Introduction
Inflammatory response is a major defense mechanism
against pathogens and chemical or mechanical injury. These
phenomena are mediated by inflammatory cells including macrophages.
Activated macrophages produce reactive oxygen species (ROS) and
nitric oxide (NO), and cause substantial oxidant injury to
surrounding tissue (1–3). Extensive laboratory and clinical
evidence indicates that chronic inflammation contributes to cancer
(4). Oxidative stress-induced
neuron injury induces a variety of neurodegenerative diseases
including Alzheimer’s and Parkinson’s disease as well as cerebral
ischemia (5).
The dried roots of Angelica gigas Nakai (AGN)
have been traditionally used in Oriental Medicine. It is known in
Korean as ‘Cham-dang-gui’. Several coumarin derivatives and a
pectic polysaccharide were isolated from AGN (6). These derivatives are known to inhibit
cancer cell adhesion and invasion (7), have anti-diabetic activity (8), suppress androgen-induced and
-independent cell proliferation, and cause anti-inflammatory
activities (9) and neuroprotective
activity (10).
Panax ginseng (PG), one of the most
well-known herbal medicines, has been commonly used in East Asia.
Total saponins and ginsenosides are the major active components of
PG (11). Ginseng has various
biological activities, including inhibition of tumor-induced
angiogenesis, prevention of tumor invasion and metastasis (12), as well as anti-infective (13), anti-diabetic (14), anti-inflammatory (15), and neuroprotective activities
(16).
Rhus verniciflua Stokes (RVS) has
traditionally been used as an ingredient in East Asian Medicine for
the treatment of gastritis, stomach cancer and atherosclerosis. The
compounds identified from RVS are as follows: Gallic acid,
protocatechuic acid, quercetin, fustin, fisetin, sulfuretin and
butein (17). RVS protects from
oxidative damage by scavenging reactive oxygen species (ROS)
(18) and it has
anti-proliferative, anti-cancer and anti-inflammatory effects
(19).
In the present study, the effect of a mixture of
three medicinal plants AGN, PG, RVS (APR) on
lipopolysaccharide (LPS)-induced inflammatory responses in the
mouse macrophage cell line RAW264.7 was evaluated. It was assessed
whether an ethanolic (EtOH) extract of APR suppresses LPS-induced
inflammatory responses in RAW264.7. The present study also
investigated whether APR exhibits anti-proliferative activity
regulating intracellular molecules associated with cell survival
and apoptosis.
Materials and methods
Cell culture
RAW264.7 mouse macrophage cells were obtained from
the Korea Cell Line Bank (Seoul, Korea). Cells were cultured in
Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10%
heat-inactivated fetal bovine serum (FBS) and 1% antibiotics
(penicillin-streptomycin) at 37°C in a 5% CO2 humidified
incubator.
Extraction of medicinal plants (APR)
Medicinal plants used in the present study were
purchased from Omniherb (Daekoo, Gyeongsangbuk-do, Korea). The
powder with a mass of 100 g (AGN and PG, root and RVS, bark) was
extracted twice with 80% (v/v) ethanol (Duksan Pharmaceutical Co.,
Ltd., Ansan, Republic of Korea) by using an Ultra-sonicator
(Branson, Danbury, CT, USA) for 30 min at room temperature. The
alcoholic extract was filtered through a 0.22 μm filter, the
solvent was evaporated at 40°C and the residue freeze-dried. The
yields of the extracts were 38.2, 26.4 and 13.7% (w/w) for AGN, PG
and RVS, respectively. The plant extract mixture was prepared as
AGN : PG : RVS = 1:1:0.1.
Cell proliferation assay
The cell proliferation rate was determined using the
water soluble tetrazolium (WST) assay following treatment with APR.
The WST assay is based on the cleavage of the yellow tetrazolium
salt to purple formazan crystals by metabolically active live
cells.
RAW264.7 cells (1×104 cells/well) were
seeded in 96-well plates, incubated overnight and treated with APR.
Following 24 h of incubation, 10 μl WST solution was added to 100
μl cell culture medium and the plates were incubated for a further
2 h. The optical density was determined at 490 nm using an ELISA
reader (Molecular Devices, Palo Alto, CA, USA).
Cell death assay
Cell death following APR treatment was determined
using the trypan blue assay. Trypan blue selectively stains dead
cells. RAW264.7 cells were treated with APR for 12 and 24 h,
respectively. The cells were then suspended and stained with trypan
blue solution (Sigma Aldrich; St. Louis, MO, USA). The cell number
was determined by counting using a hemocytometer.
Cell surface observation
Cells were seeded into 60-mm culture dishes at a
density of 3×105 cells/dish. The following day, the
cells were treated with APR for 12 h. The cell surface was observed
by capturing an image using a camera (Olympus) attached to a
microscope.
Mitochondrial membrane potential
analysis
The loss of mitochondrial membrane potential is a
specific characteristic of apoptosis.
5,5′,6,6′-Tetrachloro-1,1′,3,3′-tetraethylbenzimidazolylcarbocyanine
iodide (JC-1) is a membrane-permeable dye widely used for
determining the mitochondrial membrane potential using flow
cytometry and fluorescent microscopy. Cells were seeded into 60 mm
culture dishes at a density of 3×105 cells/dish. The
following day, the cells were treated with APR for 24 h. Cells were
harvested from each culture dish, washed with phosphate-buffered
saline (PBS), suspended in PBS containing 2 μM JC-1 and incubated
for 30 min at 37°C in the dark. The data were analyzed by
FACSCalibur flow cytometry (BD Biosciences, Franklin Lakes, NJ,
USA).
Assessment of intracellular reactive
oxygen species (ROS) levels
The molecule 2′,7′-dichlorofluorescein diacetate
(DCFH-DA) freely permeates cells, and following incorporation into
cells, is converted into the fluorescent 2,7-dichlorofluorescein
(DCF) by oxidative substances, revealing the intracellular
production of redox-active substances. It has been widely used to
investigate oxidative damage in intact cells. Cells were seeded
into 35 mm culture dishes containing glass coverslips. Following
different pretreatments, the cells were washed with PBS and
incubated with 20 μM DCFH-DA for 30 min at 37°C in the dark.
Following washing with cold PBS, the fluorescence was captured
using a laser confocal scanning microscope (LSM 510; Carl Zeiss,
Jena, Germany) and a FACSCalibur flow cytometer (BD Biosciences,
San Jose, CA, USA). DCF fluorescence was measured at an excitation
wavelength of 488 nm and emission at 515–540 nm.
RNA extraction and reverse transcription
polymerase chain reaction (RT-PCR)
Cells were collected by centrifugation and RNA was
extracted using an Invitrogen kit (Grand Island, NY, USA),
according to the manufacturer’s instructions. Primers were designed
for RT-PCR for nitric oxide synthase isoform (iNOS) and
prostaglandin endoperoxide synthase 2 (Cox-2). The primer sequences
were as follows: (sense) 5′-GGAGAGACTATCAAGATAGT-3′ and (antisense)
5′-ATGGTCAGTAGACTTTTACA-3′ for COX-2; (sense)
5′-AATGGCAACATCAGGTCGGCCATCACT-3′ and (antisense)
5′-GCTGTGTGTCACAGAAGTCTCGAACTC-3′ for iNOS; (sense)
5′-GAGGGGCCATCCACAGTCTTC-3′ and (antisense)
5′-CATCACCATCTTCCAGGAGCG-3′ for glyceraldehyde 3-phosphate
dehydrogenase (GAPDH). The sequencing involved 30 cycles with
denaturation at 94°C for 45 sec, annealing at 55°C for 45 sec and
extension at 72°C for 45 sec. The resulting PCR products were
resolved on 1% agarose gels containing ethidium bromide.
Western blot analysis
Whole cell lysates from cells treated with DMSO and
APR in the presence or absence of LPS were prepared by washing with
ice-cold PBS and lysis with a radioimmunoprecipitation assay (RIPA)
buffer. Equal amounts of protein (30 μg) from the cell lysates were
boiled for 5 min in SDS-PAGE sample buffer, resolved by 10%
SDS-PAGE, transferred to nitrocellulose membranes at 80 V for 1.5 h
and visualized using western blot analysis and
chemiluminescence.
Results
Effects of APR on cell viability
The anti-proliferative effects of APR were assessed
on RAW264.7 mouse macrophage cells. First, the inhibitory effect of
APR on cell proliferation was investigated using the WST assay
(Fig. 1a). For this purpose, the
cells were treated with APR at concentrations of 0–1,000 μg/ml and
1 μg/ml LPS for 12 h. LPS alone did not show any proliferative
activity in RAW264.7 cells. However, APR significantly inhibited
cell proliferation at concentrations of 250–1,000 μg/ml, suggesting
that APR inhibits the growth of RAW264.7 cells. Second, the cell
death rate was determined using the trypan blue assay during APR
treatment (Fig. 1b). At 24 h of
incubation, APR significantly decreased the percentage of surviving
cells.
Changes in the cellular morphology under LPS and APR
treatment were also observed (Fig.
1c). Untreated RAW264.7 cells are circular shaped. However,
following incubation with LPS, the cells exerted an anomalous shape
and became elongated. Microscopic examination of cells revealed a
reversal of LPS-induced alteration in cell morphology following
treatment with APR.
On the other hand, Fig.
1d shows the number of cell surface changes following LPS
and/or APR treatment in RAW264.7 cells. APR significantly decreased
the number of cell surface changes induced by LPS. These results
suggest that APR inhibits the proliferation of RAW264.7 cells and
blocks the LPS-induced activation of RAW264.7 cells.
APR decreases iNOS and COX-2 mRNA
expression in RAW264.7 cells
Since iNOS and ROS are mediators in inflammatory
reactions, the expression of iNOS mRNA and COX-2 mRNA in RAW264.7
cells was assessed. APR suppressed iNOS mRNA and COX-2 mRNA
expression induced by LPS in RAW264.7 cells (Fig. 2a), suggesting that APR suppresses
inflammatory reactions.
 | Figure 2APR decreased iNOS and Cox-2 mRNA
expression and suppressed intracellular ROS levels induced by LPS
in RAW264.7 cells. (A) The expression of iNOS and COX-2 mRNA was
assayed by RT-PCR. Cells were treated with 150 μg/ml APR in the
absence or presence of 1 μg/ml LPS for 12 h. APR suppressed iNOS
and COX-2 mRNA expression induced by LPS. ROS were detected by (B)
laser confocal scanning microscopy and (C) flow cytometry with
DCFH-DA. Cells were treated with 150 μg/ml APR in the absence or
presence of 1 μg/ml LPS for 6 h and incubated with DCFH-DA for 30
min. DCF fluorescence was measured using a confocal laser-scanning
microscopy and FACSCalibur. APR inhibited ROS generation induced by
LPS. APR, Angelica gigas Nakai, Panax ginseng and
Rhus verniciflua Stokes; iNOS, nitric oxide synthase
isoform; Cox-2, prostaglandin endoperoxide synthase 2; ROS,
reactive oxygen species; LPS, liposaccharide RT-PCR, reverse
transcription polymerase chain reaction; DCFH-DA,
2′,7′-dichlorofluorescein diacetate; GADPH, glyceraldehyde
3-phosphate dehydrogenase. |
APR decreases ROS levels in RAW264.7
cells
ROS levels were measured by confocal microscopy
(Fig. 2b) and
fluorescence-activated cell sorting (FACS) analysis (Fig. 2c) using DCFH-DA. Following LPS
treatment cellular ROS levels were increased. However, APR
co-treatment inhibited ROS generation induced by LPS in a
time-dependent manner.
APR affects cell cycle
The effect of APR on the cell cycle was assessed
using FACS analysis. As shown in Fig.
3a and b, APR caused G1 phase arrest at 6 and 12 h of
incubation in a time-dependent manner. Fig. 3c shows the expression of
intracellular molecules associated with cell proliferation,
assessed using western blot analysis.
 | Figure 3APR affects cell cycle. (A) Cell cycle
was analyzed using FACS. Cells were treated with APR (150 μg/ml) in
the absence or presence of 1 μg/ml LPS for 3, 6 and 12 h,
respectively. Cells were stained with propidium iodide solution and
analyzed by flow cytometry. APR caused G1 arrest. (B) G1, S and
G2/M phase fractions were quantified from DNA histograms data in A.
(C) Effect of APR on the expression of intracellular molecules in
RAW264.7 cells. Cells were treated with APR (150 μg/ml) in the
absence or presence of 1 μg/ml LPS for 3, 6 and 12 h, respectively.
Cell extracts were subjected to western blot analysis with specific
antibodies. APR had inhibitory effects on AKT, ERK, p38 and NF-κB
phosphorylation. APR, Angelica gigas Nakai, Panax
ginseng and Rhus verniciflua Stokes; FACS,
fluorescence-activated cell sorting; LPS, liposaccharide; AKT,
protein kinase B; ERK, extracellular signal-regulated kinase;
p-JNK, c-Jun terminal kinase; p38, p38 mitogen-activated protein
kinase; NF-κB, necrosis factor κB. |
APR suppresses protein kinase B (p-AKT)
and extracellular signal-regulated kinase (p-ERK) expression
induced by LPS in a time-dependent manner
c-Jun terminal kinase (JNK) phosphorylation was not
suppressed by APR, but p38 mitogen-activated protein kinases (p38)
phosphorylation induced by LPS was downregulated in a
time-dependent manner by APR. Necrosis factor κB (NF-κB)
phosphorylation induced by LPS was inhibited by APR. These results
suggest that APR inhibits cell proliferation by suppression of the
phosphorylation of AKT, ERK, p38 and NF-κB.
APR induces apoptosis through the
mitochondrial death pathway
The loss of mitochondrial membrane potential (ΔΨ) is
a hallmark of apoptosis. Mitochondrial permeability transition is
an important step in the induction of cell apoptosis. JC-1 is able
to selectively enter mitochondria and reversibly change the color
from red to green as the membrane potential decreases. Thus, cells
incubated with LPS and/or APR were stained with JC-1 and assessed
through FACS analysis. It was found that APR decreased the
mitochondrial membrane potential in RAW264.7 cells as shown in
Fig. 4a. Treatment with LPS alone
resulted in a more stable mitochondrial membrane potential (green
fluorescence, 9.61% at 12 h) compared with the control (green
fluorescence, 31.72% at 12 h). APR perturbed this stability induced
by LPS and decreased the mitochondrial membrane potential (green
fluorescence, 32.92% at 12 h), suggesting that APR induces
apoptosis through the mitochondrial death pathway.
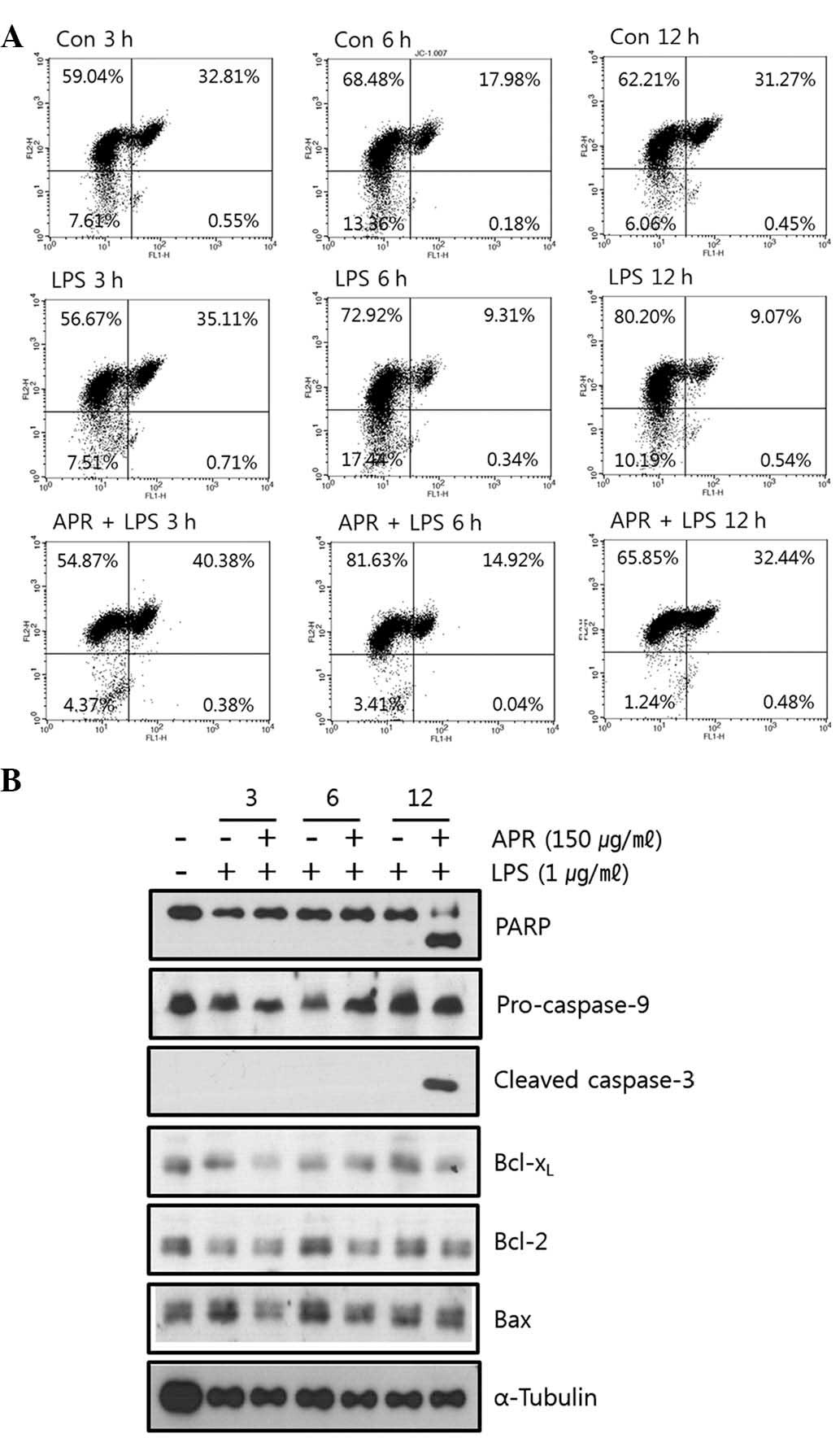 | Figure 4APR induces apoptosis through
mitochondrial death pathway. (a) Cells were treated with APR (150
μg/ml) in the absence or presence of 1 μg/ml LPS for 3, 6 and 12 h,
respectively. Following incubation with JC-1, cells were analyzed
by FACS. APR decreased the mitochondrial membrane potential. (b)
Effect of APR on caspase activity and apoptosis in RAW264.7 cells.
Cells were treated with APR (150 μg/ml) in the absence or presence
of 1 μg/ml LPS for 3, 6 and 12 h, respectively. Cell extracts were
subjected to western blot analysis with specific antibodies. APR
cleaved PARP and caspase-3. APR, Angelica gigas Nakai,
Panax ginseng and Rhus verniciflua Stokes; LPS,
liposaccharide; JC-1,
5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolylcarbocyanine
iodide; FACS, fluorescence-activated cell sorting; PARP, poly
adenosine diphosphate ribose polymerase; Bcl-xL, B-cell
lymphoma-extra large protein; Bcl-2, B-cell lymphoma 2; Bax,
BCl-2-associated protein x. |
It was also confirmed whether APR regulated the
expression of apoptosis-associated molecules. As shown in Fig. 4b, APR induced the cleavage of poly
adenosine diphosphate ribose polymerase (PARP) and caspase-3 which
are apoptotic products, confirming that APR causes apoptosis.
Discussion
The present study revealed that an ethanolic extract
of APR suppressed the LPS-induced inflammatory responses in the
mouse macrophage cell line RAW264.7.
Inflammation is a host’s protection method against
pathogens and is stimulated by diverse microbial products (20). Pro-inflammatory cytokines may
aggravate the severity of multiple inflammatory diseases (21). Diverse inflammatory agents are able
to activate NF-κB. Activation of NF-κB induces inflammation and
increases cell survival and tumor cell transformation (22). Mitogen-activated protein kinase
(MAPK) pathways are also associated with inflammation. The ERK
pathway is activated by inflammation (23).
APR effectively inhibited the growth stimulation and
activation of RAW264.7 cells induced by LPS. APR significantly
inhibited cell growth at concentrations of 250–1,000 μg/ml and
induced cell death at 150 μg/ml (24 h). It was shown that APR
negated the morphological changes of RAW264.7 cells induced by LPS.
APR decreased intracellular ROS levels and suppressed iNOS and
Cox-2 mRNA expression induced by LPS. APR decreased the
mitochondrial membrane potential and cleaved caspase-3 and PARP.
This result indicates that APR inhibits inflammation and induces
apoptosis through mitochondrial death pathways.
Since APR has an anti-inflammatory effect, it may be
used for the treatment of inflammatory diseases, including
rheumatoid arthritis and asthma (24). Transformation of a normal cell into
tumor cell is closely associated with chronic inflammation
(25), and accordingly, APR (AGN,
PG and RVS) may be a useful compound for cancer prevention.
Acknowledgements
The present study was supported by the Traditional
Korean Medicine R&D Project, Ministry of Health & Welfare,
Republic of Korea (B110043).
References
|
1
|
Oh PS, Lee SJ and Lim KT: Glycoprotein
isolated from Rhus verniciflua Stokes inhibits inflammation-related
protein and nitric oxide production in LPS-stimulated RAW 264.7
cells. Biol Pharm Bull. 30:111–116. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Park EK, Shin YW, Lee HU, Kim SS, Lee YC,
Lee BY and Kim DH: Inhibitory effect of ginsenoside Rb1 and
compound K on NO and prostaglandin E2 biosyntheses of RAW264.7
cells induced by lipopolysaccharide. Biol Pharm Bull. 28:652–656.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yang JH, Suh SJ, Lu Y, Li X, Lee YK, Chang
YC, Na MK, Choi JH, Kim CH, Son JK and Chang HW: Anti-inflammatory
activity of ethylacetate fraction of Cliona celata. Immunopharmacol
Immunotoxicol. 33:373–379. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hofseth LJ and Wargovich MJ: Inflammation,
cancer, and targets of ginseng. J Nutr. 137(1 Suppl): 183S–185S.
2007.PubMed/NCBI
|
|
5
|
Liu Q, Kou JP and Yu BY: Ginsenoside Rg1
protects against hydrogen peroxide-induced cell death in PC12 cells
via inhibiting NF-κB activation. Neurochem Int. 58:119–125.
2011.PubMed/NCBI
|
|
6
|
Shin S, Jeon JH, Park D, Jang JY, Joo SS,
Hwang BY, Choe SY and Kim YB: Anti-inflammatory effects of an
ethanol extract of Angelica gigas in a Carrageenan-air pouch
inflammation model. Exp Anim. 58:431–436. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kim JY, Yoon YD, Ahn JM, Kang JS, Park SK,
Lee K, Song KB, Kim HM and Han SB: Angelan isolated from Angelica
gigas Nakai induces dendritic cell maturation through toll-like
receptor 4. Int Immunopharmacol. 7:78–87. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kim HM, Kang JS, Park SK, Lee K, Kim JY,
Kim YJ, Hong JT, Kim Y and Han SB: Antidiabetic activity of angelan
isolated from Angelica gigas Nakai. Arch Pharm Res. 31:1489–1496.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kim WJ, Lee MY, Kim JH, Suk K and Lee WH:
Decursinol angelate blocks transmigration and inflammatory
activation of cancer cells through inhibition of PI3K, ERK and
NF-κB activation. Cancer Lett. 296:35–42. 2010.PubMed/NCBI
|
|
10
|
Shin S, Joo SS, Park D, Jeon JH, Kim TK,
Kim JS, Park SK, Hwang BY and Kim YB: Ethanol extract of
Angelica gigas inhibits croton oil-induced inflammation by
suppressing the cyclooxygenase - prostaglandin pathway. J Vet Sci.
11:43–50. 2010.
|
|
11
|
Su W, Sun AJ, Xu DL, Zhang HQ, Yang L,
Yuan LY, Jia JG, Zou YZ, Wu YL, Wang KQ and Ge JB: Inhibiting
effects of total saponins of panax ginseng on immune maturation of
dendritic cells induced by oxidized-low density lipoprotein. Cell
Immunol. 263:99–104. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Park HJ, Han ES, Park DK, Lee C and Lee
KW: An extract of Phellinus linteus grown on germinated brown rice
inhibits inflammation markers in RAW264.7 macrophages by
suppressing inflammatory cytokines, chemokines, and mediators and
up-regulating antioxidant activity. J Med Food. 13:1468–1477. 2010.
View Article : Google Scholar
|
|
13
|
Song Z, Kong KF, Wu H, Maricic N,
Ramalingam B, Priestap H, Schneper L, Quirke JM, Høiby N and Mathee
K: Panax ginseng has anti-infective activity against opportunistic
pathogen Pseudomonas aeruginosa by inhibiting quorum
sensing, a bacterial communication process critical for
establishing infection. Phytomedicine. 17:1040–1046. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kiefer D and Pantuso T: Panax ginseng. Am
Fam Physician. 68:1539–1542. 2003.PubMed/NCBI
|
|
15
|
Ahn JY, Choi IS, Shim JY, Yun EK, Yun YS,
Jeong G and Song JY: The immunomodulator ginsan induces resistance
to experimental sepsis by inhibiting Toll-like receptor-mediated
inflammatory signals. Eur J Immunol. 36:37–45. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cho SH, Chung KS, Choi JH, Kim DH and Lee
KT: Compound K, a metabolite of ginseng saponin, induces apoptosis
via caspase-8-dependent pathway in HL-60 human leukemia cells. BMC
Cancer. 9:4492009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jung CH, Jun CY, Lee S, Park CH, Cho K and
Ko SG: Rhus verniciflua stokes extract: radical scavenging
activities and protective effects on
H2O2-induced cytotoxicity in macrophage RAW
264.7 cell lines. Biol Pharm Bull. 29:1603–1607. 2006. View Article : Google Scholar
|
|
18
|
Jung CH, Kim JH, Hong MH, Seog HM, Oh SH,
Lee PJ, Kim GJ, Kim HM, Um JY and Ko SG: Phenolic-rich fraction
from Rhus verniciflua Stokes (RVS) suppress inflammatory
response via NF-κB and JNK pathway in lipopolysaccharide-induced
RAW 264.7 macrophages. J Ethnopharmacol. 110:490–497. 2007.
|
|
19
|
Hong MH, Kim JH, Lee SY, Go HY, Shin YC,
Kim SH and Ko SG: Early antiallergic inflammatory effects of
Rhus verniciflua Stokes on human mast cells. Phytother Res.
24:288–294. 2010.PubMed/NCBI
|
|
20
|
Lee HJ, Maeng K, Dang HT, Kang GJ, Ryou C,
Jung JH, Kang HK, Prchal JT, Yoo ES and Yoon D: Anti-inflammatory
effect of methyl dehydrojasmonate (J2) is mediated by the NF-κB
pathway. J Mol Med (Berl). 89:83–90. 2011.PubMed/NCBI
|
|
21
|
Tang S, Shen XY, Huang HQ, Xu SW, Yu Y,
Zhou CH, Chen SR, Le K, Wang YH and Liu PQ: Cryptotanshinone
suppressed inflammatory cytokines secretion in RAW264.7 macrophages
through inhibition of the NF-κB and MAPK signaling pathways.
Inflammation. 34:111–118. 2011.PubMed/NCBI
|
|
22
|
Reuter S, Prasad S, Phromnoi K, Ravindran
J, Sung B, Yadav VR, Kannappan R, Chaturvedi MM and Aggarwal BB:
Thiocolchicoside exhibits anticancer effects through downregulation
of NF-κB pathway and its regulated gene products linked to
inflammation and cancer. Cancer Prev Res (Phila). 3:1462–1472.
2010.PubMed/NCBI
|
|
23
|
Fan J, Liu K, Zhang Z, Luo T, Xi Z, Song J
and Liu B: Modified Si-Miao-San extract inhibits the release of
inflammatory mediators from lipopolysaccharide-stimulated mouse
macrophages. J Ethnopharmacol. 129:5–9. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jeong JB and Jeong HJ: Rheosmin, a
naturally occurring phenolic compound inhibits LPS-induced iNOS and
COX-2 expression in RAW264.7 cells by blocking NF-κB activation
pathway. Food Chem Toxicol. 48:2148–2153. 2010.PubMed/NCBI
|
|
25
|
Reuter S, Gupta SC, Chaturvedi MM and
Aggarwal BB: Oxidative stress, inflammation, and cancer: how are
they linked? Free Radic Biol Med. 49:1603–1616. 2010. View Article : Google Scholar : PubMed/NCBI
|















