Introduction
In Taiwan, head and neck squamous cell carcinoma
(HNSCC) is a highly prevalent malignancy and is associated with the
habit and common risk factor of betel nut chewing (1–3).
Clinical therapies for HNSCC patients consist of multiple-modality
treatment with surgery, radiation and multi-drug chemotherapy
(4,5). Systemic and nodal metastases are the
major causes of mortality associated with HNSCC patients (6,7).
Metastasis involves the matrix metalloproteinases (MMPs), a group
of proteolytic enzymes, which contribute in the degradation of the
basement membrane and extracellular matrix (ECM) (8–10). The
matrix metalloproteinase-2 (MMP-2) is intensely involved in the
invasion and metastasis of HNSCC. Thus, inhibition of metastasis or
downregulation of MMP-2 expression are important goals for
successful therapy (11,12).
Epidermal growth factor receptor (EGFR) is a member
of the receptor tyrosine kinase (RTK) family. EGFR is expressed in
a number of cell types, including epithelial and mesenchymal cells
(13,14). It has been reported that EGFR is
highly expressed in 80–100% of HNSCC patients, and increased
expression of EGFR is often associated with a poor prognosis in
HNSCC (15,16). The EGFR signaling pathways
contribute to the regulation of cancer cell proliferation,
angiogenesis, adhesion, migration, invasion and anti-apoptosis. In
addition, EGFR signaling is triggered by the binding of epidermal
growth factor (EGF), resulting in the dimerization of EGFR
molecules (17,18). Autophosphorylation of the EGFR
through the tyrosine kinase domains leads to the stalling of
downstream signals such as mitogen-activated protein kinases
(MAPKs) (ERK, JNK and p38), serine/threonine kinase AKT and protein
kinase C (PKC) pathways (19,20).
MAPKs are associated with the expression of the components mediated
in MMP promoter induced through AP-1, and its association with
c-fos and c-Jun (20). A number of
studies have suggested that the MAPKs play a central role in
regulating the activities of MMPs (19,20).
Several anti-metastatic agents that target EGFR or their downstream
signal have been studied in HNSCC (14–16).
Clinical studies involving HNSCC treatments have shown that the
combination of cetuximab (Erbitux) with other drugs is an EGFR
inhibitor (21–23). Cetuximab-radiation or
cetuximab-cisplatin combinations exhibited significant improvement
in adverse effects and a significant increase in survival compared
with radiation alone. However, a number of HNSCC patients
eventually manifest acquired resistance to cetuximab or cisplatin
(24).
Gefitinib (Iressa), an EGFR inhibitor, has shown
obvious in vitro and in vivo anticancer activity
through reduced EGFR expression in cancer cell lines, including
prostate, breast, ovarian, colon and HNSCC (25,26).
Preclinical therapy evidence suggests that gefitinib may enhance
anticancer activity compared to a variety of cytotoxic drugs
including platinum derivatives, taxanes, doxorubicin or topotecan
(27). Gefitinib was found to
competitively inhibit the autophosphorylation of the catalytic
domain of the EGFR (19,20). To investigate the potential of
gefitinib and the enhancement of its effects in combination with
other chemotherapeutic agents or natural products, we evaluated the
combination treatment of gefitinib and epigallocatechin gallate
(EGCG). It has been reported that EGCG has a number of biological
functions including induction of cell apoptosis, cell cycle arrest,
cell growth inhibition, anti-angiogenesis and suppression of
metastasis (28–33). EGCG is the most abundant and most
active phenolic constituent of green tea (34). EGCG, has also been extensively
studied in regards to its anticancer activity in a number of cancer
cell lines and in animal tumor models (28–33).
Several studies have also demonstrated that EGCG inhibits the
activation of the receptor tyrosine kinases, such as EGFR,
insulin-like growth factor-1 receptor (IGF-1R), vascular
endothelial growth factor receptor (VEGFR), and their downstream
effectors such as AKT and MAPKs (35,36).
Chen et al has demonstrated that EGCG inhibits cell invasion
of SCC-9 oral cancer cells through the downregulation of MMPs and
u-PA expressions (37). Thus, EGCG
may be useful as an effector for the prevention of cancer
metastasis. In the current study, we aimed to ascertain whether
that combined treatment with EGCG and gefitinib synergistically
inhibits cancer cell invasion and migration by targeting EGFR
signaling pathways. As evidenced by our results, EGCG/gefitinib
combination treatment modulates anti-metastatic effects in an HNSCC
culture system in vitro.
Materials and methods
Materials and reagents
EGCG, dimethyl sulfoxide (DMSO) and anti-actin were
obtained from Sigma-Aldrich Corp. (St. Louis, MO, USA). Gefitinib
was purchased from Toronto Research Chemicals, Inc. (North York,
ON, Canada). Dulbecco’s modified Eagle’s medium (DMEM), fetal
bovine serum (FBS), L-glutamine, penicillin-streptomycin and
trypsin-EDTA were purchased from Gibco/Life Technologies (Carlsbad,
CA, USA). The primary antibodies were obtained as follows:
antibodies for MMP-2, TIMP-2, p-ERK, p-JNK, p-p38, p-AKT and AKT
were obtained from EMD Millipore Corp. (Billerica, MA, USA);
antibodies for p-EGFR, EGFR, ERK, JNK, p38, PKCα and horseradish
peroxidase (HRP)-linked goat anti-mouse IgG, goat anti-rabbit IgG,
were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA,
USA).
Cell culture
The HNSCC CAL-27 cell line was kindly provided by Dr
Pei-Jung Lu (Graduate Institute of Clinical Medicine, National
Cheng Kung University, Tainan, Taiwan). Cells were cultured in 75
cm2 tissue culture flasks (TPP, Techno Plastic Products
AG., Trasadingen, Switzerland) with DMEM supplemented with 10% FBS,
2 mM L-glutamine, 100 units/ml penicillin and 100 μg/ml
streptomycin and grown at 37°C in a humidified 5% CO2
atmosphere and detached by 0.25% Trypsin/0.02% EDTA (38–40).
Transwell invasion assay
The invasive ability of CAL-27 cells was evaluated
using the Boyden chamber assay with Matrigel matrix-coated filters
as previously described (41,42).
Cells (1×104 cells/0.4 ml) were seeded in the upper
chamber of the Transwell inserts (8 μm pore size, EMD Millipore,
Temecula, CA, USA) pre-coated with Matrigel (BD Biosciences,
Bedford, MA, USA) and exposed to DMSO (0.5%, as a control; CTL),
EGCG (25 μM), gefitinib (10 μM) or the combination of gefitinib (10
μM) and different concentrations (25, 50 and 100 μM) of EGCG. DMEM
containing 10% FBS was placed in the lower chamber and cells for
each treatment were incubated for 48 h at 37°C in a humidified
atmosphere with 95% air and 5% CO2. Then, the
non-invasive cells in the upper chamber were removed with a cotton
swab, and the invaded cells were fixed with 4% formaldehyde for 15
min and stained with 2% crystal violet in 2% ethanol for 15 min
after being washed with PBS. The number of cells that penetrated
the membrane was counted and images were captured under a light
microscope at a magnification of ×200, as previously described
(41,42). Each experiment was repeated 3
times.
Wound-healing scratch assay
Approximately 2×105 CAL-27 cells/well
were cultured in 12-well plates after cell monolayers were attached
overnight to 80% confluency by scratching with a 200-μl pipette tip
and then incubated in the presence or absence of EGCG (25 μM),
gefitinib (10 μM) or the combination of gefitinib (10 μM) and
different concentrations (25, 50 and 100 μM) of EGCG for 48 h.
Cells that migrated into the wound region were determined and
images were captured using a phase-contrast microscope (×100) as
previously described (9,41). Five randomly chosen fields were
analyzed for each well and each experiment was performed in
triplicate.
Gelatin zymography analysis
CAL-27 cells (5×105/well) in 12-well
plates were incubated in a serum-free medium with either 25 μM EGCG
alone, 10 μM gefitinib alone or in combination with gefitinib (10
μM) and EGCG at 25, 50 and 100 μM. After a 48-h incubation,
conditioned medium was collected to perform a 10% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) containing
0.1% gelatin (Sigma-Aldrich Corp.). After electrophoresis, the gel
was washed twice with 2.5% Triton X-100 in dH2O twice
for a total of 60 min at 25°C, then were incubated in substrate
buffer (pH 7.6, 50 mM Tris, 10 mM CaCl2, 50 mM and 0.05%
Brij-35) at 37°C for 24 h. After incubation, the gel was stained
with 0.3% Coomassie brilliant blue R250 (Bio-Rad Laboratories,
Hercules, CA, USA) in 50% methanol and 10% acetic acid for 20 min
and de-staining was subsequently performed with 10% acetic acid and
30% methanol to visualize MMP-2 activity as previously described
(43,44). Bands of gelatinolytic activity were
assessed using NIH ImageJ software. The results were performed in 3
independent experiments.
Western blot analysis
The CAL-27 cells (1×107/flask) were
placed in a 75T flask and exposed to 25 μM EGCG, 10 μM gefitinib or
the combination of gefitinib (10 μM) and EGCG at 25, 50 and 100 μM
for indicated time intervals. Cells were harvested and resuspended
in lysis buffer (PRO-PREP™ protein extraction solution; iNtRON
Biotechnology, Seongnam-si, Gyeonggi-do, Korea) as previously
described (45–47). After being centrifuged at 13,000 × g
for 10 min at 4°C, the whole-cell protein extracts were collected
and quantitated using a Bio-Rad protein assay kit (Bio-Rad
Laboratories) with bovine serum albumin (BSA) as the standard. The
protein lysates were determined by 10–12% SDS-PAGE, and then
electro-transferred onto a nitrocellulose membrane using an iBlot™
Dry Blotting System (Invitrogen/Life Technologies) before being
blocked with PBS containing 0.2% Tween-20 and 5% non-fat powdered
milk for 1 h. The membrane was incubated first with antibodies
overnight and bound antibodies were detected using horseradish
peroxidase-conjugated secondary antibody, followed by Immobilon
Western Chemiluminescent HRP substrate (Millipore) and X-ray film
(GE Healthcare, Piscataway, NJ, USA). The protein abundance was
quantified and NIH ImageJ software was used to determine the band
intensity from immunoblotting analysis (47,48).
RNA purification
CAL-27 cells at a density of 1×107 cells
were placed in a 75T flask and incubated without and in combination
with gefitinib (10 μM) and EGCG (100 μM) for 24 h. Cells were
scraped and collected by centrifugation, and total RNA was
subsequently isolated using an Qiagen RNeasy Mini kit (Qiagen,
Inc., Valencia, CA, USA) after being harvested as previously
described (49,50). RNA quantity and purity were assessed
at 260 and 280 nm using a Nanodrop ND-1000 spectrophotometer
(Labtech International Ltd., East Sussex, UK).
Microarray analysis
After RNA extractions, 300 ng of each sample was
amplified and labeled using the GeneChip WT Sense Target Labeling
and Control Reagents (Affymetrix, Inc. Santa Clara, CA, USA) for
expression analysis. Thereafter, hybridization was performed
against the Affymetrix GeneChip Human Gene 1.0 ST array
(Affymetrix, Inc.). The arrays were hybridized for 17 h at 45°C and
60 rpm. Arrays were subsequently washed (Affymetrix Fluidics
Station 450; Affymetrix, Inc.) and stained with
streptavidin-phycoerythrin (GeneChip Hybridization, Wash, and Stain
kit; Affymetrix, Inc.), and were scanned on an Affymetrix
GeneChip® Scanner 3000 (Affymetrix, Inc.). Resulting
data were analyzed using Expression Console software (Affymetrix,
Inc.) with default RMA parameters. Genes regulated by EGCG and
gefitinib with a 1.2-fold change in expression were identified.
Moreover, bioinformatics analysis for these candidate genes was
determined utilizing MetaCore (GeneGo, Inc., St. Joseph, MI, USA)
as previously described (51,52).
Statistical analysis
All data represent the means ± SD from 3 independent
experiments. The differences were evaluated using the Student’s
t-test and were considered statistically significant at
P<0.001.
Results
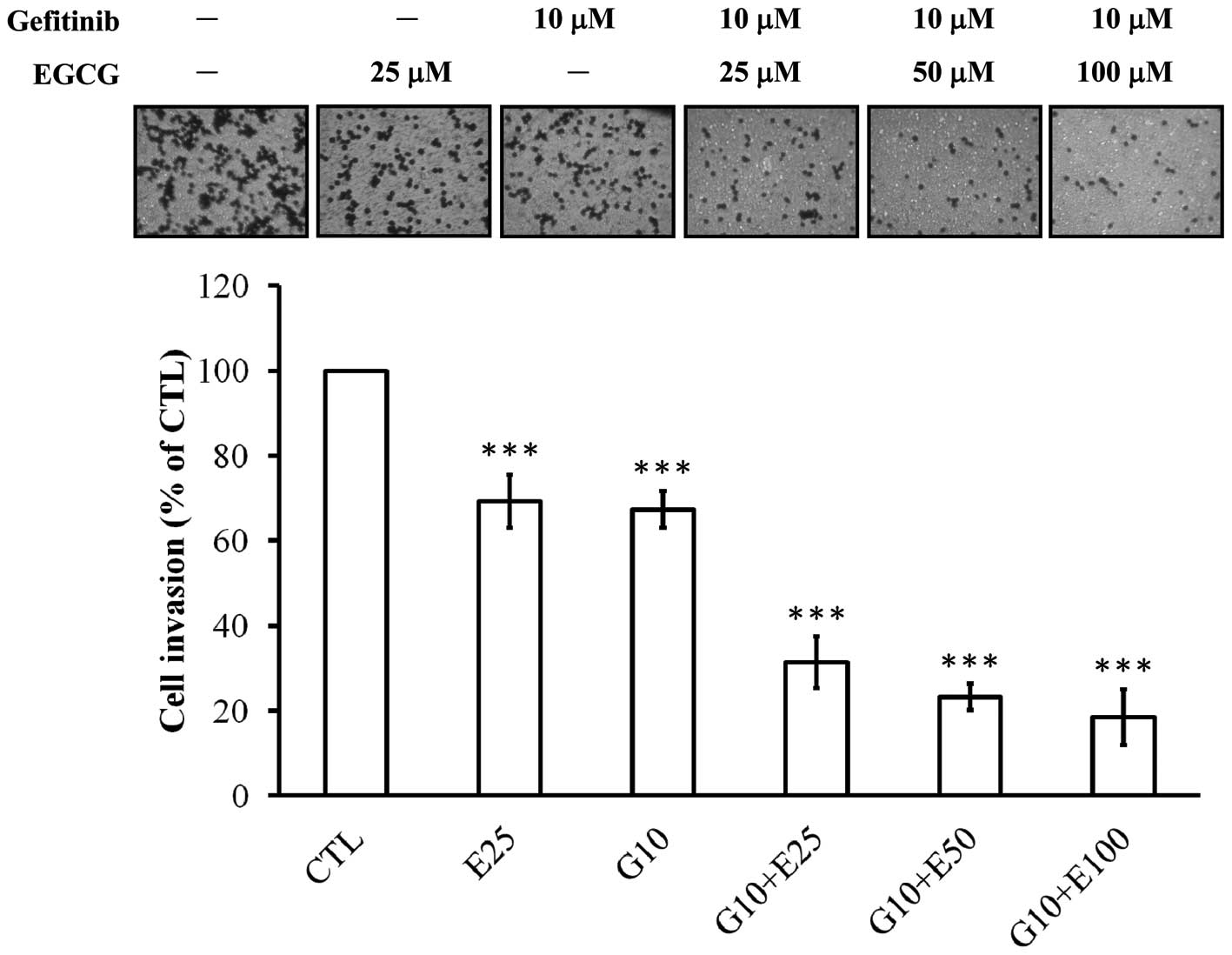
Combined or individual treatment with
EGCG and gefitinib inhibits invasive behavior of CAL-27 cells
The invasion assay revealed that CAL-27 cells
invaded the Matrigel-coated filters from the upper to the lower
chamber in the absence and presence of EGCG, gefitinib or the
combination of both compounds. Our study indicated that individual
treatment with EGCG (25 μM) or gefitinib (10 μM) alone suppressed
the cell invasive ability of CAL-27 cells at 48 h by approximately
31 and 33%, respectively (Fig. 1).
We next investigated the combined effect of gefitinib (10 μM) and
EGCG (25–100 μM). The combination of EGCG and gefitinib exhibited a
synergistic inhibition (at least by 2.12-fold) of the invasive
ability of CAL-27 cells (Fig.
1).
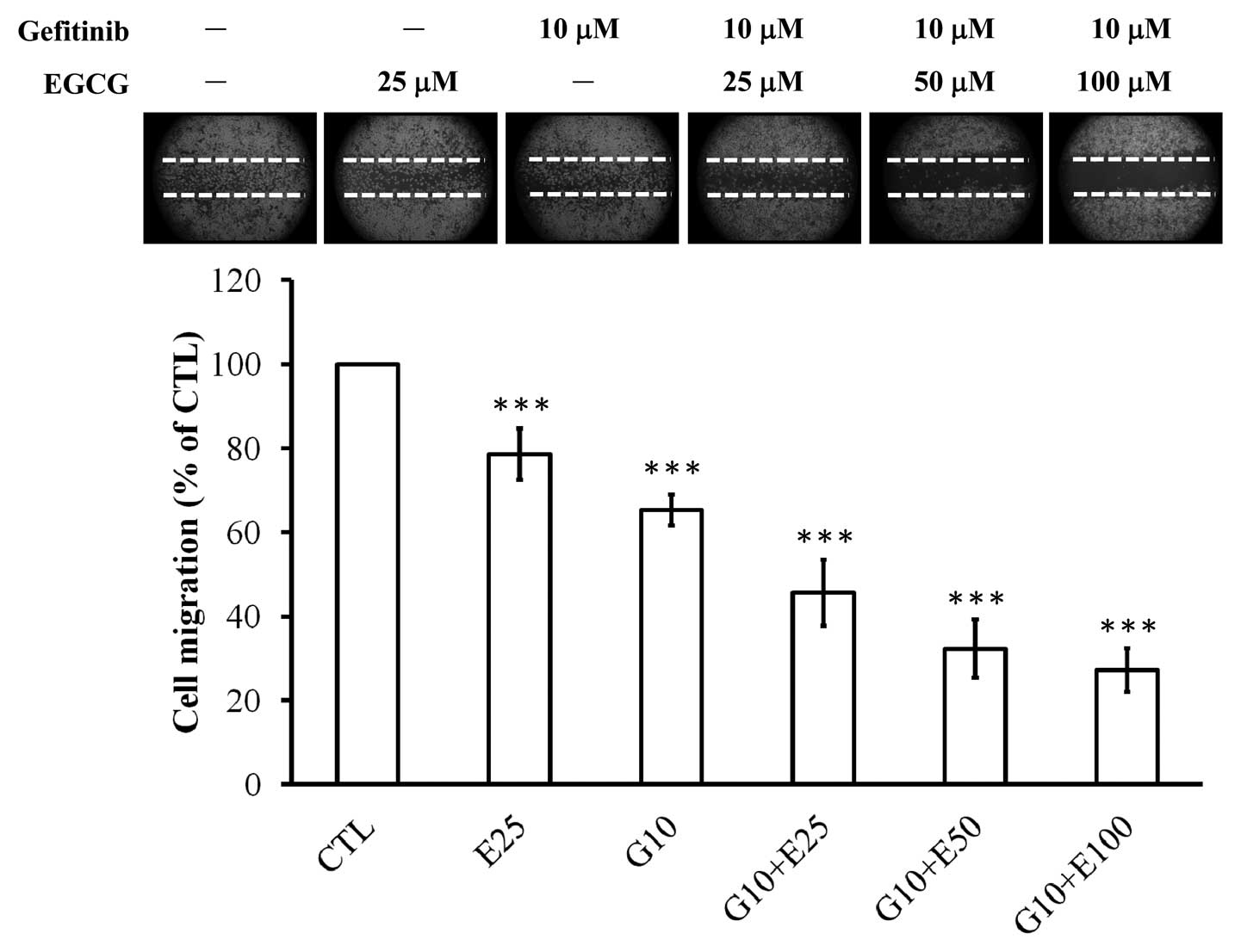
Combined or individual treatment with
EGCG and gefitinib suppresses migratory ability of CAL-27
cells
To measure the effect of cell migration we used the
wound-healing scratch assay. The ability of cells to migrate to a
wounded area in a monolayer was observed. Individual treatment of
EGCG (25 μM) and gefitinib (10 μM) had a significant inhibitory
effect on cell migration of 22 and 34%, respectively, in CAL-27
cells (Fig. 2). We also observed
that the combination treatment of gefitinib (10 μM) and EGCG
(25–100 μM) for 48 h dramatically inhibited the migration of CAL-27
cells into the wounded area; these synergistic effects were
increased by at least 1.56-fold when compared with the effects of
EGCG or gefitinib treatment alone (Fig.
2).
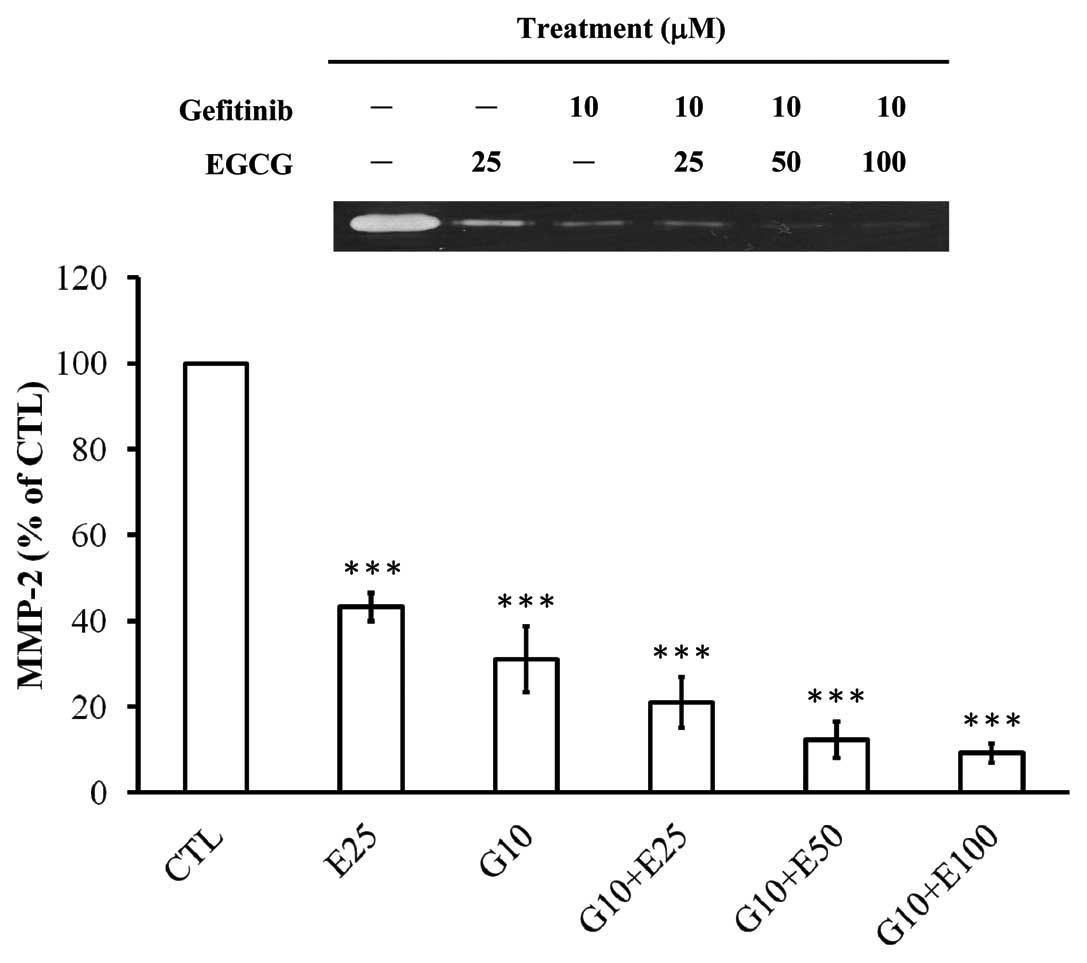
Gefitinib in combination with EGCG
synergistically attenuates the enzymatic MMP-2 activity in CAL-27
cells
We aimed to explore whether gefitinib, EGCG or the
combined treatment of both compounds influence MMP-2 activity in
the conditioned medium of CAL-27 cells. Results from the gelatin
zymographic analysis (Fig. 3)
demonstrated that co-incubation of gefitinib and EGCG (25, 50 and
100 μM) for 48 h synergistically enhanced the suppressive effect on
the activities of MMP-2 in CAL-27 cells, resulting in an additive
inhibition by at least 1.16-fold in comparison to EGCG or gefitinib
individually treated samples. However, EGCG or gefitinib
individually inhibited the enzymatic MMP-2 activity by 57 and 68%,
respectively, in CAL-27 cells.
Combined or individual exposure to EGCG
and gefitinib alters the protein expression associated with the
metastatic ability of CAL-27 cells
To investigate the expression levels of the proteins
associated with the inhibitory effects on the migration and
invasion of CAL-27 cells by the individual and combined effects of
EGCG and gefitinib, western blot analysis was applied and results
are presented in Fig. 4. Individual
or combined treatment with gefitinib and EGCG (25–100 μM) for 48 h
synergistically decreased the protein expression of MMP-2, but
significantly increased the protein levels of TIMP-2 in CAL-27
cells (Fig. 4A). We further
explored the effect of EGCG and gefitinib on upstream signaling
pathways in CAL-27 cells. Data in Fig.
4B revealed that p-EGFR protein expression was suppressed in
the CAL-27 cells after being treated or co-incubated with gefitinib
and EGCG (25, 50 and 100 μM) for 6 h. However, there was no
significant difference in EGFR levels among CAL-27 cells treated
with a combination of gefitinib and EGCG, either agent alone or the
control. Previous studies have reported that the involvement of
MAPKs may be essential for the expression of MMPs and it is
involved in cell invasion and migration during tumor metastasis
(10,53,54).
Our results indicated that gefitinib combined with EGCG
synergistically suppressed the phosphorylated protein expression of
ERK, JNK and p38 but no impact on the protein levels of ERK, JNK
and p38 in the CAL-27 cells was observed compared with the
untreated control (Fig. 4C). We
also revealed that the activation of phospho-AKT (Ser473) and PKCα
protein levels was downregulated in CAL-27 cells after exposure to
gefitinib, EGCG or co-incubation with both compounds for a 6-h
exposure (Fig. 4C).
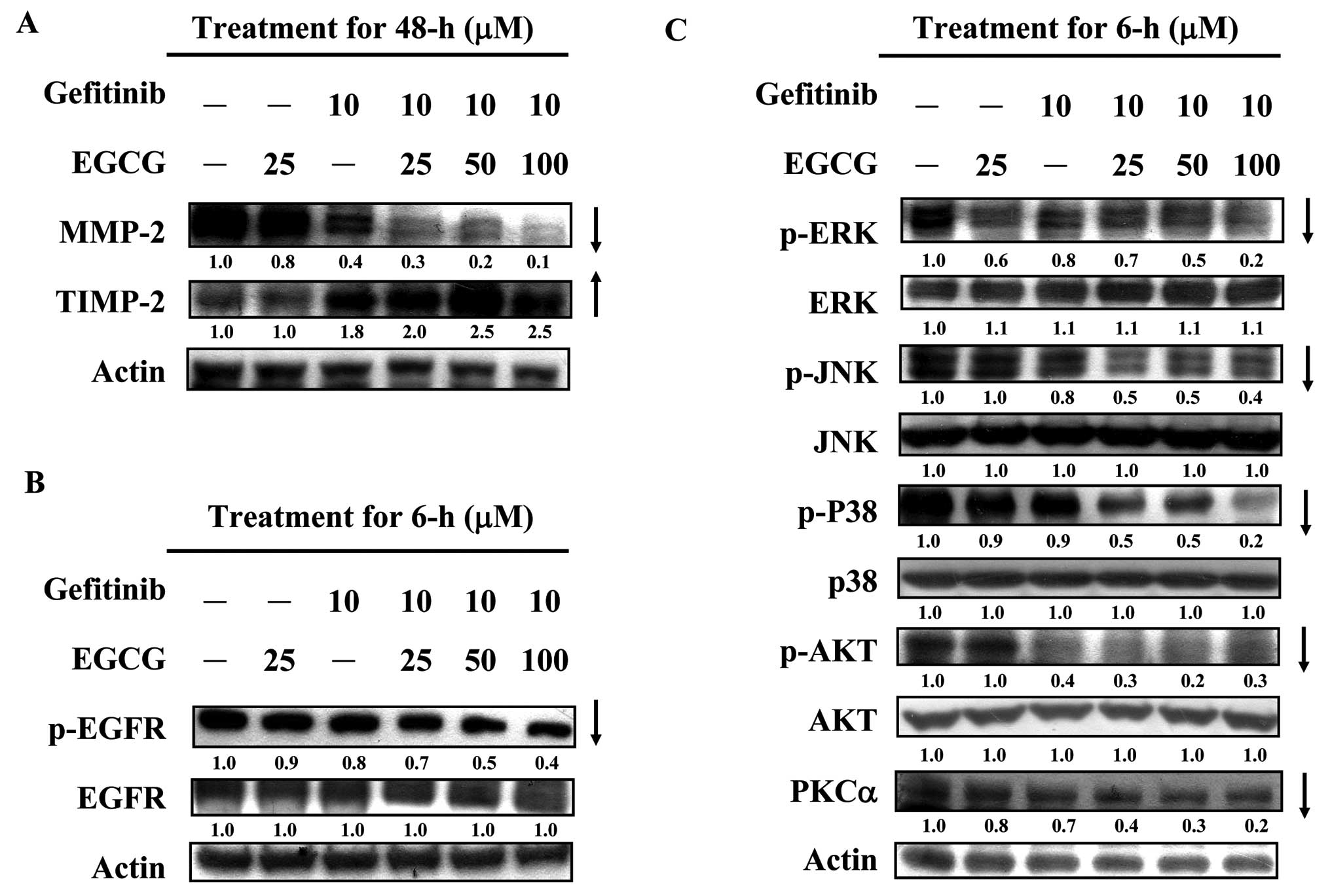 | Figure 4Individual and combined effects of
gefitinib and EGCG on the expression levels of protein associated
with metastatic ability of CAL-27 cells. Cells
(1×107/flask) were placed in a 75T flask and exposed to
25 μM EGCG, 10 μM gefitinib or the combination of gefitinib (10 μM)
and EGCG at 25, 50 and 10 μM for indicated time intervals. After
incubation, the total proteins were collected, and the proteins
levels of (A) MMP-2 and TIMP-2; (B) EGFR and p-EGFR; (C) p-ERK,
ERK, p-JNK, JNK, p-p38, p-38, p-AKT, AKT and PKCα were subjected to
western blotting as described in Materials and methods. Actin
protein level was used as the internal control for equivalent
loading. The results are representative of 3 separate
experiments. |
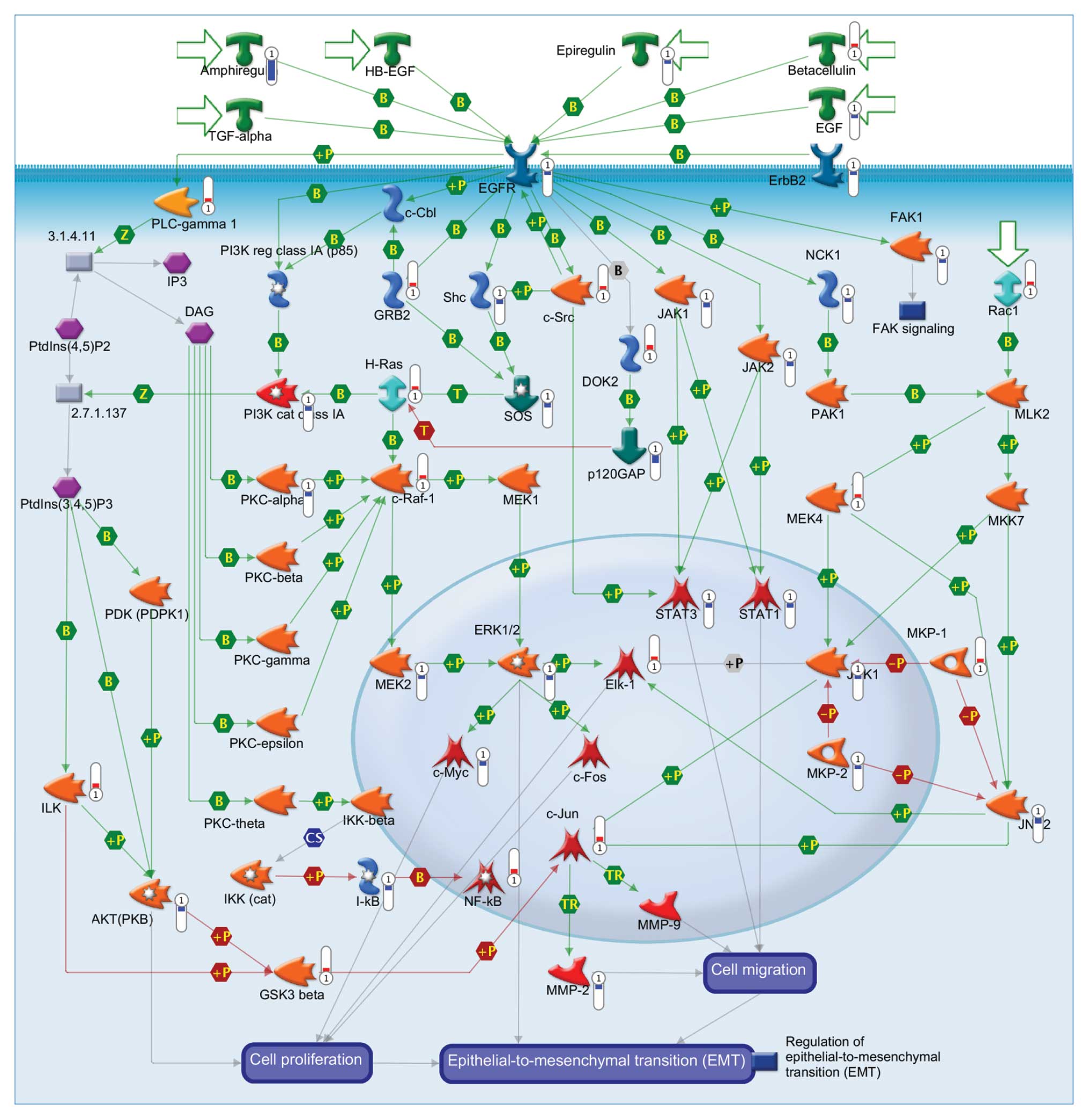
DNA microarray analysis for combined
effects of EGCG and gefitinib-altered anti-metastatic actions in
CAL-27 cells
To examine the gene expression profile in the
combined EGCG and gefitinib-treated CAL-27 cells, DNA microarray
analysis was performed after treatment for 24 h. Our data
demonstrated that 41 genes (15 genes, upregulated; 26 genes,
downregulated) were expressed using microarray analysis. As shown
in Table I, we found that the
levels of c-Jun, ILK, NF-κB, DOK2, c-Src, H-Ras, Rac1, c-Raf-1,
GRB2, betacellulin, GSK3 β, MKP-1, MEK4, Elk-1, PLC-γ 1 were
upregulated in the CAL-27 cells treated with the combination of
EGCG and gefitinib. On the other hand, expression of NCK1, ErbB2,
EGF, SOS, epiregulin, FAK1, JNK1, MEK2, MKP-2, EGFR, STAT3, ERK1/2,
Shc, c-Myc, JAK2, AKT, AKT, PKC-α, I-κB, STAT1, JAK1, MMP-2, PI3K
cat class IA, JNK2, p120GAP and amphiregulin were downregulated in
treated CAL-27 cells (Table I). The
schematic diagram shown in Fig. 5
was observed for the top scorers by the number of network pathways
from the GeneGo analysis program.
 | Table IGenes with more than a 4-fold change
in mRNA level in CAL-27 cells after 24 h treatment with gefitinib
(10 μM) and EGCG (100 μM) as identified by DNA microarray. |
Table I
Genes with more than a 4-fold change
in mRNA level in CAL-27 cells after 24 h treatment with gefitinib
(10 μM) and EGCG (100 μM) as identified by DNA microarray.
| Input IDs | Gene name | Gene symbol | Description | Fold-change |
|---|
| 7916609 | c-Jun | JUN | Transcription
factor AP-1 | 5.91 |
| 7938154 | ILK | ILK | Integrin-linked
protein kinase | 5.04 |
| 7930074 | NF-κB | NFKB2 | NF-κB | 4.62 |
| 8149638 | DOK2 | DOK2 | Docking protein
2 | 4.54 |
| 8062377 | c-Src | SRC | Proto-oncogene
tyrosine-protein kinase Src | 4.45 |
| 7945436 | H-Ras | HRAS | GTPase HRas | 4.37 |
| 8131406 | Rac1 | RAC1 | Ras-related C3
botulinum toxin substrate 1 | 4.33 |
| 8085374 | c-Raf-1 | RAF1 | RAF proto-oncogene
serine/threonine-protein kinase | 4.33 |
| 8018364 | GRB2 | GRB2 | Growth factor
receptor-bound protein 2 | 4.32 |
| 8101002 | Betacellulin | BTC |
Probetacellulin | 4.31 |
| 8089801 | GSK3 β | GSK3B | Glycogen synthase
kinase-3 β | 4.27 |
| 8115831 | MKP-1 | DUSP1 | Dual specificity
protein phosphatase 1 | 4.26 |
| 8005029 | MEK4 | MAP2K4 | Dual specificity
mitogen-activated protein kinase kinase 4 | 4.17 |
| 8172345 | Elk-1 | ELK1 | ETS
domain-containing protein Elk-1 | 4.15 |
| 8062623 | PLC-γ 1 | PLCG1 | PLC-γ 1 | 4.07 |
| 8082911 | NCK1 | NCK1 | Cytoplasmic protein
NCK1 | −4.04 |
| 8006906 | ErbB2 | ERBB2 | Receptor
tyrosine-protein kinase erbB-2 | −4.06 |
| 8096845 | EGF | EGF | Pro-epidermal
growth factor | −4.13 |
| 8051670 | SOS | SOS1 | SOS | −4.16 |
| 8095728 | Epiregulin | EREG | Proepiregulin | −4.23 |
| 8153223 | FAK1 | PTK2 | Focal adhesion
kinase 1 | −4.23 |
| 7927389 | JNK1 | MAPK8 | Mitogen-activated
protein kinase 8 | −4.28 |
| 8032761 | MEK2 | MAP2K2 | Dual specificity
mitogen-activated protein kinase kinase 2 | −4.29 |
| 8150076 | MKP-2 | DUSP4 | Dual specificity
protein phosphatase 4 | −4.32 |
| 8132860 | EGFR | EGFR | Epidermal growth
factor receptor | −4.43 |
| 8015607 | STAT3 | STAT3 | Signal transducer
and activator of transcription 3 | −4.50 |
| 8074791 | ERK1/2 | MAPK1 | ERK1/2 | −4.51 |
| 7920600 | Shc | SHC1 | SHC-transforming
protein 1 | −4.71 |
| 8148317 | c-Myc | MYC | Myc proto-oncogene
protein | −4.87 |
| 8154178 | JAK2 | JAK2 | Tyrosine-protein
kinase JAK2 | −4.96 |
| 7925531 | AKT | AKT3 | AKT (PKB) | −4.98 |
| 7981494 | AKT | AKT1 | AKT (PKB) | −5.06 |
| 8009301 | PKC-α | PRKCA | Protein kinase C α
type | −5.29 |
| 7978644 | I-κB | NFKBIA | I-κB | −5.34 |
| 8057744 | STAT1 | STAT1 | Signal transducer
and activator of transcription 1-α/β | −5.38 |
| 7916747 | JAK1 | JAK1 | Tyrosine-protein
kinase JAK1 | −5.48 |
| 7995681 | MMP-2 | MMP2 | 72 kDa type IV
collagenase | −5.65 |
| 8091009 | PI3K | cat class | IA PIK3CB PI3K cat
class IA | −5.76 |
| 8116402 | JNK2 | MAPK9 | Mitogen-activated
protein kinase 9 | −6.10 |
| 8106784 | p120GAP | RASA1 | Ras
GTPase-activating protein 1 | −10.33 |
| 8095736 | Amphiregulin | AREG | Amphiregulin | −25.41 |
Discussion
EGFR is a major target of TRKs in tumor therapy,
especially in HNSCC (15,16). Grandis et al suggested that
EGFR was overexpressed in ~90% of HNSCC tumors and overexpression
of EGFR was significantly associated with poor prognosis (56). Targeting EGFR is a strategy in
antitumor metastasis in preclinical HNSCC models. In Asia, 7% HNSCC
patients carry the EGFR mutation, and the EGFR mutation in HNSCC is
associated with altered therapeutic responses to EGFR inhibitors
(56,57). Therefore, the study of the
resistance to EGFR inhibition and combinational strategies is
required. Previous studies have shown that enhancement or
synergistic antitumor effects in both in vitro and in
vivo models of HNSCC were found when EGCG was combined with
erlotinib (58). In this study, we
demonstrated that a combined treatment of EGCG and gefitinib
synergistically inhibited invasion (Fig. 1) and migration (Fig. 2) in CAL-27 cells. In addition,
gefitinib in combination with EGCG synergistically attenuated
enzymatic activity and protein level of MMP-2 (Fig. 3). Importantly, EGCG may enhance
gefitinib-suppressed phosphorylation of EGFR in CAL-27 cells in
vitro. With the results presented in this study, the
combination of EGCG and gefitinib may be considered a useful
strategy to pursue in clinical trials.
Our earlier study discovered that EGCG induced
apoptosis through death-receptor, mitochondrial and ER stress
pathways in human adrenal tumor NCI-H295 cells (59). We also found that EGCG-provoked
apoptotic death in TSGH-8301 cells was mediated through targeting
AKT and HSP27 and modulating p-BAD, activating the intrinsic
apoptotic cascade pathway (60).
The previous study reported that EGCG and EGFR inhibitors induce
apoptosis in a number of cancer types, including HNSCC (58). Many studies have also provided
related evidence that EGCG has the potential to reverse the process
of carcinogenesis in HNSCC patients and targeted multiple signaling
pathways (such as EGFR, IGF-1R, VEGFR, MAPKs, AKT and PKC pathways)
resulting in the inhibition of cell metastasis (35,36).
Our results showed that combined exposure to EGCG and gefitinib
suppressed the protein expression of p-EGFR and inhibited the
phosphorylated protein levels of ERK, JNK, p38 and AKT associated
with metastatic actions on CAL-27 cells (Fig. 4). The combined effects of EGCG and
gefitinib altered the anti-metastatic responses of related gene
expression as observed using DNA microarray analysis. Our results
from the DNA microarray analysis demonstrated that the mRNA levels
of ErbB2, SOS, FAK1, JNK1, MEK2,
MKP-2, EGFR, STAT3, ERK1/2,
JAK2, AKT, AKT, PKC-α, JAK1,
MMP-2, PI3K cat class IA, JNK2 were
downregulated in treated CAL-27 cells (Table I). Our results suggest that EGCG may
enhance gefitinib-suppressed phosphorylation of EGFR in HNSCC
CAL-27 cells in vitro. EGCG may be developed as a new class
of chemo-preventive or chemo-therapeutic agent for HNSCC.
In conclusion, EGCG exhibited a synergistic
anti-metastatic activity when combined with gefitinib. In addition
to targeting the common EGFR downstream signaling pathways, our
study suggested novel mechanisms by which the combination of
gefitinib and EGCG results in the depletion of EGFR and ultimately
decreases both total and activated EGFR levels. Our results provide
a promising regimen for future chemoprevention and treatment of
HNSCC.
Acknowledgements
We thank the grant-in-aid DMR-101-023 from the China
Medical University Hospital. This study was also supported by the
grant NSC-101-2313-B-039-008 from the National Science Council,
Republic of China (Taiwan).
References
|
1
|
Liu SY, Lu CL, Chiou CT, et al: Surgical
outcomes and prognostic factors of oral cancer associated with
betel quid chewing and tobacco smoking in Taiwan. Oral Oncol.
46:276–282. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen PT, Kuan FC, Huang CE, et al:
Incidence and patterns of second primary malignancies following
oral cavity cancers in a prevalent area of betel-nut chewing: a
population-based cohort of 26,166 patients in Taiwan. Jpn J Clin
Oncol. 41:1336–1343. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang SC, Tsai CC, Huang ST and Hong YJ:
Betel nut chewing and related factors in adolescent students in
Taiwan. Public Health. 117:339–345. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yu FS, Yang JS, Yu CS, et al: Safrole
induces apoptosis in human oral cancer HSC-3 cells. J Dent Res.
90:168–174. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Choi Y, Kim SY, Kim SH, Yang J, Park K and
Byun Y: Inhibition of tumor growth by biodegradable microspheres
containing all-trans-retinoic acid in a human head-and-neck cancer
xenograft. Int J Cancer. 107:145–148. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Muir C and Weiland L: Upper aerodigestive
tract cancers. Cancer. 75:147–153. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Funk GF, Karnell LH, Robinson RA, Zhen WK,
Trask DK and Hoffman HT: Presentation, treatment, and outcome of
oral cavity cancer: a National Cancer Data Base report. Head Neck.
24:165–180. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sommer G, Rossa C, Chi AC, Neville BW and
Heise T: Implication of RNA-binding protein La in proliferation,
migration and invasion of lymph node-metastasized hypopharyngeal
SCC cells. PLoS One. 6:e254022011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lu Z, Lu N, Li C, et al: Oroxylin A
inhibits matrix metalloproteinase-2/9 expression and activation by
up-regulating tissue inhibitor of metalloproteinase-2 and
suppressing the ERK1/2 signaling pathway. Toxicol Lett.
209:211–220. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ni L, Feng Y, Wan H, et al:
Angiotensin-(1–7) inhibits the migration and invasion of A549 human
lung adenocarcinoma cells through inactivation of the PI3K/Akt and
MAPK signaling pathways. Oncol Rep. 27:783–790. 2012.
|
|
11
|
Kim SA, Kwon SM, Kim JA, Kang KW, Yoon JH
and Ahn SG: 5′-Nitro-indirubinoxime, an indirubin derivative,
suppresses metastatic ability of human head and neck cancer cells
through the inhibition of Integrin beta1/FAK/Akt signaling. Cancer
Lett. 306:197–204. 2011.
|
|
12
|
Liang X, Yang X, Tang Y, et al:
RNAi-mediated downregulation of urokinase plasminogen activator
receptor inhibits proliferation, adhesion, migration and invasion
in oral cancer cells. Oral Oncol. 44:1172–1180. 2008. View Article : Google Scholar
|
|
13
|
Ono M and Kuwano M: Molecular mechanisms
of epidermal growth factor receptor (EGFR) activation and response
to gefitinib and other EGFR-targeting drugs. Clin Cancer Res.
12:7242–7251. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Normanno N, De Luca A, Bianco C, et al:
Epidermal growth factor receptor (EGFR) signaling in cancer. Gene.
366:2–16. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Schuler PJ, Boeckers P, Engers R, et al:
EGFR-specific T cell frequencies correlate with EGFR expression in
head and neck squamous cell carcinoma. J Transl Med. 9:1682011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chiang WF, Liu SY, Yen CY, et al:
Association of epidermal growth factor receptor (EGFR) gene copy
number amplification with neck lymph node metastasis in
areca-associated oral carcinomas. Oral Oncol. 44:270–276. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Korner A, Mudduluru G, Manegold C and
Allgayer H: Enzastaurin inhibits invasion and metastasis in lung
cancer by diverse molecules. Br J Cancer. 103:802–811. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Matsuo M, Sakurai H and Saiki I: ZD1839, a
selective epidermal growth factor receptor tyrosine kinase
inhibitor, shows antimetastatic activity using a hepatocellular
carcinoma model. Mol Cancer Ther. 2:557–561. 2003.
|
|
19
|
Yamaoka T, Frey MR, Dise RS, Bernard JK
and Polk DB: Specific epidermal growth factor receptor
autophosphorylation sites promote mouse colon epithelial cell
chemotaxis and restitution. Am J Physiol Gastrointest Liver
Physiol. 301:G368–G376. 2011. View Article : Google Scholar
|
|
20
|
Hwang YP, Yun HJ, Choi JH, et al:
Suppression of EGF-induced tumor cell migration and matrix
metalloproteinase-9 expression by capsaicin via the inhibition of
EGFR-mediated FAK/Akt, PKC/Raf/ERK, p38 MAPK, and AP-1 signaling.
Mol Nutr Food Res. 55:594–605. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rebucci M, Peixoto P, Dewitte A, et al:
Mechanisms underlying resistance to cetuximab in the HNSCC cell
line: role of AKT inhibition in bypassing this resistance. Int J
Oncol. 38:189–200. 2011.PubMed/NCBI
|
|
22
|
Dias JD, Guse K, Nokisalmi P, et al:
Multimodal approach using oncolytic adenovirus, cetuximab,
chemotherapy and radiotherapy in HNSCC low passage tumour cell
cultures. Eur J Cancer. 46:625–635. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wagenblast J, Baghi M, Arnoldner C, et al:
Effect of bortezomib and cetuximab in EGF-stimulated HNSCC.
Anticancer Res. 28:2239–2243. 2008.PubMed/NCBI
|
|
24
|
Jouan-Hureaux V, Boura C, Merlin JL and
Faivre B: Modulation of endothelial cell network formation in vitro
by molecular signaling of head and neck squamous cell carcinoma
(HNSCC) exposed to cetuximab. Microvasc Res. 83:131–137. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lu Y, Liu P, Van den Bergh F, et al:
Modulation of gene expression and cell-cycle signaling pathways by
the EGFR inhibitor gefitinib (Iressa) in rat urinary bladder
cancer. Cancer Prev Res. 5:248–259. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Normanno N, De Luca A, Maiello MR, et al:
The MEK/MAPK pathway is involved in the resistance of breast cancer
cells to the EGFR tyrosine kinase inhibitor gefitinib. J Cell
Physiol. 207:420–427. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Milano G, Spano JP and Leyland-Jones B:
EGFR-targeting drugs in combination with cytotoxic agents: from
bench to bedside, a contrasted reality. Br J Cancer. 99:1–5. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Peng G, Wargovich MJ and Dixon DA:
Anti-proliferative effects of green tea polyphenol EGCG on
Ha-Ras-induced transformation of intestinal epithelial cells.
Cancer Lett. 238:260–270. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Collins QF, Liu HY, Pi J, Liu Z, Quon MJ
and Cao W: Epigallocatechin-3-gallate (EGCG), a green tea
polyphenol, suppresses hepatic gluconeogenesis through
5′-AMP-activated protein kinase. J Biol Chem. 282:30143–30149.
2007.PubMed/NCBI
|
|
30
|
Guo S, Yang S, Taylor C and Sonenshein GE:
Green tea polyphenol epigallocatechin-3 gallate (EGCG) affects gene
expression of breast cancer cells transformed by the carcinogen
7,12-dimethylbenz[a]anthracene. J Nutr. 135:S2978–S2986.
2005.PubMed/NCBI
|
|
31
|
Ahmed S, Wang N, Lalonde M, Goldberg VM
and Haqqi TM: Green tea polyphenol epigallocatechin-3-gallate
(EGCG) differentially inhibits interleukin-1 beta-induced
expression of matrix metalloproteinase-1 and -13 in human
chondrocytes. J Pharmacol Exp Ther. 308:767–773. 2004. View Article : Google Scholar
|
|
32
|
Annabi B, Currie JC, Moghrabi A and
Beliveau R: Inhibition of HuR and MMP-9 expression in
macrophage-differentiated HL-60 myeloid leukemia cells by green tea
polyphenol EGCg. Leuk Res. 31:1277–1284. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu L, Lai CQ, Nie L, et al: The
modulation of endothelial cell gene expression by green tea
polyphenol-EGCG. Mol Nutr Food Res. 52:1182–1192. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Siddiqui IA, Malik A, Adhami VM, et al:
Green tea polyphenol EGCG sensitizes human prostate carcinoma LNCaP
cells to TRAIL-mediated apoptosis and synergistically inhibits
biomarkers associated with angiogenesis and metastasis. Oncogene.
27:2055–2063. 2008. View Article : Google Scholar
|
|
35
|
Masuda M, Wakasaki T, Toh S, Shimizu M and
Adachi S: Chemoprevention of head and neck cancer by green tea
extract: EGCG-the role of EGFR signaling and ‘Lipid Raft’. J Oncol.
2011:5401482011.PubMed/NCBI
|
|
36
|
Khan N and Mukhtar H: Multitargeted
therapy of cancer by green tea polyphenols. Cancer Lett.
269:269–280. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chen PN, Chu SC, Kuo WH, Chou MY, Lin JK
and Hsieh YS: Epigallocatechin-3 gallate inhibits invasion,
epithelial-mesenchymal transition, and tumor growth in oral cancer
cells. J Agric Food Chem. 59:3836–3844. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jiang L, Ji N, Zhou Y, et al: CAL 27 is an
oral adenosquamous carcinoma cell line. Oral Oncol. 45:e204–e207.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Fan MJ, Lin YC, Shih HD, et al: Crude
extracts of Agaricus brasiliensis induce apoptosis in human oral
cancer CAL 27 cells through a mitochondria-dependent pathway. In
Vivo. 25:355–366. 2011.PubMed/NCBI
|
|
40
|
Chien MH, Ying TH, Hsieh YS, et al:
Dioscorea nipponica Makino inhibits migration and invasion of human
oral cancer HSC-3 cells by transcriptional inhibition of matrix
metalloproteinase-2 through modulation of CREB and AP-1 activity.
Food Chem Toxicol. 50:558–566. 2012. View Article : Google Scholar
|
|
41
|
Lai KC, Huang AC, Hsu SC, et al: Benzyl
isothiocyanate (BITC) inhibits migration and invasion of human
colon cancer HT29 cells by inhibiting matrix metalloproteinase-2/-9
and urokinase plasminogen (uPA) through PKC and MAPK signaling
pathway. J Agric Food Chem. 58:2935–2942. 2010. View Article : Google Scholar
|
|
42
|
Yu FS, Huang AC, Yang JS, et al: Safrole
induces cell death in human tongue squamous cancer SCC-4 cells
through mitochondria-dependent caspase activation cascade apoptotic
signaling pathways. Environ Toxicol. 27:433–444. 2011.
|
|
43
|
Troeberg L and Nagase H: Zymography of
metalloproteinases. Curr Protoc Protein Sci. Chapter 21(Unit 21):
152004. View Article : Google Scholar
|
|
44
|
Lin JJ, Hsu HY, Yang JS, et al: Molecular
evidence of anti-leukemia activity of gypenosides on human myeloid
leukemia HL-60 cells in vitro and in vivo using a HL-60 cells
murine xenograft model. Phytomedicine. 18:1075–1085. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chang YC, Lai TY, Yu CS, et al: Emodin
induces apoptotic death in murine myelomonocytic leukemia WEHI-3
cells in vitro and enhances phagocytosis in leukemia mice in vivo.
Evid Based Complement Alternat Med. 2011:5235962011.PubMed/NCBI
|
|
46
|
Chou ST, Peng HY, Chang CT, et al:
Zanthoxylum ailanthoides Sieb and Zucc. extract inhibits growth and
induces cell death through G2/M-phase arrest and activation of
apoptotic signals in colo 205 human colon adenocarcinoma cells.
Anticancer Res. 31:1667–1676. 2011.PubMed/NCBI
|
|
47
|
Lu CC, Yang JS, Huang AC, et al:
Chrysophanol induces necrosis through the production of ROS and
alteration of ATP levels in J5 human liver cancer cells. Mol Nutr
Food Res. 54:967–976. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Chiang JH, Yang JS, Ma CY, et al:
Danthron, an anthraquinone derivative, induces DNA damage and
caspase cascades-mediated apoptosis in SNU-1 human gastric cancer
cells through mitochondrial permeability transition pores and
Bax-triggered pathways. Chem Res Toxicol. 24:20–29. 2011.
View Article : Google Scholar
|
|
49
|
Chung JG, Chang HL, Lin WC, Yeh FT and
Hung CF: Effects of ibuprofen on arylamine N-acetyltransferase
activity in human colon tumor cells. J Appl Toxicol. 19:1–6. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Gardina PJ, Clark TA, Shimada B, et al:
Alternative splicing and differential gene expression in colon
cancer detected by a whole genome exon array. BMC Genomics.
7:3252006. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Yeh MH, Tsai TC, Kuo HP, et al: Lentiviral
short hairpin RNA screen of human kinases and phosphatases to
identify potential biomarkers in oral squamous cancer cells. Int J
Oncol. 39:1221–1231. 2011.PubMed/NCBI
|
|
52
|
Saeed AI, Sharov V, White J, et al: TM4: a
free, open-source system for microarray data management and
analysis. Biotechniques. 34:374–378. 2003.PubMed/NCBI
|
|
53
|
Yang HL, Kuo YH, Tsai CT, et al:
Anti-metastatic activities of Antrodia camphorata against human
breast cancer cells mediated through suppression of the MAPK
signaling pathway. Food Chem Toxicol. 49:290–298. 2011. View Article : Google Scholar
|
|
54
|
Deng YT and Lin JK: EGCG inhibits the
invasion of highly invasive CL1–5 lung cancer cells through
suppressing MMP-2 expression via JNK signaling and induces G2/M
arrest. J Agric Food Chem. 59:13318–13327. 2011.PubMed/NCBI
|
|
55
|
Sok JC, Coppelli FM, Thomas SM, et al:
Mutant epidermal growth factor receptor (EGFRvIII) contributes to
head and neck cancer growth and resistance to EGFR targeting. Clin
Cancer Res. 12:5064–5073. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Rubin Grandis J, Melhem MF, Barnes EL and
Tweardy DJ: Quantitative immunohistochemical analysis of
transforming growth factor-alpha and epidermal growth factor
receptor in patients with squamous cell carcinoma of the head and
neck. Cancer. 78:1284–1292. 1996.
|
|
57
|
Rubin Grandis J, Melhem MF, Gooding WE, et
al: Levels of TGF-alpha and EGFR protein in head and neck squamous
cell carcinoma and patient survival. J Natl Cancer Inst.
90:824–832. 1998.
|
|
58
|
Zhang X, Zhang H, Tighiouart M, et al:
Synergistic inhibition of head and neck tumor growth by green tea
(−)-epigallocatechin-3-gallate and EGFR tyrosine kinase inhibitor.
Int J Cancer. 123:1005–1014. 2008.
|
|
59
|
Wu PP, Kuo SC, Huang WW, et al:
(−)-Epigallocatechin gallate induced apoptosis in human adrenal
cancer NCI-H295 cells through caspase-dependent and
caspase-independent pathway. Anticancer Res. 29:1435–1442.
2009.
|
|
60
|
Chen NG, Lu CC, Lin YH, et al: Proteomic
approaches to study epigallocatechin gallate-provoked apoptosis of
TSGH-8301 human urinary bladder carcinoma cells: roles of AKT and
heat shock protein 27-modulated intrinsic apoptotic pathways. Oncol
Rep. 26:939–947. 2011.
|