Introduction
Aerobic glycolysis, first studied by Otto Warburg,
is the process whereby tumor cells use glycolysis with reduced
mitochondrial oxidative phosphorylation to metabolize glucose, even
under aerobic conditions (1). The
metabolic switch may be required to support cell growth more than
the defects in mitochondrial respiration. In certain cancer cell
lines, the most glycolytic tumor cells were demonstrated to also be
the most aggressive cells (2). In
another study, aerobic glycolysis was an essential step that
ultimately resulted in tumorigenesis, which was characterized by
enhanced activity of glycolytic enzymes and distinct changes in the
glycolytic isoenzyme (3). There were
three rate-limiting enzymes of glycolysis, consisting of
hexokinase, phosphofructokinase and pyruvate kinase (4). Among these enzymes, pyruvate kinase
catalyzed the final step of glycolysis, which is associated with
the production of adenosine triphosphate and pyruvate, and was
important during the process of glycolysis (5). Four pyruvate kinase isoenzymes are known
in mammals: Liver-type pyruvate kinase (PKL) is expressed in the
kidney and liver; protein kinase R (PKR) is expressed in red blood
cells; the M1 isoform of pyruvate kinase (PKM1) is expressed in the
majority of adult tissues; and the M2 isoform of pyruvate kinase
(PKM2) is the embryonic form, and is a splice variant of PKM1
(6). PKM2 was overexpressed in
numerous cancer cells. The expression of PKM2 was critical for
rapid growth in cancer cells (7,8). However,
inhibition of PKM2 may undesirably affect cancer cell metabolism
(9–11).
Gastric cancer is one of the most common cancers,
and is a leading cause of cancer-associated mortality in the world,
accounting for ~8% of novel cancers (12). Numerous trials have demonstrated that
PKM2 is overexpressed in gastric cancer and that the inhibition of
PKM2 has well-supported anti-tumor effects (13–15). The
critical role of PKM2 in gastric cancer suggests that identifying
novel inhibitors of PKM2 may provide a promising future in the
treatment of gastric cancer.
Proton pump inhibitors (PPIs) are highly effective
in the treatment and symptomatic relief of peptic ulcers, by
inhibiting H+,K+-adenosine triphosphate
(ATP)ase in the gastric parietal cells (16). The effects of pantoprazole (PPZ), one
type of PPI, on PKM2 were hypothesized to be associated with the
inhibitory efficacy of PPZ (17,18). Yeo
et al reported that PPZ selectively induced apoptotic cell
death in in vivo and in vitro gastric cancer models
(19), suggesting that PPIs may exert
selective anticancer effects. However, there are few relevant
studies, and the mechanisms involved remain unclear.
The present study aimed to determine whether PPZ
inhibits proliferation and selectively induces apoptosis in
SGC-7901 cells, possibly via a PKM2-mediated mechanism. The
hypothesis was examined by altering the expression of PKM2 in
vitro.
Materials and methods
Cell line and cell culture
The human gastric adenocarcinoma cell line SGC-7901
was kindly provided by the Department of Oncology, Medical School,
Drum Tower Hospital of Nanjing University (Nanjing, Jiangsu,
China). The cells were cultured in RPMI-1640 (Hyclone, Logan, UT,
USA) supplemented with 10% fetal bovine serum (Hangzhou Sijiqing
Biological Engineering Materials, Hangzhou, Zhejiang, China) and
antibiotics (100 units/ml penicillin and 100 µg/ml streptomycin) in
a humidified air with 5% CO2 at 37°C (Direct Heat
CO2) (Thermo Fisher Scientific, Waltham, MA, USA).
Drugs and experimental design
Pantoprazole sodium salts (Takeda Pharmaceuticals
International GmbH, Zurich, Switzerland) were resuspended in normal
saline (0.85%) at 5 mg/ml immediately prior to use. When the
SGC-7901 cells had reached 60–70% confluence, PPZ was added at a
final concentration of 20 µg/ml, and the cells were cultured for
additional experiments.
Transfection
The human gene encoding PKM2 was amplified using
reverse transcription-polymerase chain reaction (RT-PCR) and the
total-RNA extracted from the SGC-7901 cells. The PKM2 gene was
ligated to an internal ribosome entry site (pIRES2)-enhanced green
fluorescent protein (EGFP) plasmid (Invitrogen; Thermo Fisher
Scientific) using T4 DNA ligase following digestion by the
restricted endonucleases EcoRI and BamHI. SGC-7901
cells were transfected with the pIRES2-EGFP-PKM2 expression vector
using Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific),
according to the manufacturer's protocol for 24 h prior to PPZ
treatment.
Cell proliferation assay
The SGC-7901 cells were seeded in 100 µl of medium
per well, at a density of 1×104/well, in 96-well plates
and treated with 5 mg/ml PPZ for 24 h. The cytotoxicity of PPZ was
assessed by a cell counting kit-8 assay (KeyGen Biotech Co., Ltd.,
Nanjing, Jiangsu, China), according to the manufacturer's protocol.
The cytotoxicity was expressed as the relative cell viability,
using the following formula: Cell viability (%) = (OD of
drug-treated sample / OD of untreated sample) × 100. The experiment
was repeated 3 times.
Annexin V-fluorescein isothiocyanate
(FITC) apoptosis detection
The SGC-7901 cells were seeded into 6-well plates.
Following treatment with 5 mg/ml PPZ, the cells were trypsinized,
washed twice with phosphate-buffered saline (PBS), resuspended and
then stained with an Annexin V-FITC apoptosis detection kit
(Nanjing KeyGen Biotech Co., Ltd.). The apoptosis of cells was
analyzed using a flow cytometer (BD Biosciences, Franklin Lakes,
NJ, USA). The experiment was repeated 3 times.
Western blot analysis
The expression of PKM2 was evaluated using western
blot analysis. The cells were kept on ice for 30 min in lysate
containing 0.01% phenylmethanesulfonylfluoride, 150 mM NaCl, 50 mM
Tris-HCl, pH 8.0, 0.1% sodium dodecyl sulfate (SDS), 0.2%
ethylenediaminetetraacetic acid, 1% Triton X-100 and 1% sodium
deoxycholate, supplemented with the protease inhibitors leupeptin,
aprotinin, phenylmethylsulfonyl fluoride and sodium orthovanadate
(Roche Diagnostics, Basel, Switzerland). The lysed cells were then
centrifuged at 9,660 × g at 4°C (Centrifuge 5804R; Eppendorf,
Hamburg, Germany) for 15 min, leaving a supernatant of extracted
proteins. The protein content was estimated according to the
bicinchoninic acid protein assay kit (Nanjing KeyGen Biotech Co.,
Ltd.). In total, 50 µm of each protein sample was subjected to
western blotting on a denaturing 10% SDS-polyacrylamide gel
electrophoresis and then transferred to polyvinylidene fluoride
membranes (Immobilon-P; EMD Millipore, Billerica, MA, USA) using a
semidry transfer system (Bio-Rad Laboratories Inc., Hercules, CA,
USA). The non-specific binding was blocked by the incubation of
membranes in 1X Tris-buffered saline containing 0.05% Tween-20 with
5% nonfat dry milk for 1 h. Subsequently, the membranes were
incubated overnight at 4°C with monoclonal rabbit antibodies
against PKM2 (dilution, 1:1,000; no. 4053; Cell Signaling
Technology Inc., Danvers, MA, USA, USA) and glyceraldehyde
3-phosphate dehydrogenase (dilution, 1:3,000; no. sc-51905; Santa
Cruz Biotechnology, Inc., Dallas, TX, USA), which was used as an
internal control for protein loading. Antibody binding was detected
by incubating the blot with a horseradish peroxidase-conjugated
goat anti-rabbit antibody (dilution, 1:1,000; KPL, Gaithersburg,
MD, USA). The specific signals were visualized by enhanced
chemiluminescence (Santa Cruz Biotechnology, Inc.). Digital images
were obtained and analyzed by Quantity One V4.31 (Bio-Rad
Laboratories, Inc.).
Immunofluorescence staining
analysis
Dispersed cells were grown on glass slides within
6-well plates and were then treated with 5 mg/ml PPZ for 24 h.
Subsequent to washing with PBS 3 times, the cells were fixed with
cold acetone for 10 min at 4°C. Next, the cells were blocked with
10% normal goat serum (Wuhan Boster Biological Technology, Wuhan,
China) for 30 min and probed with the antibody against PKM2
(dilution, 1:100) at 4°C overnight. Alexa Fluor dye-conjugated
secondary antibodies (dilution, 1:100; Alexa Fluor 594) and 2 mg/ml
goat anti-rabbit IgG (H + L) highly cross-adsorbed (Molecular
Probes; Thermo Fisher Scientific) were incubated with the cells for
1 h to enable the samples to be visualized under a fluorescent
microscope (Axio Imager A1; Carl Zeiss AG, Oberkochen, Germany).
The nuclei were stained using Invitrogen
4′,6-diamidino-2-phenylindole (2 µg/ml; Thermo Fisher
Scientific).
Statistical analysis
The data were analyzed using SPSS software, version
16.0 (SPSS Inc., Chicago, IL, USA) and the independent-samples
t-test method. A P-value of <0.05 was considered to
indicate a statistically significant difference.
Results
Effects of PPZ and PKM2 on cell
proliferation
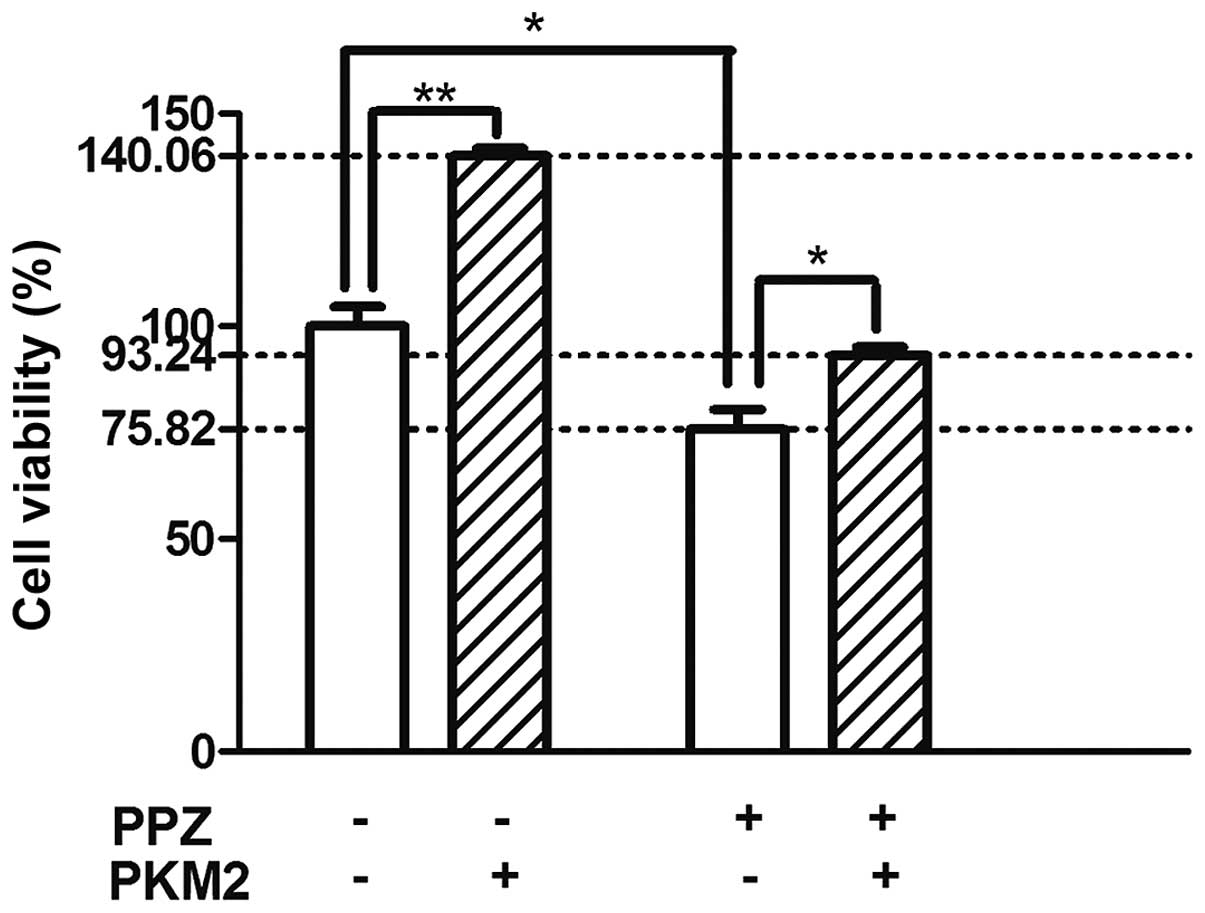
As demonstrated in Fig.
1, PPZ may inhibit the proliferation of the SGC-7901 cells. The
cell viability in the PPZ+ group was significantly
decreased compared with the PPZ− group (−24.18% vs. the
PPZ− group; P<0.05). In addition, PKM2 transfection
may notably increase cell viability (+40.06% vs. the
PKM2− group; P<0.01). As expected, the cell viability
in the PKM2+ group was increased compared with the
viability in the PKM2− group subsequent to PPZ
intervention (+17.42% vs. the PKM2− group;
P<0.05).
Effect of PPZ and PKM2 on apoptosis in
SGC-7901 cells
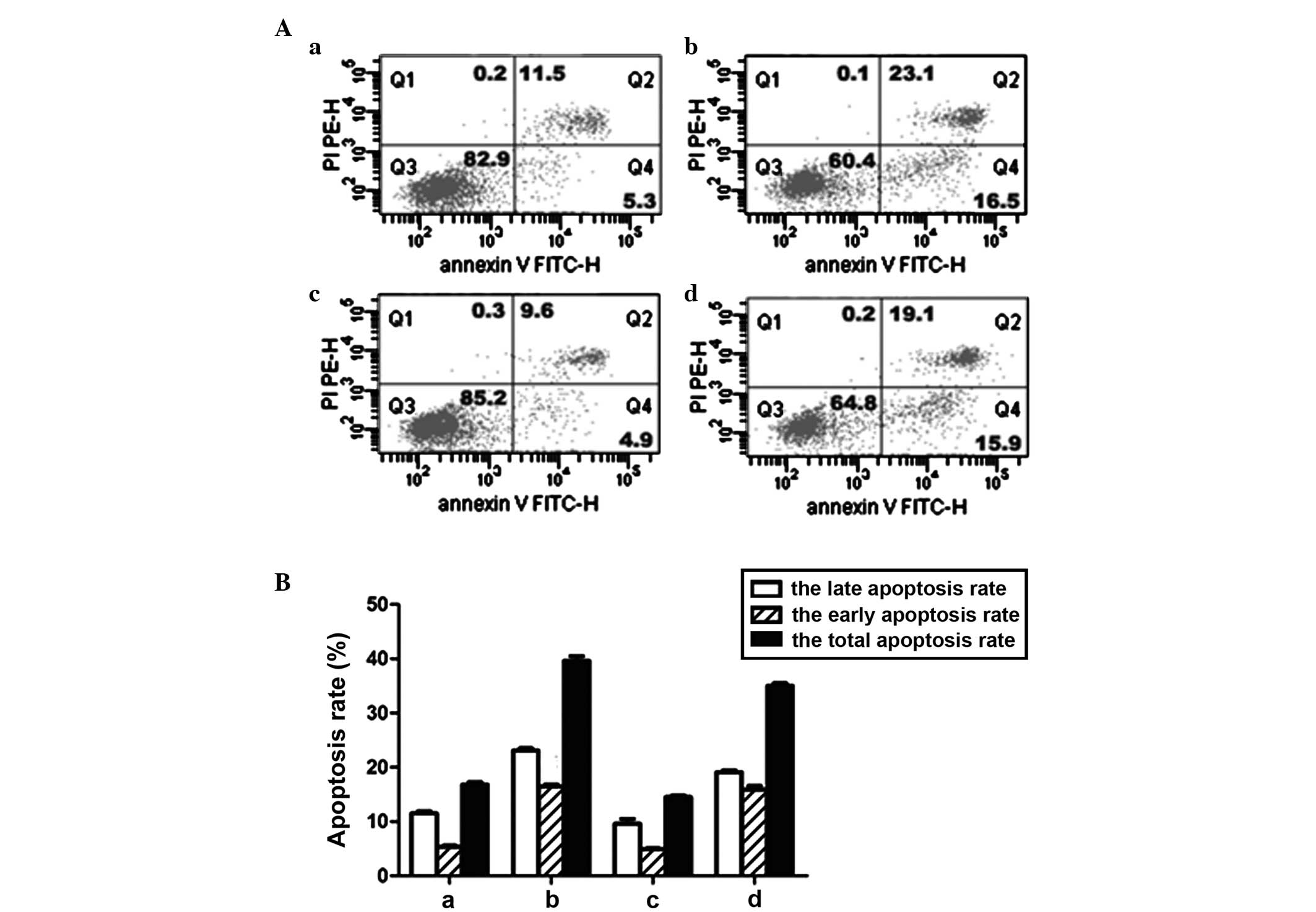
A quantitative analysis of the fluorescent signals
was performed by fluorescence-activated cell sorting (FACS). The
results were summarized in Fig. 2A and
B. As exhibited in Fig. 2B, the
early apoptosis, late apoptosis and total apoptosis rates in the
PPZ+ group were notably increased compared with the
PPZ− group (+1.2, +11.6 and +22.8%, respectively, vs.
the PPZ− group; P<0.001). Following PKM2
transfection, the early and late apoptosis rates were not affected
(P>0.05), while the total apoptosis rate was statistically
decreased compared with the PKM2− group (−2.3% vs. the
PKM2− group; P<0.05). Under PPZ intervention, there
was no difference between the early apoptosis rates of the
PKM2+ and PKM2− groups (P>0.05), while the
apoptosis rate was decreased compared with the PKM2−
group on the late apoptosis rate (−4.0% vs. the PKM2−
group; P<0.01) and total apoptosis rate (−4.6% vs. the
PKM2− group; P<0.05).
Effects of PPZ treatment and PKM2
transfection on protein expression of PKM2 in SGC-7901 cells
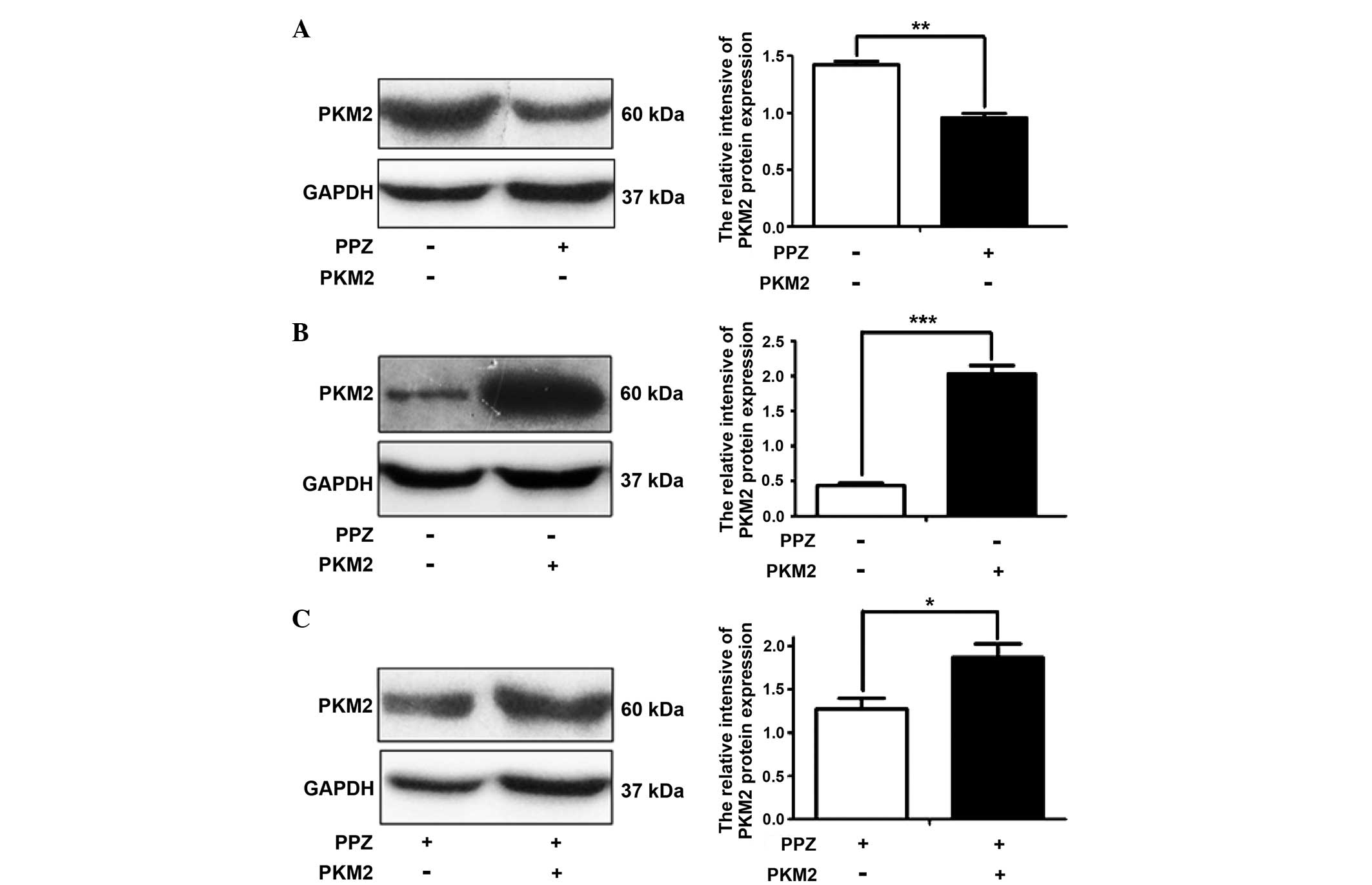
The expression of PKM2 in SGC-7901 cells was
examined using western blot analysis. Following PPZ treatment
without PKM2 transfection, the expression of PKM2 decreased when
compared to that in the control group (P<0.01) (Fig. 3A). As Fig.
3B showed, the protein expression of PKM2 notably increased
(P<0.001), which confirmed the success of PKM2 transfection.
Furthermore, the PKM2 protein expression significantly increased
following PKM2 transfection under PPZ treatment (P<0.05)
(Fig. 3C).
PPZ intervention and PKM2 transfection
affects the intracellular expression of PKM2
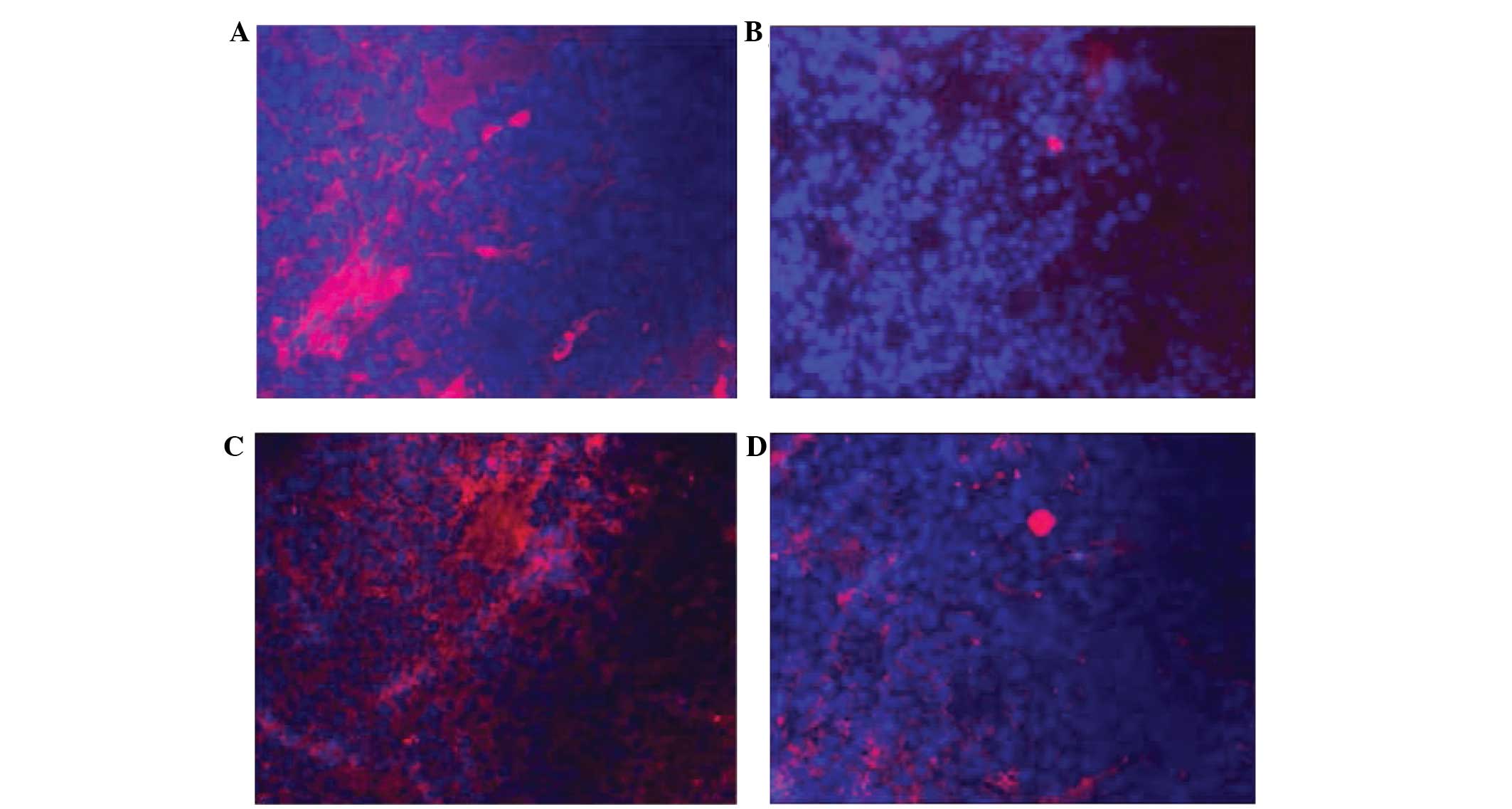
As an inhibitor of PKM2, PPZ treatment for 24 h
caused decreased intracellular expression of PKM2 (Fig. 4A and B). Subsequent to PKM2
transfection, the fluorescent intensity of PKM2 expression
significantly increased compared to the control group (Fig. 4A and C). In addition, the fluorescent
intensity of the PKM2 expression in PKM2+ group with PPZ
treatment was approximately the median (Fig. 4D).
Discussion
Although the Warburg effect was previously proposed,
this phenomenon received sufficient attention after the successful
application of positron emission tomography (PET) in the clinical
diagnosis of tumors (20,21). In recent years, the therapeutic
strategies aimed at the glycolytic pathway have been explored
extensively; and as one of the key enzymes regulating aerobic
glycolysis of tumor cells, PKM2 may be a potential target of cancer
therapy. Christofk et al reported that PKM2 promoted the
rapid growth of cancer cells by diverting glucose metabolites from
energy production to anabolic processes, which was attributed to
phosphotyrosine signaling regulation (8). Based on those results, targeting PKM2
may provide a novel method of reversing chemotherapy resistance to
cancer therapy, by inhibiting tumor growth and promoting increased
cancer cell apoptosis in vitro and in vivo (22). However, the role of PKM2 in tumor
growth remains controversial. One study suggested that PKM2 was
dispensable for tumor maintenance and growth in vivo, and
considered that other metabolic pathways occurred in this
complicated process (23). Therefore,
to clarify the issue, the present study used the gastric
adenocarcinoma SGC-7901 cell line with PPZ treatment and PKM2
transfection to observe cell proliferation and apoptosis, and
simultaneously detect the protein expression of PKM2.
PPZ may suppress proliferation and induce apoptosis
in SGC-7901 cells, which was also confirmed in a previous study
(24). In the present study, it was
observed that the expression of PKM2 decreased concurrently during
this process. It is possible that decreased PKM2 expression impeded
cancer cell growth, or the impeded cell growth reduced the protein
expression of PKM2. In order to resolve this issue, the expression
of PKM2 was increased using transfection, which demonstrated that
cancer cell proliferation was elevated and apoptosis was
restrained. Thus, it was concluded that PKM2 is crucial for cancer
cell growth, to promote proliferation and to inhibit apoptosis.
Based on this conclusion, the inhibition of PKM2 may elicit
anticancer effects, which has been reported to involve various
mechanisms, including the impairment of tumor growth, induction of
apoptotic cell death and increased sensitivity to chemotherapy
(7,11,25,26). In
the present study, increased PKM2 expression by transfection may
partially reverse the effects of PPZ in inhibiting cancer cell
proliferation and inducing apoptosis. The cancer-specific metabolic
transformation associated with therapeutic resistance is a
promising target for cancer therapy (27,28).
Response of the SGC-7901 cells was not observed subsequent to the
inhibition of PKM2 by RNA interference or small-molecule
inhibitors. An additional study may therefore be required.
The mechanisms underlying the effect of PPZ on the
expression of PKM2 remain unclear. Tumor microenvironments are
characterized by acidification and hypoxia. PPIs may inhibit the
expression of vacuolar H+-ATPases (V-ATPases), which are
important for maintaining an acidic extracellular pH (pHe) as
specific proton pumps of the cell (24). A previous study demonstrated that PPZ
treatment significantly inhibited protein expression of V-ATPases,
mechanistic target of rapamycin (mTOR) and hypoxia-inducible factor
1α (HIF-1α), accompanied by increased pHe (29). Another study demonstrated that mTOR
activation simulated hypoxic effects by inducing HIF-1α expression,
which in turn enhanced PKM2 expression through collaboration with
c-Myc-hnRNPs splicing regulators, leading to aerobic glycolysis in
tumor cells (30). Therefore, PPZ may
suppress SGC-7901 cancer cells by downregulating the
V-ATPases/mTOR/HIF-1α/PKM2 signaling pathway. Alternatively, as
benzimidazole compounds, which were found to regulate the stability
of HIF-1α through the Hsp90-Akt pathway (31), PPZ may indirectly reduce the protein
expression of PKM2 by inhibiting HIF-1α expression. Therefore, the
inhibitory effects of PPZ on the PKM2 protein may involve a variety
of factors.
In conclusion, PPZ was demonstrated to effectively
inhibit PKM2 expression in human gastric adenocarcinoma SGC-7901
cells, which was associated with cancer cell proliferation and
apoptosis. The present study has indicated a novel anticancer
mechanism, thus extending the clinical utilization of PPIs as an
anticancer drug by inhibiting the expression of PKM2 in gastric
cancer cells.
Acknowledgements
The present study was supported by grants from
Nanjing Medical Science and Technique Development Foundation (grant
nos. 81101814, 81071816 and 81272742). This study was also
supported by the Fundamental Research Funds for the Central
Universities (grant no. 21414340154).
References
|
1
|
Warburg O: On the origin of cancer cells.
Science. 123:309–314. 1956. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Simonnet H, Alazard N, Pfeiffer K, Gallou
C, Béroud C, Demont J, Bouvier R, Schägger H and Godinot C: Low
mitochondrial respiratory chain content correlates with tumor
aggressiveness in renal cell carcinoma. Carcinogenesis. 23:759–768.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jiang K, He B, Lai L, Chen Q, Liu Y, Guo Q
and Wang Q: Cyclosporine A inhibits breast cancer cell growth by
downregulating the expression of pyruvate kinase subtype M2. Int J
Mol Med. 30:302–308. 2012.PubMed/NCBI
|
|
4
|
Furtado CM, Marcondes MC, Sola-Penna M, de
Souza ML and Zancan P: Clotrimazole preferentially inhibits human
breast cancer cell proliferation, viability and glycolysis. PLoS
One. 7:e304622012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kosugi M, Ahmad R, Alam M, Uchida Y and
Kufe D: MUC1-C oncoprotein regulates glycolysis and pyruvate kinase
M2 activity in cancer cells. PLoS One. 6:e282342011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jurica MS, Mesecar A, Heath PJ, Shi W,
Nowak T and Stoddard BL: The allosteric regulation of pyruvate
kinase by fructose-1,6-bisphosphate. Structure. 6:195–210. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Christofk HR, Van der Heiden MG, Harris
MH, Ramanathan A, Gerszten RE, Wei R, Fleming MD, Schreiber SL and
Cantley LC: The M2 splice isoform of pyruvate kinase is important
for cancer metabolism and tumour growth. Nature. 452:230–233. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Christofk HR, Van der Heiden MG, Wu N,
Asara JM and Cantley LC: Pyruvate kinase M2 is a
phosphotyrosine-binding protein. Nature. 452:181–186. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang W, Xia Y, Ji H, Zheng Y, Liang J,
Huang W, Gao X, Aldape K and Lu Z: Nuclear PKM2 regulates β-catenin
transactivation upon EGFR activation. Nature. 480:118–122. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Anastasiou D, Poulogiannis G, Asara JM,
Boxer MB, Jiang JK, Shen M, Bellinger G, Sasaki AT, Locasale JW,
Auld DS, et al: Inhibition of pyruvate kinase M2 by reactive oxygen
species contributes to cellular antioxidant responses. Science.
334:1278–1283. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Goldberg MS and Sharp PA: Pyruvate kinase
M2-specific siRNA induces apoptosis and tumor regression. J Exp
Med. 209:217–224. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang LY, Liu YP, Chen LG, Chen YL, Tan L,
Liu JJ, Jazag A, Ren JL and Guleng B: Pyruvate kinase M2 plays a
dual role on regulation of the EGF/EGFR signaling via
E-cadherin-dependent manner in gastric cancer cells. PLoS One.
8:e675422013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lim JY, Yoon SO, Seol SY, Hong SW, Kim JW,
Choi SH and Cho JY: Overexpression of the M2 isoform of pyruvate
kinase is an adverse prognostic factor for signet ring cell gastric
cancer. World J Gastroenterol. 18:4037–4043. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kwon OH, Kang TW, Kim JH, Kim M, Noh SM,
Song KS, Yoo HS, Kim WH, Xie Z, Pocalyko D, et al: Pyruvate kinase
M2 promotes the growth of gastric cancer cells via regulation of
Bcl-xL expression at transcriptional level. Biochem Biophys Res
Commun. 423:38–44. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Blume H, Donath F, Warnke A and Schug BS:
Pharmacokinetic drug interaction profiles of proton pump
inhibitors. Drug Saf. 29:769–784. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shen Y, Wu Y, Chen M, Shen W, Huang S,
Zhang L and Zou X: Effects of pantoprazole as a HIF-1α inhibitor on
human gastric adenocarcinoma sgc-7901 cells. Neoplasma. 59:142–149.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tao T, Li G, Dong Q, Liu D, Liu C, Han D,
Huang Y, Chen S, Xu B and Chen M: Loss of SNAIL inhibits cellular
growth and metabolism through the miR-128-mediated
RPS6KB1/HIF-1α/PKM2 signaling pathway in prostate cancer cells.
Tumour Biol. 35:8543–8550. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yeo M, Kim DK, Kim YB, Oh TY, Lee JE, Cho
SW, Kim HC and Hahm KB: Selective induction of apoptosis with
proton pump inhibitor in gastric cancer cells. Clin Cancer Res.
10:8687–8696. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Czernin J and Phelps ME: Positron emission
tomography scanning: Current and future applications. Annu Rev Med.
53:89–112. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gambhir SS: Molecular imaging of cancer
with positron emission tomography. Nat Rev Cancer. 2:683–693. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shi HS, Li D, Zhang J, Wang YS, Yang L,
Zhang HL, Wang XH, Mu B, Wang W, Ma Y, et al: Silencing of pkm2
increases the efficacy of docetaxel in human lung cancer xenografts
in mice. Cancer Sci. 101:1447–1453. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cortés-Cros M, Hemmerlin C, Ferretti S,
Zhang J, Gounarides JS, Yin H, Muller A, Haberkorn A, Chene P,
Sellers WR and Hofmann F: M2 isoform of pyruvate kinase is
dispensable for tumor maintenance and growth. Proc Natl Acad Sci
USA. 110:489–494. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen M, Huang SL, Zhang XQ, Zhang B, Zhu
H, Yang VW and Zou XP: Reversal effects of pantoprazole on
multidrug resistance in human gastric adenocarcinoma cells by
down-regulating the V-ATPases/mTOR/HIF-1α/P-gp and MRP1 signaling
pathway in vitro and in vivo. J Cell Biochem.
113:2474–2487. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Spoden GA, Rostek U, Lechner S,
Mitterberger M, Mazurek S and Zwerschke W: Pyruvate kinase
isoenzyme M2 is a glycolytic sensor differentially regulating cell
proliferation, cell size and apoptotic cell death dependent on
glucose supply. Exp Cell Res. 315:2765–2774. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Guo W, Zhang Y, Chen T, Wang Y, Xue J,
Zhang Y, Xiao W, Mo X and Lu Y: Efficacy of RNAi targeting of
pyruvate kinase M2 combined with cisplatin in a lung cancer model.
J Cancer Res Clin Oncol. 137:65–72. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Spoden GA, Mazurek S, Morandell D, Bacher
N, Ausserlechner MJ, Jansen-Dürr P, Eigenbrodt E and Zwerschke W:
Isotype-specific inhibitors of the glycolytic key regulator
pyruvate kinase subtype M2 moderately decelerate tumor cell
proliferation. Int J Cancer. 123:312–321. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hamanaka RB and Chandel NS: Targeting
glucose metabolism for cancer therapy. J Exp Med. 209:211–215.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tamada M, Suematsu M and Saya H: Pyruvate
kinase M2: Multiple faces for conferring benefits on cancer cells.
Clin Cancer Res. 18:5554–5561. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sun Q, Chen X, Ma J, Peng H, Wang F, Zha
X, Wang Y, Jing Y, Yang H, Chen R, et al: Mammalian target of
rapamycin up-regulation of pyruvate kinase isoenzyme type M2 is
critical for aerobic glycolysis and tumor growth. Proc Natl Acad
Sci USA. 108:4129–4134. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Won MS, Im N, Park S, Boovanahalli SK, Jin
Y, Jin X, Chung KS, Kang M, Lee K, Park SK, et al: A novel
benzimidazole analogue inhibits the hypoxia-inducible factor
(HIF)-1 pathway. Biochem Biophys Res Commun. 385:16–21. 2009.
View Article : Google Scholar : PubMed/NCBI
|