Introduction
Breast cancer is one of the most common malignant
diseases among females. In China, its morbidity rate increases
annually while affecting patients at younger ages, making breast
cancer an important current health issue. However, with recent
advances in molecular oncology, cancer treatments are capable of
targeting the signaling pathways of tumors. Recently, oncologists
have been targeting the Wnt/β-catenin signaling pathway as a method
of treatment (1,2).
The Wnt signaling pathway regulates cell fate,
survival, proliferation and differentiation at various stages of
mammalian development and pathology. Activation of the canonical
Wnt signaling pathway results in the stabilization and nuclear
accumulation of β-catenin and subsequent gene transcription.
β-catenin, a multifunctional cytoplasmic protein, forms a dynamic
complex with E-cadherin and acts as a key regulator in the Wnt
signaling pathway (3–6) Several studies have demonstrated that
the abnormal expression and mutations of β-catenin are observed in
a wide range of human cancers, and are associated with
tumorigenesis (3–5). Axin is an important negative
regulator of the Wnt signaling pathway, which coordinates β-catenin
phosphorylation and degradation (7). Certain studies have demonstrated that
c-myc and cyclin D1 are important target genes for the Wnt
signaling pathway and their overexpression is associated with the
accumulation of β-catenin in numerous tumor types (3–5).
The mode of regulation of these molecules, with
regard to breast cancer, remains unclear. It is important to
determine their expression and investigate their regulation
mechanisms for the prevention, treatment, and evaluation of human
breast cancer. In the present study, the expression of axin,
β-catenin, c-myc and cyclin D1, and the mutation of the β-catenin
gene were detected. This study also examined the association
between the expression levels and the clinicopathological factors
for axin, β-catenin, c-myc and cyclin D1; axin mechanisms that
regulate β-catenin and the downstream target gene were also
analyzed.
Materials and methods
Patients and specimens
In total, 168 breast cancer tissue specimens were
collected for this study from 2005 to 2010 from the Department of
Pathology, Beijing Tiantan Hospital Affiliated Capital Medical
University (Beijing, China). Of these specimens, 82 were obtained
from the left breast and 86 from the right. The mean age of the
participants was 54.5 years (range, 27–82 years). Tumors were
divided into three groups according to size: 56 patients had a
tumor <2 cm in diameter, 91 patients had tumors 2–5 cm in
diameter and 21 patients had tumors >5 cm in diameter.
Histologically, 126 cases involved invasive ductal carcinoma, not
otherwise specified (IDC, NOS), whereas 42 cases were classified
according to other categories: 15 cases of invasive lobular
carcinomas (ILC), six tubular carcinomas, three medullary
carcinomas, eight mucinous carcinomas, three invasive papillary
carcinomas, one apocrine carcinoma and six neuroendocrine tumors
were observed. According to The Diagnosed and Treated Standard of
Breast Carcinoma (2011 version) (8), 46 cases were classified as grade 1
(well differentiated), 66 as grade 2 (moderately differentiated),
and 56 as grade 3 (poorly differentiated). In terms of clinical
tumor, node, metastasis (TNM) stage, 127 cases were stage I/II and
47 were stage III/IV. There were 43 cases of local and distant
lymph node metastasis, and 115 without metastasis. In total, 44
samples were frozen and stored at −70ºC for DNA sequencing. In
addition, 72 breast intraductal proliferative lesion and 40 normal
breast tissue samples were also obtained.
Immunohistochemistry
All specimens were fixed in 10% neutral formalin,
embedded in paraffin and cut into 4-μm sections for
immunohistochemical staining. The EnVision™ two-step method was
used (DAKO, Hamburg, Germany), as well as the following antibodies:
antibodies against β-catenin, c-myc and cyclin D1, were purchased
from Cell Signaling Technology (Danvers, MA, USA). Prior to
staining, sections were pretreated with microwaves for 18 min in a
0.01 M citrate buffer (pH 6.0) for antigen retrieval.
3,3′-Diaminobenzidine was used as a chromogen and 0.01 M
phosphate-buffered saline (pH 7.4) was substituted for primary
antibodies as a negative control.
Evaluation of immunohistochemical
staining
Under normal conditions, positive staining of
β-catenin was located within the cytomembrane. According to the
method described by Maruyama et al(9), the evaluation of β-catenin staining
is based on its expression in the cytomembrane, cytoplasm, and cell
nucleus. Normal expression was observed in >70% of the cell
expressed in the cytomembrane; reduced expression was observed in
the remainder. More than 10% cells expressed in the cytoplasm or
cell nucleus, which was regarded as ectopic expression. Reduced
expression and ectopic expression were regarded as abnormal
expression.
Positive axin expression was observed in the cell
cytoplasm. The evaluation of axin staining was based on the
staining intensity and staining cell area, according to the method
described by Nakajima et al(10). Staining intensity scores: 0, no
expression; 1, mildly positive; 2, moderately positive; and 3,
markedly positive. Staining cell area scores: 0, <10% of cells
stained; 1, 11–25% stained; 2, 26–50% stained; 3, >50% stained.
A sum of scores >2 was considered to be positive, whereas a
score of <2 was regarded as negative.
A positive expression of cyclin D1 was observed in
the cell nucleus, and c-myc was in the cell nucleus and/or
cytoplasm. The slides showed either negative (−), positive (+), or
overexpression, depending on the count of positive cells; <10%
was regarded as negative, 10–50% was regarded as positive and
>50% was regarded as overexpression.
Detection of β-catenin gene exon 3
mutation
DNA was extracted using the phenol-chloroform method
for polymerase chain reaction (PCR) amplification with the upstream
primer (5′-GCTGATTTGATGGAGTTGGA-3′) and the downstream primer
(5′-GCTACTTGTTCTTGAGTGAA-3′). PCR conditions included denaturation
at 94ºC for 3 min, amplification for 35 cycles for 40 sec at 94ºC,
annealing at 54ºC for 40 sec, and extension at 72ºC for 60 sec.
Finally, the samples underwent extension at 72ºC for 10 min,
followed by cooling at 12ºC. The size of the PCR products was 199
bp. Synthesis of PCR primers, and purification and direct
sequencing of PCR products, was performed by the Beijing Genomics
Institute (Beijing, China).
Statistical analysis
Statistical analysis was performed using the Pearson
χ2 test. The likelihood ratio and Spearman rank
correlation coefficient analysis was performed by SPSS 13.0
software (SPSS, Inc., Chicago, IL, USA). P<0.05 was considered
to indicate a statistically significant difference.
Results
Axin, β-catenin, c-myc and cyclin D1
expression in normal breast tissues, breast intraductal
proliferative lesions, and breast carcinoma tissues
In the 40 cases with normal breast tissue, the
epithelial cells revealed an equally strong membranous expression
of β-catenin protein at the cell-cell boundaries. Axin expression
was positive in 87.5% (35/40) of cases, but the expression of
cyclin D1 and c-myc were negative in all cases. In the 72 cases of
breast intraductal proliferative lesions, the abnormal rate of
β-catenin expression was 44.44% (32/72), and the expression of axin
was positive in 68.06% (49/72) of cases. The overexpression of
cyclin D1 and c-myc were observed in 33.33% (24/72) and 27.78%
(20/72) of cases, respectively. In 168 cases of breast carcinoma,
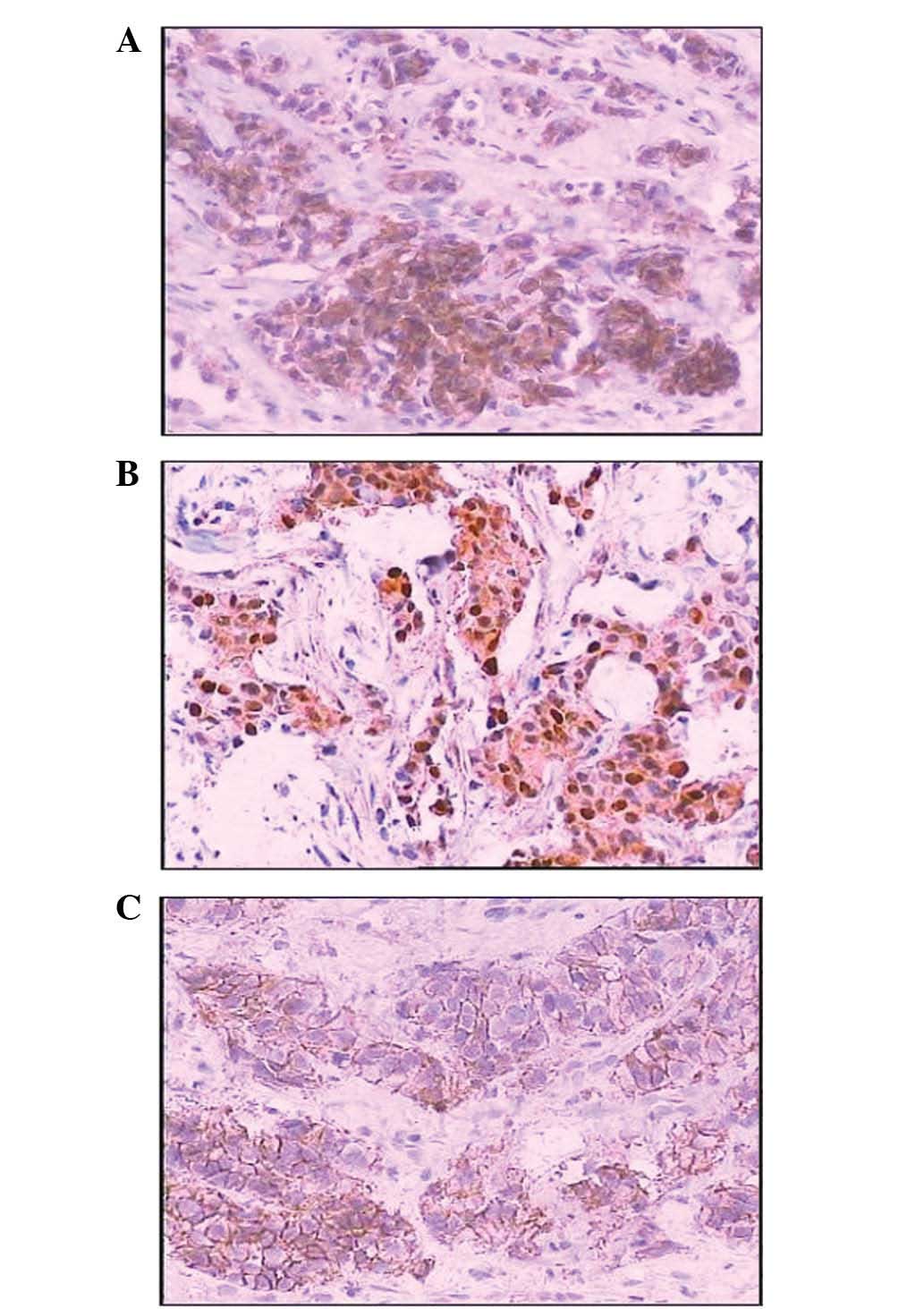
positive axin staining was observed in the cytoplasm in 47.62%
(80/168) of cases, amongst which one sample exhibited nuclear axin
expression and 11 samples showed membrane and cytoplasmic axin
expression (Fig. 1). The abnormal
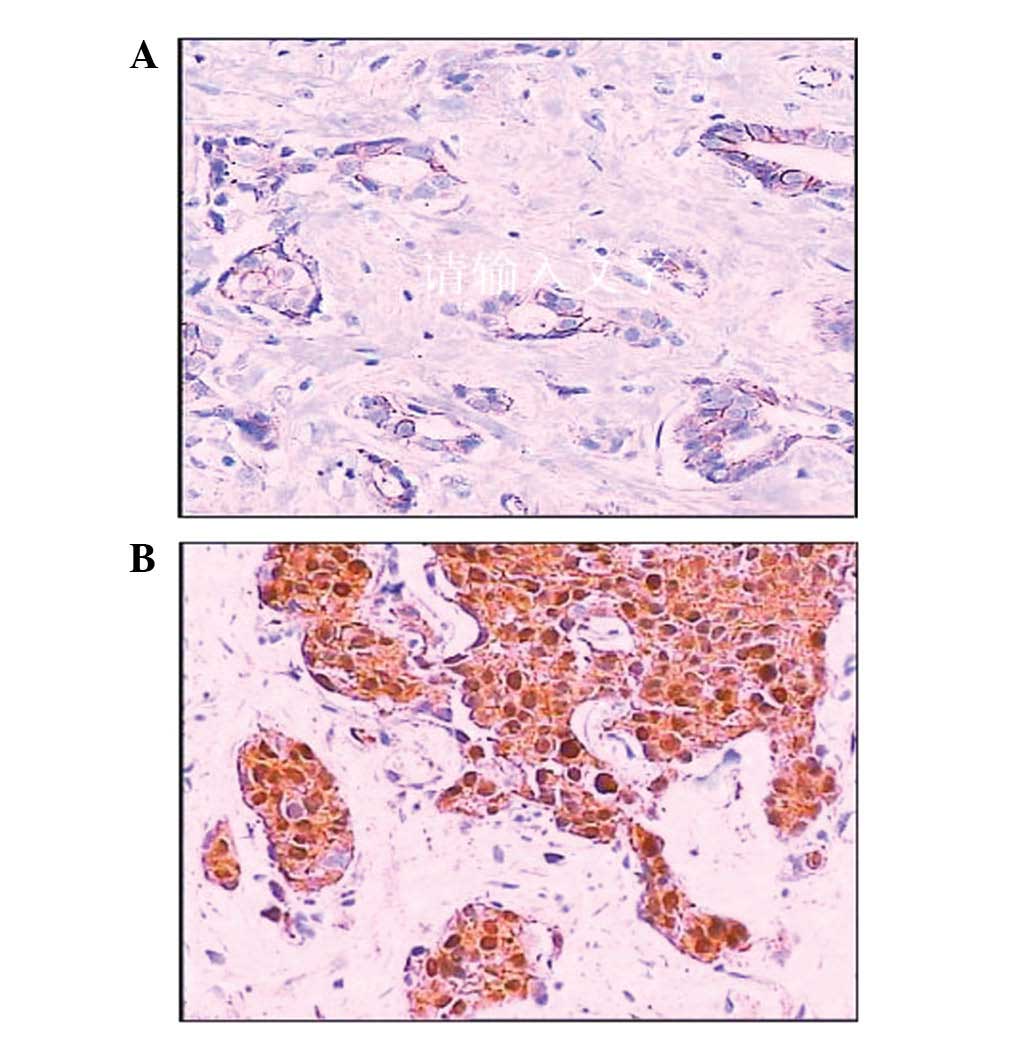
expression rate of β-catenin was 70.83% (119/168). The majority of
β-catenin expression was observed in the cytoplasm, whereas
membranous staining was reduced or disappeared (Fig. 2). The nuclear expression of
β-catenin was observed in 27.38% (46/168) of cases. The positive
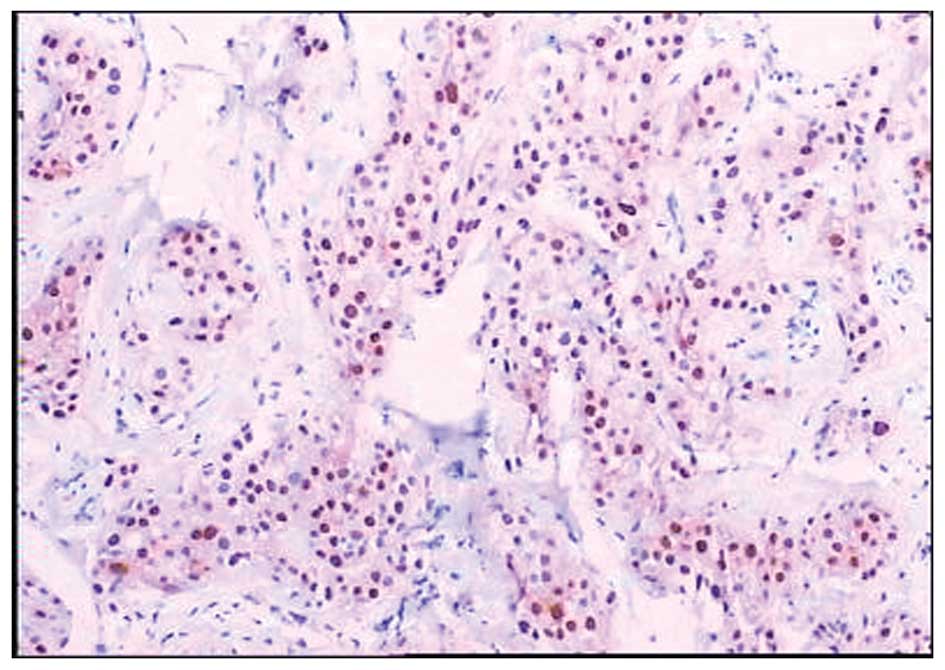
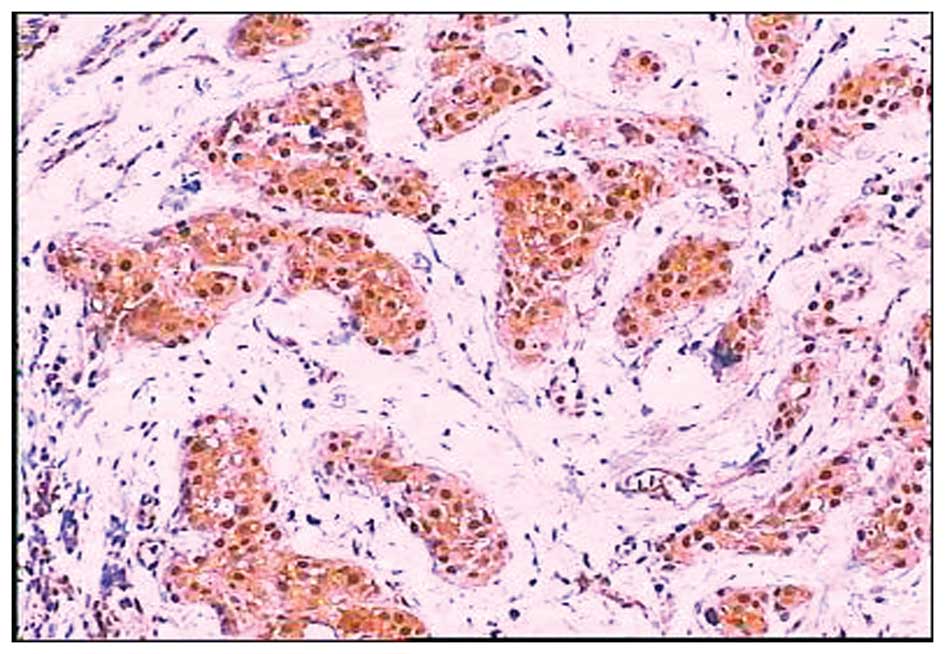
expression of cyclin D1 was observed in the cell nucleus (Fig. 3) and c-myc was observed in the cell
nucleus and/or cytoplasm (Fig. 4).
The overexpression of cyclin D1 and c-myc was observed in 52.98%
(89/168) and 50.00% (84/168) of cases, respectively.
The abnormal expression rate of β-catenin was higher
in breast carcinoma compared with breast intraductal proliferative
lesions (119/168, 70.83% vs. 32/72, 44.44%; P<0.01). Positive
axin expression was lower in breast carcinoma compared with breast
intraductal proliferative lesions (80/168, 47.62% vs. 49/72,
68.06%; P=0.004), and also compared with normal breast tissues
(80/168, 47.62% vs. 35/40, 87.50%;P<0.01). The overexpression of
cyclin D1 and c-myc was higher in breast carcinoma compared with
breast intraductal proliferative lesions (89/168, 52.98% vs. 24/72,
33.33%; P<0.01 and 84/168, 50.00% vs. 20/72, 27.78%;P<0.01),
respectively.
Correlation among axin, β-catenin, c-myc
and cyclin D1 expression, and the clinicopathological
characteristics of breast carcinoma
Axin expression was inversely correlated with tumor
size, histological grade, clinical TNM stage and lymph node
metastasis, but not with patient age or left/right location of the
breast carcinoma (Table I). The
rate of expression in tumors with a diameter of >5 cm was lower
than those 2–5 cm in diameter (5/21, 23.81% vs. 44/91, 48.35%;
P=0.041) and those <2 cm in diameter (5/21, 23.81% vs. 31/56,
55.36%; P=0.038), but no difference in axin expression was observed
between tumors with a diameter of <2 cm and those between 2–5 cm
in diameter (31/56, 55.36% vs. 44/91, 48.35%; P=0.409). The
difference in expression was also observed between tumors with
histologic grade I and grade II (37/46, 80.43% vs. 30/66, 45.45%;
P<0.01), grade II and grade III (30/66, 45.45% vs. 13/56,
23.21%; P<0.01), and grade I and grade III (37/46, 80.43% vs.
13/56, 23.21%; P<0.01). A higher expression rate of axin
expression was observed in patients with clinical TNM stage I/II of
the disease compared with those with stage III/IV of the disease
(69/127, 54.33% vs. 11/41, 26.83%; P<0.01). The positive
expression rate of axin was 24.53% (13/53) in tumors with lymph
node metastasis, in contrast to 58.26% (67/115) in tumors without
lymph node metastasis (P=0.002) (Table
I).
 | Table ICorrelation between axin, β-catenin,
cyclin D1, c-myc and clinicopathological characteristics of breast
cancer. |
Table I
Correlation between axin, β-catenin,
cyclin D1, c-myc and clinicopathological characteristics of breast
cancer.
| | Axin | β-catenin | Cyclin D1 | C-myc |
|---|
| |
|
|
|
|
|---|
| Parameter | n | Positive (%) | P-value | Abnormal (%) | P-value | Positive (%) | P-value | Positive (%) | P-value |
|---|
| Age (years) | | | | | | | | | |
| ≤50 | 65 | 28 (43.08) | 0.349 | 45 (69.23) | 0.717 | 37 (56.92) | 0.416 | 37 (56.92) | 0.154 |
| >50 | 103 | 52 (50.49) | | 74 (71.84) | | 52 (50.49) | | 47 (45.63) | |
| Site | | | | | | | | | |
| Left | 82 | 36 (43.90) | 0.346 | 63 (76.83) | 0.095 | 46 (56.10) | 0.429 | 41 (50.00) | 1.000 |
| Right | 86 | 44 (51.16) | | 56 (65.12) | | 43 (50.00) | | 43 (50.00) | |
| Size (cm) | | | | | | | | | |
| ≤2 | 56 | 31 (55.36) | 0.046 | 32 (57.14) | 0.003 | 21 (37.50) | 0.005 | 18 (32.14) | 0.005 |
| >2≤5 | 91 | 44 (48.35) | | 67 (73.63) | | 52 (57.14) | | 54 (59.34) | |
| >5 | 21 | 5 (23.81) | | 20 (95.24) | | 16 (76.19) | | 12 (57.14) | |
| Histological
type | | | | | | | | | |
| IDC | 126 | 51 (40.48) | 0.001 | 89 (70.63) | 0.922 | 65 (51.59) | 0.532 | 63 (50.00) | 1.000 |
| Others | 42 | 29 (69.05) | | 30 (71.43) | | 24 (57.14) | | 21 (50.00) | |
| Histological
grade | | | | | | | | | |
| I | 46 | 37 (80.43) | <0.01 | 30 (65.22) | 0.003 | 20 (43.48) | 0.009 | 14 (30.43) | 0.002 |
| II | 66 | 30 (45.45) | | 40 (60.61) | | 30 (45.45) | | 33 (50.00) | |
| III | 56 | 13 (23.21) | | 49 (87.50) | | 39 (69.64) | | 37 (66.07) | |
| Clinical stage
TNM | | | | | | | | | |
| I/II | 127 | 69 (54.33) | <0.01 | 82 (64.57) | 0.002 | 60 (47.24) | 0.009 | 60 (47.24) | 0.209 |
| III/IV | 41 | 11 (26.83) | | 37 (90.24) | | 29 (70.73) | | 24 (58.54) | |
| Lymph node
metastasis | | | | | | | | | |
| Positive | 53 | 13 (24.53) | 0.002 | 44 (83.02) | 0.018 | 39 (73.58) | <0.01 | 34 (64.15) | 0.013 |
| Negative | 115 | 67 (58.26) | | 75 (65.22) | | 50 (43.48) | | 50 (43.48) | |
The abnormal expression of β-catenin and the
overexpression of cyclin D1 was correlated with tumor size,
histological grade, clinical TNM stage and lymph node metastasis,
but not with patient age or the location (left/right) of the breast
carcinoma (Table I). The abnormal
expression of β-catenin in tumors >5 cm in diameter was higher
than in those 2–5 cm in diameter (20/21, 95.24% vs. 67/91, 73.63%;
P=0.032) and in those <2 cm in diameter (20/21, 95.24% vs.
32/56, 57.14%; P=0.001). It was also higher in tumors 2–5 cm in
diameter compared with those <2 cm in diameter (67/91, 73.63%
vs. 32/56, 57.14%; P=0.038). The overexpression of cyclin D1 in
tumors <2 cm was less than in those between 2–5 cm in diameter
(21/56, 37.50% vs. 52/91, 57.14%; P=0.021) and those tumors >5
cm in diameter (21/56, 37.50% vs. 16/21, 76.19%; P=0.002). However,
no difference was observed in the overexpression of cyclin D1
between tumors >5 cm and those 2–5 cm in diameter (16/21, 76.19%
vs. 52/91, 57.14%; P=0.107). There was a significant difference in
the expression of β-catenin and cyclin D1, respectively, between
tumors with a histological grade of grade II and grade III (40/66,
60.61% vs. 49/56, 87.50%; P=0.001; 30/66, 45.45% vs. 39/56, 69.64%;
P=0.007), and between those with a grade I and grade III (30/46,
65.22% vs. 49/56, 87.50%; P=0.007; 20/46, 43.48% vs. 39/56, 69.64%;
P=0.008). No difference was observed between grade I and grade II
tumors (30/46, 65.22% vs. 40/66, 60.61%; P=0.620 and 20/46, 43.48%
vs. 30/66, 45.45%; P=0.836), respectively. Significant differences
in the abnormal expression of β-catenin (82/127, 64.57% vs. 37/41,
90.24%; P=0.002) and the overexpression of cyclin D1 (60/127,
47.24% vs. 29/41, 70.73%; P=0.009) were observed between stage I/II
and III/IV of breast carcinoma. Higher rates of abnormal β-catenin
expression (44/53, 83.02% vs. 75/115, 65.22%; P=0.018) and the rate
of overexpression of cyclin D1 (39/53, 73.58% vs. 50/115, 43.48%)
were observed in the primary tumors of patients with lymph node
metastasis compared with those who did not.
The overexpression of c-myc does not correlate with
age, left/right side location of the breast carcinoma, and clinical
TNM stage, but does correlate with tumor size, histologic grade,
and lymph node metastasis (Table
I). The rate of overexpression of c-myc in tumors <2 cm in
diameter was lower than those with tumors 2–5 cm in diameter
(18/56, 32.14% vs. 54/91, 59.34%; P=0.001) and those >5 cm in
diameter (18/56, 32.14% vs. 12/21, 57.14%; P=0.045), but no
difference in expression was observed between tumors >5 cm in
diameter and those between 2–5 cm in diameter (12/21, 57.14% vs.
54/91, 59.34%; P=0.854). Significant differences in the expression
of c-myc were observed between tumors with a histological grade I
and grade II (14/46, 30.43% vs. 33/66, 50.00%; P=0.039) and between
those with grade I and grade III (14/46, 30.43% vs. 37/56, 66.07%;
P<0.01), but no difference was observed between histological
grade II and grade III tumors (33/66, 50.00% vs. 37/56, 66.07%,
P=0.074). The overexpression of c-myc in primary tumors was higher
in patients with lymph node metastasis than in those without
metastasis (34/53, 64.15% vs. 50/115, 43.48%, P=0.013).
Correlation between the abnormal
expression of β-catenin and the overexpression of cyclin D1 and
c-myc in breast carcinoma
The abnormal expression of β-catenin was observed in
74/119 (62.18%) cases with cyclin D1-overexpressing tumors and in
65/119 (54.62%) cases with c-myc overexpressing tumors. There was a
positive and significant association between the abnormal
expression of β-catenin and the overexpression of cyclin D1 in
breast cancer (r=0.288, P<0.01), but the abnormal expression was
not associated with the overexpression of c-myc in breast cancer
(P=0.062) (Table II).
| Table IICorrelation between the expression of
β-catenin, cyclin D1 and c-myc in breast cancer. |
Table II
Correlation between the expression of
β-catenin, cyclin D1 and c-myc in breast cancer.
| | Cyclin D1 | C-myc |
|---|
| |
|
|
|---|
| β-catenin | n | + | − | + | − |
|---|
| Abnormal | 119 | 74 | 45 | 65 | 54 |
| Normal | 49 | 15 | 34 | 19 | 30 |
| P-value | | <0.01 | | 0.062 | |
Correlation between the expression of
axin and β-catenin in breast carcinoma
An increased rate of axin expression was observed in
tumors with a nuclear expression of β-catenin compared with those
without β-catenin nuclear expression (8/46, 17.39% vs. 72/122,
59.02%; P<0.01), but the expression of axin was similar in
tumors with abnormal (reduced) expression of β-catenin to those
with normal β-catenin expression (57/119, 47.90% vs. 23/49, 46.94%;
P=0.910) (Table III).
| Table IIICorrelation between the expression of
axin and β-catenin in breast cancer. |
Table III
Correlation between the expression of
axin and β-catenin in breast cancer.
| β-catenin | β-catenin |
|---|
|
|
|
|---|
| Axin | Reduced expression
in membrane | Normal expression
in membrane | Positive in
nuclear | Negative in
nuclear |
|---|
| + | 57 | 23 | 8 | 72 |
| − | 62 | 26 | 38 | 50 |
| P-value | 0.910 | | <0.01 | |
Detection of β-catenin gene exon 3
mutation
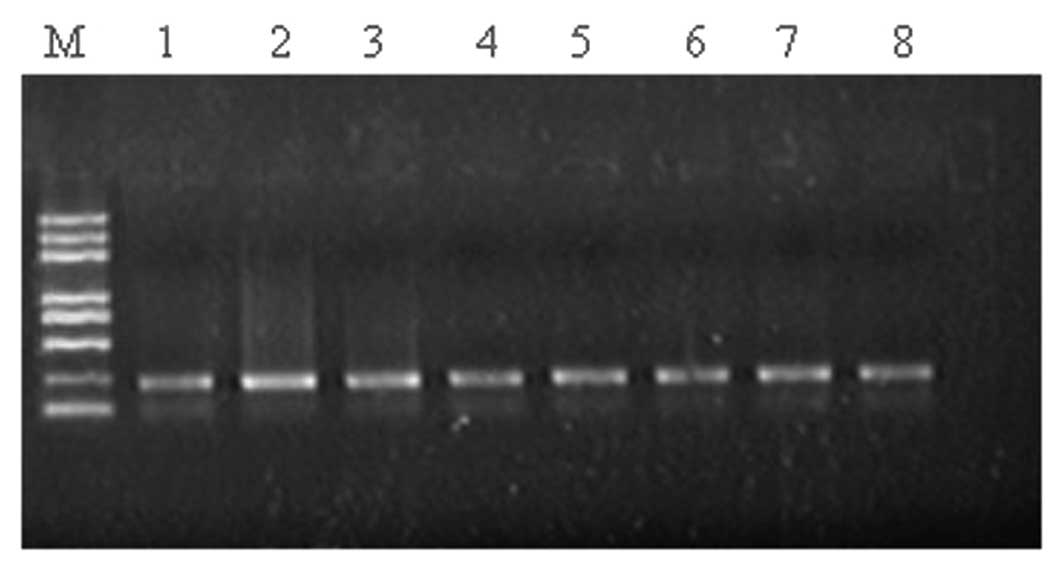
The PCR products of β-catenin displayed bands at
~200 bp (Fig. 5). However,
mutations in the β-catenin gene were not detected in 44 cases of
breast carcinoma.
Discussion
β-catenin was originally identified as a cytoplasmic
protein that links E-cadherin and α-catenin to the cytoskeleton
constituting the E-cadherin-catenin complex in order to maintain
normal epithelial polarity and intercellular adhesion, and to
regulate cellular differentiation and proliferation (6). β-catenin has been reported to be a
critical member of the canonical Wnt/β-catenin pathway, which
promotes cell fate determination, proliferation and survival. In
the absence of Wnt signals, β-catenin in the cytoplasm is
sequestered into a complex with the adenomatous polyposis coli
tumor suppressor, axin and a serine threonine glycogen synthetase
kinase-3β (GSK-3β), enabling the phosphorylation and degradation of
free β-catenin by a ubiquitin-proteasome system. The activation of
canonical Wnt signaling inhibits the degradation of β-catenin,
allowing it to accumulate within the cytoplasm and enter the
nucleus, where it interacts with Lef/Tcfs to regulate gene
expression. Wnt/β-catenin signaling has been documented in a wide
range of cancers, including colorectal and gastric cancer,
melanoma, and tumors derived from hepatic, breast and prostate
tissue. Numerous studies have suggested that the activation of the
Wnt/β-catenin signaling pathway is important in human tumorigenesis
(3–5). However, reports regarding the
expression of β-catenin in human breast cancer have been
controversial. Bankfalvi et al(11) reported that the abnormal expression
of β-catenin is an early event in breast cancer, as reduced
β-catenin membrane expression was noted during the development of
invasive carcinoma from normal breast tissue. Other studies have
reported that the abnormal expression of β-catenin was associated
with a poor prognosis in breast cancer (12–15),
and β-catenin was deemed to be an independent prognostic factor.
Chung et al(16)
demonstrated that patients with tumors coexpressing high levels of
abnormal β-catenin and p53 had poor survival outcomes, therefore
β-catenin may not be an independent prognostic factor. Wang et
al(17) observed a correlation
between β-catenin and Her2 expression, and this correlation may
synergistically promote lymph node metastasis in breast cancer. In
the present study study, >70% of abnormal β-catenin expression
was observed in breast carcinomas, which was higher than in breast
intraductal proliferative lesions. The nuclear expression rate of
β-catenin was 27.38% (46/168). Abnormal expression was
significantly associated with histological grade, clinical TNM
stage and lymph node metastasis. The abnormal expression of
β-catenin may be important in breast carcinogenesis as an
independent prognostic factor.
Mutations of β-catenin, which result in the
accumulation of β-catenin in the cytoplasm and nucleus, are
identified in a wide variety of human cancers, including colon
cancer, desmoid, gastric cancer, hepatocarcinoma, medulloblastoma,
melanoma, ovarian, pancreatic and prostate cancers (18,19).
The β-catenin gene is located on chromosome 3p21, it is 23.2 kb and
has 16 exons. Exon 3 is clustered in the domain that contains an
ubiquitination motif (codons 32 through to 37) and four conserved
GSK-3β phosphorylation sites (Ser33, Ser37, Thr41 and Ser45).
Normally, β-catenin gene mutations occur in exon 3 (18). Zhuang et al(20) identified mutations of the β-catenin
gene in 1,3-butadiene-induced B6C3F1 mice mammary adenocarcinomas
and identified missense mutations in codons 33, 34, and 41 of
β-catenin in 18% (3/17) of tumors. By contrast, such mutations
rarely exist in human breast cancer (21–23).
β-catenin mutations were detected in 45% of breast fibromatosis
cases in the study (24). PCR and
direct sequencing on DNA from 44 breast carcinoma tissue samples
was conducted, but no mutations were identified. It is possible
that the mutation in exon 3 of the β-catenin gene in human breast
cancer may be a rare event and not a predominant mechanism for
abnormal β-catenin expression. Other mechanisms may result in the
abnormal expression of β-catenin, which requires further study.
Axin was originally identified as the product of the
mouse gene termed Fused (25).
Axin, a scaffold protein, is a multidomain protein that interacts
with multiple proteins and functions as a negative regulator of Wnt
signaling by downregulating β-catenin levels. It is also important
in the JNK signaling pathway. Axin controls diverse cellular
functions in proliferation, fate determination and the suppression
of tumorigenesis (7). Several
studies have identified axin expression in various human cancers,
and have demonstrated an inverse association between axin
expression and tumor invasion and metastasis (26–28).
However, there have been few studies regarding axin expression in
human breast cancer, and knowledge regarding axin and β-catenin
expression in human breast cancer is limited. In this study, the
positive expression of axin was 47.62% in breast carcinoma, which
was lower than the positive expression in breast intraductal
proliferative lesions and in normal breast tissues. Axin expression
was inversely correlated with tumor size, histological grade,
clinical TNM stage and lymph node metastasis. The expression was
negatively correlated with β-catenin nuclear expression, but was
not correlated with the reduced membranous expression of β-catenin.
It has been postulated that axin expression may downregulate the
accumulation of β-catenin in the nucleus and inhibit the loss of
differentiation and proliferation of tumor cells in breast
carcinoma. In addition, Axin is considered to be a cytoplasmic
protein. Cong et al(29)
reported that axin functions as a molecular chaperone for β-catenin
and that the nuclear-cytoplasmic shuttling of axin regulates the
nuclear-cytoplasmic distribution of β-catenin. In the present
study, nuclear axin expression was only identified in one sample,
indicating that axin may act as a shuttle in and out of the
nucleus. However, the reason for infrequent nuclear axin expression
in tumor tissues remains unknown. Cliffe et al(30) reported that the dishevelled
protein, an upstream component of the Wnt/β-catenin pathway, may
relocate axin from the cytoplasm to the plasma membrane and allow
the subsequent inactivation of the axin complex by Wingless
signaling. In the present study, 11 samples were observed to have
membranous and cytoplasmic axin expression, thus further
investigation is required.
Cyclin D1 and c-myc are oncogenes involved in cell
proliferation, differentiation and apoptosis. As target genes of
the Wnt signaling pathway, the amplification and/or overexpression
of cyclin D1 and c-myc in tumor cells are extremely common,
indicating that they are essential components during
carcinogenesis. In breast carcinoma, Ozaki et al(31) observed a strong correlation between
the overexpression of β-catenin, and cyclin D1 and c-myc. Roh et
al(32) reported that the abnormal expression of β-catenin may
upregulate the overexpression of c-myc. Lin et al(12) observed that the abnormal expression
of β-catenin was associated with the overexpression of cyclin D1,
whereas Lim et al(13) did
not. In the present study, the levels of overexpression of cyclin
D1 and c-myc in breast carcinoma were 53 and 50%, respectively,
which were higher than those in the breast intraductal
proliferative lesions. The overexpression of cyclin D1 was
significantly correlated with tumor size, histological grade,
clinical TNM stage and lymph node metastasis, whereas the
overexpression of c-myc was correlated with tumor size,
histological grade and lymph node metastasis. There was a positive
correlation between the abnormal expression of β-catenin and the
overexpression of cyclin D1 in breast cancer, but that association
was not observed between the abnormal expression of β-catenin and
the overexpression of c-myc. The abnormal expression of β-catenin
may be key in the carcinogenesis and progression of human breast
carcinoma by upregulating the expression of cyclin D1. Cyclin D1
and c-myc overexpression may be considered to be useful markers for
determining metastasis and providing a prognosis of human breast
carcinoma and to aid in guiding treatment.
References
|
1
|
Paul S and Dey A: Wnt signaling and cancer
development: therapeutic implication. Neoplasma. 55:165–176.
2008.PubMed/NCBI
|
|
2
|
Prosperi JR and Goss KH: A Wnt-ow of
opportunity: targeting the Wnt/beta-catenin pathway in breast
cancer. Curr Drug Targets. 11:1074–1088. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Miller JR: The Wnts. Genome Biol. 3:1–15.
2002.
|
|
4
|
Polakis P: Wnt signaling and cancer. Genes
Dev. 14:1837–1851. 2000.
|
|
5
|
Lustig B and Behrens J: The Wnt signaling
pathway and its role in tumor development. J Cancer Res Clin Oncol.
129:199–221. 2003.PubMed/NCBI
|
|
6
|
Van Aken E, De Wever O, Correia da Rocha
AS and Mareel M: Defective E-cadherin/catenin complexes in human
cancer. Virchows Arch. 439:725–751. 2001.PubMed/NCBI
|
|
7
|
Luo W and Lin SC: Axin: a master scaffold
for multiple signaling pathways. Neurosignals. 13:99–113. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Böcker W: WHO classification of breast
tumors and tumors of the female genital organs: pathology and
genetics. Verh Dtsch Ges Pathol. 86:116–119. 2002.(In German).
|
|
9
|
Maruyama K, Ochiai A, Akimoto S, et al:
Cytoplasmic beta-catenin as a predictor of hematogenous metastasis
in human colorectal cancer. Oncology. 59:302–309. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nakajima M, Fukuchi M, Miyazaki T, et al:
Reduced expression of Axin correlates with tumour progression of
oesophageal squamous cell carcinoma. Br J Cancer. 88:1734–1739.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bankfalvi A, Terpe HJ, Breukelmann D, et
al: Immunophenotypic and prognostic analysis of E-cadherin and
beta-catenin expression during breast carcinogenesis and tumour
progression: a comparative study with CD44. Histopathology.
34:25–34. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lin SY, Xia W, Wang JC, et al:
Beta-catenin, a novel prognostic marker for breast cancer: its
roles in cyclin D1 expression and cancer progression. Proc Natl
Acad Sci USA. 97:4262–4266. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lim SC and Lee MS: Significance of
E-cadherin/beta-catenin complex and cyclin D1 in breast cancer.
Oncol Rep. 9:915–928. 2002.PubMed/NCBI
|
|
14
|
Dolled-Filhart M, McCabe A, Giltnane J, et
al: Quantitative in situ analysis of beta-catenin expression in
breast cancer shows decreased expression is associated with poor
outcome. Cancer Res. 66:5487–5494. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
López-Knowles E, Zardawi SJ, McNeil CM, et
al: Cytoplasmic localization of beta-catenin is a marker of poor
outcome in breast cancer patients. Cancer Epidemiol Biomarkers
Prev. 19:301–309. 2010.PubMed/NCBI
|
|
16
|
Chung GG, Zerkowski MP, Ocal IT, et al:
beta-Catenin and p53 analyses of a breast carcinoma tissue
microarray. Cancer. 100:2084–2092. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang Z, Ren Y, He JJ, et al: Relationship
between ectopic expression of Wnt/beta-catenin signaling pathway
and Her-2 overexpression in human breast carcinoma. J Xi’an
Jiaotong Univ (Med Sci). 30:587–591. 2009.
|
|
18
|
Kikuchi A: Tumor formation by genetic
mutations in the components of the Wnt signaling pathway. Cancer
Sci. 94:225–229. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
He YJ, Jia XS, Qing SKZ, et al: The study
on p53 and beta-catenin gene mutations in NK/T cell lymphoma. Chin
J Clin Oncol. 31:1385–1388. 2004.
|
|
20
|
Zhuang SM, Wiseman RW and Söderkvist P:
Frequent mutations of the Trp53, Hras1 and beta-catenin (Catnb)
genes in 1,3-butadiene-induced mammary adenocarcinomas in B6C3F1
mice. Oncogene. 21:5643–5648. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Howe LR and Brown AM: Wnt signaling and
breast cancer. Cancer Biol Ther. 3:36–41. 2004. View Article : Google Scholar
|
|
22
|
Ozaki S, Ikeda S, Ishizaki Y, et al:
Alterations and correlations of the components in the Wnt signaling
pathway and its target genes in breast cancer. Oncol Rep.
14:1437–1443. 2005.PubMed/NCBI
|
|
23
|
Kizildag S, Zengel B, Vardar E, et al:
beta-catenin gene mutation in invasive ductal breast cancer. J
BUON. 13:533–536. 2008.PubMed/NCBI
|
|
24
|
Abraham SC, Rcynolds C, Lee JH, et al:
Fibromatosis of the breast and mutations involving the
APC/beta-catenin pathway. Hum Pathol. 33:39–46. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zeng L, Fagotto F, Zhang T, et al: The
mouse Fused locus encodes Axin, an inhibitor of the Wnt signaling
pathway that regulates embryonic axis formation. Cell. 90:181–192.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xu HT, Wang L, Lin D, et al: Abnormal
beta-catenin and reduced axin expression are associated with poor
differentiation and progression in non-small cell lung cancer. Am J
Clin Pathol. 125:534–541. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ishizaki Y, Ikeda S, Fujimori M, et al:
Immunohistochemical analysis and mutational analyses of
beta-catenin, Axin family and APC genes in hepatocellular
carcinomas. Int J Oncol. 24:1077–1083. 2004.PubMed/NCBI
|
|
28
|
Jin LH, Shao QJ, Luo W, et al: Detection
of point mutations of the Axin1 gene in colorectal cancers. Int J
Cancer. 107:696–699. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cong F and Varmus H: Nuclear-cytoplasmic
shuttling of Axin regulates subcellular localization of
beta-catenin. Proc Natl Acad Sci USA. 101:2882–2887. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cliffe A, Hamada F and Bienz M: A role of
Dishevelled in relocating Axin to the plasma membrane during
wingless signaling. Curr Biol. 13:960–966. 2003. View Article : Google Scholar
|
|
31
|
Roh MS, Hong SH, Jeong JS, et al: Gene
expression profiling of breast cancer with emphasis of beta-catenin
regulation. J Korean Med Sci. 19:275–282. 2004. View Article : Google Scholar : PubMed/NCBI
|