Introduction
Colorectal carcinoma (CRC) is one of the most common
types of solid malignancy and a main cause of cancer-related
mortality globally. Colorectal carcinogenesis is a complex
multistep process that involves progressively disrupting intestinal
epithelial cell proliferation, differentiation, apoptosis and
survival mechanisms (1,2). Mitogen-activated protein kinases
(MAPKs), members of a large family of serine-threonine kinases,
form major cell proliferation signaling pathways from the cell
surface to the nucleus (3), and can
be activated in response to a wide range of extracellular stimuli
(4). The first member of this
family to be characterized was extracellular signal-regulated
kinase (ERK). The ERK pathway is important for cell survival and
proliferation, and a number of key growth factors and
proto-oncogenes act via this cascade, transducing the signals to
promote cell growth and differentiation (5). For example, activated Ras activates
Raf-1, which in turn phosphorylates and activates MAPK/ERK kinase
(MEK), resulting in activation of ERK, the only known substrate of
MEK. Analysis of established cell lines indicates that constitutive
activation of MAPK is induced in tumors in a tissue-specific
manner; for example, tumor cells derived from CRC tissues exhibit a
particularly high frequency of ERK activation (6). Furthermore, increasing evidence
indicates that the ERK pathway is important for regulating
apoptosis and cell proliferation, and that its activation is
involved in the tumorigenesis, progression and oncogenic behavior
of CRC (7–12).
Raf-1 kinase inhibitory protein (RKIP), a member of
the phosphatidylethanolamine binding protein family, was initially
identified as an endogenous inhibitor of the ERK pathway. By
binding to Raf-1 or MEK, RKIP interferes with MEK activation by
Raf-1, thus inhibiting ERK activation (13). Our previous studies demonstrated
that RKIP downregulation significantly contributes to the
invasiveness and metastatic potential of a number of different
types of human carcinoma, including CRC (14–19);
for example, it was identified that RKIP expression was
significantly reduced in metastatic tumor tissues compared with
corresponding primary tumor and non-neoplastic tissues (15,17,18,20).
In addition, a reduction in RKIP expression levels was associated
with lymphovascular invasion, as well as nodal and distant
metastases, increased local recurrence, advanced tumor stage and
decreased survival rate (14–16,18).
Furthermore, RKIP downregulation appeared to correlate with
aggressive oncogenic behavior. Thus, the data indicated that RKIP
may act as a suppressor of metastasis in CRC.
However, our previous studies determined conflicting
results regarding the association between RKIP expression and
metastasis in specific CRC patients, for example, hepatic
colorectal metastasis (HCM) was histopathologically diagnosed in
CRC patients whose tumors exhibited positive RKIP immunoreactivity
(19). This finding indicated that
metastatic properties are preserved in a certain group of tumor
cells despite the presence of the metastatic suppressor RKIP.
Furthermore, previous studies demonstrated that ERK can be
phosphorylated regardless of RKIP status (21,22).
Based on these data, we hypothesized that the ERK pathway may be
activated in RKIP-expressing HCM tissues, and that the activation
of this pathway may be involved in the invasive process of HCM
cells.
Therefore, the present study investigated whether
phosphorylated ERK (pERK) is detectable in RKIP-expressing HCM
tissues. In addition, the association between pERK expression and
various clinicopathological characteristics of HCM patients was
evaluated to assess its clinical value.
Patients and methods
Patient and tissue specimens
Human HCM tissue samples were obtained from 68
consecutive patients (46 male and 22 female; median age, 61 years;
mean age, 60.1 years) who underwent surgery at the Kyung Hee
University Hospital (Seoul, Korea) between January 2005 and
December 2012. All of the patients met the following criteria for
hepatic resection with intent to cure: i) No signs of extrahepatic
metastases in pre-operative imaging, including chest X-ray,
abdominal ultrasonography and abdominopelvic computed tomography;
ii) HCM that allowed adequately sized, well-vascularized hepatic
remnants to remain following hepatic resection; and iii) medical
fitness for major hepatic resection. Only patients whose metastases
were resectable on presentation were included in the present study.
The resected lesions were stained with hematoxylin and eosin and
reviewed by two independent pathologists, who selected the most
representative slide from each case for subsequent
immunohistochemical staining. The selected tissues were fixed in
10% neutral buffered formalin for 24–72 h at room temperature for
paraffin embedding prior to analysis. Furthermore, the following
clinicopathological data was assessed in each case: Patient age and
gender, size, number and distribution of HCMs, HCM recurrence and
survival data obtained from follow-up after hepatic resection.
Informed consent was obtained from all participants. The study was
approved by the institutional review board of the Republic of Korea
Air Force Aerospace Medical Center (Cheongju, Korea).
Immunohistochemistry
RKIP and pERK protein expression was assessed by
immunohistochemistry using the Bond Polymer Intense Detection
System (Leica Biosystems, Seoul, Korea), according to the
manufacturer’s instructions. Briefly, 4-μm sections of
formalin-fixed, paraffin-embedded tissue were deparaffinized with
Bond Dewax solution (Leica Biosystems) and an antigen retrieval
procedure was performed using Bond Epitope Retrieval solution 1
(cat. no. AR9961; Leica Biosystems) for 30 min at 100°C. Endogenous
peroxidases were blocked by incubation with hydrogen peroxide for 5
min and the sections were incubated for 15 min at ambient
temperature with a 1:200 dilution of rabbit polyclonal anti-RKIP
(cat. no. sc-32623; Santa Cruz Biotechnology, Inc., Santa Cruz, CA,
USA) or rabbit monoclonal anti-pERK (phospho-p44/42 MAPK
[Thr202/Tyr204; clone, 20G11]; cat. no. 4376S; Cell Signaling
Technology, Inc., Beverly, MA, USA). Staining was performed using a
biotin-free polymeric horseradish peroxidase-linker antibody
conjugate system with the Bond Max automatic slide stainer (Leica
Biosystems), and the slides were visualized with
3,3′-diaminobenzidine (DAB) solution [1 mm DAB, 50 mm Tris-HCl
buffer (pH 7.6) and 0.006% H2O2]. In
addition, the nuclei were counterstained with hematoxylin and the
slides were dehydrated through a series of graded alcohols (70, 90
and 100%), cleared in xylene (Sigma Aldrich, St. Louis, MO, USA)
and sealed with coverslips. Positive control samples consisted of
healthy colonic mucosa for RKIP, and prostate carcinoma and
malignant melanoma for pERK (23,24).
The negative control was prepared by substituting non-immune serum
for primary antibody and resulted in no detectable staining.
Evaluation of immunohistochemical
staining
Two experienced independent pathologists, who were
blinded to the clinicopathological data and outcomes of the
patients, examined the immunohistochemical expression of RKIP and
pERK in the stained sections. Immunoreactivity for RKIP and pERK
was analyzed using previously described semi-quantitative scoring
methods (14–20), and the scores of the two
pathologists were compared, with discrepancies resolved by
re-examination by the two pathologists to achieve a consensus
score. The final score was the sum of the scores for the percentage
of positive tumor cells (0, 0%; 1, ≤25%; 2, 25–50%; and 3, >50%)
and the staining intensity (0, absent; 1, faint; 2, moderate; 3,
strong). Additionally, expression scoring was follows: 0–2,
negative expression; 3–4, weak expression; and 5–6, positive
expression. All the immunostaining scores were determined at the
center, intermediate zone and invasive front of the tumor, as well
as at subcellular locations.
Statistical analysis
Fisher’s exact test was performed to determine
whether pERK expression was associated with each HCM
clinicopathological characteristic, and a survival analysis was
used to examine the prognostic significance of pERK expression in
the HCM patients by producing survival curves using the
Kaplan-Meier method and applying a log-rank test to analyze any
statistical differences. All statistical analyses were performed
using SPSS software (version 15.0; SPSS Inc., Chicago, IL, USA) and
P<0.05 was used to indicate a statistically significant
difference.
Results
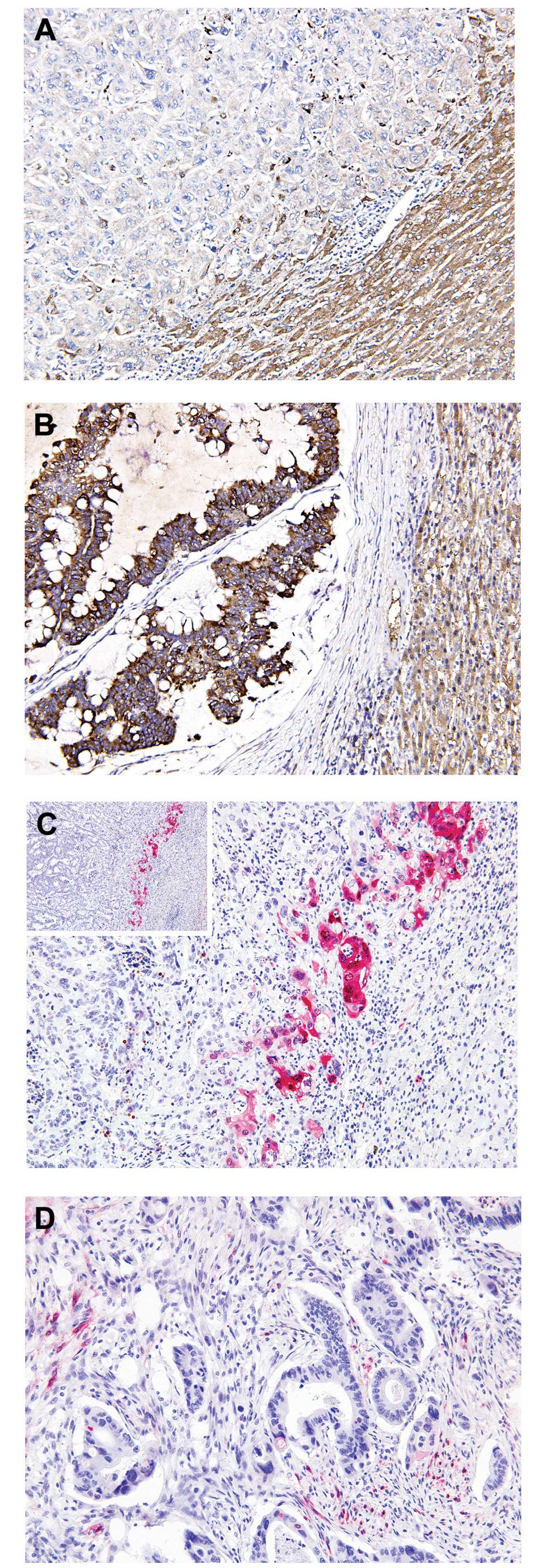
Overall, RKIP immunoreactivity appeared to be
predominantly cytoplasmic and homogeneous throughout the tumor,
with protein expression levels determined to be negative in 36.8%
(25/68; Fig. 1A), weak in 30.8%
(21/68) and positive in 32.4% (22/68; Fig. 1B) of samples. Of the 22
RKIP-expressing cases, seven (31.8%) exhibited intense nuclear and
moderate cytoplasmic pERK immunoreactivity (Fig. 1C), while the remaining 15 cases
(68.2%) exhibited no pERK immunoreactivity (Fig. 1D). In contrast to RKIP expression,
pERK immunoreactivity was heterogeneous and strongly positive pERK
protein expression was exclusively detected at the invasive tumor
front, whereas the tumor center and intermediate zones did not
display pERK immunoreactivity. Furthermore, only cells that faced
the hepatic parenchyma expressed pERK, forming a band-like pattern
(Fig. 1C). This unique localization
of pERK was accompanied by loss of the epithelial phenotype, and
characterized by the detachment of small isolated clusters of tumor
cells and a dedifferentiated morphology at the invasive front. By
contrast, cells in the center and intermediate zones of the tumor
exhibited a typical epithelial growth pattern characterized by
adherent polarized cells forming clear trabecular or tubular
structures. In addition, pERK staining exhibited strong staining
intensity in the nucleus, consistent with ERK activity being
highest in this subcellular location.
The association between pERK expression at the
invasive front and the clinicopathological characteristics of HCM
is indicated in Table I. It was
observed that pERK expression at the invasive front was
significantly associated with recurrent HCM (P<0.001); all
patients with pERK-positive tumors developed recurrent HCM, while
in patients who did not develop recurrent HCM, pERK expression was
absent. The prediction of recurrent HCM was associated with pERK
expression at the invasive front, with 87.5% sensitivity, 100%
specificity, a 100% positive predictive value and a 93.3% negative
predictive value.
 | Table IAssociation between pERK expression at
the invasive front and clinicopathological characteristics of HCM
patients. |
Table I
Association between pERK expression at
the invasive front and clinicopathological characteristics of HCM
patients.
| pERK status | |
|---|
|
| |
|---|
| Characteristic | Positive (n=7) | Negative (n=15) | P-value |
|---|
| Age, n (%) |
| ≥61 years | 4 (57.1) | 6 (40.0) | 0.652 |
| <61 years | 3 (42.9) | 9 (60.0) | |
| Gender, n (%) |
| Male | 5 (71.4) | 10 (66.7) | 1.000 |
| Female | 2 (28.6) | 5 (33.3) | |
| HCM, n (%) |
| Single | 4 (57.1) | 11 (73.3) | 0.630 |
| Multiple | 3 (42.9) | 4 (26.7) | |
| HCM size, n
(%) |
| ≥2.5 cm | 3 (42.9) | 8 (53.3) | 1.000 |
| <2.5 cm | 4 (57.1) | 7 (46.7) | |
| HCM distribution, n
(%) |
| Unilobar | 6 (85.7) | 12 (80.0) | 1.000 |
| Bilobar | 1 (14.3) | 3 (20.0) | |
| HCM recurrence, n
(%) |
| Present | 7 (100.0) | 1 (6.7) | <0.001a |
| Absent | 0 (0.0) | 14 (93.3) | |
| Median survival
(range), monthsb | 26 (1–62) | 28 (6–85) | 0.853 |
Discussion
Histopathological studies have demonstrated that
invasiveness, characterized by the loss of epithelial
differentiation and the acquisition of a mesenchymal phenotype, is
predominantly observed at the periphery of CRC tumors (25–27),
with cells at the invasive front considered to exhibit a higher
malignant potential than the inner areas of the same tumor.
Previous studies utilized CRC cell lines and surgical specimens to
demonstrate that the ERK pathway is critical for the invasive and
proliferative properties of these tumors (6,25,28,29).
Furthermore, pERK was expressed at higher levels in more advanced
stages of CRC, demonstrating the importance of this molecule in CRC
progression, invasion and metastasis (2). In the present study, the selective
localization of pERK was identified at the invasive front of a
subpopulation of metastasized CRC cells, but not in the inner, more
differentiated areas of the tumor, and pERK expression was
accompanied by the loss of the epithelial phenotype. This pERK
expression pattern is consistent with the results of the
aforementioned studies and possibly reflects the high biological
aggressiveness of tumor cells at the invasive edge. Activation of
the ERK pathway initiates transcription, which leads to the
expression of matrix metalloproteinase and myosin light-chain
kinase, inducing degradation of the basement membrane and promoting
cell migration and invasion (30–32).
Additionally, recent studies into the functional role of pERK in
carcinomas revealed that pERK upregulation augmented cell motility
on extracellular matrix components, increased Matrigel invasion and
promoted the growth of the tumor (33–35).
To the best of our knowledge, the distinct distribution of pERK at
the invasive front of HCM has yet to be reported, and the findings
of the present study indicate that pERK is significant in the HCM
invasive process in vivo, which supports the previous
finding that activation of the ERK pathway is necessary for the
induction of epithelial cell dedifferentiation (36).
Furthermore, the current study detected intense pERK
immunoreactivity in RKIP-expressing metastatic tumors. Although
oncogenic pathways other than the ERK cascade may contribute to CRC
cell dissemination in RKIP-expressing tumors, the possibility that
the ERK pathway may be involved in the metastatic process
regardless of RKIP status must be considered, thus, the present
study proposes a number of possible explanations for this result.
Firstly, the ERK cascade is not a simple linear pathway, but has
numerous positive and negative regulatory components, which
function at all levels of the cascade (37). For example, a previous study
demonstrated that RKIP regulates Raf-1, but not B-Raf, indicating
that B-Raf is able to phosphorylate and activate MEK and ERK
independent of RKIP (22). Papin
et al (21) reported that
B-Raf displayed higher MEK kinase activity compared with Raf-1.
Based on these data, the present study proposes that BRAF
mutations may be involved in ERK activation in RKIP-expressing
tumors. Secondly, in addition to the predominant MEK activators
Raf-1 and B-Raf, MEK kinase-1 and A-Raf are able to phosphorylate
MEK and thus activate the ERK pathway, although the biochemical
potency of A-Raf is much weaker than that of Raf-1 or B-Raf
(38). Finally, MEK is capable of
autophosphorylation, resulting in an increase in MEK kinase
activity (39). Thus, although the
observation of pERK expression at the invasive front of
RKIP-expressing HCMs indicates that the activation of the ERK
pathway contributes to the invasive process in RKIP-expressing
metastatic tumor cells, additional studies are required to clarify
or disprove the findings of the present study.
The treatment of primary CRC with surgical
resection, combined with chemotherapy or radiation therapy in
certain cases, is curative in a number of patients, however, almost
half will develop HCM during the disease course (40). Furthermore, even when hepatic
resection is performed with curative intent, the majority of
patients experience tumor recurrence (41); one-third of which develop recurrent
metastases isolated to the liver. Repeat hepatic resection for
recurrent HCM appears to be warranted in carefully selected
patients due to the reasonable survival expectations, similar to
those of patients undergoing single hepatic resection (42). In this regard, identifying markers
based on biological determinants may facilitate the improvement and
earlier stratification of patients according to recurrence risk,
and aid in selecting patients who may benefit from repeated hepatic
resection. The present study demonstrated that pERK expression was
significantly associated with recurrent HCM, and high sensitivity,
specificity and predictive values indicated that pERK expression at
the invasive front of HCM may be used as a novel biomarker for the
prediction of recurrent HCM.
In conclusion, the present study identified the
unique, heterogeneous expression of pERK in HCM and its association
with metastatic recurrence. The selective upregulation of pERK at
the invasive tumor front indicates that activation of the ERK
pathway is involved in the invasive process of HCM cells. In
addition, pERK expression at the invasive front of HCM was strongly
associated with HCM recurrence, with high sensitivity, specificity
and predictive values. These findings indicate that pERK expression
may be used as a novel predictive biomarker of metastatic
recurrence and an effective target for therapeutic strategies
against RKIP-expressing HCM.
References
|
1
|
Calvert PM and Frucht H: The genetics of
colorectal cancer. Ann Intern Med. 137:603–612. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fang JY and Richardson BC: The MAPK
signalling pathways and colorectal cancer. Lancet Oncol. 6:322–327.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dong C, Davis RJ and Flavell RA: MAP
kinases in the immune response. Annu Rev Immunol. 20:55–72. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lewis TS, Shapiro PS and Ahn NG: Signal
transduction through MAP kinase cascades. Adv Cancer Res.
74:49–139. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Troppmair J, Bruder JT, Munoz H, et al:
Mitogen-activated protein kinase/extracellular signal-regulated
protein kinase activation by oncogenes, serum, and
12-O-tetradecanoylphorbol-13-acetate requires Raf and is necessary
for transformation. J Biol Chem. 269:7030–7035. 1994.PubMed/NCBI
|
|
6
|
Hoshino R, Chatani Y, Yamori T, et al:
Constitutive activation of the 41-/43-kDa mitogen-activated protein
kinase signaling pathway in human tumors. Oncogene. 18:813–822.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Remacle-Bonnet MM, Garrouste FL, Heller S,
et al: Insulin-like growth factor-I protects colon cancer cells
from death factor-induced apoptosis by potentiating tumor necrosis
factor alpha-induced mitogen-activated protein kinase and nuclear
factor kappaB signaling pathways. Cancer Res. 60:2007–2017.
2000.PubMed/NCBI
|
|
8
|
Wang X, Wang Q, Hu W and Evers BM:
Regulation of phorbol ester-mediated TRAF1 induction in human colon
cancer cells through a PKC/RAF/ERK/NF-kappaB-dependent pathway.
Oncogene. 23:1885–1895. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ukegawa JI, Takeuchi Y, Kusayanagi S and
Mitamura K: Growth-promoting effect of muscarinic acetylcholine
receptors in colon cancer cells. J Cancer Res Clin Oncol.
129:272–278. 2003.PubMed/NCBI
|
|
10
|
Vial E and Marshall CJ: Elevated ERK-MAP
kinase activity protects the FOS family member FRA-1 against
proteasomal degradation in colon carcinoma cells. J Cell Sci.
116:4957–4963. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Im E and Martinez JD: Ursodeoxycholic acid
(UDCA) can inhibit deoxycholic acid (DCA)-induced apoptosis via
modulation of EGFR/Raf-1/ERK signaling in human colon cancer cells.
J Nutr. 134:483–486. 2004.PubMed/NCBI
|
|
12
|
Sebolt-Leopold JS, Dudley DT, Herrera R,
et al: Blockade of the MAP kinase pathway suppresses growth of
colon tumors in vivo. Nat Med. 5:810–816. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yeung K, Seitz T, Li S, et al: Suppression
of Raf-1 kinase activity and MAP kinase signalling by RKIP. Nature.
401:173–177. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim HS, Kim GY, Lim SJ and Kim YW:
Expression of Raf-1 kinase inhibitory protein in extrahepatic bile
duct carcinoma. Korean J Pathol. 44:234–242. 2010. View Article : Google Scholar
|
|
15
|
Kim HS, Kim GY, Lim SJ and Kim YW: Loss of
Raf-1 kinase inhibitory protein in pancreatic ductal
adenocarcinoma. Pathology. 42:655–660. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim HS, Kim GY, Lim SJ and Kim YW: Raf-1
kinase inhibitory protein expression in thyroid carcinomas. Endocr
Pathol. 21:253–257. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kim HS, Kim GY, Lim SJ, Park YK and Kim
YW: Reduced expression of Raf-1 kinase inhibitory protein is a
significant prognostic marker in patients with gallbladder
carcinoma. Hum Pathol. 41:1609–1616. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim HS, Lee SH, Won KY, et al: Expression
of Raf-1 kinase inhibitory protein in carcinoma of the ampulla of
Vater. Virchows Arch. 460:61–68. 2012. View Article : Google Scholar
|
|
19
|
Kim HS, Park SJ, Lee KY, Park YK and Kim
YW: Reduced Raf-1 kinase inhibitor protein expression predicts less
favorable outcomes in patients with hepatic colorectal metastasis.
Oncol Rep. 28:161–171. 2012.PubMed/NCBI
|
|
20
|
Kim HS, Won KY, Kim GY, et al: Reduced
expression of Raf-1 kinase inhibitory protein predicts regional
lymph node metastasis and shorter survival in esophageal squamous
cell carcinoma. Pathol Res Pract. 208:292–299. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Papin C, Denouel-Galy A, Laugier D,
Calothy G and Eychene A: Modulation of kinase activity and
oncogenic properties by alternative splicing reveals a novel
regulatory mechanism for B-Raf. J Biol Chem. 273:24939–24947. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Trakul N, Menard RE, Schade GR, Qian Z and
Rosner MR: Raf kinase inhibitory protein regulates Raf-1 but not
B-Raf kinase activation. J Biol Chem. 280:24931–24940. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Royuela M, Arenas MI, Bethencourt FR, et
al: Regulation of proliferation/apoptosis equilibrium by
mitogen-activated protein kinases in normal, hyperplastic, and
carcinomatous human prostate. Hum Pathol. 33:299–306. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhuang L, Lee CS, Scolyer RA, et al:
Activation of the extracellular signal regulated kinase (ERK)
pathway in human melanoma. J Clin Pathol. 58:1163–1169. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Corn BW, Kovner F, Bek S, et al: ERK
signaling in colorectal cancer: a preliminary report on the
expression of phosphorylated ERK and the effects of radiation
therapy. Am J Clin Oncol. 31:255–258. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Brabletz T, Jung A, Reu S, et al: Variable
beta-catenin expression in colorectal cancers indicates tumor
progression driven by the tumor environment. Proc Natl Acad Sci
USA. 98:10356–10361. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bates RC and Mercurio AM: The
epithelial-mesenchymal transition (EMT) and colorectal cancer
progression. Cancer Biol Ther. 4:365–370. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhu F, Zykova TA, Kang BS, et al:
Bidirectional signals transduced by TOPK-ERK interaction increase
tumorigenesis of HCT116 colorectal cancer cells. Gastroenterology.
133:219–231. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Levidou G, Saetta AA, Gigelou F, et al:
ERK/pERK expression and B-raf mutations in colon adenocarcinomas:
correlation with clinicopathological characteristics. World J Surg
Oncol. 10:472012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Klemke RL, Cai S, Giannini AL, et al:
Regulation of cell motility by mitogen-activated protein kinase. J
Cell Biol. 137:481–492. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
McCawley LJ, Li S, Wattenberg EV and
Hudson LG: Sustained activation of the mitogen-activated protein
kinase pathway. A mechanism underlying receptor tyrosine kinase
specificity for matrix metalloproteinase-9 induction and cell
migration. J Biol Chem. 274:4347–4353. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kajanne R, Miettinen P, Mehlem A, et al:
EGF-R regulates MMP function in fibroblasts through MAPK and AP-1
pathways. J Cell Physiol. 212:489–497. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Vial E, Sahai E and Marshall CJ: ERK-MAPK
signaling coordinately regulates activity of Rac1 and RhoA for
tumor cell motility. Cancer Cell. 4:67–79. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Viala E and Pouysségur J: Regulation of
tumor cell motility by ERK mitogen-activated protein kinases. Ann
NY Acad Sci. 1030:208–218. 2004. View Article : Google Scholar
|
|
35
|
Roberts PJ and Der CJ: Targeting the
Raf-MEK-ERK mitogen-activated protein kinase cascade for the
treatment of cancer. Oncogene. 26:3291–3310. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xie L, Law BK, Chytil AM, et al:
Activation of the Erk pathway is required for TGF-beta1-induced EMT
in vitro. Neoplasia. 6:603–610. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dhillon AS, Hagan S, Rath O and Kolch W:
MAP kinase signalling pathways in cancer. Oncogene. 26:3279–3290.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wu X, Noh SJ, Zhou G, Dixon JE and Guan
KL: Selective activation of MEK1 but not MEK2 by A-Raf from
epidermal growth factor-stimulated Hela cells. J Biol Chem.
271:3265–3271. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gardner AM, Vaillancourt RR, Lange-Carter
CA and Johnson GL: MEK-1 phosphorylation by MEK kinase, Raf, and
mitogen-activated protein kinase: analysis of phosphopeptides and
regulation of activity. Mol Biol Cell. 5:193–201. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ballantyne GH and Quin J: Surgical
treatment of liver metastases in patients with colorectal cancer.
Cancer. 71(Suppl): 4252–4266. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Nordlinger B, Vaillant JC, Guiguet M, et
al: Survival benefit of repeat liver resections for recurrent
colorectal metastases: 143 cases. Association Francaise de
Chirurgie. J Clin Oncol. 12:1491–1496. 1994.PubMed/NCBI
|
|
42
|
Petrowsky H, Gonen M, Jarnagin W, et al:
Second liver resections are safe and effective treatment for
recurrent hepatic metastases from colorectal cancer: a
bi-institutional analysis. Ann Surg. 235:863–871. 2002. View Article : Google Scholar : PubMed/NCBI
|