Introduction
Cervical cancer is the second most common cancer in
women globally, and is a significant cause of morbidity and
mortality (1,2). According to the US Center for Disease
Control and Prevention, there are >11,000 new cases of cervical
cancer diagnosed every year in the United States, and ~4,000 people
die annually as a result of this disease (3). Although invasive cervical cancer rates
have decreased steadily over recent decades, due to a lack of
screening programs, cervical cancer is predominantly detected
during advanced stages (ІІB and ІІІB) in developing countries
(4). Approximately half of patients
with advanced cervical cancer will develop recurrence or metastases
within the initial two years following completion of therapy
(5).
Molecular and epidemiological studies have revealed
that human papillomavirus (HPV) infection may be a prerequisite for
cervical cancer development, and host genetic variations that
control cell division and the maintenance of genome integrity (for
example, DNA repair) may determine the risk of an individual
developing cervical cancer (6–8). In order
to reveal a patient's genetic susceptibility to cervical cancer, it
is necessary to identify novel molecular markers that are able to
predict the development of cervical cancer.
Breast cancer 1, early onset (BRCA1)-interacting
protein 1 (BRIP1), which is also known as BRCA1-associated
C-terminal helicase 1, and located on chromosome 17q23, is part of
the DEAH helicase family. BRIP1 directly interacts with the BRCA1 C
terminus domain of BRCA1, and is important in DNA damage repair
(9–13). The specific interaction between
phosphorylated BRIP1 and BRCA1 is regulated by the cell cycle and
is crucial for DNA damage-induced checkpoint control during the
G2-to-M-phase transition (14,15). Given
the crucial function of BRIP1 in the regulation of normal cell
cycle progression and DNA repair, the BRIP1 gene represents a good
candidate for prediction of genetic susceptibility to cancer.
Accumulating evidence has suggested that BRIP1 may have an
anti-oncogenic role, and downregulation of BRIP1 has been observed
in multiple types of cancer (16–19).
The aims of the present study were to evaluate the
role of BRIP1 in the tumorigenic properties of cervical cancer, and
to identify whether the Ras homolog gene family, member A
guanosine-5′-triphosphate-ase (RhoA GTPase) has a role in the
antitumor effects of BRIP1.
Materials and methods
Reagents
Monoclonal rabbit anti-human BRIP1 (cat. no. 4578;
1:1,000), cleaved caspase-3 (cat. no. 9661; 1:1,000), B-cell
lymphoma 2 (cat. no. 2870; 1:1,000), tubulin (cat. no. 5335;
1:1,000), RhoA-GTP (cat. no. 2461; 1:1,000), RhoA (cat. no. 2117;
1:1,000), Ras-related C3 botulinum toxin substrate 1 (Rac1)-GTP
(cat. no. 8815; 1:1,000) and monoclonal mouse anti-human Rac1 (cat.
no. 4651; 1:1,000) antibodies were obtained from Cell Signaling
Technology, Inc. (Danvers, MA, USA). Lysophosphatidic acid (LPA)
was obtained from Cayman Chemical Co. (Ann Arbor, MI, USA).
Patient and tissue samples
Human cervical carcinoma samples (n=225) were
randomly collected from surgical specimens in the Department of
Obstetrics and Gynecology, Xijing Hospital (Xi'an, China) between
2012 and 2013. None of the patients had received radiotherapy or
chemotherapy prior to surgery. Additionally, normal cervical
specimens were obtained from 30 patients with uterine fibroids, who
had undergone total hysterectomy at Xijing Hospital. All tissue
samples were either immediately frozen in liquid nitrogen for later
use in reverse transcription-polymerase chain reaction (RT-PCR)
analysis, or fixed in 10% formalin and embedded in paraffin for
immunohistochemical analysis. Pathologists at Xijing Hospital
verified the diagnoses of the tissue samples. All samples were
obtained with written informed consent, and the ethics committee of
Xijing Hospital approved the protocol for the present study.
Cell culture
Normal cervix cell lines (End1 and Ect1) and a
cervical cancer cell line (HeLa) were purchased from American Type
Culture Collection (Manassas, VA, USA). The cells were maintained
in Dulbecco's modified Eagle's medium (DMEM) with high glucose
(Gibco; Thermo Fisher Scientific, Waltham, MA, USA) supplemented
with 10% fetal bovine serum (FBS; Hyclone; GE Healthcare Life
Sciences, Logan, UT, USA) at 37°C in a cell culture incubator with
an atmosphere of 5% CO2.
RT-qPCR
Total RNA was extracted using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Waltham, MA, USA), and
first-strand complementary DNA was reverse transcribed from RNA
using the Reverse Transcription System kit (Promega Corp., Madison,
WI, USA). RT-qPCR was performed using a LightCycler96® (Roche
Diagnostics GmbH, Mannheim, Germany). PCR cycling conditions were
as follows: 40 cycles at 98°C for 30 sec, 58°C for 90 sec, and 72°C
for 30 sec, with a final extension step at 72°C for 10 min. The
messenger RNA (mRNA) expression levels, which were normalized
against glyceraldehyde 3-phosphate dehydrogenase (GAPDH), were
calculated and expressed as 2−ΔΔCq. The primer sequences
utilized were as follows: BRIP1 sense, 5′-CAATGCCCGTGCTGTCA-3′ and
antisense, 5′-ATCTGCTGCCGTACCCATTTA-3′; GAPDH sense,
5′-GCACCGTCAAGGCTGAGAAC-3′ and antisense, 5′-TGGTGAAGACGCCAGTGG
A-3′ (Jie Li Biology, Shanghai, China).
Immunohistochemistry
Paraffin-embedded tissue sections (8-µm thick) were
dewaxed, rehydrated and immersed in methanol containing 0.3%
hydrogen peroxide for 30 min in order to block any endogenous
peroxidase activity. Subsequently, the slides were incubated at
89°C in 10 mM sodium citrate buffer (pH 6.0) for 30 min for antigen
retrieval. Primary rabbit anti-human BRIP1 polyclonal antibodies
(cat. no. HPA005474; Sigma-Aldrich, St. Louis, MO, USA) were
diluted to 1:100 and incubated with the tissue sections overnight
at 4°C. The slides were subsequently incubated with anti-rabbit
horseradish peroxidase-conjugated secondary antibodies (Nichirei
Bioscience Inc., Tokyo, Japan) for 1 h. Finally, the visualization
signal was developed by incubation in 3,3′-diaminobenzidine in
buffered substrate (Nichirei Bioscience) for 5 min.
Plasmid construction and
transfection
In order to generate the BRIP1 recombinant plasmid,
the full-length translated region of the BRIP1 gene was amplified
using PCR with the following primers: forward,
5′-GCAATGTCTTCAATGTGGTCT-3′ and reverse,
5′-GGATTTTACTTAAAACCAGGAAA-3′. PCR was performed under the
following conditions: 30 cycles at 98°C for 15 sec, 58°C at 90 sec,
and 72°C for 20 sec, with a final extension step at 72°C for 5 min.
The resulting PCR amplicons of BRIP1 were cloned into the pGEM®-T
vector (Promega Corp.). The correct clones were confirmed via
sequencing. The cells were grown in Opti-MEM medium (Invitrogen;
Thermo Fisher Scientific) for 24 h prior to transfection. The cells
were transfected with plasmid (200 ng) using the Lipofectamine™
2000 reagent (Invitrogen; Thermo Fisher Scientific) according to
the manufacturer's protocol. The following assays were performed 48
h subsequent to transfection.
Cell proliferation assays
HeLa cells were plated into 96-well microplates
(Beyotime Institute of Biotechnology, Haimen, China) at a density
of 103 cells/well. Cell proliferation was evaluated
using the Cell Proliferation kit I, containing
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT;
Roche Diagnostics GmbH) and was performed according to the
manufacturer's protocol. The absorbance value (A) at 570 nm was
measured using a Benchmark microplate reader (Bio-Rad Laboratories,
Inc., Hercules, CA, USA).
Transwell migration assays
Transwell migration assays were performed with
6.5-mm diameter cell culture inserts (8-µm pore size; Corning Life
Sciences, Corning, NY, USA) in 24-well culture plates. The
Transwell inserts were coated with Matrigel (80 µg/well; BD
Biosciences, San Jose, CA, USA) for the invasion assays. The cells
(5×104/well) were added to the upper chamber in 200 µl
of serum-free DMEM high glucose medium with 0.1% bovine serum
albumin (Gibco; Thermo Fisher Scientific), and subsequently placed
into 24-well plates in DMEM-high glucose medium with 10% FBS.
Following 24 h of incubation, non-migrated cells were removed from
the upper surface with cotton buds, and the cells that migrated to
the lower surface were fixed using cold methanol, stained with
crystal violet (Sigma-Aldrich) and counted using light microscopy
(Olympus BX53; Olympus Corporation, Tokyo, Japan).
Cell cycle assays
For the cell cycle assays, cells were seeded at a
density of 1×105 cells into 6-cm tissue culture dishes
(Clontech Laboratories, Mountain View, CA, USA) and cells were
synchronized at the G1 phase by serum starvation for 12 h.
Following 24 h of incubation, cells were washed using cold
phosphate-buffered saline (PBS) and fixed with 70% cold alcohol
overnight at 4°C. The fixed cells were collected, washed with PBS
and stained using propidium iodide (PI; Sigma-Aldrich) in the
presence of RNase (Sigma-Aldrich). Flow cytometric (FCM;
FACSCanto™; BD Biosciences) analysis was used to determine cell
cycle distribution with the aid of ModFit LT software (Verity
Software House, Topsham, ME, USA).
Apoptosis assays
Cell apoptosis was evaluated by FCM using an Annexin
V-fluorescein isothiocyanate (FITC) apoptosis kit (Cell Signaling
Technology) according to the manufacturer's protocol. Briefly, the
cells were washed, resuspended with binding buffer, incubated with
Annexin V-FITC and PI buffers for 15 min at 4°C in the dark and
subsequently analyzed using FCM.
Western blot analysis
The cells were lysed on ice with RIPA buffer
(Sigma-Aldrich). The protein concentration of the lysed cells was
measured using the bicinchoninic acid protein assay reagent (Pierce
Biotechnology, Inc., Rockford, IL, USA). Protein extracts (50 µg)
were separated using 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis at 100 V, transferred onto polyvinylidene fluoride
membranes (Invitrogen; Thermo Fisher Scientific) and analyzed by
western blotting with monoclonal rabbit anti-human BRIP1 (cat. no.
4578; 1:1,000), cleaved caspase-3 (cat. no. 9661; 1:1,000), B-cell
lymphoma 2 (cat. no. 2870; 1:1,000), tubulin (cat. no. 5335;
1:1,000), RhoA-GTP (cat. no. 2461; 1:1,000), RhoA (cat. no. 2117;
1:1,000), Ras-related C3 botulinum toxin substrate 1 (Rac1)-GTP
(cat. no. 8815; 1:1,000) and monoclonal mouse anti-human Rac1 (cat.
no. 4651; 1:1,000) antibodies (Cell Signaling Technology, Inc).
Bands were detected with an enhanced chemiluminescence kit (GE
Healthcare Life Sciences, Chalfont, UK) and visualized using the
ChemiDoc™ XRS system (Bio-Rad Laboratories, Inc.).
Statistical analysis
The data obtained in the present study was analyzed
using the statistical package SPSS version 17.0 (SPSS Inc.,
Chicago, IL, USA). Data are expressed as the mean ± standard
deviation from at least three independent experiments. For values
that were normally distributed, a direct comparison between two
groups was performed using an unpaired Student's t-test, and
one-way analysis of variance with Dunnett's post-hoc test was
utilized to compare the means of three or more groups. P<0.05
was considered to indicate a statistically significant
difference.
Results
Expression of BRIP1 in human cervical
cancer tissues
In order to investigate the characteristics of BRIP1
expression in normal cervix and cervical cancer tissues, 30 normal
cervix tissue samples and 225 cervical cancer samples were analyzed
using RT-qPCR. The clinicopathological data from the 255 cervical
cancer samples is summarized in Table
I. Decreased BRIP1 mRNA expression was observed in the cervical
cancer patient group. Furthermore, the reduction in BRIP1
expression correlated significantly with unfavorable variables,
including the International Federation of Gynecologists and
Obstetricians (FIGO) stage and the presence of lymph node
metastases. However, no significant differences were observed
between the groups when compared according to any other
clinicopathological feature, including age, histological type and
level of differentiation.
 | Table I.Correlation of BRIP1 expression with
clinicopathological features in cervical cancer. |
Table I.
Correlation of BRIP1 expression with
clinicopathological features in cervical cancer.
| Clinicopathological
feature | No. of samples | Relative BRIP1
expression | P-value |
|---|
| Tissue type |
|
|
|
| Normal
cervix | 30 | 1.215±0.249 |
|
|
Carcinoma | 225 | 0.587±0.176 | 0.002 |
| Age, years |
|
|
|
|
>40 | 136 | 0.647±0.134 |
|
| ≤40 | 89 | 0.556±0.146 | 0.035 |
| Histologic type |
|
|
|
| Squamous
cancer | 125 | 0.595±0.204 |
|
|
Adenocarcinoma | 79 | 0.573±0.108 | 0.418 |
|
Other | 21 | 0.571±0.127 | 0.580 |
| Degree of
differentiation |
|
|
|
| Well | 35 | 0.615±0.122 |
|
|
Moderate | 108 | 0.633±0.143 | 0.539 |
|
Poor | 82 | 0.645±0.173 | 0.783 |
| FIGO stage |
|
|
|
| I | 141 | 0.778±0.103 |
|
| II | 72 | 0.623±0.098 | 0.022 |
|
III–IV | 12 | 0.402±0.189 | 0.009 |
| Lymph node
metastasis |
|
|
|
|
Negative | 132 | 0.678±0.132 |
|
|
Positive | 93 | 0.439±0.139 | 0.003 |
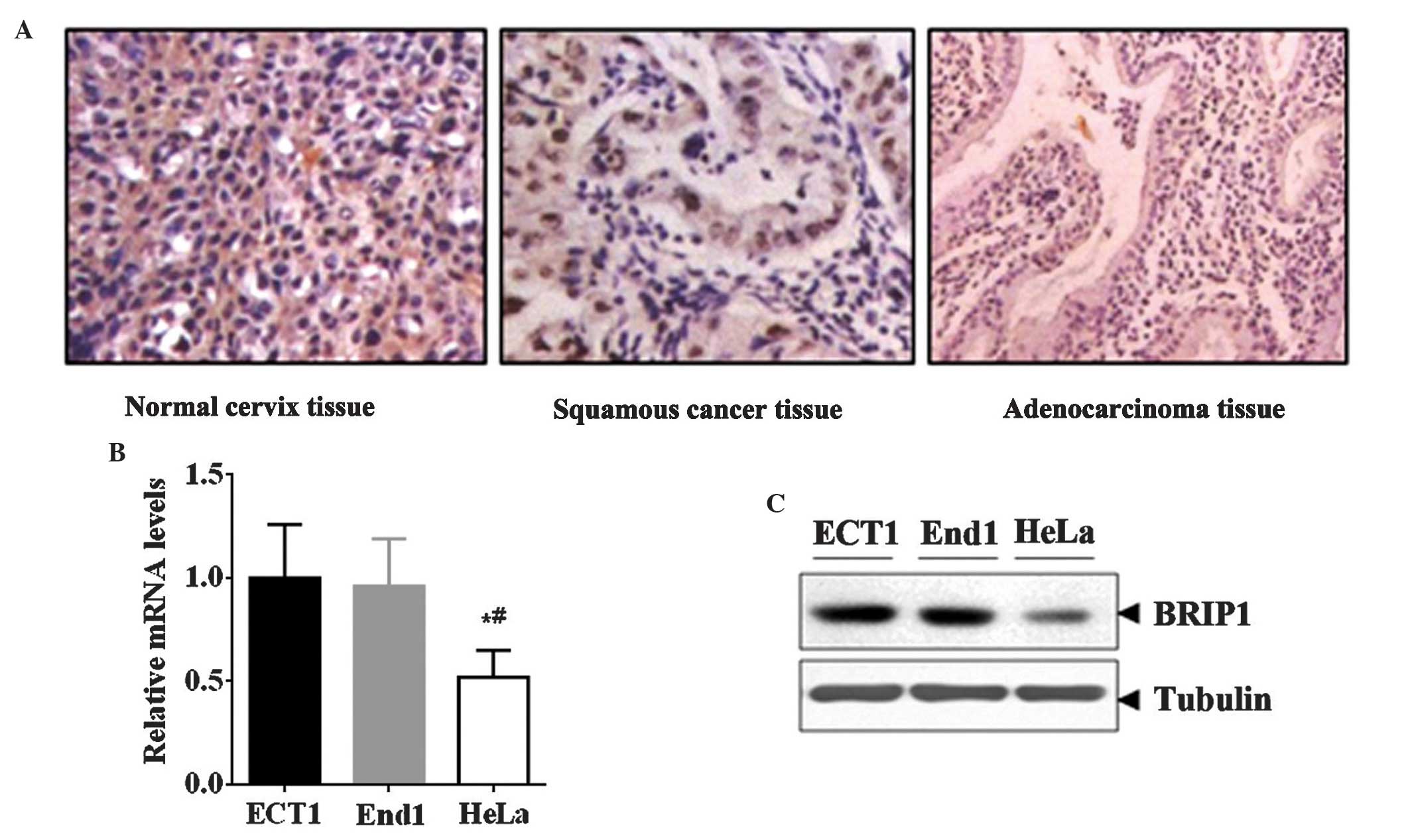
In addition, immunostaining data revealed that the
expression of BRIP1 protein was markedly decreased in the cervical
cancer samples (squamous cancer and adenocarcinoma) compared with
the normal cervix tissue samples (Fig.
1A). Furthermore, the expression of BRIP1 was detected in
normal cervical cell lines (End1 and Ect1) and a cervical cancer
cell line (HeLa). In line with these results, HeLa cells exhibited
lower BRIP1 mRNA and protein levels compared with End1 and Ect1
cells (P<0.05; Fig. 1B and C).
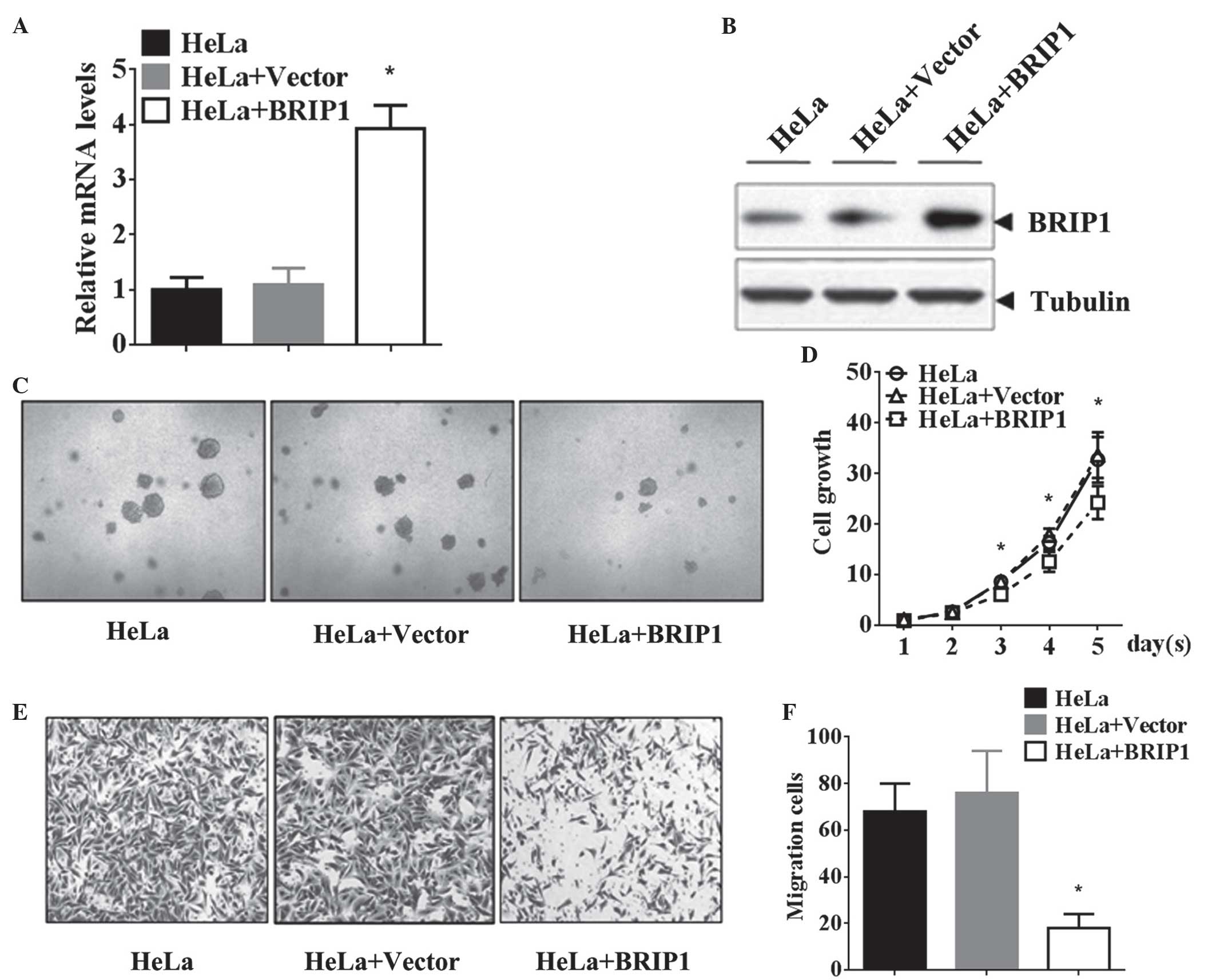
Overexpression of BRIP1 inhibits the
tumorigenic properties of HeLa cells
In order to elucidate the role of BRIP1 in cervical
cancer, BRIP1 was overexpressed in HeLa cells via transfection with
the BRIP1 recombinant plasmid. RT-qPCR and western blot analyses
revealed that BRIP1 mRNA and protein levels were markedly elevated
following BRIP1 recombinant plasmid transfection, compared with the
control groups (P<0.05; Fig. 2A and
B). The MTT proliferation assay results revealed that BRIP1
overexpression significantly suppressed the growth of HeLa cells in
a time-dependent manner (P<0.05; Fig.
2C and D). Similarly, ectopic expression of BRIP1 attenuated
the invasive ability of HeLa cells, led to a reduction in the
number of invasive cells (P<0.05; Fig.
2E and F). Additionally, BRIP1 overexpression in HeLa cells
decreased the cell adhesion rate at the indicated time points (20,
40 and 60 min; Table II).
Collectively, the results of the present study confirmed that BRIP1
overexpression significantly inhibited the tumorigenic properties
of HeLa cells in vitro.
| Table II.Impact of BRIP1 expression on cell
adhesion, n=5. |
Table II.
Impact of BRIP1 expression on cell
adhesion, n=5.
|
| Cell adhesion rate,
% |
|---|
|
|
|
|---|
| Group | 20 min | 40 min | 60 min |
|---|
| HeLa | 11.5±0.12 | 35.7±2.33 | 53.8±2.13 |
| HeLa+Vector | 10.3±0.98 | 38.6±3.18 | 51.5±1.48 |
| HeLa+BRIP1 |
8.6±1.24a |
21.7±2.41a |
38.6±2.37a |
| P-value | 0.003 | 0.001 | 0.001 |
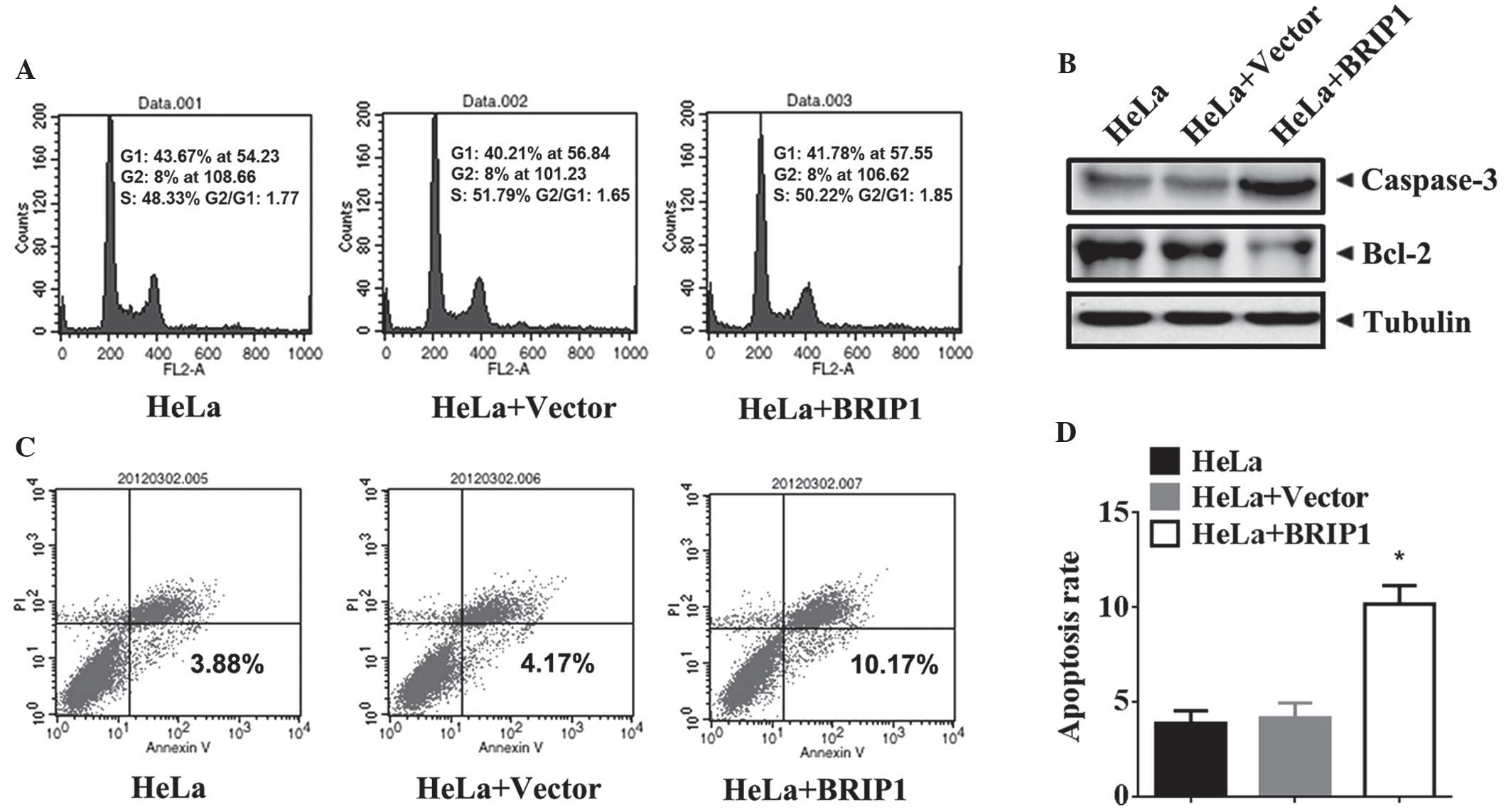
BRIP1 induces apoptosis in HeLa
cells
In order to identify the antitumorigenic mechanism
of BRIP1 in cervical cancer, the cell cycle and cell apoptosis were
analyzed in HeLa cells following transfection with BRIP1 and
control vectors. The results demonstrate that ectopic expression of
BRIP1 in HeLa cells did not affect the cell cycle distribution
(Fig. 3A). In addition, the levels of
pro-apoptotic proteins (caspase-3 and Bcl-2) were detected in
BRIP1-overexpressing HeLa cells. Western blot analysis demonstrated
that BRIP1 overexpression upregulated cleaved caspase-3 and
downregulated Bcl-2 protein in HeLa cells (Fig. 3B). Moreover, BRIP1 overexpression in
HeLa cells significantly increased the percentage of apoptotic
cells (P<0.05; Fig. 3C and D).
Thus, BRIP1 overexpression-induced cell apoptosis may suppress the
growth of HeLa cells.
RhoA GTPases are critical for the
antitumor effects of BRIP1 in HeLa cells
The alteration of Rho GTPase activity in
BRIP1-overexpressing HeLa cells was assessed. The western blotting
results demonstrated that BRIP1 overexpression was able to
significantly reduce the levels of RhoA-GTP in HeLa cells (Fig. 4A). In order to determine whether the
inhibitory tumorigenic properties obtained by BRIP1 overexpression
were associated with the activity of RhoA GTPase,
BRIP1-overexpressing HeLa cells were treated with the RhoA GTPase
agonist LPA, and their tumorigenic properties were then evaluated.
As demonstrated in Fig. 4B, the
growth of the BRIP1-overexpressing HeLa cells was markedly elevated
following LPA treatment (P<0.05). Similar results were observed
in the Transwell migration assay (P<0.05; Fig. 4C). The apoptosis assay revealed that
LPA treatment decreased the levels of cleaved caspase-3, increased
Bcl-2 protein levels (Fig. 4D) and
reversed the apoptotic inhibitory effect of BRIP1 overexpression
(P<0.05; Fig. 4E).
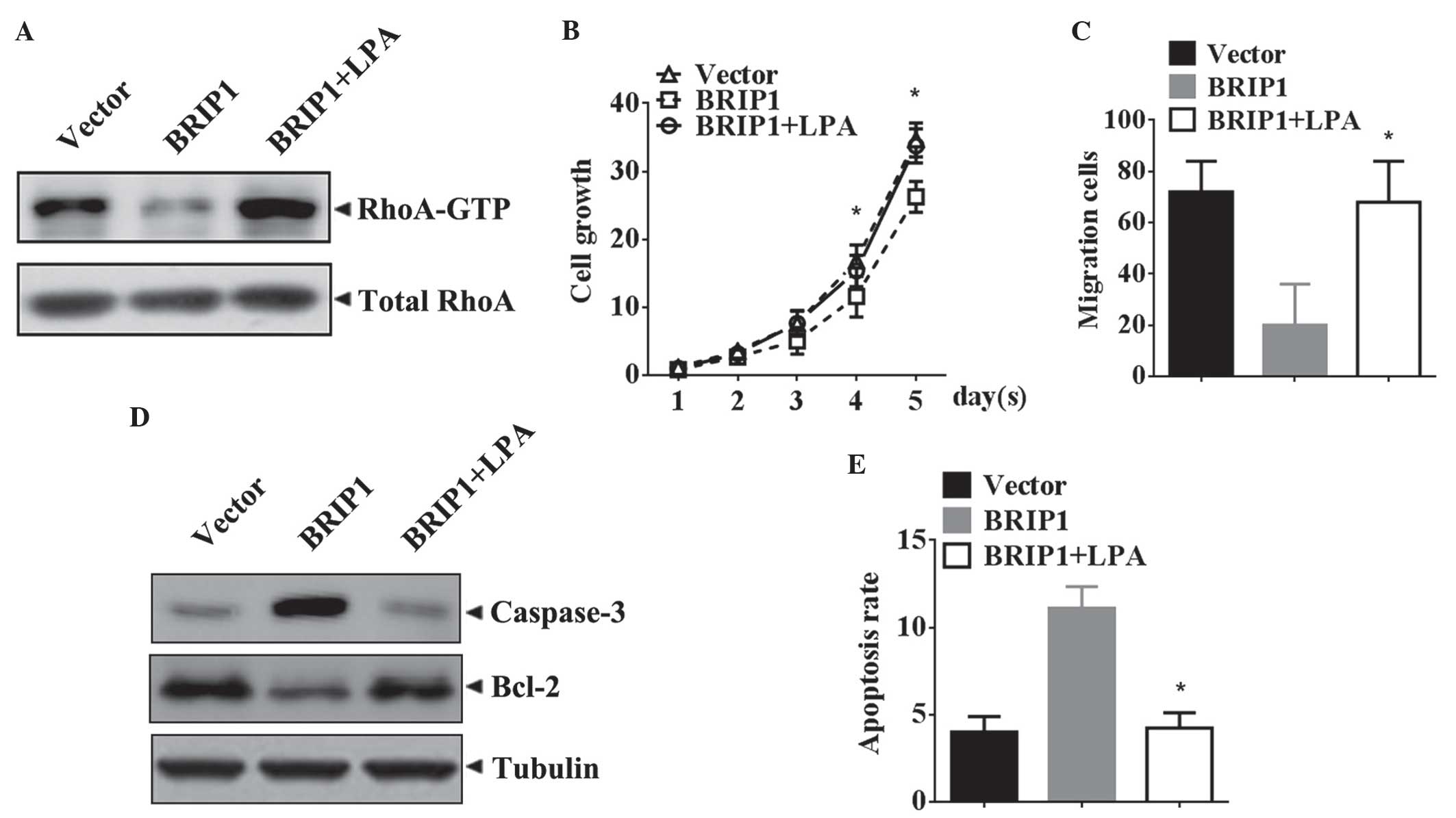 | Figure 4.Attenuation in tumorigenic properties
obtained by BRIP1 overexpression is reversed by LPA, an agonist of
RhoA GTPase. Forty-eight hours subsequent to the transfection of
HeLa cells with BRIP1, the cells were pretreated with saline or LPA
(5 µM) for 24 h. (A) Expression of RhoA-GTP and total RhoA was
analyzed by western blotting. (B) Cell growth was monitored by
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide.
Absorbance at day 0 was assigned a value of 1. Data are expressed
as the mean±standard deviation (n=3). *P<0.05 vs. the
BRIP1+saline group. (C) Quantification of transmembrane HeLa cells.
Data are expressed as the mean±standard deviation (n=3). *P<0.05
vs. the BRIP1+saline group. (D) Western blot analysis of cleaved
caspase-3 and Bcl-2 expression. Tubulin was analyzed as the loading
control. (E) Quantification of the rate of apoptosis of HeLa cells.
Data are expressed as the mean±standard deviation (n=3). *P<0.05
vs. the BRIP1+saline group. BRIP1, breast cancer 1, early
onset-interacting protein 1; LPA, Lysophosphatidic acid; Bcl-2,
B-cell lymphoma 2; RhoA GTP, Ras homolog gene family, member A
guanosine-5′-triphosphatease. |
Discussion
Cervical cancer remains a significant health problem
for women in developing countries (20). Although there have been significant
advances in the diagnosis of cervical cancer over the last decade,
cervical cancer has a high mortality rate, as the majority of
patients are initially diagnosed in the advanced stages of the
disease (4,21). The treatment for pre-invasive lesions
is typically based on surgery, and for invasive cervical cancers,
the treatment consists of surgery and/or radiation and
cisplatin-based chemotherapy (22,23).
However, a large proportion of patients will go on to develop drug
resistance during treatment (24).
In-depth investigations concerning the molecular mechanisms of
cervical cancer are required to promote the development of novel
effective molecular targeted drugs.
The BRIP1 gene encodes a helicase that interacts
with BRCA1, which is crucial for DNA repair and genomic stability
(14). Studies have suggested that
BRIP1 possesses an anti-oncogenic role, and that frequently
observed single nucleotide polymorphisms (SNPs) in the BRIP1 gene
may be associated with susceptibility to breast, prostate and
ovarian cancer (18,25,26).
Current evidence has revealed that SNPs in the BRIP1 gene may be
capable of influencing cervical cancer susceptibility and that
BRIP1 gene variations are potentially implicated in the development
and progression of cervical cancer (14,15). The
results of the present study support the observations made in
previous studies, and furthermore, have identified that BRIP1 is
downregulated in cervical carcinoma tissue. In addition,
significant correlations were observed between the reduction in
BRIP1 expression and unfavorable variables, including FIGO stage
and presence of lymph node metastases. Functionally, the
plasmid-mediated overexpression of BRIP1 inhibited the
carcinogenesis of cervical cancer, which suggested that the BRIP1
gene may possess the role of a tumor suppressor in cervical cancer
development.
The present study additionally identified that BRIP1
overexpression suppressed RhoA GTPase activity in cervical cancer
cells, and that the inhibitory tumorigenic properties obtained as a
result of BRIP1 overexpression were significantly reversed by the
activation of RhoA GTPase. RhoA GTPase is part of the Ras
superfamily of small GTPases, and its activation can induce the
assembly of basal stress fibers and cytoskeleton-mediated changes
in cell motility (27). A number of
previous studies have supported the role of RhoA GTPase in cell
proliferation, adhesion, apoptosis, cell polarity, invasion and
metastasis (28–30). A previous study provided evidence that
RhoA GTPase may be associated with metastasis of cervical cancer
and that its functioning is mediated by a mitogen-activated protein
kinase/extracellular signal-regulated kinase-dependent mechanism
(31,32), implying that the involvement of
alterations in the activation of RhoA GTPase in events may have a
role in cervical cancer carcinogenesis. Overall, BRIP1 suppressed
the tumorigenic properties of cervical cancer via the
downregulation of RhoA GTPase activity.
In conclusion, the results of the present study may
provide some insight into the functioning of BRIP1 in cervical
cancer. BRIP1 may act as an essential DEAH helicase that is capable
of regulating the tumorigenic properties of cervical cancer by
controlling RhoA GTPase. BRIP1 may prove to be a potential tumor
suppressor gene in cervical cancer and may assist with uncovering
further associations between RhoA GTPase and cervical cancer. BRIP1
may constitute a promising candidate for cervical cancer
therapy.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81101959 and
81101958).
References
|
1
|
Arbyn M, Castellsagué X, de Sanjosé S,
Bruni L, Saraiya M, Bray F and Ferlay J: Worldwide burden of
cervical cancer in 2008. Ann Oncol. 22:2675–2686. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J and
Thun MJ: Cancer statistics, 2009. CA Cancer J Clin. 59:225–249.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Balacescu O, Balacescu L, Tudoran O, Todor
N, Rus M, Buiga R, Susman S, Fetica B, Pop L, Maja L, et al: Gene
expression profiling reveals activation of the FA/BRCA pathway in
advanced squamous cervical cancer with intrinsic resistance and
therapy failure. BMC Cancer. 14:2462014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lippert TH, Ruoff HJ and Volm M: Intrinsic
and acquired drug resistance in malignant tumors. The main reason
for therapeutic failure. Arzneimittelforschung. 58:261–264.
2008.PubMed/NCBI
|
|
6
|
Benjamin I, Saigo P, Finstad C, Takahashi
H, Federici M, Rubin SC and Boyd J: Expression and mutational
analysis of P53 in stage IB and IIA cervical cancers. Am J Obstet
Gynecol. 175:1266–1271. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kim JW, Roh JW, Park NH, Song YS, Kang SB
and Lee HP: Polymorphism of TP53 codon 72 and the risk of cervical
cancer among Korean women. Am J Obstet Gynecol. 184:55–58. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Singh H, Sachan R, Devi S, Pandey SN and
Mittal B: Association of GSTM1, GSTT1, and GSTM3 gene polymorphisms
and susceptibility to cervical cancer in a North Indian population.
Am J Obstet Gynecol. 198:303.e1–6. 2008. View Article : Google Scholar
|
|
9
|
Cantor SB, Bell DW, Ganesan S, Kass EM,
Drapkin R, Grossman S, Wahrer DC, Sgroi DC, Lane WS, Haber DA and
Livingston DM: BACH1, a novel helicase-like protein, interacts
directly with BRCA1 and contributes to its DNA repair function.
Cell. 105:149–160. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yu X, Chini CC, He M, Mer G and Chen J:
The BRCT domain is a phospho-protein binding domain. Science.
302:639–642. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cantor S, Drapkin R, Zhang F, Lin Y, Han
J, Pamidi S and Livingston DM: The BRCA1-associated protein BACH1
is a DNA helicase targeted by clinically relevant inactivating
mutations. Proc Natl Acad Sci USA. 101:2357–2362. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Peng M, Litman R, Jin Z, Fong G and Cantor
SB: BACH1 is a DNA repair protein supporting BRCA1 damage response.
Oncogene. 25:2245–2253. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tu Z, Aird KM, Bitler BG, Nicodemus JP,
Beeharry N, Xia B, Yen TJ and Zhang R: Oncogenic RAS regulates
BRIP1 expression to induce dissociation of BRCA1 from chromatin,
inhibit DNA repair, and promote senescence. Dev Cell. 21:1077–1091.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ma XD, Cai GQ, Zou W, Huang YH, Zhang JR,
Wang DT and Chen BL: First evidence for the contribution of the
genetic variations of BRCA1-interacting protein 1 (BRIP1) to the
genetic susceptibility of cervical cancer. Gene. 524:208–213. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ma XD, Cai GQ, Zou W, Huang YH, Zhang JR,
Wang DT and Chen BL: BRIP1 variations analysis reveals their
relative importance as genetic susceptibility factor for cervical
cancer. Biochem Biophys Res Commun. 433:232–236. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Narayan G, Arias-Pulido H, Nandula SV,
Basso K, Sugirtharaj DD, Vargas H, Mansukhani M, Villella J, Meyer
L, Schneider A, et al: Promoter hypermethylation of FANCF:
Disruption of Fanconi Anemia-BRCA pathway in cervical cancer.
Cancer Res. 64:2994–2997. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Levitus M, Waisfisz Q, Godthelp BC, de
Vries Y, Hussain S, Wiegant WW, Elghalbzouri-Maghrani E,
Steltenpool J, Rooimans MA, Pals G, et al: The DNA helicase BRIP1
is defective in Fanconi anemia complementation group J. Nat Genet.
37:934–935. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Song H, Ramus SJ, Kjaer SK, Hogdall E,
Dicioccio RA, Whittemore AS, McGuire V, Hogdall C, Jacobs IJ,
Easton DF, et al: Tagging single nucleotide polymorphisms in the
BRIP1 gene and susceptibility to breast and ovarian cancer. PLoS
One. 2:e2682007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rafnar T, Gudbjartsson DF, Sulem P,
Jonasdottir A, Sigurdsson A, Jonasdottir A, Besenbacher S, Lundin
P, Stacey SN, Gudmundsson J, et al: Mutations in BRIP1 confer high
risk of ovarian cancer. Nat Genet. 43:1104–1107. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Comparetto C and Borruto F: Cervical
cancer screening: A never-ending developing program. World J Clin
Cases. 3:614–624. 2015.PubMed/NCBI
|
|
21
|
Schiffman M, Wentzensen N, Wacholder S,
Kinney W, Gage JC and Castle PE: Human papillomavirus testing in
the prevention of cervical cancer. J Natl Cancer Inst. 103:368–383.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Salicrú SR, de la Torre JF and Gil-Moreno
A: The surgical management of early-stage cervical cancer. Curr
Opin Obstet Gynecol. 25:312–319. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Movva S, Rodriguez L, Arias-Pulido H and
Verschraegen C: Novel chemotherapy approaches for cervical cancer.
Cancer. 115:3166–3180. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Casagrande N, De Paoli M, Celegato M,
Borghese C, Mongiat M, Colombatti A and Aldinucci D: Preclinical
evaluation of a new liposomal formulation of cisplatin, lipoplatin,
to treat cisplatin-resistant cervical cancer. Gynecol Oncol.
131:744–752. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kote-Jarai Z, Jugurnauth S, Mulholland S,
Leongamornlert DA, Guy M, Edwards S, Tymrakiewitcz M, O'Brien L,
Hall A, Wilkinson R, et al: UKGPCS Collaborators; British
Association of Urological Surgeons' Section of Oncology: A
recurrent truncating germline mutation in the BRIP1/FANCJ gene and
susceptibility to prostate cancer. Br J Cancer. 100:426–430. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wong MW, Nordfors C, Mossman D,
Pecenpetelovska G, Avery-Kiejda KA, Talseth-Palmer B, Bowden NA and
Scott RJ: BRIP1, PALB2, and RAD51C mutation analysis reveals their
relative importance as genetic susceptibility factors for breast
cancer. Breast Cancer Res Treat. 127:853–859. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Leve F and Morgado-Díaz JA: Rho GTPase
signaling in the development of colorectal cancer. J Cell Biochem.
113:2549–2559. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tas PW, Gambaryan S and Roewer N: Volatile
anesthetics affect the morphology of rat glioma C6 cells via RhoA,
ERK, and Akt activation. J Cell Biochem. 102:368–376. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Struckhoff AP, Rana MK and Worthylake RA:
RhoA can lead the way in tumor cell invasion and metastasis. Front
Biosci (Landmark Ed). 16:1915–1926. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu M, Lang N, Chen X, Tang Q, Liu S,
Huang J, Zheng Y and Bi F: miR-185 targets RhoA and Cdc42
expression and inhibits the proliferation potential of human
colorectal cells. Cancer Lett. 301:151–160. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hamadmad SN and Hohl RJ: Erythropoietin
stimulates cancer cell migration and activates RhoA protein through
a mitogen-activated protein kinase/extracellular signal-regulated
kinase-dependent mechanism. J Pharmacol Exp Ther. 324:1227–1233.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
He M, Cheng Y, Li W, Liu Q, Liu J, Huang J
and Fu X: Vascular endothelial growth factor C promotes cervical
cancer metastasis via up-regulation and activation of
RhoA/ROCK-2/moesin cascade. BMC Cancer. 10:1702010. View Article : Google Scholar : PubMed/NCBI
|