Introduction
Hepatocellular carcinoma (HCC) is the fifth most
common type of malignant tumor worldwide and the third greatest
cause of cancer-related mortality (1–4). HCC
is one of the most aggressive human malignancies and previous data
showed that the five-year survival rate remained extremely poor
(5). HCC is a hypervascular solid
cancer characterized by a high degree of drug resistance (6). Multidrug resistance (MDR) against
chemotherapeutic agents is key in the failure of HCC therapy
(7).
MDR protects cancer cells against a variety of drugs
with different structures and functions. MDR of cancer cells is a
predominant problem in cancer chemotherapy. A number of mechanisms
have been established for the development of MDR, including
expression changes of genes involved in apoptosis, increased enzyme
activity of the glutathione-mediated detoxification pathways and
activation of adenosine triphosphate (ATP) binding cassette
transporters (8,9). The classical mechanism of MDR
development involves the overexpression of P-glycoprotein (P-gp), a
plasma membrane transporter extrudes chemotherapeutic agents from
tumor cells (10–12). Significant effort has been aimed at
the development of MDR reversers. A significant number of compounds
have been identified to function as inhibitors of P-gp and have
been tested for their reversal activities on MDR tumor cells
(13–16). However, a number of these compounds
are currently under clinical evaluation as the majority of tested
compounds exhibit unpredictable pharmacokinetic interactions,
toxicity or insufficient efficacy (17). Novel therapeutic strategies,
including agents that are effective, safe and exhibit low toxicity
are required to reverse the MDR of tumor cells.
Flavonoids, a type of plant polyphenol, are well
established for analgesic, physiological antipyretic and
anti-inflammatory activities and have attracted significant
attention due to their antitumor activities and ability to bind
with P-gp (18–21). Icaritin, a prenylflavonoid, is a
hydrolytic product of icariin, which is derived from the plant
Herba Epimedium which is commonly used in traditional
Chinese medicine. Icaritin exhibits a variety of pharmacological
and biological activities, including antirheumatic and
antidepressant activities; the stimulation of cardiac and neuronal
differentiation (22,23); prevention of steroid-associated
osteonecrosis (24); induction of
human prostate carcinoma PC-3; breast cancer MCF-7 cell growth
inhibition; and cell apoptosis (25,26).
In the present study, the multidrug-resistant
HepG2/ADR subclones were developed and treated with icaritin. The
reversal effect of icaritin on MDR tumor cells was investigated
using the adriamycin (ADR) sensitive HepG2 cells and ADR resistant
HepG2/ADR cells.
Materials and methods
Cell culture and icaritin treatment
HepG2 [a human hepatocellular carcinoma (HCC) cell
line] was purchased from KeyGen (Nanjing, China). Cells were
cultured with Dulbecco’s modified Eagle’s medium (Gibco-BRL,
Carlsbad, CA, USA) supplemented with 10% newborn calf serum
(Gibco-BRL) at 37°C in a humidified atmosphere containing 5%
CO2. An MDR human HCC cell line, HepG2/ADR, was
developed by culturing HepG2 cells in the presence of increasing
concentrations of ADR (0.02, 0.05, 0.1 mg/l; Hisun Pharmaceutical
Co. Ltd., Zhejiang, China). Resistant cells were selected and
resistance was maintained by culturing the cells in medium
supplemented with 0.1 mg/l ADR and labeled HepG2/ADR (0.1).
Stock solution of icaritin (purity, >98%; Yousi
Biotechnology Inc., Shanghai, China), which was further diluted
with cell culture medium before each experiment, was prepared in
dimethyl sulfoxide (DMSO) at a concentration of 10 mM at −20°C. The
final concentration of DMSO in culture was <0.1%.
Measurement of cellular sensitivity to
anticancer drugs
The 3-(4,
5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide(MTT;
Sigma-Aldrich, St. Louis, MO, USA] assay was used to determine drug
sensitivity. HepG2 and HepG2/ADR cells were seeded into 96-well
plates at a concentration of 5 × 103 cells/200 μl/well.
Cells were incubated at 37°C in a humidified 5% CO2
incubator. Following 24 h treatment with specific concentrations of
the anticancer drugs ADR, vincristine, cisplatin and
5-fluorouracil, plates were returned to standard tissue incubator
conditions for an additional 4 h. Medium was removed and cells were
solubilized in 150 μl DMSO. The intensity of formazan was measured
at 490 nm using an automated microplate spectrophotometer (iMark;
Bio-Rad, Hercules, CA, USA). The survival rate was calculated as
(OD value of the treated group/OD value of untreated group) × 100%.
Assays were performed in triplicate in three independent
experiments.
Analysis of MDR reversal activity
Viability of HepG2 and HepG2/ADR cells following
treatment with ADR in the presence (1, 15, 30 μM ) or absence of
icaritin was analyzed by an MTT assay. Following plotting of the
dose-response curve, the IC50, the concentration of drug
inhibiting 50% of cells, was calculated, from which reversal fold
was calculated.
Effect of icaritin on intracellular ADR
accumulation
HepG2 and HepG2/ADR cells were incubated with ADR in
the presence (1, 15, 30 μM ) or absence of icaritin for 4 h. ADR
accumulation in HepG2 and HepG2/ADR cells was assessed by
fluorescence spectrophotometery. The fluorescence was generated
when HepG2 cells were treated with ADR, and the fluorescence
intensity was positively associated with ADR accumulation. ADR
accumulation in HepG2 cells and HepG2/ADR cells was assessed using
a fluorescence spectrophotometer (excitation wavelength, 470 nm;
emission wavelength, 590 nm; F-7000, Hitachi, Chiyoda, Japan).
Quantitative polymerase chain reaction
(qPCR) assay
Cells were frozen in liquid nitrogen and stored at
−80°C for use in qPCR experiments. MDR1 mRNA expression levels were
quantified by qPCR. Total cellular RNA was extracted using TRIzol
reagent (Invitrogen Life Technologies, Carlsbad, CA, USA) according
to the manufacturer’s instructions. Primer sequences used were as
follows: sense: 5′-CATCGAGTCACTGCCTAATAAATA-3′ and
antisense:5′-GCTTCTTGGACAACCTTTTCACT-3′ for MDR1; and sense:
5′-CCTCTATGCCAACACAGTGC-3′ and antisense:
5′-GTACTCCTGCTTGCTGATCC-3′ for β-actin. PCR was performed for 35
cycles, each cycle comprised of denaturation at 95°C for 45 sec,
annealing at 52°C for 45 sec and extension at 72°C for 45 sec,
prior to a final extension at 72°C for 10 min. MDR1 mRNA levels
were analyzed by one-step qPCR with RNA-direct™ SYBR-Green Realtime
PCR Master mix (Toyobo, Osaka, Japan), according to the
manufacturer’s instructions. The amplification was monitored on an
ABI PRISM 7500 real-time PCR apparatus (Applied Biosystems,
Carlsbad, CA, USA).
Western blot analysis
Cells were lysed with ice-cold lysis buffer [50 mM
Tris-HCl (pH 7.4) 150 mM NaCl, 1 mM MgCl2, 100 μg/ml
PMSF and 1% Triton X-100] for 30 min on ice. Total proteins were
dissolved in the supernatant following centrifugation at 13,225 × g
for 5 min at 4°C and protein concentrations were measured in the
supernatants (Protein Assay Dye; Bio-Rad, Hercules, CA, USA). Equal
quantities (40 μg) of lysate proteins were separated on 10%
SDS-PAGE gels and electrophoretically transferred onto
polyvinylidene fluoride membranes. Following blocking with 5%
non-fat dry milk in Tris-buffered saline with Tween-20 (TBST)
buffer [10 mM Tris (pH 7.5) 150 mM NaCl and 0.05% Tween-20] for 2 h
at room temperature, membranes were probed with A 1:1,000 dilution
of anti-target protein (Santa Cruz Biotechnology Inc., Santa Cruz,
CA, USA) or anti-β-actin antibodies (Sigma-Aldrich) at 4°C
overnight, followed by incubation in a 1:5,000 dilution of
horseradish peroxidase-coupled secondary antibody (Sigma-Aldrich).
Protein bands were detected using an enhanced chemiluminescence
detection system (ChemiDoc; Bio-Rad). Band intensity was quantified
by BandScan 5.0 software (Glyko, Hayward, CA, USA). All western
blot analyses were performed at least three times.
Statistical analysis
SPSS version 16.0 software was used (SPSS Inc.,
Chicago, IL, USA). Each assay was performed a minimum of three
times. Data are expressed as the mean ± SD; Student’s t-test and
one-way analysis of variance were used for statistical analyses.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Development of multidrug-resistant
HepG2/ADR subclones
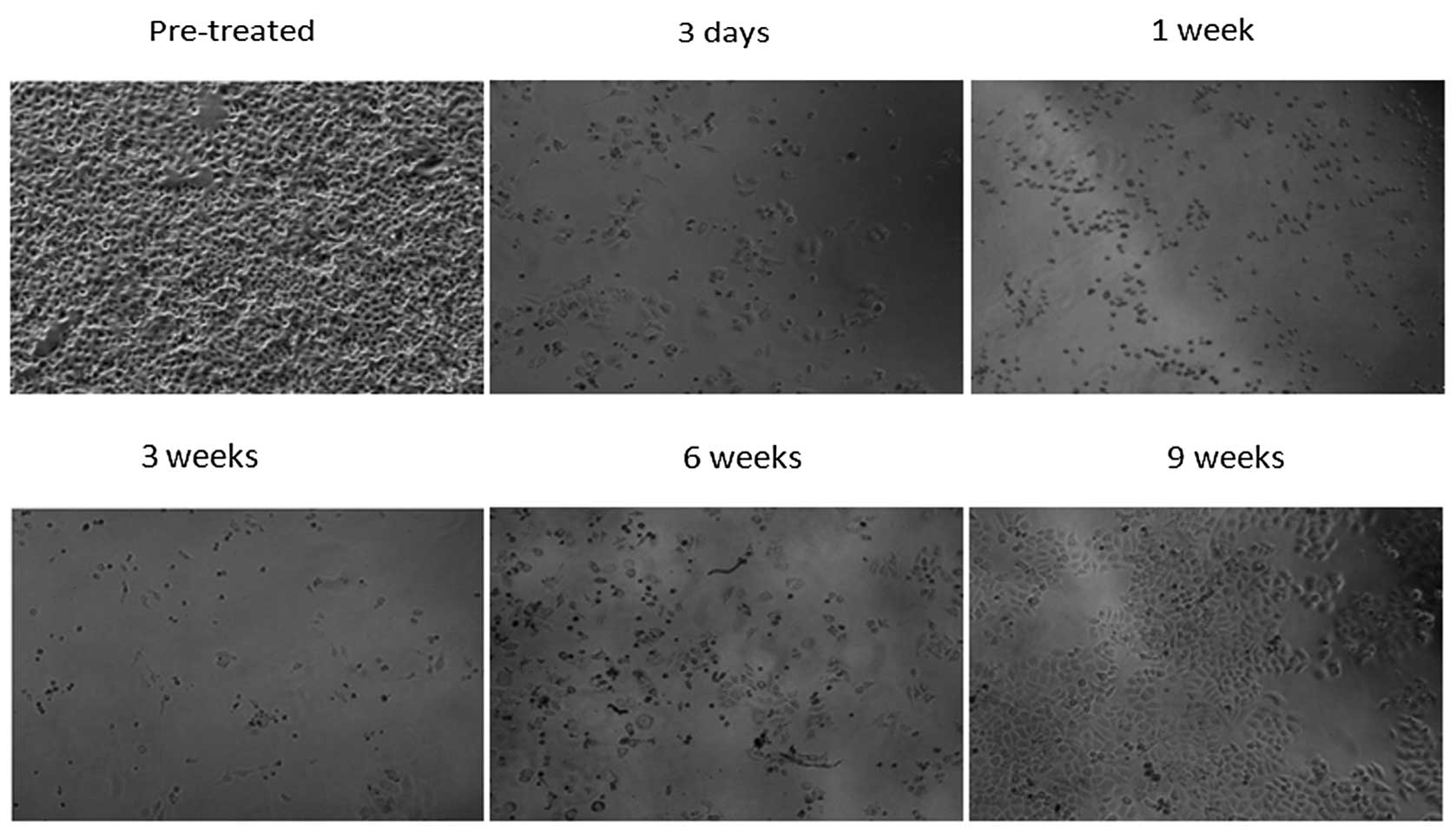
MDR was developed in HepG2 cells by treatment with
increasing concentrations of ADR. HepG2 cells began to exhibit cell
death 24–48 h following treatment with high concentrations of ADR.
Therefore, 0.02 mg/l ADR was added to HepG2 cells and the
morphological changes were observed in cultured cells. Higher
concentrations of ADR were added to the medium once HepG2/ADR
(0.02) cell death was not observed and morphological changes became
stable. A HepG2/ADR (0.1) cell was produced, which required a
minimum of 8–10 weeks culturing (Fig.
1).
Determination of MDR
HepG2/ADR (0.1) cells were investigated for their
resistance against other anticancer drugs using MTT. HepG2/ADR
cells were observed to be resistant to ADR and to multiple
anticancer drugs, including vincristine, cisplatin and
5-fluorouracil. The IC50 of these drugs in HepG2/ADR
cells was significantly higer compared with that in non-resistant
HepG2 cells (Table I). HepG2/ADR
cells were ~25-fold more resistant to ADR in comparison with HepG2
cells.
 | Table IDetermination of IC50 of
various anticancer drugs. |
Table I
Determination of IC50 of
various anticancer drugs.
| Anticancer drugs | HepG2 | HepG2/ADR | Resistant fold |
|---|
| Adriamycin | 0.024±0.007 | 0.596±0.063 | 24.83 |
| Vincristine | 0.012±0.003 | 0.267±0.034 | 22.25 |
| Cisplatin | 0.035±0.42 | 1.285±0.125 | 36.71 |
| 5-Fluorouracil | 0.812±0.102 | 63.092±2.174 | 77.70 |
Icaritin-mediated reversal of HepG2/ADR
cell resistance to ADR
MDR modulating activity of the derivatives was
evaluated by an MTT assay using human HCC cells, HepG2 and ADR
resistant HCC cells (HepG2/ADR). As shown in Table I, the IC50 values of ADR
on HepG2 and HepG2/ADR cells was 0.024±0.007 mg/l and 0.596±0.063
mg/l, respectively, when treated for 48 h. The MDR of HepG2/ADR
cells was 24.83-fold higher compared with the sensitive HepG2
cells. Icaritin was capable of reversing MDR and the sensitivity of
the HepG2/ADR cells to ADR was ~1.65, 2.50 and 7.18 fold higher
when the cells were treated with 1, 15 and 30 μM icaritin (Table II). The results indicated that
icaritin significantly reverses the cytotoxicity of ADR to
HepG2/ADR cells in a dose-dependent manner.
| Table IIEffects of icaritin on the
cytotoxicity of adriamycin to HepG2/ADR cells. |
Table II
Effects of icaritin on the
cytotoxicity of adriamycin to HepG2/ADR cells.
| Concentration,
μM | HepG2/ADR
(IC50) | Reversal fold |
|---|
| 0 | 0.596±0.063 | 1 |
| 1 | 0.362±0.112 | 1.65 |
| 15 | 0.238±0.085 | 2.50 |
| 30 | 0.083±0.011 | 7.18 |
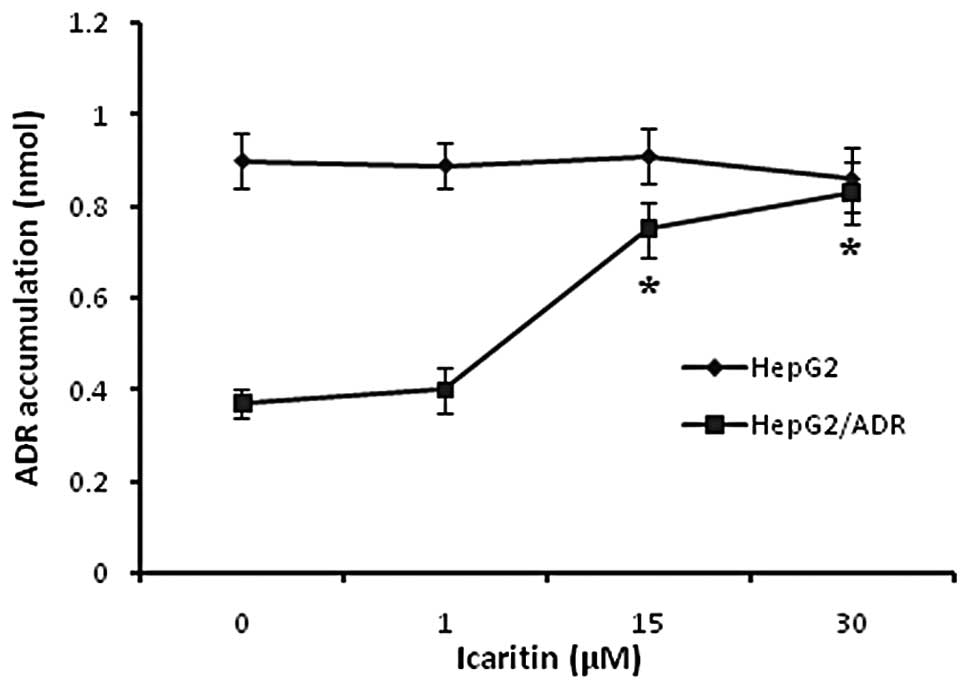
Effect of icaritin on intracellular ADR
accumulation
To investigate the mechanism of the MDR reversal
activity of icaritin, the intracellular ADR accumulation was
examined. As shown in Fig. 2, the
intracellular ADR accumulation in HepG2/ADR cells was 50% lower
compared with that in HepG2 cells. The treatment of icaritin
increased the intracellular ADR accumulation in HepG2/ADR cells at
a specific range. However, it was observed that 0–30 μM icaritin
did not trigger a significant effect in drug sensitive HepG2 cells.
It was hypothesized that the mechanism of the MDR reversal activity
of icaritin may have an association with the increase of
intracellular ADR accumulation.
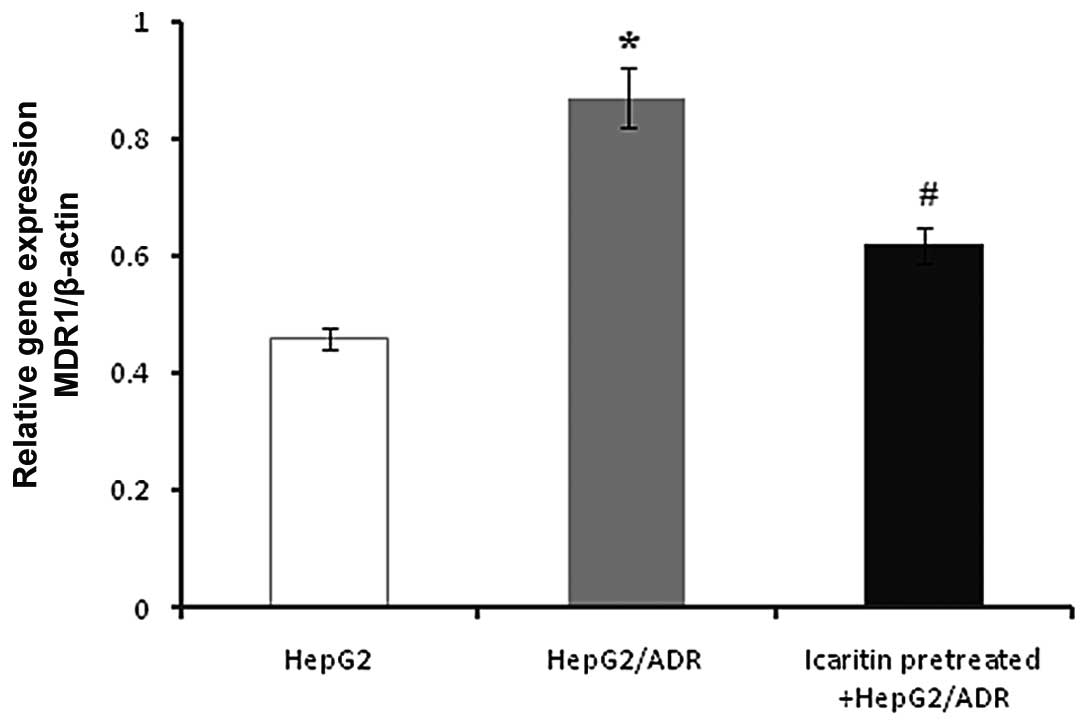
mRNA expression of the MDR1 gene
To determine whether icaritin altered the expression
of the MDR1 gene, mRNA expression of the MDR1 gene was investigated
(Fig. 3). A higher level of MDR1
expression was detected in HepG2/ADR compared with HepG2 cells.
However, when treated with icaritin, the MDR1 level was
significantly decreased in HepG2/ADR cells.
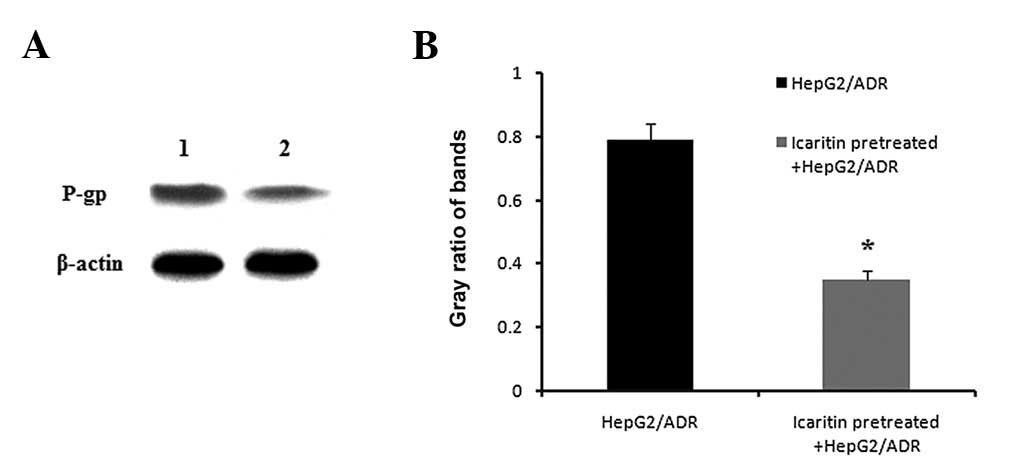
Downregulation of P-gp protein expression
by icaritin in HepG2/ADR cells
The present study showed that icaritin inhibits the
expression of the MDR1 gene in HepG2/ADR cells (Fig. 3). To investigate whether the
mechanism of icaritin on HepG2/ADR cells was responsible for the
expression of P-gp, western blot analysis was performed. Results in
Fig. 4 showed that the expression
of P-gp was significantly repressed by icaritin in HepG2/ADR
cells.
Discussion
MDR of cancer cells is a significant problem in
cancer chemotherapy. P-gp is a plasma membrane transporter, which
extrudes chemotherapeutic drugs from cells using ATP hydrolysis as
an energy source. Overexpression of P-gp has been observed in
numerous cancer cells with MDR (27,28).
The aim of the present study was to identify an effective MDR
reversing agent from Chinese traditional medicine and to gain an
insight into its reversal effect and the molecular mechanisms of
that effect.
A number of tumor cells were observed to develop MDR
when cells were treated with drugs over a period of time (29,30).
ADR is a chemotherapeutic drug, principally used for the treatment
of solid tumors, including HCC (31). It functions via specific
mechanisms; however, resistance to ADR develops in a broad range of
cell lines and results in the loss of therapeutic efficacy
(32,33). Thus, ADR was selected to develop
the current drug resistant HCC cell subclones by culturing the
cells in the presence of increasing concentrations of ADR. Although
the HepG2/ADR cell line was developed by treating the cells with
ADR only, it was observed that multiple drug resistance was
achieved. The IC50 of specific anticancer drugs to the
HepG2/ADR (0.1) subclone was higher compared with the parental
HepG2; 24.83-fold for ADR, 22.25-fold for vincristine, 36.71-fold
for cisplatin and 77.70-fold for 5-fluorouracil. The results showed
that the acquired MDR of the HepG2/ADR (0.1) subclone was
stable.
Icaritin is an active ingredient derivative from the
traditional Chinese medical plant Herba Epimedium. Icaritin
exhibits a variety of pharmacological and biological activities,
including antitumor activity and also induces cell apoptosis in a
number of types of tumor cells (34). However, the effect of icaritin on
the reversal of MDR and its molecular mechanism was unclear.
Therefore, the reversal activity of icaritin on MDR and the
molecular mechanisms involved in this process were investigated. As
shown in Table II, icaritin
treatment resulted in a significant reversal of resistance to ADR
at a concentration of 1, 15 and 30 μM. In addition, the
icaritin-mediated reversal of HepG2/ADR cell resistance to ADR was
observed in a dose-dependent manner. Furthermore, treatment of
icaritin increased the intracellular ADR accumulation in HepG2/ADR
cells at a specific range. It was hypothesized that the mechanism
of the MDR reversal activity of icaritin may be associated with the
increase of intracellular ADR accumulation.
The MDR resistance was due, in part, to the
appearance of P-gp, which exerts an efficient pumping action to
extrude the drugs from tumor cells. The mechanism of P-gp is cell-
or tissue-specific, including the excretion of chemicals into the
bile duct or kidney tubules, protection against toxic xenobiotics
by blocking absorption by the intestine, efflux of steroid hormones
and cholesterol from feces and prevention of chemicals that enter
into the brain through the blood-brain barrier (35,36).
The current results showed that icaritin significantly increased
the intracellular accumulation of ADR and decreased the expression
of the MDR1 level in HepG2/ADR cells compared with drug sensitive
HepG2 cells. It was observed that icaritin may significantly
downregulate the expression of P-gp. Reduction of P-gp expression
may be proposed as a mechanism for specific agents or modulators to
reverse MDR phenotype.
In conclusion, HepG2/ADR multidrug-resistant HCC
cell subclones were successfully established. The current
observations indicate that icaritin effectively reverses MDR, via
the increase of intracellular ADR accumulation, and downregulation
of the MDR1 gene and P-gp protein expression. The present results
indicate that icaritin is a novel and potent MDR reversal agent,
and may be a promising drug for tumor chemotherapy.
Acknowledgements
This work was supported by the Science and
Technology Development Foundation of Nanjing Medical University of
China (No.2011NJMU150), Natural Science Foundation of Jiangsu
province of China (No.BK2009451) and National Natural Science
Foundation of China (No. 30972913).
References
|
1
|
McKillop IH, Moran DM, Jin X and Koniaris
LG: Molecular pathogenesis of hepatocellular carcinoma. J Surg Res.
136:125–135. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Llovet JM, Burroughs A and Bruix J:
Hepatocellular carcinoma. Lancet. 362:1907–1917. 2003. View Article : Google Scholar
|
|
3
|
Trevisani F, Cantarini MC, Wands JR and
Bernardi M: Recent advances in the natural history of
hepatocellular carcinoma. Carcinogenesis. 29:1299–1305. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kusakabe A, Tanaka Y, Orito E, Sugauchi F,
Kurbanov F, Sakamoto T, Shinkai N, Hirashima N, Hasegawa I, Ohno T,
et al: A weak associatioon between occult HBV infection and non-B
non-C hepatocellular carcinoma in Japan. J Gastroenterol.
42:298–305. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
El-Serag HB, Siegel AB, Davila JA, Shaib
YH, Cayton-Woody M, McBride R and McGlynn KA: Treatment and
outcomes of treating of hepatocellular carcinoma among Medicare
recipients in the United States: a population-based study. J
Hepatol. 44:158–166. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wakamatsu T, Nakahashi Y, Hachimine D,
Seki T and Okazaki K: The combination of glycyrrhizin and
lamivudine can reverse the cisplatin resistance in hepatocellular
carcinoma cells through inhibition of multidrug
resistance-associated proteins. Int J Oncol. 31:1465–1472.
2007.
|
|
7
|
Pérez-Tomás R: Multidrug resistance:
retrospect and prospects in anti-cancer drug treatment. Curr Med
Chem. 13:1859–1876. 2006.PubMed/NCBI
|
|
8
|
Clarke R, Currier S, Kaplan O, Lovelace E,
Boulay V, Gottesman MM and Dickson RB: Effect of P-glycoprotein
expression on sensitivity to hormones in MCF-7 human breast cancer
cells. J Natl Cancer Inst. 84:1506–1512. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Choi C: ABC transporters as multidrug
resistance mechanisms and the development of chemosensitizers for
their reversal. Cancer Cell Int. 5:302005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cole SP, Bhardwaj G, Gerlach JH, Mackie
JE, Grant CE, Almquist KC, Stewart AJ, Kurz EU, Duncan AM and
Deeley RG: Overexpression of a transporter gene in a
multidrug-resistant human lung cancer cell line. Science.
258:1650–1654. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cox DS, Scott KR, Gao H, Raje S and
Eddington ND: Influence of multidrug resistance (MDR) proteins at
the blood-brain barrier on the transport and brain distribution of
enaminone anticonvulsants. J Pharm Sci. 90:1540–1552. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Coley HM: Mechanisms and strategies to
overcome chemotherapy resistance in metastatic breast cancer.
Cancer Treat Rev. 34:378–390. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xiang W, Gao A, Liang H, Li C, Gao J, Wang
Q, Shuang B, Zhang J, Yan Y and Wang X: Reversal of
P-glycoprotein-mediated multidrug resistance in vitro by milbemycin
compounds in adriamycin-resistant human breast carcinoma
(MCF-7/adr) cells. Toxicol in Vitro. 24:1474–1481. 2010. View Article : Google Scholar
|
|
14
|
Xu HB, Xu LZ, Li L, Fu J and Mao XP:
Reversion of P-glycoprotein-mediated multidrug resistance by
guggulsterone in multidrug-resistant human cancer cell lines. Eur J
Pharmacol. 694:39–44. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hoffmeyer S, Burk O, von Richter O, Arnold
HP, Brockmöller J, Johne A, Cascorbi I, Gerloff T, Roots I,
Eichelbaum M and Brinkmann U: Functional polymorphisms of the human
multidrug-resistance gene: multiple sequence variations and
correlation of one allele with P-glycoprotein expression and
activity in vivo. Proc Natl Acad Sci USA. 97:3473–3478. 2000.
View Article : Google Scholar
|
|
16
|
Gottesman MM and Ling V: The molecular
basis of multidrug resistance in cancer: the early years of
P-glycoprotein research. FEBS Lett. 580:998–1009. 2006.PubMed/NCBI
|
|
17
|
Li J, Xu LZ, He KL, Guo WJ, Zheng YH, Xia
P and Chen Y: Reversal effects of nomegestrol acetate on multidrug
resistance in adriamycin-resistant MCF7 breast cancer cell line.
Breast Cancer Res. 3:253–263. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Middleton E Jr, Kandaswami C and
Theoharides TC: The effects of plant flavonoids on mammalian cells:
implications for inflammation, heart disease, and cancer. Pharmacol
Rev. 52:673–751. 2000.PubMed/NCBI
|
|
19
|
Li YL, Gan GP, Zhang HZ, Wu HZ, Li CL,
Huang YP, Liu YW and Liu JW: A flavonoid glycoside isolated from
Smilax china L. rhizome in vitro anticancer effects on human
cancer cell lines. J Ethnopharmacol. 113:115–124. 2007.
|
|
20
|
Diaz JG, Carmona AJ, Torres F, Quintana J,
Estévez F and Herz W: Cytotoxic activities of flavonoid glycoside
acetates from Consolida oliveriana. Planta Med. 74:171–174.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wesołowska O: Interaction of
phenothiazines, stilbenes and flavonoids with multidrug
resistance-associated transporters, P-glycoprotein and MRP1. Acta
Biochim Pol. 58:433–448. 2011.PubMed/NCBI
|
|
22
|
Wang Z, Wang H, Wu J, Zhu D, Zhang X, Ou
L, Yu Y and Lou Y: Enhanced co-expression of beta-tubulin III and
choline acetyltransferase in neurons from mouse embryonic stem
cells promoted by icaritin in an estrogen receptor-independent
manner. Chem Biol Interact. 179:375–385. 2009. View Article : Google Scholar
|
|
23
|
Wo YB, Zhu DY, Hu Y, Wang ZQ, Liu J and
Lou YJ: Reactive oxygen species involved in prenylflavonoids,
icariin and icaritin, initiating cardiac differentiation of mouse
embryonic stem cells. J Cell Biochem. 103:1536–1550. 2008.
View Article : Google Scholar
|
|
24
|
Zhang G, Qin L, Sheng H, Wang XL, Wang YX,
Yeung DK, Griffith JF, Yao XS, Xie XH, Li ZR, Lee KM and Leung KS:
A novel semisynthesized small molecule icaritin reduces incidence
of steroid-associated osteonecrosis with inhibition of both
thrombosis and lipid-deposition in a dose-dependent manner. Bone.
44:345–356. 2009. View Article : Google Scholar
|
|
25
|
Wang ZQ and Lou YJ:
Proliferation-stimulating effects of icaritin and desmethylicaritin
in MCF-7 cells. Eur J Pharmacol. 504:147–153. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Huang X, Zhu D and Lou Y: A novel
anticancer agent, icaritin, induced cell growth inhibition, Gl
arrest and mitochondrial transmenbrane potential drop in human
prostate carcinoma PC-3 cells. Eur J Pharmacol. 564:26–36. 2007.
View Article : Google Scholar
|
|
27
|
Hall MD, Brimacombe KR, Varonka MS,
Pluchino KM, Monda JK, Li J, Walsh MJ, Boxer MB, Warren TH, Fales
HM and Gottesman MM: Synthesis and structure-activity evaluation of
isatin-β-thiosemicarbazones with improved selective activity toward
multidrug-resistant cells expressing P-glycoprotein. J Med Chem.
54:5878–5889. 2011.
|
|
28
|
Huang C, Xu D, Xia Q, Wang P, Rong C and
Su Y: Reversal of P-glycoprotein-mediated multidrug resistance of
human hepatic cancer cells by Astragaloside II. J Pharm Pharmacol.
64:1741–1750. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yan F, Wang XM, Liu ZC, Pan C, Yuan SB and
Ma QM: JNK1, JNK2, and JNK3 are involved in P-glycoprotein-mediated
multidrug resistance of hepatocellular carcinoma cells.
Hepatobiliary Pancreat Dis Int. 9:287–295. 2010.PubMed/NCBI
|
|
30
|
Eicher C, Dewerth A, Kirchner B, Warmann
SW, Fuchs J and Armeanu-Ebinger S: Development of a drug resistance
model for hepatoblastoma. Int J Oncol. 38:447–454. 2010.PubMed/NCBI
|
|
31
|
Minotti G, Menna P, Salvatorelli E, Cairo
G and Gianni L: Anthracyclines: molecular advances and
pharmacologic developments in antitumor activity and
cardiotoxicity. Pharmacol Rev. 56:185–229. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Barrand MA, Heppell-Parton AC, Wright KA,
Rabbitts PH and Twentyman PR: A 190-kilodalton protein
overexpressed in non-P-glycoprotein-containing multidrug-resistant
cells and its relationship to the MRP gene. J Natl Cancer Inst.
86:110–117. 1994. View Article : Google Scholar
|
|
33
|
Mehta K: High levels of transglutaminase
expression in doxorubicin-resistant human breast carcinoma cells.
Int J Cancer. 58:400–406. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tong JS, Zhang QH, Huang X, Fu XQ, Qi ST,
Wang YP, Hou Y, Sheng J and Sun QY: Icaritin causes sustained
ERK1/2 activation and induces apoptosis in human endometrial cancer
cells. PLoS One. 6:e167812011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Borst P and Schinkel AH: What have we
learnt thus far from mice with disrupted P-glycoprotein genes? Eur
J Cancer. 32:985–990. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kumar YS, Adukondalu D, Sathish D, Vishnu
YV, Ramesh G, Latha AB, Reddy PC, Sarangapani M and Rao YM:
P-Glycoprotein- and cytochrome P-450-mediated herbal drug
interactions. Drug Metabol Drug Interact. 25:3–16. 2010. View Article : Google Scholar : PubMed/NCBI
|