Introduction
Prostate cancer is one of the leading causes of
cancer-associated mortality in males, accounting for >240,000
new cancer cases and 28,000 fatalities annually in males in the
United States (1,2). Although effective surgical and radiation
treatments exist for clinically localized prostate cancer, the
majority of patients with metastatic prostate cancer eventually
succumb to the disease (3).
Therefore, in order to develop more effective outcomes for the
diagnosis and treatment of prostate cancer, articulation of the
genetic underpinning and novel therapeutic targets are critically
required.
Recently, the presence and function of muscarinic
acetylcholine receptors (mAChRs) in the human prostate have aroused
wide concern (4,5). mAChR and its ligands have been found to
play key roles in regulating cellular proliferation and cancer
progression (6). mAChRs are
preferentially localized to the glandular epithelium of the
prostate and promote the paracrine/autocrine actions within the
prostate gland, which are critical for cancer cell survival,
proliferation and migration (4,6,7). mAChRs consist of five distinct subtypes
(M1-M5), and the M3 mAChR (also known as CHRM3) has been found to
be a key member involved in prostate cancer. The stimulation of
CHRM3 is strongly associated with the tumor growth of prostate
carcinomas (4). Additionally, CHRM3
can effectively mediate the contractions of the mouse prostate
elicited by acetylcholine (8). A
great deal of attention has been focused on the roles of CHRM3 in
prostate cancer, however, the signal transduction by CHRM3 in the
pathophysiology of prostate cancer is not well understood.
Therefore, investigation of the signal transduction by CHRM3 will
herald a prominent expansion in our understanding of the molecular
mechanisms of prostate cancer.
The microarray data of GSE3325 has been used to
reveal critical genomic regions in the prostate tumor
microenvironment for investigating novel biomarkers (9) or to reveal signatures of the metastatic
progression of prostate cancer (10).
In contrast to previous findings, the present study downloaded the
GSE3325 microarray data and utilized comprehensive bioinformatics
methods to identify the differentially-expressed genes (DEGs)
associated with prostate cancer. Additionally, functional
enrichment analysis of the DEGs was performed, and a
protein-protein interaction (PPI) network and microRNA
(miRNA)-target DEG network was constructed. The study aimed to
elucidate the potential mechanisms used during signal transduction
by CHRM3 in prostate cancer.
Materials and methods
Affymetrix microarray data
The microarray data of GSE3325 were downloaded from
the Gene Expression Omnibus database (http://www.ncbi.nlm.nih.gov/geo/), based on the
platform of GPL570 (Affymetrix Human Genome U133 Plus 2.0 Array;
Affymetrix Inc., Santa Clara, CA, USA). The gene expression files
were deposited by Varambally et al (10). A total of 19 samples were applied to
the development of the array data, including 6 benign, 7 clinically
localized primary and 6 metastatic prostate cancer tissues. Each
category contained 2 pooled samples. In order to better investigate
the molecular pathogenesis of prostate cancer, pooled samples were
discarded, and the expression profiles analyzed in this study were
derived from 9 samples, including 5 clinically localized primary
prostate cancer and 4 benign prostate tissues.
Data preprocessing and DEG
screening
The raw data were first preprocessed by the robust
multiarray average algorithm (11)
with application of Affy package (12) in R language. The gene expression
matrix of the samples was then acquired.
The DEGs in the primary prostate cancer tissues
compared with the benign controls were screened by Limma package
(13,14) in R language. The P-value was adjusted
using Student's t-test (13) in
Limma. Fold-change (FC) of the gene expression was also observed
for differential-expression test. The DEGs with adjusted P-values
<0.05 and |log2 FC| >1 were considered to be
significant.
Gene Ontology (GO) and pathway
enrichment analysis
The GO database (15)
is a large-scale collection of gene annotation terms. The Kyoto
Encyclopedia of Genes and Genomes (KEGG) (16) is a pathway-related database for
classification of correlating gene sets into their respective
pathways. The Database for Annotation Visualization and Integrated
Discovery (DAVID) (17) is an online
tool used for systematically associating the functional terms with
large gene or protein lists.
In order to analyze the function of DEGs, GO
annotation associated with biological process (BP) and KEGG pathway
enrichment analysis of DEGs by DAVID online tool were performed.
P<0.05 and gene counts >2 were set as the threshold.
PPI network construction
The Search Tool for the Retrieval of Interacting
Genes (STRING) (18) is an online
database that provides comprehensive information on predicted and
experimental interactions of proteins in a given cell. The
interactions of protein pairs are displayed with a combined score.
In the present study, the DEGs were mapped into the STRING database
to construct a PPI network with a combined score of protein pairs
of >0.4 as the cut-off value. In addition, the connectivity
degree of each node in the PPI network was calculated and the hub
node was then identified (19).
Prediction of miRNAs associated with
DEGs
The Web-based Gene Set Analysis Toolkit (WebGestalt)
(20) is a web-based integrated
system for analyzing large-scale gene sets in various functional
categories, such as transcription factor and miRNA targets. In
order to screen the potential miRNAs associated with DEGs, the DEG
lists were uploaded to the WebGestalt system, and a miRNA-target
DEG analysis was performed by WebGestalt. Enriched gene counts of
≥2 and rawP<0.05 were defined as the cut-off value.
Then miRNA-target DEG network composed of miRNAs and
their target DEGs was visualized in the application of Cytoscape
(21) software.
Integration of PPI network and
miRNA-target DEG network
In the present study, the miRNA-target DEGs and PPI
networks were further integrated, and the PPI pair-miRNA network
composed of miRNAs and their target protein pairs was established
with Cytoscape (21) software.
Results
DEG screening
Using the cut-off value of adjusted P-values
<0.05 and |log2 FC| >1, a total of 224 DEGs in the
primary prostate cancer tissues compared with the benign controls
were screened, including 113 upregulated and 111 downregulated
genes.
GO and pathway enrichment
analysis
GO and pathway analyses were performed for the
upregulated and downregulated genes, respectively. The
overrepresented GO-BP terms of the upregulated DEGs were mainly
associated with amine transport, amino acid transport and the
enzyme-linked receptor protein signaling pathway (Table I). The downregulated DEGs were mainly
involved in the regulation of cell proliferation, the response to
organic substances and the negative regulation of the nitrogen
compound metabolic process (Table
II).
 | Table I.GO-BP terms and KEGG pathways
enriched by upregulated genes. |
Table I.
GO-BP terms and KEGG pathways
enriched by upregulated genes.
| A, GO-BP terms |
|
|
|
|---|
|
|---|
| Term | Description | Count | P-value |
|---|
| GO:0015837 | Amine
transport | 5 |
5.95×10−3 |
| GO:0006865 | Amino acid
transport | 4 |
1.78×10−2 |
| GO:0007167 | Enzyme-linked
receptor protein signaling pathway | 7 |
1.85×10−2 |
| GO:0051674 | Localization of
cell | 6 |
4.03×10−2 |
| GO:0048870 | Cell motility | 6 |
4.03×10−2 |
| GO:0035239 | Tube
morphogenesis | 4 |
4.32×10−2 |
| GO:0001501 | Skeletal system
development | 6 |
4.62×10−2 |
| GO:0035295 | Tube
development | 5 |
4.63×10−2 |
|
| B, Enriched KEGG
pathways |
|
|
|
|
| Term | Description | Count | P-value |
|
| hsa04012 | ErbB signaling
pathway | 3 |
8.21×10−2 |
| hsa05216 | Thyroid cancer | 2 |
1.48×10−1 |
| hsa00512 | O-glycan
biosynthesis | 2 |
1.53×10−1 |
| hsa04514 | CAMs | 3 |
1.64×10−1 |
| hsa00071 | Fatty acid
metabolism | 2 |
1.99×10−1 |
| hsa05219 | Bladder cancer | 2 |
2.08×10−1 |
| hsa05213 | Endometrial
cancer | 2 |
2.51×10−1 |
| hsa05221 | Acute myeloid
leukemia | 2 |
2.75×10−1 |
| hsa03320 | PPAR signaling
pathway | 2 |
3.19×10−1 |
| hsa04810 | Regulation of actin
cytoskeleton | 3 |
3.33×10−1 |
| hsa04350 | TGF-β signaling
pathway | 2 |
3.84×10−1 |
| hsa04060 | Cytokine-cytokine
receptor interaction | 3 |
4.27×10−1 |
| hsa05200 | Pathways in
cancer | 3 |
5.48×10−1 |
| hsa04310 | Wnt signaling
pathway | 2 |
5.71×10−1 |
| hsa04144 | Endocytosis | 2 |
6.45×10−1 |
| hsa04510 | Focal adhesion | 2 |
6.78×10−1 |
| hsa04080 | Neuroactive
ligand-receptor interaction | 2 |
7.66×10−1 |
| hsa04010 | MAPK signaling
pathway | 2 |
7.80×10−1 |
| hsa04740 | Olfactory
transduction | 2 |
8.86×10−1 |
| Table II.Top 10 GO-BP terms and KEGG pathways
enriched by downregulated genes. |
Table II.
Top 10 GO-BP terms and KEGG pathways
enriched by downregulated genes.
| A, GO-BP terms |
|
|
|
|---|
|
|---|
| Term | Description | Count | P-value |
|---|
| GO:0042127 | Regulation of cell
proliferation | 11 |
9.36×10−3 |
| GO:0010033 | Response to organic
substance | 9 |
3.99×10−2 |
| GO:0051172 | Negative regulation
of nitrogen compound metabolic process | 8 |
2.16×10−2 |
| GO:0031327 | Negative regulation
of cellular biosynthetic process | 8 |
3.13×10−2 |
| GO:0009890 | Negative regulation
of biosynthetic process | 8 |
3.45×10−2 |
| GO:0044092 | Negative regulation
of molecular function | 7 |
9.21×10−3 |
| GO:0008015 | Blood
circulation | 6 |
3.29×10−3 |
| GO:0003013 | Circulatory system
process | 6 |
3.29×10−3 |
| GO:0000122 | Negative regulation
of transcription from RNA polymerase II promoter | 6 |
1.44×10−2 |
| GO:0045892 | Negative regulation
of transcription, DNA-dependent | 6 |
4.31×10−2 |
|
| B, The enriched
KEGG pathways |
|
|
|
|
| Term | Description | Count | P-value |
|
| hsa00982 | Drug
metabolism | 3 |
3.14×10−2 |
| hsa04360 | Axon guidance | 3 |
1.14×10−1 |
| hsa00980 | Metabolism of
xenobiotics by cytochrome P450 | 2 |
2.39×10−1 |
| hsa04512 | ECM-receptor
interaction | 2 |
3.19×10−1 |
| hsa04350 | TGF-β signaling
pathway | 2 |
3.28×10−1 |
| hsa04916 | Melanogenesis | 2 |
3.64×10−1 |
| hsa04630 | Jak-STAT signaling
pathway | 2 |
5.10×10−1 |
| hsa04020 | Calcium signaling
pathway | 2 |
5.56×10−1 |
| hsa04510 | Focal adhesion | 2 |
6.05×10−1 |
| hsa05200 | Pathways in
cancer | 2 |
7.85×10−1 |
The upregulated DEGs were significantly enriched in
the ErbB signaling pathway, thyroid cancer and O-glycan
biosynthesis (Table I), while
downregulated DEGs were significantly enriched in drug metabolism,
axon guidance and metabolism of xenobiotics by cytochrome P450
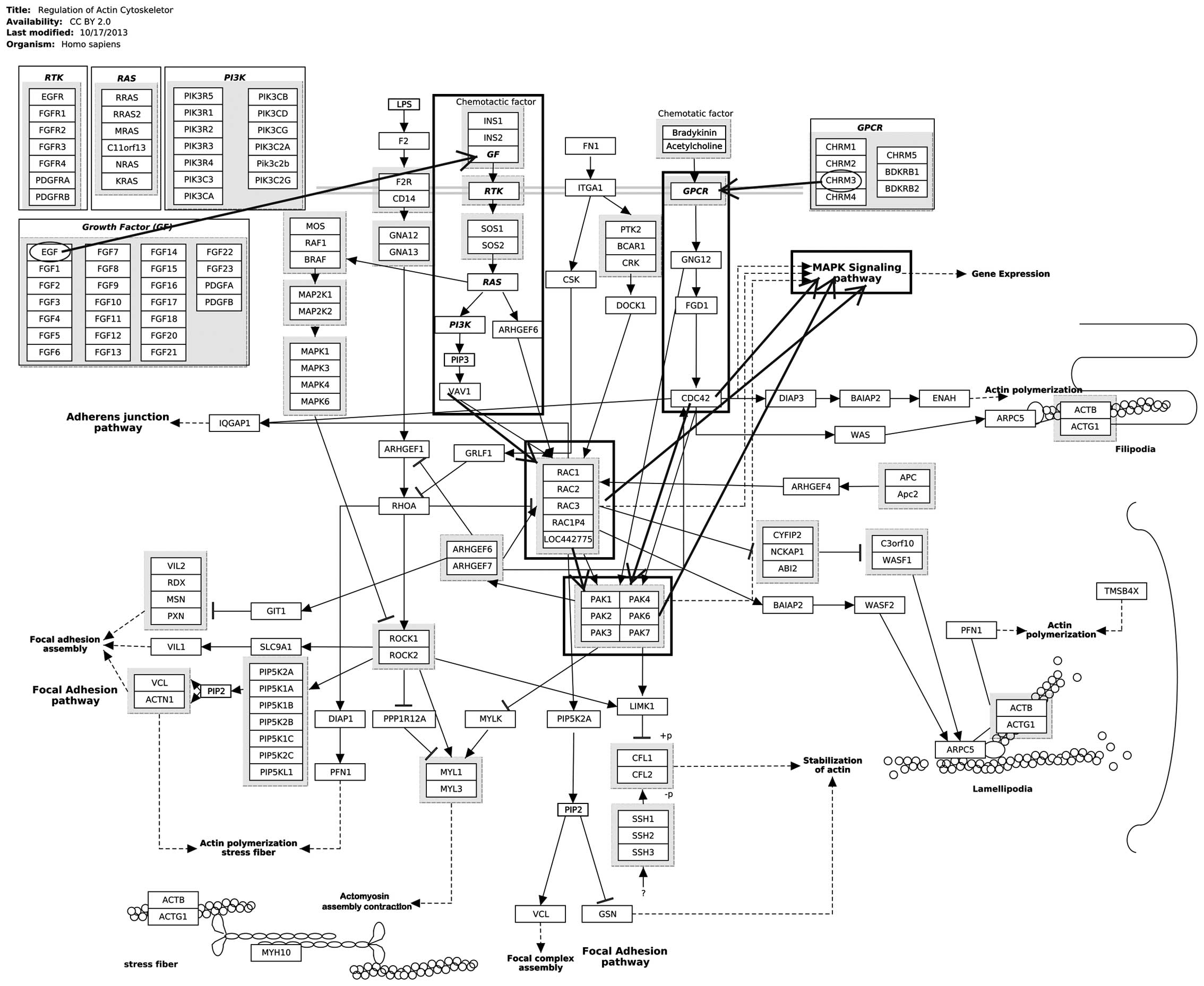
(Table II). Notably, CHRM3 and
epidermal growth factor (EGF) were significantly enriched in
regulation of the actin cytoskeleton (Fig. 1). EGF and v-myc avian myelocytomatosis
viral oncogene homolog (Myc) were enriched in the mitogen-activated
protein kinase (MAPK) signaling pathway.
PPI network analysis
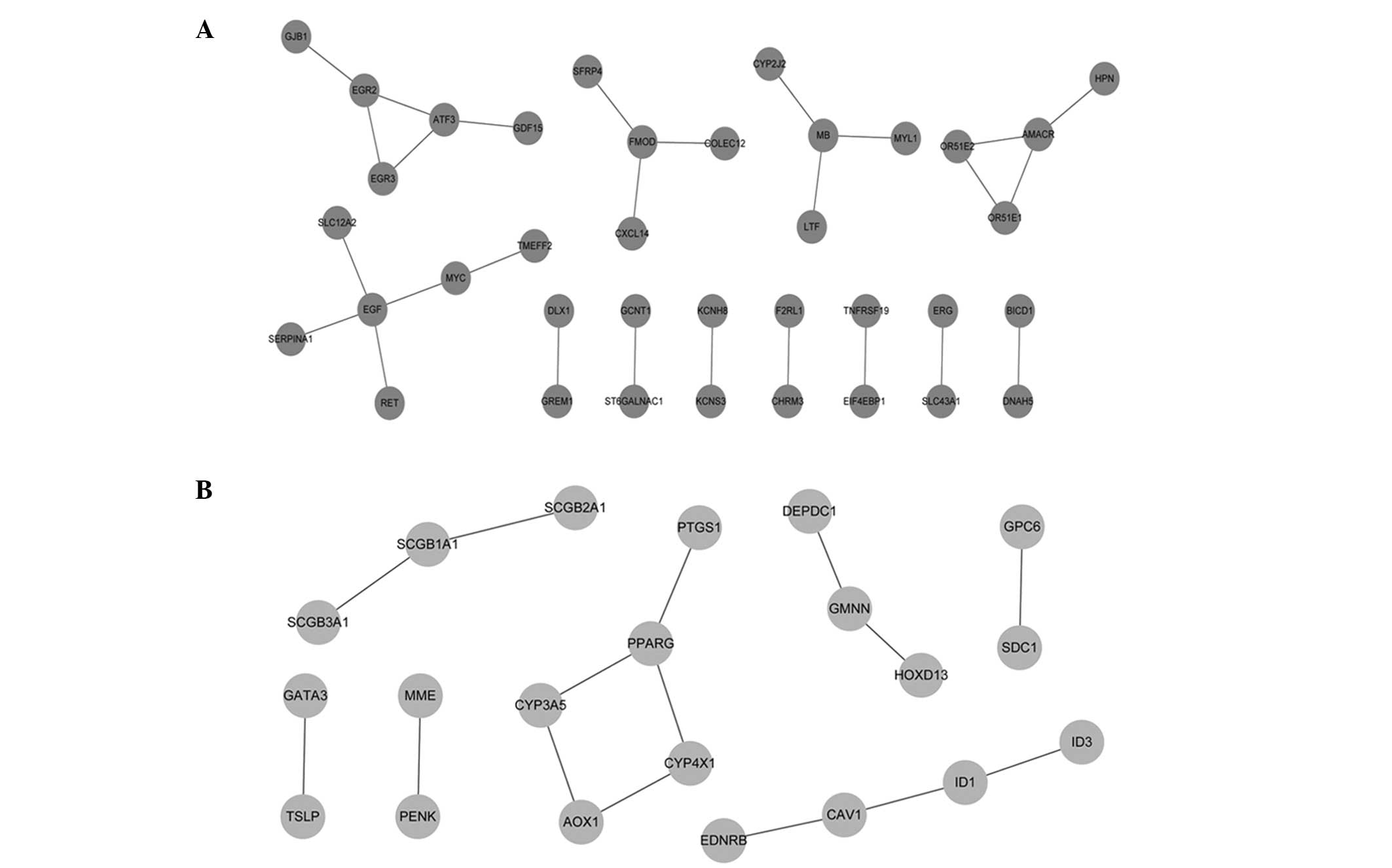
Based on the STRING database, a total of 27
upregulated PPIs (Fig. 2A) and 15
downregulated PPIs (Fig. 2B) with a
combined score of >0.4 were obtained. The upregulated nodes with
a connectivity degree of >2 were EGF (4), myoglobin (MB) (3), α-methylacyl-CoA racemase (3), early growth response 2 (3), fibromodulin (3) and activating transcription factor 3
(3). The downregulated node with a
connectivity degree of >2 was peroxisome proliferator-activated
receptor γ.
Prediction of miRNAs associated with
DEGs
The miRNAs associated with DEGs were predicted by
WebGestalt. A total of 11 upregulated miRNAs, including miR-23A,
miR-23B, miR-219 and miR-143, and 4 downregulated miRNAs, including
miR-193a, miR-193b, miR-124a and miR-302c, were selected (Table III). Notably, miR-34b could interact
with Myc directly.
| Table III.Predicted miRNAs associated with
DEGs. |
Table III.
Predicted miRNAs associated with
DEGs.
| A, Upregulated |
|
|
|---|
|
|---|
| miRNA | Count | P-value |
|---|
| hsa_GACAATC,
miR-219 | 5 |
1.10×10−3 |
| hsa_TCATCTC,
miR-143 | 5 |
1.20×10−3 |
| hsa_AATGTGA,
miR-23a, miR-23b | 8 |
2.00×10−3 |
| hsa_ACTGCCT,
miR-34b | 5 |
6.20×10−3 |
| hsa_AAGCAAT,
miR-137 | 5 |
6.30×10−3 |
| hsa_GAGCCAG,
miR-149 | 4 |
7.60×10−3 |
| hsa_TCTGATA,
miR-361 | 3 |
1.34×10−2 |
| hsa_CATGTAA,
miR-496 | 4 |
1.41×10−2 |
| hsa_GCAAGAC,
miR-431 | 2 |
2.33×10−2 |
| hsa_TACTTGA,
miR-26a, miR-26b | 5 |
2.33×10−2 |
| hsa_CAGTGTT,
miR-141, miR-200a | 5 |
2.61×10−2 |
|
| B,
Downregulated |
|
|
|
| miRNA | Count | P-value |
|
| hsa_GGCCAGT,
miR-193a, miR-193b | 3 |
1.11×10−2 |
| hsa_TGCCTTA,
miR-124a | 7 |
2.22×10−2 |
| hsa_ATGTTAA,
miR-302c | 4 |
3.31×10−2 |
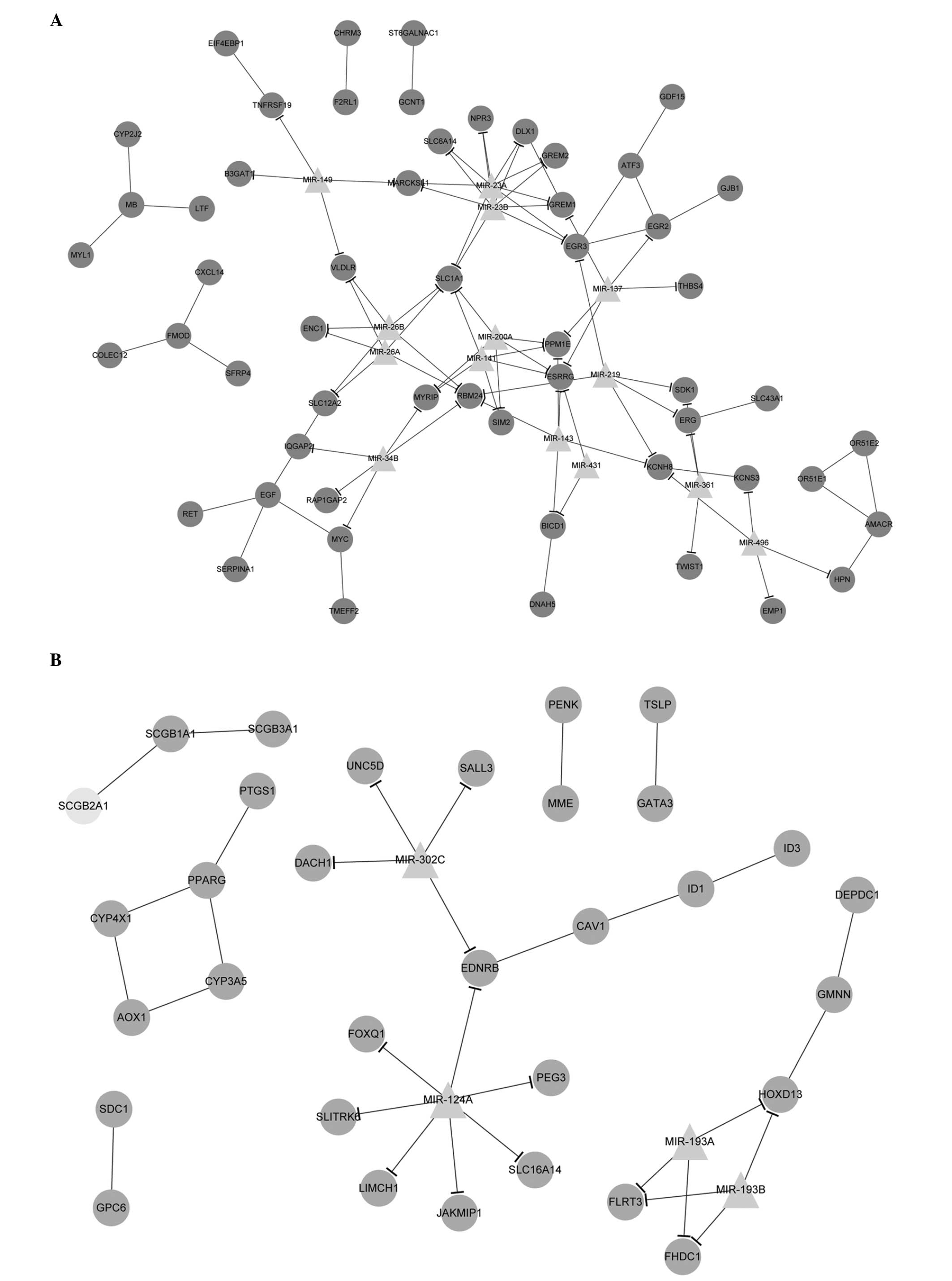
Integration of PPI and miRNA-target
DEG networks
With Cytoscape software, the PPI and miRNA-target
DEG networks were successfully integrated together. The upregulated
and downregulated PPI pair-miRNA networks with significant protein
pairs and their corresponding miRNAs were also established
(Fig. 3).
Discussion
In the present study, the bioinformatics approach
was used to investigate the potential mechanisms used during signal
transduction by CHRM3 in prostate cancer. The results showed that
CHRM3 and EGF were significantly enriched in regulation of actin
cytoskeleton. EGF and Myc were enriched in the MAPK signaling
pathway. Notably, as shown in Fig. 1,
the activation of the regulation of the actin cytoskeleton may
induce the activation of MAPK signaling pathway to a certain
extent, thus promoting the progression of prostate cancer.
Therefore, these DEGs and the aforementioned two pathways may be
potential mechanisms during signal transduction by CHRM3 in
prostate progression.
EGF, with the highest degree of connectivity, was
the hub protein in the PPI network. Growth factors (GFs), including
EGF, and their transmembrane receptor tyrosine kinases (RTKs) play
important roles in the growth, proliferation, migration and
differentiation of various human tumor cells (22,23). The
EGF receptor (EGFR) family of RTKs is formed by four members:
EGFR/ErbB1, human epidermal growth factor (HER)2/ErbB2, HER3/ErbB3
and HER4/ErbB4 (24). Increased
expression and/or amplification of EGFR and HER2 have been recorded
in a range of human cancer types (24). Additionally, stimulation with EGF
results in the activation of the lipoprotein kinase,
phosphatidylinositol 3-kinase (PI3K), which phosphorylates
phosphatidylinositol 4,5-bisphosphate, generating the second
messenger phosphatidylinositol (3,4,5)-trisphosphate (25). PI3K-Akt signaling pathway activation
promotes prostate cancer cell metastasis and invasion (26,27). In
the present study, the PI3K-Akt signaling pathway activated by EGF
was a part of the regulation of the actin cytoskeleton. Therefore,
the present results are in line with previous findings and suggest
that EGF may play roles in the metastasis and invasion of prostate
cancer via regulation of the actin cytoskeleton.
Furthermore, EGFR and HER2 have been identified to
be essential pathway elements in the signaling from G
protein-coupled receptors (GPCRs) (24). CHRM3 belongs to the GPCRs and is
coupled to MAPK via EGFR (28). GPCRs
may interact with Rho guanosine triphosphatases, including RhoA and
Cdc42, which play roles in regulating cell motility (29). Moreover, GPCRs can induce EGFR
transactivation, thus generating signals defining the required
biological response (24). The roles
of EGFR are as aforementioned. In addition, previous findings have
strongly linked the excessive activation of the GPCR and RTK
pathways to prostate cancer metastasis (26). Therefore, as shown in Fig. 1, CHRM3 may activate the regulation of
the actin cytoskeleton via EGFR or GPCRs for the promotion of the
metastasis of prostate cancer.
Notably, the highly-conserved MAPK signaling pathway
can be activated by EGFR (22,24,30).
Previous studies have also suggested that the activation of the
MAPK cascade can be mediated by GPCRs via several distinct pathways
(31). Moreover, the MAPK signaling
pathway may be activated by means of a PI3K-dependent feedback loop
in human cancer (32). The
PI3K/Akt/mTOR and MAPK signaling pathways are often observed in
prostate tumors (33). Therefore,
CHRM3 may be further involved in the MAPK signaling pathway via the
roles of EGFR or the activation of the PI3K-Akt signaling pathway.
Additionally, MAPK signaling pathway activation is necessary for
inhibitor of DNA binding 1-induced serum-independent prostate
cancer cell growth (34). Targeting
MAPK signaling pathway activated by AKT/mTOR and MEK/ERK can
inhibit the progression of prostate cancer in humans (35). Taken together, these results and the
present study results suggest that CHRM3 may play important roles
in prostate cancer progression via activating the MAPK signaling
pathway.
In addition to EGF, Myc was found to be
significantly enriched in the MAPK signaling pathway in the present
study. Myc is a basic helix-loop-helix leucine zipper
transcriptional factor that functions in cell proliferation,
differentiation and death (36). An
increased Myc gene copy number is found in human prostate cancer,
and Pim-1 kinase can cooperate with Myc in tumorigenesis (37). c-Myc may link v-ets avian
erythroblastosis virus E26 oncogene homolog to a major oncogenic
pathway in prostate cancer (38). In
addition, Myc could interact with miR-34b directly in the present
study. miR-34 mediates androgen receptor-dependent p53-induced
apoptosis in prostate cancer (39).
The study by Corney et al confirmed that miR-34b is a target
of p53 and plays important roles in the control of cell
proliferation (40). Furthermore, the
MAPK/heterogeneous nuclear ribonucleoprotein K pathway controls the
oncogenic potential of breakpoint cluster region/Abelson murine
leukemia viral oncogene homolog 1 (BCR/ABL) oncoprotein via the
regulation of Myc mRNA translation (41). Therefore, Myc may play an important
role in prostate cancer via the MAPK signaling pathway, and the
present results are consistent with the findings that the MAPK
signaling pathway may be the key mechanism used during signal
transduction by CHRM3 in prostate cancer.
In conclusion, EGF and Myc may play significant
roles in the progression of prostate cancer via regulation of the
actin cytoskeleton and the MAPK signaling pathway. CHRM3 may
activate these two pathways via EGFR or GPCRs in prostate cancer
progression. Thus, regulation of the actin cytoskeleton, the MAPK
signaling pathway and the two key factors of EGF and Myc may be
crucial mechanisms during signal transduction by CHRM3 in prostate
cancer. The present findings shed new light on the molecular
mechanism of prostate cancer and have implications for future
research. However, a relatively small sample size and no
experimental validation are limitations to the present study, and
further genetic studies are required to confirm these
observations.
Acknowledgements
The present study was supported by the Doctoral Fund
of the Ministry of Education of China (grant no.
20120131120070).
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Varambally S, Dhanasekaran SM, Zhou M,
Barrette TR, Kumar-Sinha C, Sanda MG, Ghosh D, Pienta KJ, Sewalt
RG, Otte AP, et al: The polycomb group protein EZH2 is involved in
progression of prostate cancer. Nature. 419:624–629. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rayford W, Noble MJ, Austenfeld MA, Weigel
J, Mebust WK and Shah GV: Muscarinic cholinergic receptors promote
growth of human prostate cancer cells. Prostate. 30:160–166. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Witte LP, Chapple CR, de la Rosette JJ and
Michel MC: Cholinergic innervation and muscarinic receptors in the
human prostate. Eur Urol. 54:326–334. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shah N, Khurana S, Cheng K and Raufman
J-P: Muscarinic receptors and ligands in cancer. Am J Physiol Cell
Physiol. 296:C221–C232. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ventura S, Pennefather J and Mitchelson F:
Cholinergic innervation and function in the prostate gland.
Pharmacol Ther. 94:93–112. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
White CW, Short JL, Haynes JM, Matsui M
and Ventura S: Contractions of the mouse prostate elicited by
acetylcholine are mediated by M(3) muscarinic receptors. J
Pharmacol Exp Ther. 339:870–877. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ashida S, Orloff MS, Bebek G, Zhang L,
Zheng P, Peehl DM and Eng C: Integrated analysis reveals critical
genomic regions in prostate tumor microenvironment associated with
clinicopathologic phenotypes. Clin Cancer Res. 18:1578–1587. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Varambally S, Yu J, Laxman B, Rhodes DR,
Mehra R, Tomlins SA, Shah RB, Chandran U, Monzon FA, Becich MJ, et
al: Integrative genomic and proteomic analysis of prostate cancer
reveals signatures of metastatic progression. Cancer Cell.
8:393–406. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Irizarry RA, Hobbs B, Collin F,
Beazer-Barclay YD, Antonellis KJ, Scherf U and Speed TP:
Exploration, normalization, and summaries of high density
oligonucleotide array probe level data. Biostatistics. 4:249–264.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gautier L, Cope L, Bolstad BM and Irizarry
RA: Affy - analysis of Affymetrix GeneChip data at the probe level.
Bioinformatics. 20:307–315. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Smyth GK: Linear models and empirical
bayes methods for assessing differential expression in microarray
experiments. Stat Appl Genet Mol Biol. 3:Article 3. 2004.PubMed/NCBI
|
|
14
|
Smyth GK: Limma: linear models for
microarray data. Bioinformatics and Computational Biology Solutions
using R and Bioconductor. Gentleman R, Carey V, Dudoit S, Irizarry
R and Huber W: (New York, NY). Springer. 397–420. 2005. View Article : Google Scholar
|
|
15
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: The Gene Ontology Consortium: Gene ontology: Tool for the
unification of biology. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kanehisa M and Goto S: KEGG: Kyoto
encyclopedia of genes and genomes. Nucleic Acids Res. 28:27–30.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huang DW, Sherman BT, Tan Q, Collins JR,
Alvord WG, Roayaei J, Stephens R, Baseler MW, Lane HC and Lempicki
RA: The DAVID Gene Functional Classification Tool: A novel
biological module-centric algorithm to functionally analyze large
gene lists. Genome Biol. 8:R1832007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
von Mering C, Huynen M, Jaeggi D, Schmidt
S, Bork P and Snel B: STRING: A database of predicted functional
associations between proteins. Nucleic Acids Res. 31:258–261. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
He X and Zhang J: Why do hubs tend to be
essential in protein networks? PLoS Genet. 2:e882006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang B, Kirov S and Snoddy J: WebGestalt:
An integrated system for exploring gene sets in various biological
contexts. Nucleic Acids Res. 33(Web Server): W741–8. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kohl M, Wiese S and Warscheid B:
Cytoscape: Software for visualization and analysis of biological
networks. Methods Mol Biol. 696:291–303. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yarden Y: The EGFR family and its ligands
in human cancer. signalling mechanisms and therapeutic
opportunities. Eur J Cancer. 37(Suppl 4): S3–S8. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gschwind A, Fischer OM and Ullrich A: The
discovery of receptor tyrosine kinases: Targets for cancer therapy.
Nat Rev Cancer. 4:361–370. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Prenzel N, Fischer OM, Streit S, Hart S
and Ullrich A: The epidermal growth factor receptor family as a
central element for cellular signal transduction and
diversification. Endocr Relat Cancer. 8:11–31. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bjorge JD, Chan T-O, Antczak M, Kung H-J
and Fujita DJ: Activated type I phosphatidylinositol kinase is
associated with the epidermal growth factor (EGF) receptor
following EGF stimulation. Proc Natl Acad Sci USA. 87:3816–3820.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Qin J, Xie Y, Wang B, Hoshino M, Wolff DW,
Zhao J, Scofield MA, Dowd FJ, Lin MF and Tu Y: Upregulation of
PIP3-dependent Rac exchanger 1 (P-Rex1) promotes prostate cancer
metastasis. Oncogene. 28:1853–1863. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shukla S, Maclennan GT, Hartman DJ, Fu P,
Resnick MI and Gupta S: Activation of PI3K-Akt signaling pathway
promotes prostate cancer cell invasion. Int J Cancer.
121:1424–1432. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Slack BE: The m3 muscarinic acetylcholine
receptor is coupled to mitogen-activated protein kinase via protein
kinase C and epidermal growth factor receptor kinase. Biochem J.
348:381–387. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nie D, Guo Y, Yang D, Tang Y, Chen Y, Wang
MT, Zacharek A, Qiao Y, Che M and Honn KV: Thromboxane A2 receptors
in prostate carcinoma: Expression and its role in regulating cell
motility via small GTPase Rho. Cancer Res. 68:115–121. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Katz M, Amit I and Yarden Y: Regulation of
MAPKs by growth factors and receptor tyrosine kinases. Biochim
Biophys Acta. 1773:1161–1176. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Naor Z, Benard O and Seger R: Activation
of MAPK cascades by G-protein-coupled receptors: The case of
gonadotropin-releasing hormone receptor. Trends Endocrinol Metab.
11:91–99. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Carracedo A, Ma L, Teruya-Feldstein J,
Rojo F, Salmena L, Alimonti A, Egia A, Sasaki AT, Thomas G, Kozma
SC, et al: Inhibition of mTORC1 leads to MAPK pathway activation
through a PI3K-dependent feedback loop in human cancer. J Clin
Invest. 118:3065–3074. 2008.PubMed/NCBI
|
|
33
|
Wang J, Kobayashi T, Floc'h N, Kinkade CW,
Aytes A, Dankort D, Lefebvre C, Mitrofanova A, Cardiff RD, McMahon
M, et al: B-Raf activation cooperates with PTEN loss to drive c-Myc
expression in advanced prostate cancer. Cancer Res. 72:4765–4776.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ling M-T, Wang X, Ouyang X-S, Lee TK, Fan
TY, Xu K, Tsao SW and Wong YC: Activation of MAPK signaling pathway
is essential for Id-1 induced serum independent prostate cancer
cell growth. Oncogene. 21:8498–8505. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kinkade CW, Castillo-Martin M, Puzio-Kuter
A, Yan J, Foster TH, Gao H, Sun Y, Ouyang X, Gerald WL,
Cordon-Cardo C, et al: Targeting AKT/mTOR and ERK MAPK signaling
inhibits hormone-refractory prostate cancer in a preclinical mouse
model. J Clin Invest. 118:3051–3064. 2008.PubMed/NCBI
|
|
36
|
Grandori C, Cowley SM, James LP and
Eisenman RN: The Myc/Max/Mad network and the transcriptional
control of cell behavior. Annu Rev Cell Dev Biol. 16:653–699. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ellwood-Yen K, Graeber TG, Wongvipat J,
Iruela-Arispe ML, Zhang J, Matusik R, Thomas GV and Sawyers CL:
Myc-driven murine prostate cancer shares molecular features with
human prostate tumors. Cancer Cell. 4:223–238. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sun C, Dobi A, Mohamed A, Li H,
Thangapazham RL, Furusato B, Shaheduzzaman S, Tan SH, Vaidyanathan
G, Whitman E, et al: TMPRSS2-ERG fusion, a common genomic
alteration in prostate cancer activates C-MYC and abrogates
prostate epithelial differentiation. Oncogene. 27:5348–5353. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rokhlin OW, Scheinker VS, Taghiyev AF,
Bumcrot D, Glover RA and Cohen MB: MicroRNA-34 mediates
AR-dependent p53-induced apoptosis in prostate cancer. Cancer Biol
Ther. 7:1288–1296. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Corney DC, Flesken-Nikitin A, Godwin AK,
Wang W and Nikitin AY: MicroRNA-34b and MicroRNA-34c are targets of
p53 and cooperate in control of cell proliferation and
adhesion-independent growth. Cancer Res. 67:8433–8438. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Notari M, Neviani P, Santhanam R, Blaser
BW, Chang JS, Galietta A, Willis AE, Roy DC, Caligiuri MA, Marcucci
G, et al: A MAPK/HNRPK pathway controls BCR/ABL oncogenic potential
by regulating MYC mRNA translation. Blood. 107:2507–2516. 2006.
View Article : Google Scholar : PubMed/NCBI
|