Introduction
Whilst patients with a well-confined primary tumor
may be treated by surgical removal of the tumor, metastatic
diseases are frequently incurable, and are the primary cause of
cancer morbidity and mortality (1,2). It is
estimated that metastasis is responsible for ≤90% of mortalities
from solid cancers (1,2). Colorectal cancer is one of the major
causes of cancer-related mortality worldwide, despite various
combinations of current treatments, including surgery,
chemotherapy, and/or radiation. This failure is primarily due to
the resistance to existing anticancer therapies for metastatic
tumor cells, which may be originate from cancer stem cells
(3). Therefore, it is necessary to
understand the characteristics of metastatic cancers in order to
develop new therapeutic approaches.
Cancer stem cells (CSCs) are thought to be
responsible for cancer initiation, progression, metastasis,
recurrence and drug resistance (4).
As a source of migratory/invasive cells in primary tumors, CSCs
appear to be essential for the formation of distant metastases
(3). As CSCs are naturally
resistant to a variety of therapeutic insults (5), metastatic cells may also be resistant
to therapy. Indeed, resistance to apoptosis appears to be an
important component of metastasis. It has been demonstrated that
tumor cells with resistance to apoptosis are more likely to
successfully establish metastases (6,7), and
the resistance to apoptosis is increased during metastatic
dissemination of colon cancer (8).
Therefore, highly metastatic tumors may comprise a greater
proportion of CSCs compared with primary tumors.
Cancer immunotherapies have been considered as a
potential approach to eliminate CSCs, due to the reported role of
immune effector cells, including CD8+ T cells and natural killer
(NK) cells, in the prevention of tumor formation (9,10).
Little has been established with regard to the susceptibility of
CSCs to NK cell-mediated lysis. NK cell-mediated target cell lysis
and apoptosis is a cell-contact-dependent process, which acts via
the release of cytotoxic granules containing perforin and
granzymes, and death receptor (DR) pathways, including TNF-related
apoptosis-inducing ligand (TRAIL) receptors or Fas (11,12);
this may be regulated by a balance between activatory and
inhibitory signals, and cell death signals (13,14).
In the present study, the clonogenicity in soft
agar, and susceptibility to NK cells of clonogenic colon cancer
cells was compared between the colorectal cancer cell line KM12C
and its highly metastatic sublines, KM12SM and KM12L4A.
Materials and methods
Cell lines and reagents
The poorly metastatic KM12C cells (University of
Texas MD Anderson Cancer Center, Houston, TX, USA), which were
established from a primary colorectal carcinoma classified as
Dukes, and the highly metastatic KM12SM and KM12L4A cells (both
obtained from University of Texas MD Anderson Cancer Center), which
were derived from KM12C cells (15), were maintained at 37°C in humidified
atmosphere of 5% CO2, in Dulbecco’s modified Eagle’s
medium (Gibco, Grand Island, NY, USA) supplemented with 10% fetal
bovine serum (FBS), 2mM L-glutamine, 100 units/ml penicillin and
100 μg/ml streptomycin. NK-92 cells were maintained in α-minimum
essential medium (Lonza, Walkersville, MD, USA) containing 12.5%
FBS, 12.5% horse serum (Gibco), 2 mM L-glutamine, 0.1 mM
2-mercaptoethanol, and 400 U/ml recombinant human interleukin-2
(Novartis, Surrey, UK). Anti-major histocompatibility complex (MHC)
class I polypeptide-related sequence A/B (MICA/B; cat. no. MAB1300)
and anti-UL16 binding protein (ULBP)1/2/3 (cat. no’s. MAB1380,
MAB1298 and MAB1517) antibodies were purchased from R&D
Systems, Inc. (Shanghai, China). Allophycocyanin (APC)-conjugated
anti-mouse IgG, and fluorescein isothiocyanate (FITC)-conjugated
anti-CD44 and anti-epithelial cell adhesion molecule(EpCAM)-PerCP
Cy5.5 antibodies were purchased from BD Biosciences (Franklin
Lakes, NJ, USA). Phycoerythrin (PE)-conjugated anti-CD133 antibody
was purchased from Miltenyi Biotec Inc. (Cambridge, MA, USA) and
mouse monoclonal antibodies against TRAIL-R1 and R2 were purchased
from Enzo Life Sciences (Farmingdale, NY, USA).
Flow cytometric analysis
Flow cytometry was conducted to evaluate the surface
expression levels of CD133, CD44, EpCAM and NKG2D ligands. Cells
were harvested, washed twice with cold phosphate buffered saline,
and subsequently incubated with the appropriate antibodies,
according to the manufacturer’s instructions, for 30 min at 4°C in
the dark: For the evaluation of CSC marker expression, cells were
stained with PE-conjugated anti-CD133, FITC-conjugated anti-CD44
and PerCP-conjugated EpCAM; to assess the expression of NKG2D
ligands and TRAIL receptors, cells were stained with anti-MICA/B,
ULBP1, ULBP2 and ULBP3 primary antibodies, and anti-DR4 and DR5
primary antibodies, followed by incubation with APC-conjugated
secondary antibody. Flow cytometric analysis was performed using
FACSCalibur or FACSCanto II (BD Biosciences), and analyzed with
CellQuest version 3.1 software (BD Biosciences) or BD FACSDiva
version 2.1.6 software (BD Biosciences), respectively. Normalized
mean fluorescence intensity (MFI) was calculated by subtracting the
MFI of the isotype control from the MFI of the specific antibody
(16,17).
RNA isolation and reverse transcription
quantitative polymerase chain reaction (PCR)
Total RNA was isolated using the RNeasy mini kit
(Qiagen, Valencia, CA, USA) and reverse transcribed using M-MLV
Reverse Transcriptase (Promega Corporation, Madison, WI, USA) and
Random Primers (Takara Biotechnology Co., Ltd., Dalian, China).
Real time-PCR was conducted with PowerSybr Green Master Mix
(Applied Biosystems, CA, USA) according to the manufacturer’s
instructions, using specific primers for sex determining region
Y-box 2 (SOX-2; forward, 5′-GAG ACC GAG CTG AAG CCG CC-3′; reverse,
5′-GCC CAG GCG CTT GCT GAT CT-3′), octamer-binding transcription
factor 4 (OCT-4; forward, 5′-AAC TCC GAT GGG GCC TCC CC-3′;
reverse, 5′-CTC GAG CCC AAG CTG CTG GG-3′), NANOG (forward, 5′-AGC
CAG AAG GCC TCA GCA CC-3′; reverse, 5′-GGC CTT CCC CAG CAG CTT
CC-3′) and β-actin (forward, 5′-CAG AGC AAG AGA GGC ATC CT-3′;
reverse, 5′-TTG AAG GTC TCA AAC ATG AT-3′). The relative transcript
copy number per sample was normalized to β-actin and calculated
using the 2[−ΔΔC(T)] method (18).
Soft agar colony formation assay
The soft agar colony formation assay was conducted
using 96-well culture plates. Each well was coated with 50 μl
bottom agar mixture in medium containing 10% FBS and 0.5% agar
(Sigma-Aldrich, St. Louis, MO, USA). The bottom layer was overlaid
with 50 μl top agar mixture in medium containing 10% FBS, 0.35%
agar, and 300 cells (KM12C, KM12SM or KM12l4A). Following a
two-week incubation at 37°C in a humidified atmosphere of 5%
CO2, colonies larger than 100 μm in diameter were
counted with an inverted microscope (Olympus CKX41) equipped with a
camera (Olympus DP72) and image analyzer (DP2-BSW). Colony forming
efficiency was defined as (colonies per well)/(cells seeded per
well). For NK92-mediated cytotoxicity, a soft agar colony formation
assay was performed following co-culture of the target cells with
NK-92 cells for four hours at various effector to target ratios
(0:1, 0.1:1, 1:1, 5:1 and 10:1).
Statistical analysis
Data are presented as the mean ± standard errors.
For comparison of groups, an unpaired Student’s t-test was
performed. P<0.05 was considered to indicate a statistically
significant difference in all experiments.
Results
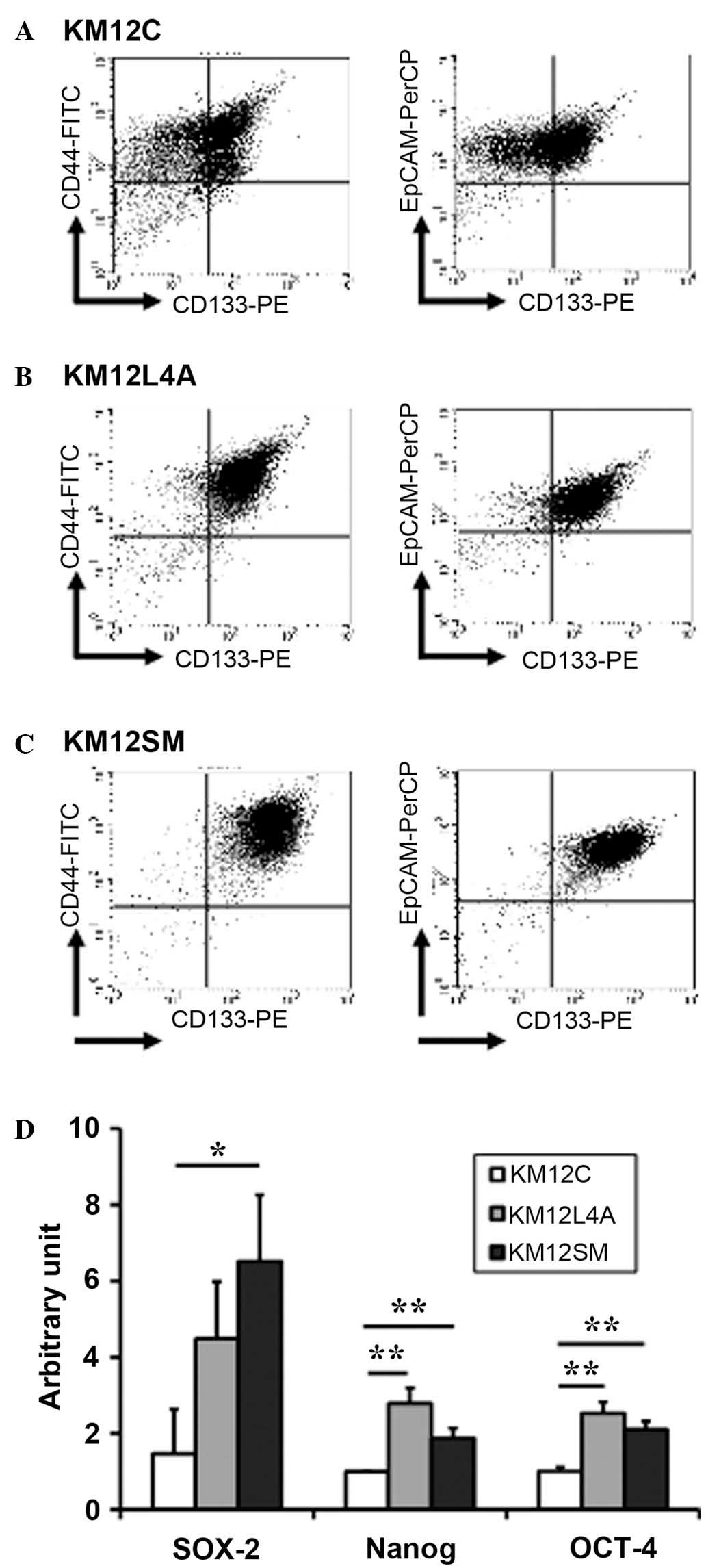
Comparison of CSC markers between primary
and metastatic colon cancer cells
To examine whether highly metastatic cancer cell
populations contain a greater proportion of CSCs, the surface
expression of several putative colon CSC markers was determined in
KM12C (Fig. 1A), KM12L4A (Fig. 1B) and KM12SM (Fig. 1C) cells. The majority of KM12L4A and
KM12SM cells were positive for CD44 (96.89 and 99.19%,
respectively), CD133 (91.9 and 98.29%, respectively) and EpCAM
(97.39 and 98.81%, respectively). Although a high proportion of
KM12C cells were also positive for CD44 and EpCAM, CD133 expression
was only detected in ~55% of these cells. In addition, the mRNA
levels of SOX-2, NANOG and OCT-4, which are essential for
maintaining self-renewal (19),
were higher in KM12L4a and KM12SM compared with that in KM12C cells
(Fig. 1D). These results indicate
that highly metastatic cancer cells may have more cancer stem-like
cells than primary cancer cells.
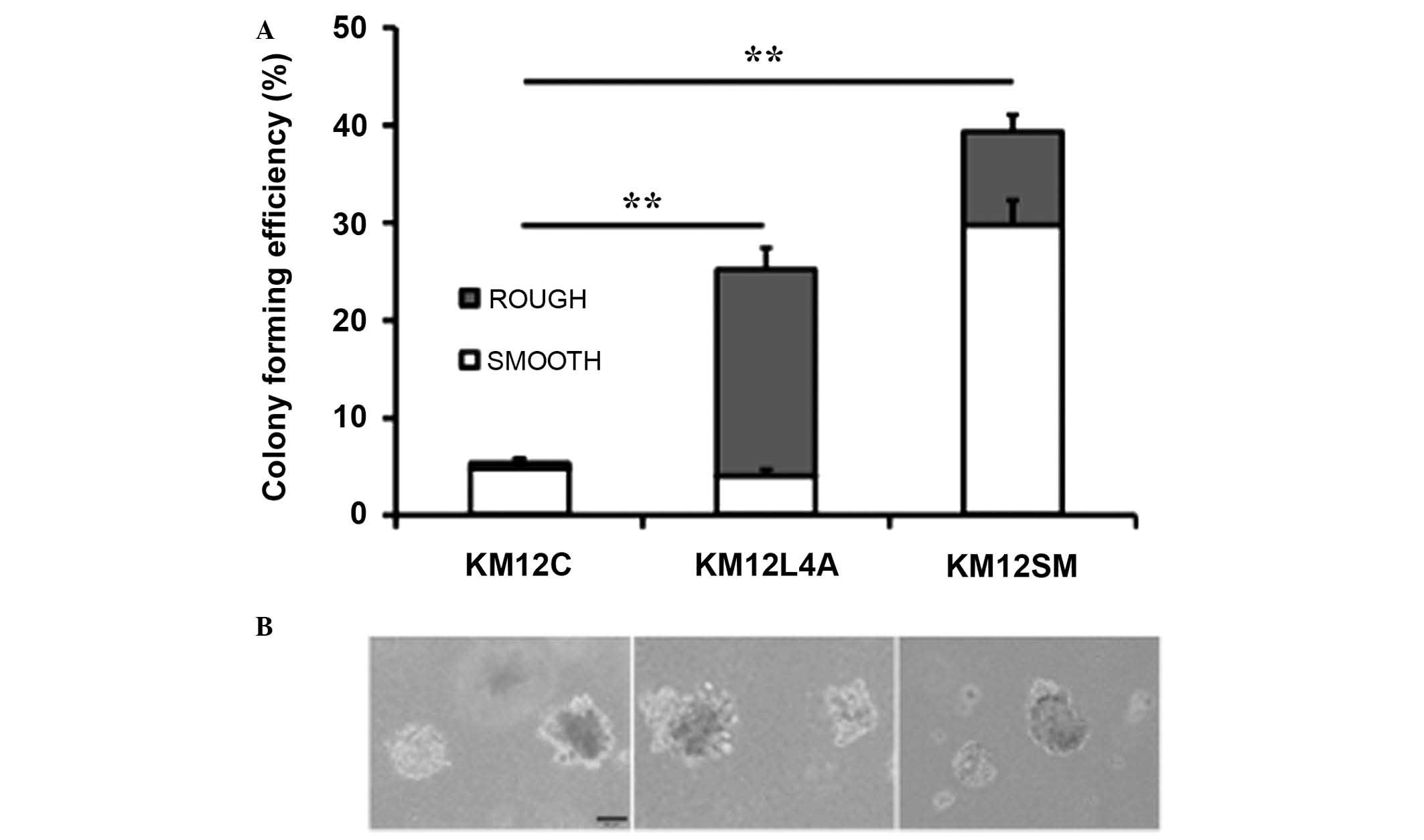
Clonogenicity of metastatic colon cancer
cells
As colony-formation in soft agar is thought to be
characteristic of cancer stem cells (20), the clonogenicity of KM12C cells and
their highly metastatic sublines was investigated. The
clonogenicity of KM12C, KM12L4A and KM12SM cells was approximately
5.4, 25.2 and 39.3%, respectively (Fig.
2). These results were consistent with the levels of putative
cancer stem cell markers and stemness genes measured by flow
cytomtery. However, colony morphology differed between these
sublines. The majority of KM12C colonies possessed a smooth surface
with a spheroid appearance whilst KM12L4A and KM12SM colonies cells
exhibited mixed morphologies. KM12L4A cell colonies had
predominantly rough surfaces, with the appearance of spherical
cellular aggregates, and KM12SM cells colonies had predominantly
smooth surfaces.
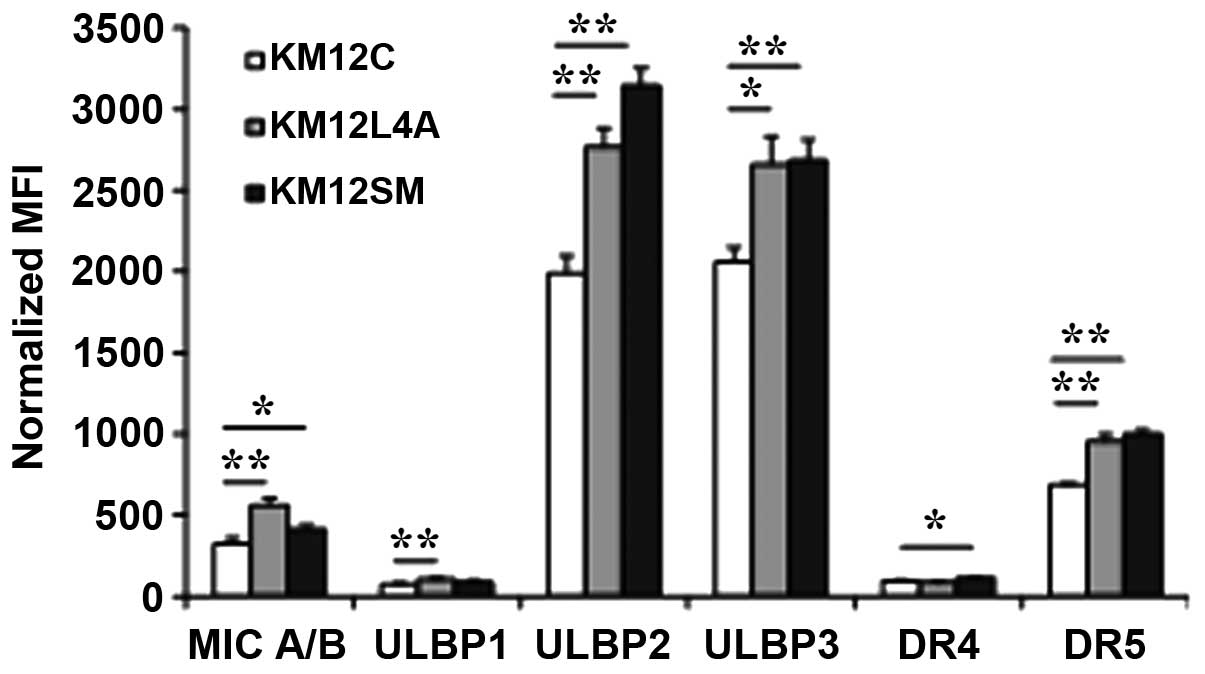
Comparison of NKG2D ligands and death
receptors between primary and metastatic colon cancer cells
The susceptibility of metastatic cancer cells to NK
cell-mediated lysis was investigated by comparing the levels of
activating NKG2D ligands and of TRAIL receptors (DR4 and DR5)
between KM12C, KM12L4A and KM12SM cells (Fig. 3). The surface expression levels of
MICA/B, and ULBP2 and 3 were significantly higher in KM12L4A
(P=0.00175, 0.00116 and 0.01597, respectively) and KM12SM cells
(P=0.02149, 0.00019 and 0.00258) compared with KM12C cells. For
ULBP1, the level of expression was higher in KM12L4A cells than in
KM12C cells. The level of DR5 was also significantly higher in
KM12L4A and KM12SM cells than in KM12C cells. For DR4, the level
was higher in KM12SM cells (P=0.00054) than in KM12C cells. These
data indicate that the levels of NKG2D ligands and death receptors
were generally higher in the metastatic KM12L4A and KM12SM cells
than in the primary KM12C cells, suggesting that metastatic colon
cancer cells may not be resistant to NK cell-mediated lysis.
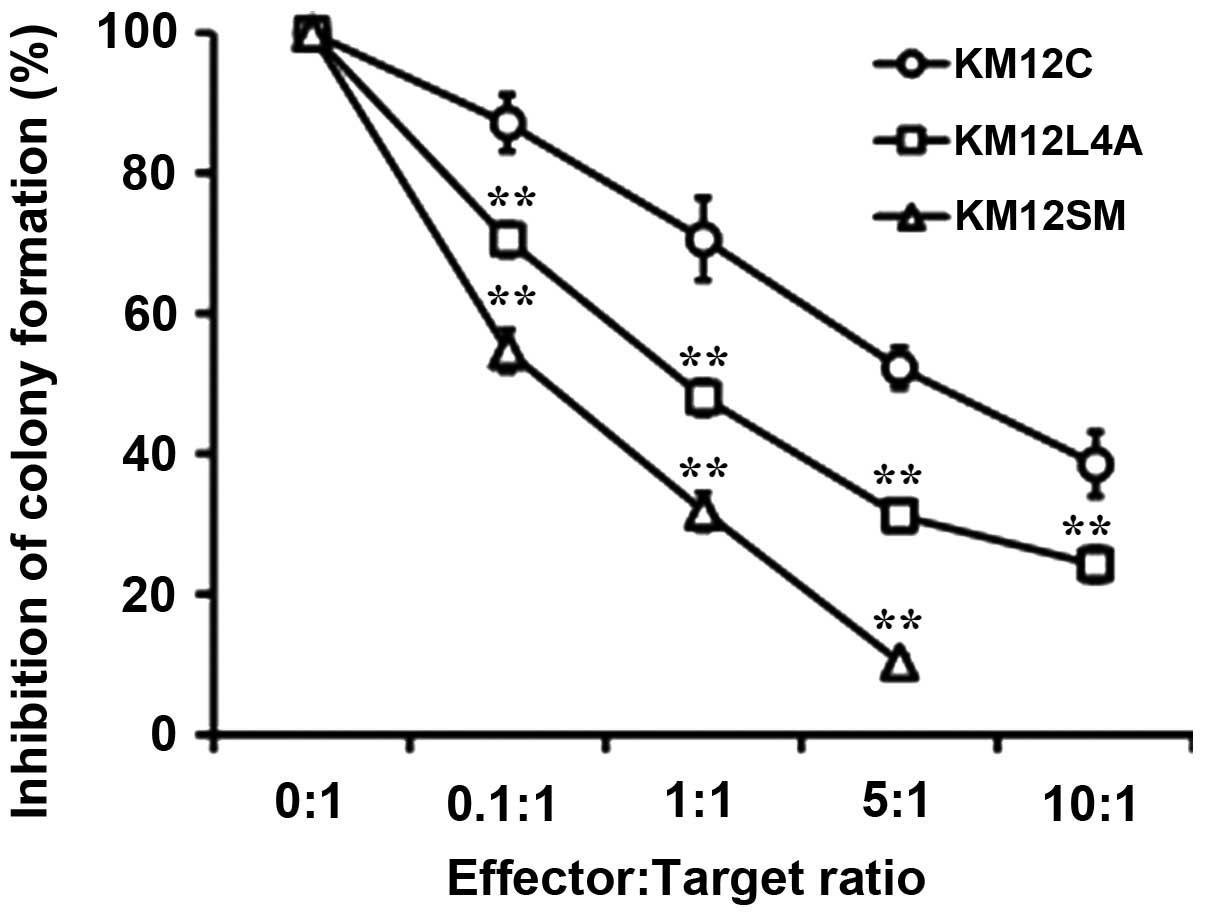
Susceptibility of metastatic colon cancer
cells to NK92 cells
The susceptibility to NK92 cells was compared
between the primary KM12C cells and the metastatic sublines
(Fig. 4). Although the
clonogenicity of the KM12L4A and KM12SM cells was higher than that
of KM12C cells, the susceptibility to NK92 cells of the KM12L4A and
KM12SM cells was also significantly higher than that of KM12C
cells. These results suggest that the metastatic cancer cell
populations, which may contain a greater proportion of cancer
stem-like cells, are not necessarily resistant to NK cell-mediated
lysis.
Discussion
Metastasis is responsible for ≤90% of mortalities
from solid cancers (1,2), primarily due to its systemic nature
and the resistance of disseminated tumor cells to existing
anticancer therapies (3). As CSCs
are thought to be responsible for metastasis and therapy-resistance
(4), the current study investigated
the levels of CSC markers, the clonogenicity in soft agar and the
susceptibility to NK cells of metastatic colon cancer cells,
including the primary KM12C cell line and the highly metastatic
sublines, KM12SM and KM12L4A, which have previously been shown to
be resistant to anticancer drugs and ionizing radiation (21).
Currently, few definitive stem cell markers are
available for the investigation of CSCs in solid tumors. In colon
cancers, certain putative cancer stem cell markers, including
CD133, CD44 and EpCAM, have been used to identify colon CSCs
(5,22). In the current study, a high
percentage of KM12L4A and KM12SM cells were positive for all
markers, including CD44, CD133 and EpCAM. The majority of KM12C
cells were also positive for CD44 and EpCAM, however, only ~55% of
these cells were positive for CD133. In addition, the mRNA levels
of stemness genes, including SOX-2, Nanog and OCT-4 were higher in
KM12L4A and KM12SM than in KM12C cells. Consistently, increased
clonogenicity of the two highly metastatic sublines in soft agar
was observed compared with KM12C cells. KM12SM and KM12L4A were
derived from the parental, poorly metastatic cell line, KM12C,
following injection into the cecum and spleen of nude mice to
produce spontaneous and experimental hepatic metastases,
respectively (15). KM12SM cells
were selected for their growth in the cecum, invasion into the
circulation, and survival and growth in the parenchyma of the
liver; KM12L4A cells were selected for their survival in the
circulation and survival and growth in the liver. Each subline
exhibited a near tetraploid number of chromosomes, in contrast to
the parental KM12C cells, which were near diploid with a minor
tetraploid subpopulation (15,23).
Additionally, a variety of metastasis-associated genes have been
reported to be upregulated and downregulated in KM12SM and KM12L4A
cells compared with in KM12C cells (24). We hypothesize that during the
genetic evolution of KM12SM and KM12L4A cells, the stemness of
these cells may increase due to an increase in the levels of
stemness genes. These genes are associated with distant recurrence
and poor disease-free survival in colorectal cancer following
chemoradiotherapy (25), and
colony-forming activity in soft agar, which is associated with the
characteristics of cancer stem cells, which have been suggested to
be the cell renewal source of a neoplasm and the seeds for the
metastatic spread of cancer (20).
Therefore, the increased levels of putative markers of CSCs and
stemness genes may be associated with the increased clonogenicity
of the highly metastatic KM12SM and KM12L4A cells, suggesting that
highly metastatic colon cancer cell populations may contain a
greater proportion of cancer stem-like cells.
CD133 was used in the initial studies on colon CSCs,
revealing that CD133+ cells isolated from colorectal cancer
exhibited the properties of self-renewal and high tumorigenic
potential, compared with CD133- cells, which were unable to
initiate tumor growth (26,27). It has been demonstrated that CD133
is associated with enhanced colony formation in 2D and 3D culture
in colorectal cancer cells (28).
In the present study, the highly metastatic KM12SM and KM12L4A
cells, which exhibited higher levels of CD133, had greater
clonogenicity compared with the poorly metastatic KM12C cells.
However, the reliability of CD133 as a marker of colon CSCs is
controversial as it has been demonstrated that CD133+ and
CD133-metastatic tumor subpopulations formed colonospheres in in
vitro cultures and were capable of long-term tumorigenesis in a
NOD/SCID serial xenotransplantation model (29,30).
Dalerba et al (31)
demonstrated that the ability to engraft in vivo in
immunodeficient mice was restricted to a minority subpopulation of
CD44+ epithelial cells with high levels of EpCAM expression. In the
current study, the majority of cells of the three KM12 series
sublines were EpCAM+ and CD44+. Therefore, CSC markers other than
CD133, CD44 and EpCAM may be necessary to identify CSCs in KM12
cell populations.
The loss of MHC molecules is often observed in
advanced metastatic cancer cells, rendering tumor cells resistant
to CD8+ T-cell-mediated cytotoxicity (32). The levels of NKG2D ligands (which
can be recognized by other T-cell subsets, including γδ T cells and
NK cells) (33) and of TRAIL
receptors (which induce apoptosis in transformed cells but not in
normal cells) (12) may therefore
affect the susceptibility of the highly metastatic colon cancer
cells to NK cells. In the present study, the levels of NKG2D
ligands and DR4/5 were generally higher in the highly metastatic
KM12L4A and KM12SM cells compared with that in the primary KM12C
cells, and this result was consistent with the increased
susceptibility to NK92 cells of the KM12L4A and KM12SM clonogenic
cells compared with the KM12C clonogenic cells. However, the
clonogenicity of KM12L4A and KM12SM cells was markedly higher than
that of KM12C cells. NK cells are essential in the control of
tumors with upregulated ligands for NK activation receptors and/or
loss of MHC-I molecules (13). The
NKG2D activation receptor binds to a group of ligands that includes
MICA, MICB, and the family of ULBP molecules in humans; the
expression of these molecules may be induced in cells under a
variety of stresses including transformation, heat shock, oxidative
stresses or DNA damage (34–37).
High expression of MIC or RAET1G has been shown to be associated
with prolonged survival of patients with colorectal tumors
(38). It has also been
demonstrated that activated NK cells with membrane-bound TRAIL
enhance NK cell cytotoxicity against neuroblastoma cells (39). In addition, colorectal
carcinoma-derived cancer-initiating cells (CICs) were more
susceptible to freshly purified allogeneic NK cells than the
non-CIC counterpart of the tumors, due to the higher expression of
ligands for NKp30 and NKp44 in the natural cytotoxicity receptor
group of activating NK receptors in CICs (40). Therefore, the results of the present
study suggest that metastatic cancer cells, which may consist of a
greater number of cancer stem-like cells, are not necessarily
resistant to NK cell-mediated lysis, and the levels of NKG2D
ligands and TRAIL receptors may affect the susceptibility of highly
metastatic colon cancer cells to NK-mediated lysis. However,
further studies using other metastatic cancer models are required
to generalize this hypothesis.
Acknowledgements
This work was supported by a 2-Year Research Grant
of Pusan National University.
References
|
1
|
Gupta GP and Massagué J: Cancer
metastasis: building a framework. Cell. 127:679–695. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chaffer CL and Weinberg RA: A perspective
on cancer cell metastasis. Science. 331:1559–1564. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fanali C, Lucchetti D, Farina M, et al:
Cancer stem cells in colorectal cancer from pathogenesis to
therapy: controversies and perspectives. World J Gastroenterol.
20:923–42. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen K, Huang YH and Chen JL:
Understanding and targeting cancer stem cells: therapeutic
implications and challenges. Acta Pharmacol Sin. 34:732–740. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Clevers H: The cancer stem cell: premises,
promises and challenges. Nat Med. 17:313–319. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xie K, Huang S, Dong Z, Gutman M and
Fidler IJ: Direct correlation between expression of endogenous
inducible nitric oxide synthase and regression of M5076 reticulum
cell sarcoma hepatic metastases in mice treated with liposomes
containing lipopeptide CGP 31362. Cancer Res. 55:3123–3131.
1995.PubMed/NCBI
|
|
7
|
Wong CW, Lee A, Shientag L, et al:
Apoptosis: an early event in metastatic inefficiency. Cancer Res.
61:333–338. 2001.PubMed/NCBI
|
|
8
|
Oliver L, Cordel S, Barbieux I, et al:
Resistance to apoptosis is increased during metastatic
dissemination of colon cancer. Clin Exp Metastasis. 19:175–180.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ljunggren HG and Malmberg KJ: Prospects
for the use of NK cells in immunotherapy of human cancer. Nat Rev
Immunol. 7:329–339. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Malmberg KJ, Bryceson YT, Carlsten M, et
al: NK cell-mediated targeting of human cancer and possibilities
for new means of immunotherapy. Cancer Immunol Immunother.
57:1541–1552. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Smyth MJ, Cretney E, Kelly JM, et al:
Activation of NK cell cytotoxicity. Mol Immunol. 42:501–510. 2005.
View Article : Google Scholar
|
|
12
|
Micheau O, Shirley S and Dufour F: Death
receptors as targets in cancer. Br J Pharmacol. 169:1723–1744.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Long EO, Kim HS, Liu D, Peterson ME and
Rajagopalan S: Controlling natural killer cell responses:
integration of signals for activation and inhibition. Annu Rev
Immunol. 31:227–258. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chan CJ, Smyth MJ and Martinet L:
Molecular mechanisms of natural killer cell activation in response
to cellular stress. Cell Death Differ. 21:5–14. 2014. View Article : Google Scholar
|
|
15
|
Morikawa K, Walker SM, Nakajima M, Pathak
S, Jessup JM and Fidler IJ: Influence of organ environment on the
growth, selection, and metastasis of human colon carcinoma cells in
nude mice. Cancer Res. 48:6863–6871. 1988.PubMed/NCBI
|
|
16
|
Elkins K, Zheng B, Go M, et al: FcRL5 as a
target of antibody-drug conjugates for the treatment of multiple
myeloma. Mol Cancer Ther. 11:2222–2232. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hennel R, Brix N, Seidl K, et al: Release
of monocyte migration signals by breast cancer cell lines after
ablative and fractionated γ-irradiation. Radiat Oncol. 9:852014.
View Article : Google Scholar
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
19
|
Jaenisch R and Young R: Stem cells, the
molecular circuitry of pluripotency and nuclear reprogramming.
Cell. 132:567–582. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hamburger AW and Salmon SE: Primary
bioassay of human tumor stem cells. Science. 197:461–463. 1977.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Um JH, Kwon JK, Kang CD, et al:
Relationship between antiapoptotic molecules and metastatic potency
and the involvement of DNA-dependent protein kinase in the
chemosensitization of metastatic human cancer cells by epidermal
growth factor receptor blockade. J Pharmacol Exp Ther.
311:1062–1070. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Alison MR, Lim SM and Nicholson LJ: Cancer
stem cells: problems for therapy? J Pathol. 223:147–161. 2011.
View Article : Google Scholar
|
|
23
|
Camps J, Morales C, Prat E, et al: Genetic
evolution in colon cancer KM12 cells and metastatic derivates. Int
J Cancer. 110:869–874. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
De Lange R, Burtscher H, Jarsch M and
Weidle UH: Identification of metastasis-associated genes by
transcriptional profiling of metastatic versus non-metastatic colon
cancer cell lines. Anticancer Res. 21:2329–2339. 2001.PubMed/NCBI
|
|
25
|
Saigusa S, Tanaka K, Toiyama Y, et al:
Correlation of CD133, OCT4, and SOX2 in rectal cancer and their
association with distant recurrence after chemoradiotherapy. Ann
Surg Oncol. 16:3488–3498. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
O’Brien CA, Pollett A, Gallinger S and
Dick JE: A human colon cancer cell capable of initiating tumour
growth in immunodeficient mice. Nature. 445:106–110. 2007.
View Article : Google Scholar
|
|
27
|
Ricci-Vitiani L, Lombardi DG, Pilozzi E,
et al: Identification and expansion of human
colon-cancer-initiating cells. Nature. 445:111–115. 2007.
View Article : Google Scholar
|
|
28
|
Elsaba TM, Martinez-Pomares L, Robins AR,
et al: The stem cell marker CD133 associates with enhanced colony
formation and cell motility in colorectal cancer. PLoS One.
5:e107142010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shmelkov SV, Butler JM, Hooper AT, et al:
CD133 expression is not restricted to stem cells, and both CD133+
and CD133- metastatic colon cancer cells initiate tumors. J Clin
Invest. 118:2111–2120. 2008.PubMed/NCBI
|
|
30
|
LaBarge MA and Bissell MJ: Is CD133 a
marker of metastatic colon cancer stem cells? J Clin Invest.
118:2021–2024. 2008.PubMed/NCBI
|
|
31
|
Dalerba P, Dylla SJ, Park IK, et al:
Phenotypic characterization of human colorectal cancer stem cells.
Proc Natl Acad Sci USA. 104:10158–10163. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Terunuma H, Deng X, Dewan Z, Fujimoto S
and Yamamoto N: Potential role of NK cells in the induction of
immune responses: implications for NK cell-based immunotherapy for
cancers and viral infections. Int Rev Immunol. 27:93–110. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Fernández-Messina L, Reyburn HT and
Valés-Gómez M: Human NKG2D-ligands: cell biology strategies to
ensure immune recognition. Front Immunol. 3:2992012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kim JY, Son YO, Park SW, et al: Increase
of NKG2D ligands and sensitivity to NK cell-mediated cytotoxicity
of tumor cells by heat shock and ionizing radiation. Exp Mol Med.
38:474–484. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kim JY, Bae JH, Lee SH, et al: Induction
of NKG2D ligands and subsequent enhancement of NK cell-mediated
lysis of cancer cells by arsenic trioxide. J Immunother.
31:475–486. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bae JH, Kim SJ, Kim MJ, et al:
Susceptibility to natural killer cell-mediated lysis of colon
cancer cells is enhanced by treatment with epidermal growth factor
receptor inhibitors through UL16-binding protein-1 induction.
Cancer Sci. 103:7–16. 2012. View Article : Google Scholar
|
|
37
|
Raulet DH, Gasser S, Gowen BG, et al:
Regulation of ligands for the NKG2D activating receptor. Annu Rev
Immunol. 31:413–441. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
McGilvray RW, Eagle RA, Watson NF, et al:
NKG2D ligand expression in human colorectal cancer reveals
associations with prognosis and evidence for immunoediting. Clin
Cancer Res. 15:6993–7002. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sheard MA, Asgharzadeh S, Liu Y, et al:
Membrane-bound TRAIL supplements natural killer cell cytotoxicity
against neuroblastoma cells. J Immunother. 36:319–329. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Tallerico R, Todaro M, Di Franco S, et al:
Human NK cells selective targeting of colon cancer-initiating
cells: a role for natural cytotoxicity receptors and MHC class I
molecules. J Immunol. 190:2381–2390. 2013. View Article : Google Scholar : PubMed/NCBI
|