Introduction
Hepatocellular carcinoma (HCC) is the most common
primary malignant tumor of the liver and, overall, one of the most
frequently diagnosed malignancies worldwide, accounting for >1
million annual mortalities (1). A
large number of risk factors for HCC have been established,
including hepatic viral infections, environmental exposure, alcohol
consumption, genetic metabolic diseases, smoking, cirrhosis and
oral contraceptives (2–4). Furthermore, comparative studies have
identified that males are 2–8 times more likely to develop HCC
compared with females in low- and high-incidence areas (5–7).
Previous studies support that the androgen receptor
(AR) is a major factor responsible for the gender disparity
observed in HCC (7,8). The AR, also known as the nuclear
receptor subfamily 3, group C, member 4, is a 90-kDa
ligand-dependent transcription factor, which regulates the
expression of target genes. As it belongs to the nuclear receptor
superfamily, the AR gene contains three primary functional domains:
i) the N-terminal transactivation domain; ii) the DNA-binding
domain; and iii) the androgen-binding domain (9). The main function of the AR is to mediate
the effects of androgens in important pathological and
physiological processes, including the formation and evolution of
cancer (10,11). Following androgen-mediated activation,
the AR translocates to the nucleus and binds to the androgen
response element (ARE) of its target genes in order to activate
their expression (9,10). In certain conditions, the AR has been
identified to be activated by non-androgenic pathways. Furthermore,
previous studies have reported that AR can be activated by a number
of other molecules, including interleukin-6, β-catenin and
acetylcholine (ACh) (12–14).
The expression of AR has been investigated in HCC
cell lines and tissues (15–17). Previous studies have demonstrated that
the AR protein and mRNA expression levels were >1.5-fold higher
in HCC tissues, and that AR overexpression was associated with the
incidence of HCC (15,17,18). In
addition, the results of AR function analyses in mouse models have
suggested that AR overexpression promotes tumorigenicity (19,20). By
contrast, the suppression of AR by small interfering RNA has been
identified to induce cell cycle arrest at the G1 phase
and inhibit HCC cell growth (18). In
addition, previous data have indicated that the AR is able to
modulate the expression and activity of a number of molecules
within HCC cells (18,21,22). These
molecules are associated with cellular proliferation, tumor growth
and/or metastasis. Notably, studies on transgenic mouse models have
demonstrated that AR may play a negative role in HCC metastasis
(22). The aforementioned findings
indicate that AR exhibits marked oncogenic properties in HCC
initiation and development in the early stages of the disease, but
functions as a suppressor of metastasis in late-stage HCC.
Targeting AR in order to treat HCC at different stages may
represent an improved therapeutic approach compared with androgen
ablation therapy. The present study reviewed the available
literature regarding the significance of AR in
hepatocarcinogenesis.
AR alterations in HCC
Significant efforts have been made in order to
investigate the expression of the AR in HCC tissue specimens and
cell lines (16,23,24). In
early studies, the percentage positivity for AR expression in
tissue specimens was found to vary significantly. These variations
may have been the result of using indirect binding assays, which
were not sufficiently sensitive to detect AR expression (16). Table I
summarizes the findings of recent studies that used human tissue
samples.
 | Table I.Overexpression of androgen receptor
in human tumors. |
Table I.
Overexpression of androgen receptor
in human tumors.
| First author
(Ref.) | Cases investigated,
n | Cases with elevated
AR, n | Year |
|---|
| Feng et al
(18) | 33 | 19 | 2011 |
| Chen et al
(25) | 20 | 16 | 2012 |
| Ma et al
(22) | 10 | 7 | 2008 |
| Barone et al
(15) | 20 | 20 | 2009 |
AR is expressed in a variety of HCC cell lines,
including THLE-2, HepG2, Huh7, SNU-387 and LM-3 (18,21). A
higher AR expression has been detected in cells that exhibit a
larger number of malignant features (21). The difference in AR expression between
HCC cells with high and low metastatic potential suggests that a
high expression of AR may contribute to HCC cell invasion and
migration (21). In previous studies
that also analyzed control tissues, a higher AR expression was
evident in tumoral tissues compared with peritumoral tissues,
revealing that the disruption in AR homeostasis was associated with
the incidence of HCC (15,18). Overexpression of AR in HCC cells may
lead to the dysregulation of several molecules associated with
cellular proliferation, tumor growth and/or metastasis (21,22,25).
Findings from clinical HCC specimens revealed that an elevation in
the levels of microRNA (miR)-216a and cell cycle-related kinase
(CCRK) was positively correlated with an increase in the AR protein
level, and that the overexpression of miR-216a and CCRK was
associated with poor patient prognoses (18,25). By
contrast, Ma et al (22)
established that AR overexpression was only present in tumors with
a size of <3 cm (22). In
addition, these authors identified a low expression of AR in severe
HCC lesions (22). These results
appear to be in agreement with those of Zhu et al (26), who demonstrated that the expression of
AR was higher in peritumoral tissues compared with tumor tissues
(26). Due to these conflicting
results regarding the expression of AR in HCCs, a detailed
investigation into the association between AR expression and tumor
grade during the progression of HCC is required.
At present, the mechanisms underlying AR-induced
hepatocarcinogenesis remain to be elucidated. In the following
sections, recent advances in molecular research that have been
conducted in order to understand the role of AR in
hepatocarcinogenesis are reviewed. The roles of AR are also shown
in Fig. 1.
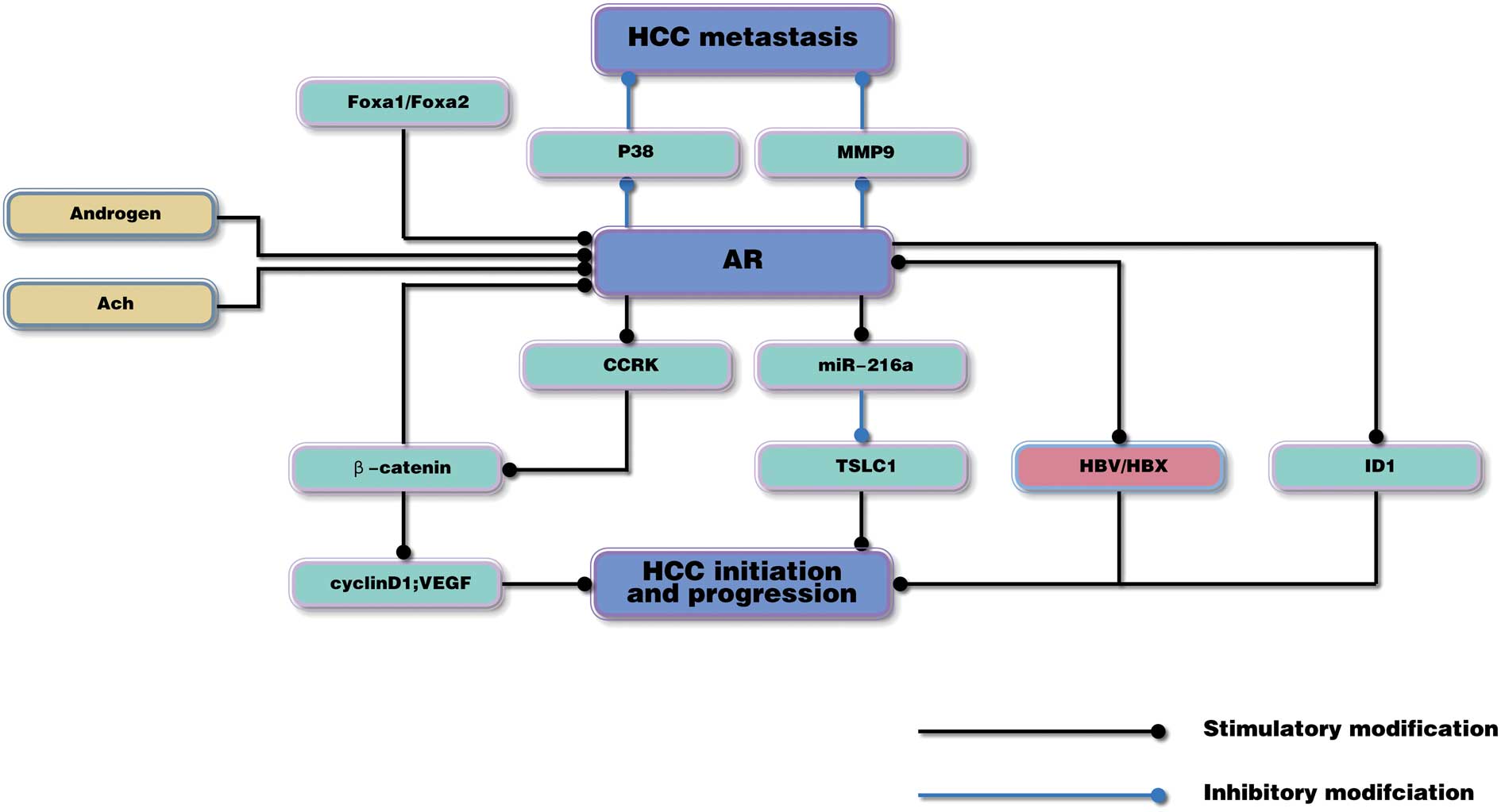 | Figure 1.AR may have a dual role, promoting
HCC initiation in the early stages, but suppressing HCC metastasis
in the advanced stages of the disease. AR, androgen receptor; HCC,
hepatocellular carcinoma; ACh, acetylcholine; Fox, forkhead box;
MMP9, matrix metalloproteinase 9; CCRK, cell cycle-related kinase;
miR-216a, microRNA-216a; TSLC1, tumor suppressor in lung cancer 1;
HBV, hepatitis B virus; HBX, HBV X protein; ID1, inhibitor of
differentiation 1; VEGF, vascular endothelial growth factor. |
AR promotes hepatocarcinogenesis
AR and the Wnt signaling pathway
HCC is a heterogeneous cancer involving the aberrant
activation of Wnt signaling (27).
β-Catenin is an important effector molecule in the Wnt signaling
pathway (28). An abnormal regulation
of β-catenin has been described in up to 90% of HCC cases, which
has been identified as a major early carcinogenic event that occurs
during the development of HCC (29).
The crosstalk between AR and Wnt/β-catenin signaling has been
established by previous genetic studies (18,30,31).
Feng et al (18) revealed that AR may be able to activate
β-catenin during heptocarcinogenesis by modulating CCRK. CCRK is a
novel cyclin-dependent kinase-activating kinase that serves as an
important regulator of tumorigenicity in a number of human
malignancies (32). Ligand-activated
AR has been demonstrated to stimulate the transcription of CCRK in
human liver and HCC cells, while the presence of ARE was identified
in the CCRK promoter region. In addition, the AR-induced CCRK was
demonstrated to activate β-catenin signaling, which is known to be
associated with glycogen synthase kinase 3β phosphorylation. RNA
interference-mediated ablation of CCRK or β-catenin resulted in a
significant suppression of AR-induced G1/S cell cycle
progression. Furthermore, the aberrant expression of CCRK in HCC
cells led to an increase in the levels of AR, whilst the
downregulation of β-catenin attenuated the effects of CCRK on AR.
This suggests that a cycle exists within HCC cells: AR stimulates
CCRK expression in order to induce the activity of β-catenin,
whilst β-catenin acting downstream of CCRK induces the expression
and activity of AR. An upregulation in the expression levels of AR,
CCRK and active β-catenin was observed in primary HCC specimens,
which further supports the role of the AR/CCRK/β-catenin-positive
regulatory circuit in hepatocarcinogenesis (18).
AR and miR-216a
miRNAs are small, non-coding RNA molecules, which
inhibit the expression levels of target genes. Previous studies
have suggested that miRNAs may be another group of host genetic
factors that are associated with hepatocarcinogenesis (33,34).
Increasing evidence demonstrated that dysregulated miRNA expression
is commonly observed in HCCs (35).
However, the mechanism through which a disturbance in cellular
miRNAs affects HCC initiation and progression remains to be
elucidated. A study by Chen et al (25) identified that ligand-stimulated AR was
a regulator of miR-216a and lead to enhanced tumorigenesis.
Furthermore, these authors revealed that the level of miR-216a was
significantly higher in male HCC patients (25). This male-predominant increase in the
expression of miR-216a suggests that the crosstalk between AR and
miR-216a may be involved in hepatocarcinogenesis. In vivo
studies have identified that the ligand-stimulated AR can bind to
ARE site residues within the promoter regions and stimulate the
transcription of primary miR-216a, resulting in an increase in the
expression of miR-216a (25). This
increase stimulates the proliferation, migration and invasion of
liver cancer cells through a decrease in the expression of tumor
suppressor in lung cancer 1 (TSLC1), a protein associated with
tumor invasion and metastasis (36,37). These
results are in agreement with the finding that AR and miR-216a were
concordantly overexpressed in clinical specimens, while an inverse
correlation between TSLC1 and miR-216a was also identified in these
liver tissues (25). The presence of
the hepatitis B virus (HBV) X protein (HBx) can further augment the
effect of AR-mediated miR-216a transcription, due to its known
ability to enhance AR activity (38,39).
Notably, recent studies have indicated that miRNAs, including
miR-224, miR-221, miR-122a and miR-223, demonstrate deregulated
expression patterns in male HCC patients (25). Therefore, the dysregulation in the
expression of these miRNAs is hypothesized to be, at least in part,
mediated by AR. Further investigation is required in order to
validate this hypothesis.
AR and HBV
HBV is a hepatotropic virus, which chronically
infects ∼350 million individuals worldwide (40). Increasing evidence has established a
clear association between persistent HBV infection and the
development of HCC (41–43). Previous studies have revealed that the
long-term risk of developing HCC was markedly increased in patients
with higher serum HBV DNA concentrations. Furthermore, antiviral
therapies have demonstrated a reduction in the occurrence of HCC
following a decrease in the serum levels of HBV DNA (44). Using mice transfected with the entire
HBV genome, which productively produced HBV in their liver, Tian
et al (45) demonstrated that
HBV replicated ∼2 times more efficiently in male mice compared with
female mice. The higher replication efficiency of HBV detected in
the male mice provided an explanation for the gender discrepancy
observed in cases of HCC. Further studies have indicated that
androgen and AR are important for HBV replication (42). In total, two AREs exist within the HBV
genome. The castration of male mice or the introduction of
mutagenesis to remove these two AREs results in a reduction in the
level of HBV surface antigen, DNA and RNA. This observation
confirms the importance of androgen and AR in the mediation of HBV
replication.
A study by Wu et al (46) indicated that an aberrant
overexpression of AR in HCC cells cooperates with HBV to enhance
cell growth and invasion in vitro, and initiate HCC in
vivo. Further studies have demonstrated that HBx, a HBV-encoded
protein required for efficient viral replication, enhanced the gene
transactivation activity of AR in the presence of androgens
(38,39). These data suggest that AR induces a
positive feedback response upon HBV production, in order to mediate
hepatocarcinogenesis as follows: i) The activated AR increases HBV
replication by binding to the ARE within the HBV genome; ii) the
increased level of HBV RNA leads to the production of more HBV
viral antigen and DNA; iii) the upregulated expression of HBx
following AR-enhanced HBV RNA transcription ultimately promotes AR
transactivation; and iv) the increased viral antigens then
cooperate with AR to increase the process of
hepatocarcinogenesis.
AR and forkhead box (FOX)A1/2
FOX proteins are a family of transcription factors,
which regulate the expression of target genes involved in cellular
differentiation, growth and proliferation (47). The family consists of the FOXA1,
FOXA2, and FOXA3 proteins. Previous studies using FOXA-null mice
demonstrated the combined importance of FOXA1 and FOXA2 in liver
cell specification and differentiation, and identified a potential
role for FOXA1/2 in human liver diseases (48–50).
A recent study investigated the association between
FOXA1/2 and AR transactivation (51).
The results demonstrated that a FOXA1/2 deficiency significantly
reduced the tumor size in male mice following the administration of
N-nitrosodiethylamine, which suggests that FOXA1/2 is required in
order for AR to promote tumor growth. Genomic distribution analysis
revealed that the majority of AR-associated genes were also bound
by FOXA1 or FOXA2. The FOXA1/2 and AR binding sites were located
close together at the regulatory regions of the common target
genes. A functional annotation of the FOXA/AR dual target genes
identified that AR and FOXA1/2 promote tumor growth via a number of
pathways, including the cell cycle, DNA replication and cell growth
and proliferation. These results indicated that FOXA1/2 and AR are
involved in the control of gene expression in HCC and are involved
in the promotion of HCC. This finding is further supported by the
observation that the co-occupancy of FOXA1/2 and AR markedly
increased in the livers of male mice following carcinogen
treatment. By contrast, mutation of FOXA1/2 prevented AR from
binding to the common FOXA/AR targets and attenuated the AR-induced
proliferation and malignant transformation of cells (51).
AR and inhibitor of differentiation 1
(ID1)
Previous genetic studies have revealed that the ID1
gene is a potential downstream target of AR in HCC cells (21). ID1 belongs to the basic helix-loop
helix family of genes, which have been detected in multiple
malignant tumors. In addition, the expression levels of these genes
have been associated with invasive features of cancer (52,53).
Increasing evidence has indicated that ID1 is upregulated in HCC
samples and induces cellular proliferation. Overexpression of ID1
has been identified as a marker of unfavorable prognoses in HCC
patients (54,55). A study by Ao et al (21) demonstrated that the androgen agonist,
R1881, increased the migration and invasion of AR-positive HCC
cells through the stimulation of ID1. By contrast, the treatment of
AR-negative HCC cells with R1881 resulted in no change in the mRNA
and protein levels of ID1. These findings suggest that activated AR
is important for the induction of ID1. Furthermore, a depletion in
ID1 levels has been revealed to attenuate AR-induced cell migration
and invasion, which indicates that ID1 is a major mediator of
AR-induced cell migration and invasion. However, the mechanism
underlying ID1 expression induced by activated AR remains to be
elucidated. Chromatin immunoprecipitation assays have revealed that
AR does not bind directly to the ID1 promoter, which suggests that
the increased in ID1 expression that is induced by AR may be a
secondary response (21). Further
investigation is required in order to determine the molecular
mechanisms underlying AR-induced ID1 expression.
Non-androgen-mediated AR signals in
hepatocarcinogenesis
A previous study reported that AR-positive HCC cells
are able to grow in the absence of androgen, and that the addition
of dihydrotestosterone (DHT) had little effect on AR-induced cell
growth (20). Another study, which
stably transfected functional AR into AR-negative HCC cells,
revealed that cell growth was increased in the absence of androgen
(46). These observations indicate
that non-androgen-mediated AR signals may also be involved in
hepatocarcinogenesis.
Increasing evidence suggests that HBx is able to
enhance AR activity and increase AR-mediated cell transformation in
the presence and absence of DHT (39,46), which
may explain the higher incidence of male HBV-associated HCC
(38). Nie et al (14) revealed that ACh promotes the migration
and invasion of HCC cells by activating the AR transcriptional
activity and upregulating AR expression. Previous studies have
demonstrated that the ACh degradation enzyme, acetylcholinesterase,
is downregulated in HCC. This can lead to an increase in the level
of ACh, which subsequently activates the ACh receptor (AChR) and
promotes the proliferation of HCC cells (56). In addition, Nie et al (14) demonstrated that the expression of AR
was upregulated by ACh in a dose-dependent manner in HCC cells, and
that the upregulated expression of AR was inhibited by the AChR
antagonist, mecamylamine. Furthermore, downregulation of AR was
identified to significantly reverse the ACh-induced migration and
invasion of HCC cells, which indicates that AR is a major mediator
of ACh-induced tumorigenicity. Considering the role of
non-androgen-mediated AR signals in hepatocarcinogenesis and the
observation that androgen ablation therapy in treating HCC provided
inconsistent results (57,58), it may be hypothesized that the AR,
rather than androgens, represent an improved therapeutic approach
to target HCC.
AR suppresses HCC metastasis
AR is known to be involved in the promotion of the
genesis and evolution of HCC. However, a previous study revealed
that AR may function as a suppressor of metastasis in advanced
stages of HCC (22). Using an
AR-knockout mouse model with carcinogen-induced HCC, Ma et
al (22) demonstrated that mice
lacking hepatic AR developed HCC later, but exhibited a more severe
malignant phenotype with a higher risk of lung metastasis and
succumbed to the disease earlier. Further biochemical studies
indicated that AR may suppress HCC metastasis via multiple
signaling pathways (22).
p38 kinases are a class of mitogen-activated protein
kinases, which convert extracellular stimuli into a number of
cellular responses, including survival, proliferation, migration
and differentiation (52).
Deregulation of p38 has been identified to be involved in
hepatocarcinogenesis (59,60). Ma et al (22) revealed that a higher AR expression may
enhance cell anoikis by decreasing the phosphorylation and
inhibiting the activity of p38, thereby suppressing metastasis.
However, the mechanism underlying p38 dephosphorylation induced by
AR remains to be elucidated. In addition to regulating HCC invasion
by modulating p38 phosphorylation, AR has been identified to
regulate the expression of matrix metalloproteinase (MMP)9. MMP9 is
the founding member of the MMP family, which is comprised of ≥40
proteins (61). There is clear
evidence to indicate that the expression of MMP9 is higher in cases
of HCC with metastasis (62,63). In addition, molecular analysis has
revealed that AR downregulates MMP9 expression through the
inhibition of nuclear factor-κB signaling (22). These effects of AR on HCC metastasis
suggest that AR is involved in a wide range of cellular signaling
pathways, and has dual roles at different disease stages.
AR as a promising target for cancer
therapy
A number of clinical trials that used various
anti-androgens to treat HCC have yielded poor results (57,58). For
instance, a small-scale phase II clinical trial using flutamide
failed to establish any evidence supporting improved survival.
Similar results were also observed in large-scale population
studies using leuprorelin and flutamide (57,64).
Several hypotheses exist that may be able to explain these
controversial results observed in clinical trials and basic
studies. The current literature suggests that androgen-mediated and
non-androgen-mediated AR signals are involved in
hepatocarcinogenesis, which indicates that the expression of AR
affects tumor growth, whereas the androgen concentration does not
have a similar effect (14,20,46). The
anti-androgens used in clinical trials were developed in order to
compete with androgens for binding to AR. In addition, increasing
evidence suggests that AR is a negative regulator of metastasis in
advanced stages of HCC (22). Yet the
majority of anti-androgen based trials included patients with
advanced stage and unresectable HCCs; therefore, anti-androgens are
likely to be less effective in such cases. On the basis of the
aforementioned evidence, it is conceivable to hypothesize that AR
may be a promising candidate for molecular-targeted HCC therapy.
Targeting the AR should be stage-dependent by blocking the action
of AR during the early stages and upregulating the expression of AR
at the metastatic stage of the disease.
Conclusions
Following the identification of the critical role of
AR signaling in hepatocarcinogenesis and the involvement of
additional nuclear hormone receptor signals, HCC should be
considered as an endocrine-associated neoplasm (18,20).
Although the specific mechanisms underlying AR regulation in HCC
remain to be elucidated, various findings can be merged in order to
provide a perspective of the oncogenic properties of AR in HCC
(18,20–22).
Androgenic and non-androgenic pathways have been demonstrated to
activate the AR during hepatocarcinogenesis (14,18,46). AR
exhibits marked oncogenic properties in the early stages of HCC
development, but may suppress HCC metastasis in the advanced stages
of HCC (22). Several upstream and
downstream targets of AR involved in mediating cellular
proliferation and transformation have been identified in HCC
(14,21,22,25). The
primary objective of these studies was to translate these findings
into clinical applications. The available results support the
hypothesis that targeting the AR, rather than the androgen, may
provide an improved therapeutic approach for the treatment of
HCC.
References
|
1
|
Yang JD and Roberts LR: Hepatocellular
carcinoma: A global view. Nat Rev Gastroenterol Hepatol. 7:448–458.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
El-Serag HB: Epidemiology of viral
hepatitis and hepatocellular carcinoma. Gastroenterology.
142:1264–1273. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ascha MS, Hanouneh IA, Lopez R, Tamimi TA,
Feldstein AF and Zein NN: The incidence and risk factors of
hepatocellular carcinoma in patients with nonalcoholic
steatohepatitis. Hepatology. 51:1972–1978. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rahbari NN, Mehrabi A, Mollberg NM, et al:
Hepatocellular carcinoma: current management and perspectives for
the future. Ann Surg. 253:453–469. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Altekruse SF, McGlynn KA and Reichman ME:
Hepatocellular carcinomaincidence, mortality and survival trends in
the United States from 1975 to 2005. J Clin Oncol. 27:1485–1491.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bruix J and Sherman M: American
Association for the Study of Liver Diseases: Management of
hepatocellular carcinoma: an update. Hepatology. 53:1020–1022.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yeh SH and Chen PJ: Gender disparity of
hepatocellular carcinoma: the roles of sex hormones. Oncology.
78(Suppl 1): 172–179. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ruggieri A, Barbati C and Malorni W:
Cellular and molecular mechanisms involved in hepatocellular
carcinoma gender disparity. Int J Cancer. 127:499–504. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gelmann EP: Molecular biology of the
androgen receptor. J Clin Oncol. 20:3001–3015. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Simental JA, Sar M, Lane MV, French FS and
Wilson EM: Transcriptional activation and nuclear targeting signals
of the human androgen receptor. J Biol Chem. 266:510–518.
1991.PubMed/NCBI
|
|
11
|
Zegarra-Moro OL, Schmidt LJ, Huang H and
Tindall DJ: Disruption of androgen receptor function inhibits
proliferation of androgen-refractory prostate cancer cells. Cancer
Res. 62:1008–1013. 2002.PubMed/NCBI
|
|
12
|
Truica CI, Byers S and Gelmann EP:
Beta-catenin affects androgen receptor transcriptional activity and
ligand specificity. Cancer Res. 60:4709–4713. 2000.PubMed/NCBI
|
|
13
|
Yeh S, Lin HK, Kang HY, Thin TH, Lin MF
and Chang C: From HER2/Neu signal cascade to androgen receptor and
its coactivators: a novel pathway by induction of androgen target
genes through MAP kinase in prostate cancer cells. In: Proc Natl
Acad Sci USA. 96. pp. 5458–5463. 1999; View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nie H, Cao Q, Zhu L, Gong Y, Gu J and He
Z: Acetylcholine acts on androgen receptor to promote the migration
and invasion but inhibit the apoptosis of human hepatocarcinoma.
PLoS One. 8:e616782013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Barone M, Margiotta M, Scavo MP, et al:
Possible involvement of androgen receptor alterations in
hepatocarcinogenesis. Dig Liver Dis. 41:665–670. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kalra M, Mayes J, Assefa S, Kaul AK and
Kaul R: Role of sex steroid receptors in pathobiology of
hepatocellular carcinoma. World J Gastroenterol. 14:5945–5961.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Villa E: Androgen receptor alterations in
hepatocarcinogenesis. Dig Liver Dis. 41:632–633. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Feng H, Cheng AS, Tsang DP, et al: Cell
cycle-related kinase is a direct androgen receptor-regulated gene
that drives β-catenin/T cell factor-dependent hepatocarcinogenesis.
J Clin Invest. 121:3159–3175. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zender L and Kubicka S: Androgen receptor
and hepatocarcinogenesis: what do we learn from HCC mouse models?
Gastroenterology. 135:738–740. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ma WL, Hsu CL, Wu MH, et al: Androgen
receptor is a new potential therapeutic target for the treatment of
hepatocellular carcinoma. Gastroenterology. 135:947–955. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ao J, Meng J, Zhu L, et al: Activation of
androgen receptor induces ID1 and promotes hepatocellular carcinoma
cell migration and invasion. Mol Oncol. 6:507–515. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ma WL, Hsu CL, Yeh CC, et al: Hepatic
androgen receptor suppresses hepatocellular carcinoma metastasis
through modulation of cell migration and anoikis. Hepatology.
56:176–185. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vizoso FJ, Rodriguez M, Altadill A, et al:
Liver expression of steroid hormones and Apolipoprotein D receptors
in hepatocellular carcinoma. World J Gastroenterol. 13:3221–3227.
2007.PubMed/NCBI
|
|
24
|
Tavian D, De Petro G, Pitozzi A, Portolani
N, Giulini SM and Barlati S: Androgen receptor mRNA
under-expression in poorly differentiated human hepatocellular
carcinoma. Histol Histopathol. 17:1113–1119. 2002.PubMed/NCBI
|
|
25
|
Chen PJ, Yeh SH, Liu WH, et al: Androgen
pathway stimulates microRNA-216a transcription to suppress the
tumor suppressor in lung cancer-1 gene in early
hepatocarcinogenesis. Hepatology. 56:632–643. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhu R, Zhang JS, Zhu YZ, et al:
HBx-induced androgen receptor expression in HBV-associated
hepatocarcinoma is independent of the methylation status of its
promoter. Histol Histopathol. 26:23–35. 2011.PubMed/NCBI
|
|
27
|
Dahmani R, Just PA and Perret C: The
Wnt/β-catenin pathway as a therapeutic target in human
hepatocellular carcinoma. Clin Res Hepatol Gastroenterol.
35:709–713. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lee JM, Yang J, Newell P, et al: β-Catenin
signaling in hepatocellular cancer: Implications in inflammation,
fibrosis, and proliferation. Cancer Lett. 343:90–97. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nault JC and Zucman-Rossi J: Genetics of
hepatobiliary carcinogenesis. Semin Liver Dis. 31:173–187. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lee E, Madar A, David G, Garabedian MJ,
Dasgupta R and Logan SK: Inhibition of androgen receptor and
β-catenin activity in prostate cancer. In: Proc Natl Acad Sci USA.
110. pp. 15710–15715. 2013; View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li Y, Zheng Y, Izumi K, et al: Androgen
activates β-catenin signaling in bladder cancer cells. Endocr Relat
Cancer. 20:293–304. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tian Y, Wan H and Tan G: Cell
cycle-related kinase in carcinogenesis. Oncol Lett. 4:601–606.
2012.PubMed/NCBI
|
|
33
|
Takahashi K, Yan I, Wen HJ and Patel T:
microRNAs in liver disease: from diagnostics to therapeutics. Clin
Biochem. 46:946–952. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gailhouste L and Ochiya T: Cancer-related
microRNAs and their role as tumor suppressors and oncogenes in
hepatocellular carcinoma. Histol Histopathol. 28:437–451.
2013.PubMed/NCBI
|
|
35
|
Guo J and Friedman SL: The expression
patterns and clinical significance of microRNAs in liver diseases
and hepatocellular carcinoma. Curr Pharm Des. 19:1262–1272. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liang QL, Chen GQ, Li ZY and Wang BR:
Function and histopathology of a cell adhesion molecule TSLC1 in
cancer. Cancer Invest. 29:107–112. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang J, Ning J, Geng J, Cui B and Dong X:
Down-regulation of tumor suppressor in lung cancer 1 (TSLC1)
expression correlates with poor prognosis in patients with colon
cancer. J Mol Histol. 43:715–721. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chiu CM, Yeh SH, Chen PJ, et al: Hepatitis
B virus X protein enhances androgen receptor-responsive gene
expression depending on androgen level. In: Proc Natl Acad Sci USA.
104. pp. 2571–2578. 2007; View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zheng Y, Chen WL, Ma WL, Chang C and Ou
JH: Enhancement of gene transactivation activity of androgen
receptor by hepatitis B virus X protein. Virology. 363:454–461.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lok AS and McMahon BJ: Chronic hepatitis
B: update 2009. Hepatology. 50:661–662. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xu C, Zhou W, Wang Y and Qiao L: Hepatitis
B virus-induced hepatocellular carcinoma. Cancer Lett. 345:216–222.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Bharadwaj M, Roy G, Dutta K, Misbah M,
Husain M and Hussain S: Tackling hepatitis B virus-associated
hepatocellular carcinoma - the future is now. Cancer Metastasis
Rev. 32:229–268. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Pollicino T, Saitta C and Raimondo G:
Hepatocellular carcinoma: the point of view of the hepatitis B
virus. Carcinogenesis. 32:1122–1132. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Giacomin A, Cazzagon N, Sergio A, Vanin V
and Farinati F: Hepatitis B virus-related hepatocellular carcinoma:
primary, secondary, and tertiary prevention. Eur J Cancer Prev.
20:381–388. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Tian Y, Kuo CF, Chen WL and Ou JH:
Enhancement of hepatitis B virus replication by androgen and its
receptor in mice. J Virol. 86:1904–1910. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wu MH, Ma WL, Hsu CL, et al: Androgen
receptor promotes hepatitis B virus-induced hepatocarcinogenesis
through modulation of hepatitis B virus RNA transcription. Sci
Transl Med. 2:32ra352010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Xia L, Huang W, Tian D, et al: Forkhead
box Q1 promotes hepatocellular carcinoma metastasis by
transactivating ZEB2 and VersicanV1 expression. Hepatology.
59:958–973. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Xia L, Huang W, Tian D, et al: ACP5, a
direct transcriptional target of FoxM1, promotes tumor metastasis
and indicates poor prognosis in hepatocellular carcinoma. Oncogene.
33:1395–1406. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Liang C, Chen W, Zhi X, et al: Serotonin
promotes the proliferation of serum-deprived hepatocellular
carcinoma cells via upregulation of FOXO3a. Mol Cancer. 12:142013.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wang W, He S, Ji J, Huang J, Zhang S and
Zhang Y: The prognostic significance of FOXQ1 oncogene
overexpression in human hepatocellular carcinoma. Pathol Res Pract.
209:353–358. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Li Z, Tuteja G, Schug J and Kaestner KH:
Foxa1 and Foxa2 are essential for sexual dimorphism in liver
cancer. Cell. 148:72–83. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Su Y, Gao L, Teng L, et al: Id1 enhances
human ovarian cancer endothelial progenitor cell angiogenesis via
PI3K/Akt and NF-κB/MMP-2 signaling pathways. J Transl Med.
11:1322013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Guo Q, Guo P, Mao Q, et al: ID1 affects
the efficacy of radiotherapy in glioblastoma through inhibition of
DNA repair pathways. Med Oncol. 30:3252013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ding R, Han S, Lu Y, et al: Overexpressed
Id-1 is associated with patient prognosis and HBx expression in
hepatitis B virus-related hepatocellular carcinoma. Cancer Biol
Ther. 10:299–307. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Damdinsuren B, Nagano H, Kondo M, et al:
Expression of Id proteins in human hepatocellular carcinoma:
relevance to tumor dedifferentiation. Int J Oncol. 26:319–327.
2005.PubMed/NCBI
|
|
56
|
Zhao Y, Wang X, Wang T, et al:
Acetylcholinesterase, a key prognostic predictor for hepatocellular
carcinoma, suppresses cell growth and induces chemosensitization.
Hepatology. 53:493–503. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Groupe d'Etude et de Traitement du
Carcinome Hépatocellulaire: Randomized trial of leuprorelin and
flutamide in male patients with hepatocellular carcinoma treated
with tamoxifen. Hepatology. 40:1361–1369. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Forbes A, Wilkinson ML, Iqbal MJ, Johnson
PJ and Williams R: Response to cyproterone acetate treatment in
primary hepatocellular carcinoma is related to fall in free 5
alpha-dihydrotestosterone. Eur J Cancer Clin Oncol. 23:1659–1664.
1987. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Iyoda K, Sasaki Y, Horimoto M, et al:
Involvement of the p38 mitogen-activated protein kinase cascade in
hepatocellular carcinoma. Cancer. 97:3017–3026. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Koul HK, Pal M and Koul S: Role of p38 MAP
kinase signal transduction in solid tumors. Genes Cancer.
4:342–359. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Zhang J, Zhang D, Wu GQ, Feng ZY and Zhu
SM: Propofol inhibits the adhesion of hepatocellular carcinoma
cells by upregulating microRNA-199a and downregulating MMP-9
expression. Hepatobiliary Pancreat Dis Int. 12:305–309. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Gao J, Ding F, Liu Q and Yao Y: Knockdown
of MACC1 expression suppressed hepatocellular carcinoma cell
migration and invasion and inhibited expression of MMP2 and MMP9.
Mol Cell Biochem. 376:21–32. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Zhang M, Dai C, Zhu H, et al: Cyclophilin
A promotes human hepatocellular carcinoma cell metastasis via
regulation of MMP3 and MMP9. Mol Cell Biochem. 357:387–395. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Chao Y, Chan WK, Huang YS, et al: Phase II
study of flutamide in the treatment of hepatocellular carcinoma.
Cancer. 77:635–639. 1996. View Article : Google Scholar : PubMed/NCBI
|














