. Previous studies have
shown that a polymorphonuclear (PMN) activation in thrombosis is
present, and that PMN activation may be important in the etiologies
of thrombosis (14,15). CXC chemokines possess a potent
chemotactic activity on neutrophils. Twelve genes encoding CXC
chemokines and mRNA expression levels of Cxcl1, Cxcl2, Cxcl6,
Cxcl13 and Cxcl14 were significantly upregulated in PE patients
compared with controls, suggesting that a high expression of
chemokines may induce neutrophil aggregation. The CC chemokines
RANTES/CCL5, MIP-1α/CCL3 and MIP-1β/CCL4 produced by CD8+ T cells
induce the inhibition of human immunodeficiency virus (HIV)
(16). During chronic viral
infection, virus-specific CD8 T cells generated low levels of
cytokines and the cytotoxic ability of CD8 T cells was decreased in
RANTES knockout mice (17).
Deficiency of MIP-1α led to the recovery of higher levels of
infectious viruses in comparison with fully competent mice in
paramyxovirus infection (18).
During mouse hepatitis virus infection, the cytokine production and
cytolytic activity of CD8+ T cells were reduced in CCL3 knockout
mice (19). Our results
demonstrate the presence of lower mRNA expression levels of Ccl3,
Ccl4 and Ccl5 in PE patients, which suggests that a reduced
antiviral ability in PE patients may exist.
The interaction of costimulatory receptor CD137 (TNF
receptor superfamily, member 9) and its ligand CD137L (TNF
superfamily, member 9), which is involved in the TNF superfamily,
is important in immune regulation (20). Blockade of the CD137-CD137L pathway
significantly reduced the percentage of CD3+ CD8+ T cells in mice
transplanted with donor bone marrow cells plus primary MLC spleen T
cells (21). It has been reported
that 4-1BB (also known as CD137)-deficient mice have enhanced
susceptibility to Listeria monocytogenes (intracellular bacteria)
infections (22). It has also been
shown that the cytotoxic lymphocyte (CTL) response to the influenza
virus was reduced in mice lacking 4-1BBL (23). In this study, mRNA expression
levels of CD137 and CD137L were upregulated in PE patients,
suggesting that PE patients may be more susceptible to infection.
TNF may interfere with viral replication in several ways (24). TNF has two receptors, TNF receptor
1 (TNFR1) and 2 (TNFR2) (24). The
TNF/TNFR pathway is important in the induction of apoptosis
(25). It has been demonstrated
that TNFR1 and TNFR2 induced evident apoptosis, including CD4 and
CD8 T cells during HIV infection (26). In our study, mRNA expression levels
of TNFR1 and TNFR2 were significantly greater in PE patients than
controls. Thus, we hypothesize that peripheral T-cell apoptosis
induced by TNFR1 and TNFR2 may exist, leading to a decreased immune
function and enhanced susceptibility to virus expression in PE
patients.
GM-CSF is important to the immune system. GM-CSF is
capable of affecting the balance of Th1/Th2. It has been reported
that mice lacking GM-CSF were not able to control an
aerosol-delivered infection with Mycobacterium tuberculosis
and died from severe necrosis in the lungs as it was not able to
express a Th1 response (27). In
this study, mRNA expression levels of GM-CSF were significantly
downregulated in PE patients, suggesting that there may be
decreased immune function and a reduced antipathogenic
micro-organism ability in PE patients.
VEGF-A (also known as VEGF) is an important member
of the VEGF family. PE-induced endothelium injury is important in
the sequelae following PE (28).
In this study, mRNA expression levels of VEGF in PE patients were
significantly higher than controls, suggesting that the level of
VEGF mRNA may increase compensatively to promote endothelial cell
repair and vascular remodeling following PE-induced endothelium
injury. Three types of isoform have been identified in mammals;
TGFβ1, TGFβ2 and TGFβ3 (29). All
three isoforms have similar properties in vitro, however,
TGFβ1 is dominantly expressed in the immune system (30). It has been reported that the level
of TGFβ1 increases during cytomegalovirus and hepatitis B virus
infection (31,32). It has been shown that in HIV
infection, TGFβ is capable of suppressing the cellular immune
response and the humoral immune response (33). In this study, mRNA expression
levels of TGF were upregulated significantly in PE patients,
suggesting that a decreased immune function in PE patients may
exist and this disease may be associated with viral infections.
From the characteristics of a variety of cytokine
mRNA expression levels in PE patients, we conclude that the immune
function and the ability of clearing viruses, intracellular
bacteria and parasites are reduced in PE patients. Previously, we
reported downregulated gene expression associated with NK and T
cells in patients with PE (34),
and the number of CD3+ and CD8+ T cells were significantly
downregulated in patients with APE and CTEPH (2,3). In
this study, we detected and analyzed the mRNA expression levels of
cytokines in patients with PE and conclude that an immune
dysfunction in PE patients exists, which is similar with the
results of previously performed cytology tests. Our results may
provide evidence for the etiology and pathogenesis of VTE.
Acknowledgements
This study was supported by the ‘12th Five Year’
National Science and Technology Supporting Program
(2011BAI11B16).
References
|
1
|
Heit JA: The epidemiology of venous
thromboembolism in the community. Arterioscler Thromb Vasc Biol.
28:370–372. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang L, Song H, Gong Z, Duan Q and Liang
A: Acute pulmonary embolism and dysfunction of CD3+ CD8+ T cell
immunity. Am J Respir Crit Care Med. 184:13152011.PubMed/NCBI
|
|
3
|
Haoming S, Lemin W, Zhu G, et al: T
cell-mediated immune deficiency or compromise in patients with
CTEPH. Am J Respir Crit Care Med. 183:417–418. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sang Y, Rowland RR, Hesse RA and Blecha F:
Differential expression and activity of the porcine type I
interferon family. Physiol Genomics. 42:248–258. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Day SL, Ramshaw IA, Ramsay AJ and
Ranasinghe C: Differential effects of the type I interferons
alpha4, beta, and epsilon on antiviral activity and vaccine
efficacy. J Immunol. 180:7158–7166. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Abbas AK and Lichtman AH: Cellular and
Molecular Immunology. 5th edition. Saunders; Philadelphia, PA: 562.
2005
|
|
7
|
Bray M: The role of the Type I interferon
response in the resistance of mice to filovirus infection. J Gen
Virol. 82:1365–1373. 2001.PubMed/NCBI
|
|
8
|
van den Broek MF, Müller U, Huang S,
Zinkernagel RM and Aguet M: Immune defence in mice lacking type I
and/or type II interferon receptors. Immunol Rev. 148:5–18.
1995.PubMed/NCBI
|
|
9
|
Font A and Closa JM: Ultrasonographic
localization of a caudal vena cava thrombus in a dog with
leishmaniasis. Vet Radiol Ultrasound. 38:394–396. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Komazaki Y, Sakakibara Y, Sakashita H,
Miyazaki Y and Inase N: Pulmonary thromboembolism with pulmonary
tuberculosis. Kekkaku. 86:717–722. 2011.(In Japanese).
|
|
11
|
Constant SL and Bottomly K: Induction of
Th1 and Th2 CD4+ T cell responses: the alternative approaches. Annu
Rev Immunol. 15:297–322. 1997.
|
|
12
|
Romagnani S: Biology of human TH1 and TH2
cells. J Clin Immunol. 15:121–129. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kidd P: Th1/Th2 balance: the hypothesis,
its limitations, and implications for health and disease. Altern
Med Rev. 8:223–246. 2003.PubMed/NCBI
|
|
14
|
LoPresti R, Ferrara F, Canino B, Montana M
and Caimi G: Deep venous thrombosis: leukocyte rheology at baseline
and after in vitro activation. Haemostasis. 30:168–173.
2000.PubMed/NCBI
|
|
15
|
Esposito CJ, Popescu WM, Rinder HM,
Schwartz JJ, Smith BR and Rinder CS: Increased leukocyte-platelet
adhesion in patients with graft occlusion after peripheral vascular
surgery. Thromb Haemost. 90:1128–1134. 2003.PubMed/NCBI
|
|
16
|
Cocchi F, DeVico AL, Garzino-Demo A, Arya
SK, Gallo RC and Lusso P: Identification of RANTES, MIP-1 alpha,
and MIP-1 beta as the major HIV-suppressive factors produced by
CD8+ T cells. Science. 270:1811–1815. 1995.
|
|
17
|
Crawford A, Angelosanto JM, Nadwodny KL,
Blackburn SD and Wherry EJ: A role for the chemokine RANTES in
regulating CD8 T cell responses during chronic viral infection.
PLoS Pathog. 7:e10020982011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Domachowske JB, Bonville CA, Gao JL,
Murphy PM, Easton AJ and Rosenberg HF: The chemokine
macrophage-inflammatory protein-1 alpha and its receptor CCR1
control pulmonary inflammation and antiviral host defense in
paramyxovirus infection. J Immunol. 165:2677–2682. 2000. View Article : Google Scholar
|
|
19
|
Trifilo MJ, Bergmann CC, Kuziel WA and
Lane TE: CC chemokine ligand 3 (CCL3) regulates CD8(+)-T-cell
effector function and migration following viral infection. J Virol.
77:4004–4014. 2003.
|
|
20
|
Vinay DS and Kwon BS: Immunotherapy
targeting 4-1BB and its ligand. Int J Hematol. 83:23–28. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li CH, Xu KL, Pan XY and Du B: Study on
control of graft-versus-host disease by blocking CD137-CD137L
ligand costimulatory pathway in mice. Zhonghua Xue Ye Xue Za Zhi.
28:93–97. 2007.(In Chinese).
|
|
22
|
Lee SC, Ju SA, Pack HN, et al: 4-1BB
(CD137) is required for rapid clearance of Listeria monocytogenes
infection. Infect Immun. 73:5144–5151. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
DeBenedette MA, Wen T, Bachmann MF, et al:
Analysis of 4-1BB ligand (4-1BBL)-deficient mice and of mice
lacking both 4-1BBL and CD28 reveals a role for 4-1BBL in skin
allograft rejection and in the cytotoxic T cell response to
influenza virus. J Immunol. 163:4833–4841. 1999.PubMed/NCBI
|
|
24
|
Herbein G and O’Brien WA: Tumor necrosis
factor (TNF)-alpha and TNF receptors in viral pathogenesis. Proc
Soc Exp Biol Med. 223:241–257. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Micheau O and Tschopp J: Induction of TNF
receptor I-mediated apoptosis via two sequential signaling
complexes. Cell. 114:181–190. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
de Oliveira Pinto LM, Garcia S, Lecoeur H,
Rapp C and Gougeon ML: Increased sensitivity of T lymphocytes to
tumor necrosis factor receptor 1 (TNFR1)- and TNFR2-mediated
apoptosis in HIV infection: relation to expression of Bcl-2 and
active caspase-8 and caspase-3. Blood. 99:1666–1675.
2002.PubMed/NCBI
|
|
27
|
Gonzalez-Juarrero M, Hattle JM, Izzo A, et
al: Disruption of granulocyte macrophage-colony stimulating factor
production in the lungs severely affects the ability of mice to
control Mycobacterium tuberculosis infection. J Leukoc Biol.
77:914–922. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wan J, Lu LJ, Miao R, et al: Alterations
of bone marrow-derived endothelial progenitor cells following acute
pulmonary embolism in mice. Exp Biol Med (Maywood). 235:989–998.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Govinden R and Bhoola KD: Genealogy,
expression, and cellular function of transforming growth
factor-beta. Pharmacol Ther. 98:257–265. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li MO, Wan YY, Sanjabi S, Robertson AK and
Flavell RA: Transforming growth factor-beta regulation of immune
responses. Annu Rev Immunol. 24:99–146. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Haagmans BL, Teerds KJ, van den
Eijnden-van Raaij AJ, Horzinek MC and Schijns VE: Transforming
growth factor beta production during rat cytomegalovirus infection.
J Gen Virol. 78(Pt 1): 205–213. 1997.PubMed/NCBI
|
|
32
|
Yoo YD, Ueda H, Park K, et al: Regulation
of transforming growth factor-beta 1 expression by the hepatitis B
virus (HBV) X transactivator. Role in HBV pathogenesis. J Clin
Invest. 97:388–395. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kekow J, Wachsman W, McCutchan JA, et al:
Transforming growth factor-beta and suppression of humoral immune
responses in HIV infection. J Clin Invest. 87:1010–1016. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gong Z, Liang AB, Wang LM, et al: The
expression and significance of immunity associated genes mRNA in
patients with pulmonary embolism. Zhonghua Nei Ke Za Zhi.
48:666–669. 2009.(In Chinese).
|