Introduction
Lung cancer is the leading cause of cancer-related
death throughout the world (1).
Since the lack of symptoms and little help from traditional X-rays
on early stage lung cancer, most patients are diagnosed with
advanced stage disease and thus have poor prognosis. Therefore,
there is an urgent need to explore new biomarkers to diagnose and
therapeutic strategies to treat this disease at its early
stages.
Previous studies showed that a great majority of
NSCLC highly expressed estrogen receptor beta (ERβ) (2–6). Human
normal bronchial and alveolar epithelial cells were also reported
to express ERβ, which was considered to contribute to the
maintenance of normal lung tissue (7). Niikawa et al first reported
that intratumoral estradiol (an activator of ERβ) concentration was
significantly higher than that of corresponding non-neoplastic lung
tissues, and positively associated with tumor size and Ki-67
labeling index (LI) in ER-positive NSCLC tissues (8). Aromatase (estrogen synthetase)
expression was also detected in NSCLC tissues (9,10). It
was reported that activated ERβ signaling promoted carcinogenisis
through stimulating abnormal cell proliferation (8,11).
Steroid hormones play important physiological roles
in cell differentiation, development, and homeostasis through their
binding to specific receptors of the nuclear receptor superfamily
(12–16). Some studies showed that human
testicular orphan nuclear receptor-4 (TR4) repressed the
ERβ-mediated transactivation of its downstream signal pathways in
lung cancer cells and indicated that TR4 might be a potential tumor
suppressor gene (17). We
previously found that TR4-associated protein (TRA16) suppressed the
TR4-mediated ERβ activity and its downstream signaling by
interrupting binding of TR4 with TR4 response element (TR4RE) and
blocking TR4 dimerization in lung cancer cells (18), suggesting that TRA16 plays a role in
lung cancer development.
TR2 is highly homologous with TR4 and both were
reported to act as transcriptional factors to regulate gene
expression in embryonic stem cells and early embryos (19). TR2 was also shown to repress the
ERβ-mediated transactivation of its downstream signal pathway in
lung cancer cells (20). Thus, this
study further investigated the role of TRA16 in NSCLC and the
relationships among TRA16, TR2 and ERβ. We demonstrate that TRA16
is highly expressed in NSCLC tissues, and its expression level is
increased with increased malignancy and negatively correlated with
survival of NSCLC patients. Furthermore, we show that TRA16
promotes cancer cell growth through activating ERβ and blocking TR2
activity.
Materials and methods
Patients, tissue specimens, and cell
lines
This study was approved by Institutional Review
Committee of Beijing Cancer Hospital. As summarized in Table I, a total of 154 NSCLC specimens
were obtained from patients who underwent surgical resection from
1995 to 2000 in the Department of Thoracic Surgery II of Beijing
Cancer Hospital. The median overall survival time was obtained from
all the patients examined and the follow-up was from September 1995
till September 2005. The median follow-up time was 38.9 months
(from 1.5 to 109 months). Lung tissues from 12 benign lung disease
patients and 2 thoracic injury patients were collected as controls.
All the patients and controls gave informed consent for the
study.
 | Table IClinicopathological characteristics of
patients. |
Table I
Clinicopathological characteristics of
patients.
| Characteristic | No. of patients
(%) |
|---|
| Gender |
| Male | 113 (73) |
| Female | 41 (27) |
| Age |
| ≤60 | 68 (44) |
| >60 | 86 (56) |
| Histology |
| Squamous
carcinoma | 73 (47) |
|
Adenocarcinoma | 70 (45) |
| Others | 11 (7) |
| TNM stage |
| I | 54 (35) |
| II | 54 (35) |
| IIIA | 46 (30) |
| Tumor size |
| T1 | 18 (12) |
| T2 | 62 (40) |
| T3 | 74 (48) |
| Lymph node
metastasis |
| No | 105 (68) |
| Yes | 49 (32) |
| Tumor thrombus |
| No | 131 (85) |
| Yes | 23 (15) |
|
Differentiation |
| Poor | 66 (43) |
| Moderate | 46 (30) |
| High | 39 (25) |
NSCLC cell types included squamous carcinoma,
adenocarcinoma (including bronchioloalveolar carcinoma), large cell
carcinoma, adenosqumous carcinoma, and salivary adenocarcinoma.
Benign diseases included inflammatory pseudotumor, tuberculosis,
sclerotic hemangioma, and harmatoma. The tissue samples were
histologically confirmed by two independent pathologists. Monkey
kidney cell line COS-1 was obtained from the American Type Culture
Collection and maintained in DMEM with 10% fetal bovine serum (FBS)
(Gibco).
Tissue microarray and immunohistochemical
(IHC) analysis
Tissue microarray was constructed as described
previously using a manual tissue arrayer (Beecher Instruments)
(21). Briefly, H&E slides of
each tissue sample were reviewed by two pathologists. A tumor
tissue block was chosen for the tissue microarray. Scarred, myxoid,
and hypocellular areas were avoided. Two tissue cores with a
diameter of 0.6-mm were taken from each block and inserted into the
blank tissue microarray block. The whole set of tissue array
contained two chips (A and B), in which chip A had 78 NSCLCs and 3
benign controls and chip B contained 76 NSCLCs and 11 benign
controls.
The tissue microarrays were deparaffinized in xylene
and ethanol. Antigen retrieval was done by heating the slides in an
autoclave at 120°C for 3 min in citric acid buffer. The primary
antibody was mouse monoclonal anti-TRA16 (22) (1:200 dilution in PBS) or rabbit
monoclonal anti-ERβ (1:100 dilution in PBS, Dako). The
immunostaining was carried out using the EnVision method (Dako)
according to the manufacturer’s instructions. Brown staining was
considered positive. Normal mouse IgG was used instead of the
primary antibody as a negative control.
Stable transfection of Cos-1 cells and
methyl thiazolyl tetrazolium (MTT) assay
Cos-1 cells, which are TR2-negative and express very
low level of TRA16, were transfected with pBig or pBig-TRA16 using
SuperFect (Qiagen). The cells were then selected using 100 μg/ml
hygromycin B to generate stable clones, which were confirmed by
reporter gene assay. The established stable pBig-TRA16-Cos-1 cells
(5×104) per well were seeded in 12-well plates. After 12
h, the medium was changed to DMEM medium with 10% FBS for another 4
days and then treated with doxycycline (6 μg/ml) or DMSO for
induction of TRA16 expression for 24 h.
For the MTT assay, 2×103 of Cos-1 cells
per well were plated in 96-well plates with RPMI-1640 medium
containing 10% FBS and incubated at 37°C overnight. After 1, 2, 3,
or 4 days, 200 μl of MTT (5 mg/ml, Sigma) was added to each well
for 3-h incubation, then 2 ml of 0.04 M HCl in isopropyl alcohol
was added to each well to stop the reaction. After 5 min of
incubation at room temperature, the absorbance was read at 570
nm.
Transient transfection and mammalian two
hybrid assays
Cos-1 cells (2.5×104) per well were
plated in 12-well plates. The next day, the medium was changed to a
medium containing 10% charcoal-stripped serum to deprive the cells
of steroid hormones. The cells were then transfected the following
day. Transfections were performed using SuperFect (Qiagen)
according to the manufacturer's instructions. The cells were
transiently cotransfected with reporter plasmids pERE-TK-Luc (a
gift from Dr M. Nichols at University of Pittsburgh, Pittsburgh,
PA) and ERβ with or without TRA16. After 24 h, the cells were
incubated with 10 nM estrogen or androgen if necessary for 24 h and
then harvested using lysis buffer (Promega) to detect the
interaction between TRA16 and ERβ.
To further prove the interaction between TRA16 and
ERβ, the Cos-1 cells were transiently co-transfected with reporter
plasmid pG5-Luc and each of GAL4DBD, VP16, VP16-TRA16, and GAL4-ERβ
in mammalian two hybrid system. As negative controls, the Cos-1
cells were transiently cotransfected with PPRE-Luc, MMTV-Luc,
PSA-Luc, ARE-Luc, and the pRL-CMV (Promega, Madison, WI) to confirm
the specifity of the interaction between TRA16 and ERβ.
To assess the interaction between TRA16 and TR2, we
adopted three approaches. First, we transiently co-transfected TR2
and TRA16 or TR2 alone into Cos-1 cells to measure the inhibition
of TR2 transcriptional activity by TRA16. Second, pBig-TRA16-Cos-1
and pBig-Cos-1 stable cells were transiently co-transfected with
HCR-1-Luc reporter and TR2 for 24 h, then transfected cells were
treated with doxycycline to induce TRA16 expression, and measure
the TR2 activity. Third, we transiently co-transfected the reporter
plasmid pG5-Luc and each of GAL4DBD, VP16, VP16-TR2, and GAL4-TRA16
in mammalian two-hybrid system to further prove the interaction
between TRA16 and TR2. The luciferase activity was measured using
the Dual-Luciferase System (Promega) by TD 20/20 luminometer.
Values of the luciferase activity were corrected for protein
concentration and presented as the mean ± SD of three independent
experiments.
Immunoprecipitation assay
Cos-1 cells were transiently transfected with TR2
and either TRA16 or pcDNA4 control for 48 h and then harvested and
dissolved in lysis buffer (1% Nonidet P-40, 10% glycerol, 135 mM
NaCl, 40 mM Tris, pH 7.4, 1 mM phenylmethylsulfonyl fluoride, 1 mM
dithiothreitol, and 1X protease inhibitor cocktail Roche). Cell
lysates containing 500 μg of proteins were precleared with 20 μl of
protein A/G Plus-agarose and 1.0 μg of normal mouse IgG (Santa Cruz
Biotechnology) for 30 min. The supernatant was then mixed with a
1:100 dilution of mouse TR2 antibody at 4°C for 2 h, followed by
adding protein A/G Plus-agarose and incubated for another 2 h.
Immunoprecipitates obtained by spinning down protein A/G
Plus-agarose were washed with PBS for three times and separated on
SDS-8% PAGE. After transferring to the membrane, the protein was
detected by anti-His-tag or anti-TR2 antibodies.
Immunocytofluorescence assay
Cos-1 cells transiently co-transfected with TR2 and
TRA16 were seeded on two-well Lab Tek Chamber slides (Nalge) for 48
h. Immunostaining was performed as described previously (18) by incubating with mouse anti-TRA16
monoclonal antibody and/or rabbit anti-TR2 monoclonal antibody, and
then followed by incubating with either fluorescein-conjugated goat
anti-mouse or anti-rabbit antibodies. Coverslips were fixed on the
glass slides with a drop of DAPI to stain the nucleus. The slides
were observed under 400-fold magnification of a fluorescence
microscope or confocal fluorescence microscope.
Data analysis
SPSS v11.0 software was used to perform statistical
analysis. Chi-square test was used to analyze the assocation
between TRA16 expression and different clinical factors in NSCLC
patients. Univariate and multivariate Cox regression hazards models
were used to analyze the correlation of individual factors with
overall survival of NSCLC patients. Log-rank test and Kaplan-Meier
survival analysis were used to examine the effect of individual
factors on prognosis of NSCLC patients. Values of P<0.05 were
considered significant.
Results
TRA16 is highly expressed in non-small
cell lung cancer
We previously found that TRA16 was highly expressed
in lung adenocarcinoma cell line H1299 (18). To investigate the expression of
TRA16 in NSCLC tissues, we performed tissue microarrays with 154
NSCLCs, in which 134 NSCLC samples were qualified for data analysis
(Table II). The results of TRA16
tissue microarray and its expression level and distribution in
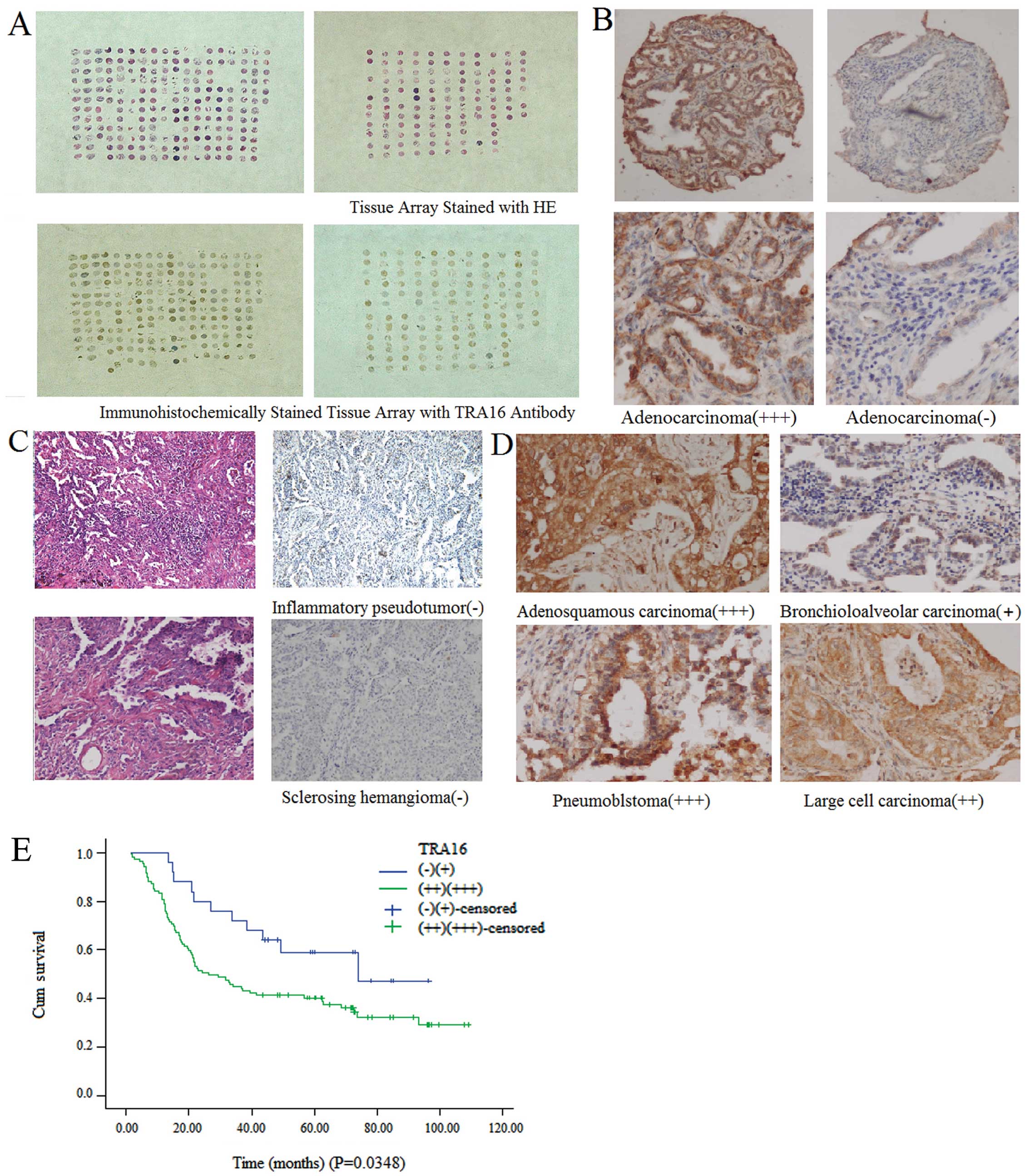
different NSCLC cell types are exemplified in Fig. 1A-D. We found that all the control
samples were TRA16-negative, but 120 lung cancer samples were
TRA16-positive (+, ++, or +++) with the positive rate of 89.55%
(120/134) (P<0.001). When categorizing the samples with the
staining intensity of ++ or +++ as strong positive, there were 105
strong positive samples accounting for 78.36% (105/143) of the
NSCLCs. The 105 samples included 55 adenocarcinoma (83.3%, 55/66),
43 squamous carcinoma (71.7%, 43/60). No correlation of TRA16
expression with NSCLC cell types was observed (P=0.230). The
Chi-square test indicated that the TRA16 expression was correlated
with TNM stage (P=0.016).
| Table IIThe expression level of TRA16 in
human lung clinical samples. |
Table II
The expression level of TRA16 in
human lung clinical samples.
| TRA16
expression | ++/+++ (%) | −/+ | Total | P |
|---|
| Control | 0 (0) | 14 | 14 | <0.001 |
| Benign
disease | 0 (0) | 12 | 12 | |
| Inflammatory
pseudotumor | 0 (0) | 4 | 4 | |
| Tuberculosis | 0 (0) | 4 | 4 | |
| Sclerotic
hemangioma | 0 (0) | 2 | 2 | |
| Harmatoma | 0 (0) | 2 | 2 | |
| Normal | 0 (0) | 2 | 2 | |
| Lung cancer cell
type | 105 (78.36) | 29 | 134 | 0.230 |
| Squmous cell | 43 (71.67) | 17 | 60 | |
|
Adenocarcinoma | 55 (83.33) | 11 | 66 | |
| Others | 7 (87.50) | 1 | 8 | |
| TNM stage | 128 (83.12) | 26 | 154 | 0.016 |
| I | 39 (72.22) | 15 | 54 | |
| II | 46 (85.18) | 8 | 54 | |
| IIIA | 43 (93.48) | 3 | 46 | |
| ER expression | 56 (78.87) | 15 | 71 |
<0.001 |
| ++/+++ | 34 (97.14) | 1 | 35 | |
| −/+ | 22 (61.11) | 14 | 36 | |
TRA16 expression is correlated with
prognosis of NSCLC patients
An univariate Cox regression analysis of
clinicopathologic parameters and TRA16 expression showed that TNM
stage, tumor size, lymph node metastasis, tumor thrombus, cell
differentiation, and TRA16 expression were significantly associated
with overall survival of NSCLC patients (Table III). The multivariate Cox
regression analysis showed that lymph node metastasis, tumor
thrombus formation, poor cell differentiation, and high expression
of TRA16 were independent prognostic factors in NSCLC (Table III). The Kaplan-Meier analysis
showed that the patients with higher TRA16 expression had
significantly shorter overall survival than those with lower TRA16
expression (P=0.0348) (Fig. 1E),
similarly to the standard clinical prognostic factors of TNM stage,
lymph node metastasis, and tumor differentiation.
| Table IIIUnivariate and multivariate Cox
regression analysis of clinicopathological and genetic factors for
prognosis in NSCLC patients. |
Table III
Univariate and multivariate Cox
regression analysis of clinicopathological and genetic factors for
prognosis in NSCLC patients.
| Clinicopathological
and genetic factors | Patient no. | Median OS
(month) | P-value |
|---|
|
|---|
| Univariate
analysis | Multivariate
analysis |
|---|
| Gender | | | 0.2372 | |
| Male | 113 | 37 | | |
| Female | 41 | 63 | | |
| Age | | | 0.8500 | |
| ≤60 | 68 | 39 | | |
| >60 | 86 | 34 | | |
| Smoking
history | | | 0.7769 | |
| No | 92 | 37 | | |
| Yes | 62 | 39 | | |
| Histopathological
classification | | | 0.6300 | |
| Squamous cell | 73 | 38 | | |
| Adenocacinona | 70 | 37 | | |
| Others | 11 | 41 | | |
| TNM stage | | | 0.0000 | |
| I | 54 | 93 | | |
| II | 54 | 42 | | |
| IIIA | 46 | 17 | | |
| Tumor size | | | 0.0093 | |
| T1 | 18 | 93 | | |
| T2 | 62 | 63 | | |
| T3 | 74 | 23 | | |
| Lymph node
metastasis | | | 0.0000 | 0.007 |
| No | 105 | 73 | | |
| Yes | 49 | 21 | | |
| Tumor thrombus in
vana | | | 0.0177 | 0.007 |
| No | 131 | 49 | | |
| Yes | 23 | 21 | | |
| Cell
differentiation | | | 0.0151 | 0.000 |
|
Well-differentiated | 39 | 93 | | |
|
Moderately-differentiated | 46 | 42 | | |
|
Poorly-differentiated | 66 | 22 | | |
| TRA16
expression | | | 0.0348 | 0.020 |
| (−) (+) | 25 | 65 | | |
| (++) (+++) | 109 | 26 | | |
TRA16 expression is correlated with ERβ
expression in NSCLC tissues
Our previous studies demonstrated that TRA16
inhibited the function of TR4 in lung cancer cell line H1299 and
resultantly relieved TR4-repressed ERβ activity (18). To further determine the relationship
between TRA16 and ERβ in NSCLC, we analyzed the expressions of
TRA16 and ERβ in 71 NSCLC tissues and 2 benign samples using the
tissue microarrays. We found that among the 71 NSCLC tumors, 35
cases were ERβ-positive with 22 cases (+), 9 cases (++), and 4
cases (+++), while 36 cases and two benign controls were
ERβ-negative. When using the TRA16 expression to stratify the 71
cases into no or low (+) level group (n=15) and high level (++ −
+++) group (n=56) and then analyzing the difference of the ERβ
expression between the two groups, we found that the ERβ expression
in the high level group of TRA16 was significantly higher than that
in the no or low level group, while only one of 15 samples in the
no or low level group of TRA16 showed ERβ-positive (P<0.001)
(Table II), suggesting their
functional association.
TRA16 enhances cell proliferation in
Cos-1 cells
The increased expression of TRA16 in NSCLC indicated
that TRA16 could play an important role in lung cancer development.
Thus, we performed the MTT assay to study the effect of TRA16 on
cell growth by transfecting doxycycline-inducible TRA16 in pBig
vector into Cos-1 cells. We found that the doxycycline-treated
group had a significantly higher cell growth than the DMSO-treated
control group.
Interaction of TRA16 and ERβ enhances ERβ
activity
To further explore the functional association
between TRA16 and ERβ, we performed several reporter gene assays by
transient transfection of different combinations of plasmids into
Cos-1 cells (Fig. 2). We found that
when transiently co-transfecting ERβ response element (ERE)-Luc
reporter and ERβ with or without TRA16 into Cos-1 cells, after
adding 10 nM of estradiol (E2) to stimulate ERβ
activity, TRA16 significantly enhanced the ERβ activity in Cos-1
cells compared with the ERβ alone (Fig.
2A). Further, in the mammalian two-hybrid assay by transfecting
the reporter plasmid pG5-Luc and each of GAL4DBD, VP16, VP16-TRA16,
and GAL4-ERβ in various combinations as indicated into Cos-1 cells,
we found that after the stimulation of E2, the
co-presence of TRA16 and ERβ significantly enhanced the ERβ
activity in Cos-1 cells (Fig. 2B),
indicating that TRA16 interacted with ERβ.
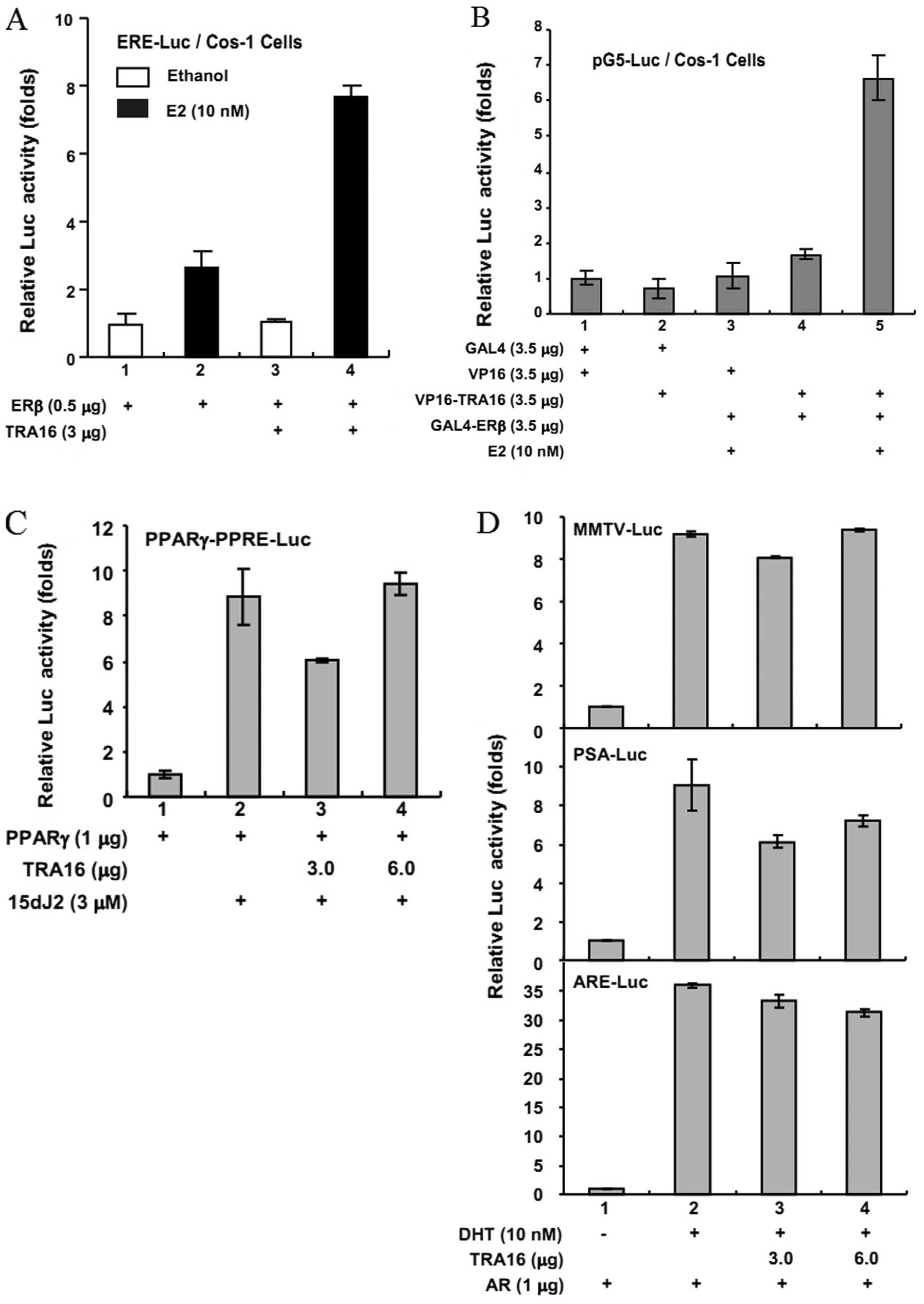 | Figure 2The interaction of TRA16 and ERβ on
the ERβ activity in mammalian Cos-1 cells. (A) Cos-1 cells were
transiently co-transfected with 3 μg of ERE-Luc reporter, 0.5 μg of
ERβ, and 3 μg of TRA16 expression plasmids for 24 h, then treated
with either 10 nM E2 or ethanol vehicle for the other 24
h to measure the luciferase activities. (B) Cos-1 cells were
transiently co-transfected with 3 μg of reporter plasmid pG5-Luc
and 3.5 μg each of GAL4DBD, VP16, VP16-TRA16, and GAL4-ERβ in
various combinations, then treated with either 10 nM E2
or ethanol vehicle for 24 h to measure luciferase activities. (C)
Cos-1 cells were transiently co-transfected with 3 μg of reporter
plasmid PPRE-LUC, 1 μg of PPARγ, and different doses (3.0 and 6.0
μg) of TRA16, then treated with 3 μM of 15dJ2 (an activator of
PPARγ) for another 24 h to measure luciferase activities. (D) Cos-1
cells were transiently co-transfected with 3 μg of different
reporter plasmids (MMTV-Luc, PSA-Luc, or ARE-Luc), and both AR and
TRA16 (ratio: 1:3 and 1:6) for 24 h, and then treated with 10 nM of
DHT to measure luciferase activities. |
To clarify the specificity of the association of
TRA16 and ERβ, similar but negative control experiments were
performed by co-transfecting TRA16 and PPAR or androgen receptor
(AR) in Cos-1 cells. As shown in Fig.
2C and D, TRA16 had little effect on activation of both PPAR
and AR.
TRA16 interacts with TR2 enhancing ERβ
activity
We previously demonstrated that TRA16 bound to TR4
and blocked TR4 dimerization and thus inhibited TR4 activity in
lung cancer cells (18). TR2 and
TR4 are highly homologous and TR2 can suppress ERβ-mediated cell
growth in lung cancer cells in vitro(19,20).
To assess a potential relationship between TRA16 and TR2, we
performed similar reporter gene assays as described above. We found
that when HCR-1-Luc reporter gene and TR2 with or without TRA16
were co-transfected into Cos-1 cells, TRA16 strongly inhibited TR2
transcriptional activity in Cos-1 cells (Fig. 3A). Similarly, the
doxycycline-inducible assay using the pBig-Cos-1 and
pBig-TRA16-Cos-1 cell system showed that after treatment of 6 μg/ml
doxycycline, TR2 activity was significantly lower in
pBig-TRA16-Cos-1 cells than that in pBig-Cos-1 cells (Fig. 3B). Moreover, the mammalian
two-hybrid assay demonstrated that the co-expression of TRA16 and
TR2 significantly enhanced the reporter Luc activity compared with
TR2 or TRA16 alone (Fig. 3C),
suggesting the association of TRA16 and TR2.
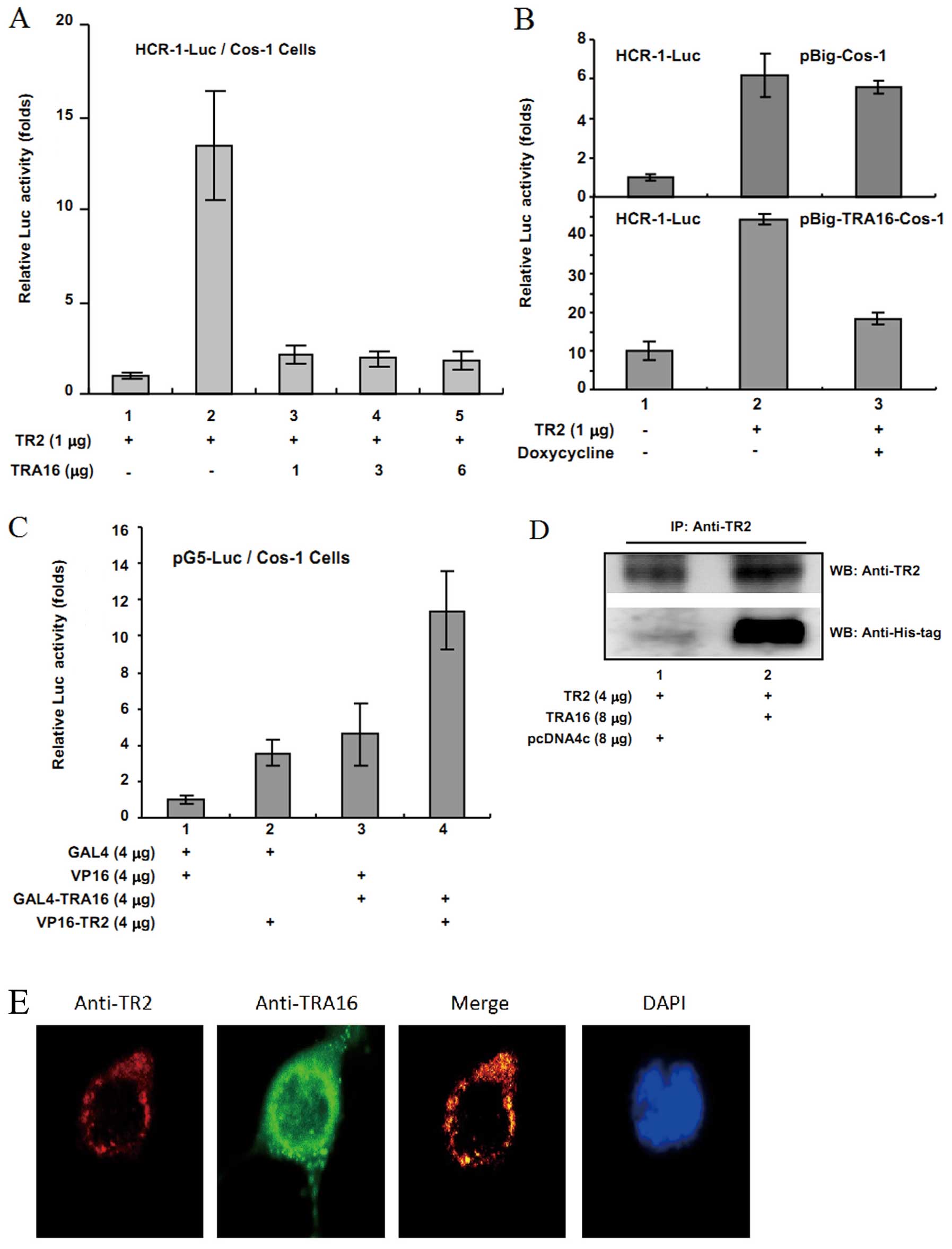 | Figure 3Interaction between TRA16 and TR2
detected by different methods. (A) Cos-1 cells were transiently
co-transfected with 3 μg of HCR-1-Luc reporter, 1 μg of TR2, and
increased amount of TRA16 expression plasmid, and 24 h after
transfection, the luciferase activity was measured. (B)
pBig-TRA16-Cos-1 and pBig-Cos-1 stable cells were transiently
co-transfected with 3 μg of HCR-1-Luc reporter, and 1 μg of TR2 for
24 h, then treated with either 6 μg/ml doxycycline or DMSO vehicle
for another 24 h, and the luciferase activity was measured. (C)
Cos-1 cells were transiently co-transfected with 4 μg of reporter
plasmid pG5-Luc and 4 μg each of GAL4DBD, VP16, VP16-TR2, and
GAL4-TRA16 in various combinations as indicated, and then cultured
for 24 h to measure the luciferase activities. (D) Cos-1 cells were
transiently co-transfected with 4 μg TR2 and 8 μg either TRA16 or
pcDNA4c for 48 h, then the cell lysates were harvested, precleared
and added with anti-TR2 antibody and protein G-agarose beads. The
immunoprecipitates obtained were subjected to 8% SDS-PAGE to detect
the pull-down protein. (E) Cos-1 cells were transiently
co-transfected with TR2 and TRA16, then stained with anti-TRA16 or
anti-TR2 antibody, and then corresponding second antibodies. DAPI
was added to stain the nucleus. The cells were then observed under
the fluorescent microscopy. The green signal represents TRA16, the
red signal represents TR2, the yellow signal represents
co-localization of TRA16 and TR2, and the blue signal represents
the DAPI-stained nucleus. |
Furthermore, the immunoprecipitation assay showed
that TRA16 directly interacted with TR2 (Fig. 3D), and TRA16 and TR2 were
co-localized in the nuclear membrane by immunocytofluorescence
assay (Fig. 3E). Taken together,
the findings suggest that TRA16 interact with TR2 to release
TR2-inhbitory role in ERβ activity in addition to its direct
stimulation of ERβ signaling.
Discussion
Our study reports that TRA16 is highly expressed in
NSCLCs and its expression level is negatively correlated with the
overall survival of NSCLC patients, and it enhances ERβ signaling
pathway by direct stimulation of ERβ activity and through
suppressing TR2-inhibitory effect on ERβ for cancer cell growth,
suggesting TRA16 is a promising diagnostic and prognostic biomarker
and potential target in NSCLC.
Tumor biomarkers, such as CEA, CA125, NSE,
CYFRA21-1, and p53, are currently used in cancer diagnosis and
prognosis, but few of them are ideal for NSCLC because of low
sensitivity and specificity (23–25).
For stage I NSCLC patients, the five-year survival rate exceeds
60%, but the patients with metastatic spread to regional or distant
sites, who account for at least three quarters of lung cancer
patients at the time of diagnosis, have only approximately 15%
five-year survival rate (26).
Therefore, if the NSCLC can be diagnosed in early stages, the
patient survival rate and time could be significantly improved.
This study demonstrates that TRA16 is highly expressed in
approximately 90% of NSCLCs, in which over 71% of stage I NSCLC
overexpress TRA16, which is not expressed in lung benign diseases
and normal lung tissues, indicating its potential for NSCLC
diagnosis, especially for early stage NSCLC.
Furthermore, our findings of the correlation of
increased expression of TRA16 with increased malignancy of NSCLC
demonstrate that TRA16 could be a valuable biomarker to monitor the
progression of the tumor, and provide physicians with useful
information to evaluate the severity of the disease and to choose
proper treatments for NSCLC patients.
For a long time, researchers have proposed that
sexual hormone, such as progesterone and estrogen, may play
important roles in lung cancer carcinogenesis and development
(27,28), which was supported by recent
clinical findings of abnormal expression of ERβ (2–6) and
increased intratumoral estrodial concentration in NSCLC tissues
(8). There are several possible
mechanisms that may be involved in the abnormal activation of
hormone related signaling transduction pathways. First, the high
serum hormone level or high hormone receptor expression level can
up-regulate downstream signaling pathways to stimulate target cell
proliferation (8). Second, the
mutation of hormone receptors may increase the ligand binding
affinity or be activated by other ligands (29,30).
Third, the abnormality of hormone receptor co-factors can active
the downstream signaling transduction pathway, and then promote
oncogenesis (31).
TRA16 belongs to the co-factors of nuclear receptors
(NRs) with which it interacts to affect the transcriptional
activity of NRs. Our findings that the concordant expressions of
TRA16 and ERβ in NSCLC and their interaction suggest that TRA16 may
serve as an upstream positive regulator of ERβ in NSCLC. Moreover,
we have observed the interaction between TRA16 and TR2. Therefore,
possible interactions among TRA16, TR2, TR4, and ERβ could be
proposed. TR2 and TR4 inhibit ERβ signaling pathways. In addition
to its direct stimulation of ERβ, TRA16 also directly interacts
with TR2 and TR4 that results in the release of ERβ from TR2 and
TR4 inhibition, which consequently leads to the tumorigenesis.
Since it is highly and specifically expressed in tumor tissues,
TRA16 may be used as a tumor-specific therapeutic target for lung
cancer treatments in the form of anti-TRA16 antibody or TRA16
shRNA.
Therefore, there are still many questions to be
answered, such as what are the upstream regulators of TRA16 and its
interaction sites with ERβ and TR2, and whether the TRA16
expression can be detected in circulating cancer cells in patient
blood that could be used to easily screen and diagnose patients
with NSCLC, all of which will be investigated in future
studies.
Acknowledgements
This study was supported by National Institutes of
Health Grant DK56784, a George Whipple Professorship Endowment,
Beijing Natural Science Foundation, and National Natural Science
Foundation. We thank Dr P. Monaghan-Nichols from University of
Pittsburgh School of Medicine for kindly providing the reporter
plasmids, Dr Wanzhong Zou from Peking University Health Science
Center of Pathology, Mrs. E. Jie, and Mrs. Hong Du from Peking
University Cancer Hospital for their excellent technical
assistance.
Abbreviations:
|
ERβ
|
estrogen receptor β
|
|
TR2
|
testicular orphan nuclear receptor
2
|
|
TRA16
|
TR4-associated protein 16
|
|
NSCLC
|
non-small cell lung cancer
|
|
OS
|
overall survival
|
|
E2
|
estradiol
|
|
NRs
|
nuclear receptors
|
|
LI
|
labeling index
|
|
TR4RE
|
TR4 response element
|
|
FBS
|
fetal bovine serum
|
|
MTT
|
methyl thiazolyl tetrazolium
|
References
|
1
|
Siegel R, Ward E, Brawley O and Jemal A:
Global Cancer Statistics, 2011 The impact of eliminating
socioeconomic and racial disparities on premature cancer deaths. CA
Cancer J Clin. 61:212–236. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kawai H, Ishii A, Washiya K, Konno T, Kon
H, Yamaya C, Ono I, Minamiya Y and Ogawa J: Estrogen receptor α and
β are prognostic factors in non-small cell lung cancer. Clin Cancer
Res. 11:5084–5089. 2005.
|
|
3
|
Schwartz AG, Prysak GM, Murphy V, Lonardo
F, Pass H, Schwartz J and Brooks S: Nuclear estrogen receptor β in
lung cancer: expression and survival differences by sex. Clin
Cancer Res. 11:7280–7287. 2005.
|
|
4
|
Siegfried JM, Hershberger PA and Stabile
LP: Estrogen receptor signaling in lung cancer. Semin Oncol.
36:524–534. 2005. View Article : Google Scholar
|
|
5
|
Mollerup S, Jørgensen K, Berge G and
Haugen A: Expression of estrogen receptors α and β in human lung
tissue and cell lines. Lung Cancer. 37:153–159. 2002.
|
|
6
|
Omoto Y, Kobayashi Y, Nishida K, Tsuchiya
E, Eguchi H, Nakagawa K, Ishikawa Y, Yamori T, Iwase H, Fujii Y,
Warner M, Gustafsson JA and Hayashi S: Expression, function, and
clinical implications of the estrogen receptor β in human lung
cancers. Biochem Biophys Res Commun. 285:340–347. 2001.
|
|
7
|
Patrone C, Cassel T, Pettersscon K, Piao
Y, Piao Y, Cheng G, Ciana P, Maggi A, Warner M, Gustafsson J and
Nord M: Regulation of postnatal lung development and homeostasis by
estrogen receptor β. Mol Cell Biol. 23:8542–8552. 2003.
|
|
8
|
Niikawa H, Suzuki T, Miki Y, Suzuki S,
Nagasaki S, Akahira J, Honma S, Evans DB, Hayashi S, Kondo T and
Sasano H: Intratumoral estrogens and estrogen receptors in human
non small cell lung carcinoma. Clin Cancer Res. 14:4417–4426. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mah V, Seligson D, Li A, Marquez D,
Wistuba I, Elshimali Y, Fishbein M, Chia D, Pietras R and Goodglick
L: Aromatase expression predicts survival in women with early-stage
non-small cell lung cancer. Cancer Res. 21:10484–10490. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Weinberg O, Marquez-Garban D, Fishbein M,
Goodglick L, Garban H, Dubinett S and Pietras R: Aromatase
inhibitors in human lung cancer therapy. Cancer Res.
24:11287–11291. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Stabile L, Gaither Davis A, Gubish C,
Hopkins T, Luketich J, Christie N, Finkelstein S and Siegfried J:
Human non-small cell lung tumors and cells derived from normal lung
express both estrogen receptor α and β and show biological
responses to estrogen. Cancer Res. 62:2141–2150. 2002.
|
|
12
|
Chang C, Kokontis J and Liao ST: Molecular
cloning of human and rat complementary DNA encoding androgen
receptors. Science. 240:324–326. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Evans RM: The steroid and thyroid hormone
receptor superfamily. Science. 240:889–895. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Beato M: Gene regulation by steroid
hormones. Cell. 56:335–344. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang LH, Tsai SY, Cook RG, Beattie WG,
Tsai MJ and O'Malley BW: Coup transcription factor is a member of
the steroid receptor superfamily. Nature. 340:163–166. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tsai MJ and O'Malley BW: Molecular
mechanisms of action of steroid/thyroid receptor superfamily
members. Annu Rev Biochem. 63:451–486. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shyr CR, Hu YC, Kim E and Chang C:
Modulation of estrogen receptor-mediated transactivation by orphan
receptor TR4 in MCF-7 cells. J Biol Chem. 277:14622–14628. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang Y, Wang X, Dong T, Kim E, Lin WJ and
Chang C: Identification of a novel testicular orphan
receptor-4(TR4)-associated protein as repressor for the selective
suppression of TR4-mediated transactivation. J Biol Chem.
278:7709–7717. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shyr CR, Kang HY, Tsai MY, Liu NC, Ku PY,
Huang KE and Chang C: Roles of testicular orphan nuclear receptors
2 and 4 in early embryonic development and embryonic stem cells.
Endocrinology. 150:2454–2462. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hu YC, Shyr CR, Che W, Mux M, Kim E and
Chang C: Suppression of estrogen receptor-mediated transcription
and cell growth by interaction with TR2 orphan receptor. J Biol
Chem. 277:33571–33579. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kononen J, Bubendorf L, Kallionimeni A,
Barlund M, Schraml P, Leighton S, Torhorst J, Mihatsch MJ, Sauter G
and Kallioniemi OP: Tissue microarrays for high-throughput
molecular profiling of tumor specimens. Nat Med. 4:844–847. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zheng QF, Dong B, Sun Y, Du H, Xiong HC,
Wu N, Chen JF, Sun L, Ao J, Li JY, Ji JF and Yang Y: Expression of
TR4-associated protein in non-small cell lung cancer. Beijing
Daxuexue Bao. 39:472–475. 2007.(in Chinese).
|
|
23
|
Schneider J, Velcovsky HG, Morr H, Katz N,
Neu K and Eigenbrodt E: Comparison of the tumor markers tumor
M2-PK, CEA, CYFRA 21-1, NSE and SCC in the diagnosis of lung
cancer. Anticancer Res. 20:5053–5058. 2000.PubMed/NCBI
|
|
24
|
Salgia R, Harpole D, Herndon JE II, Pisick
E, Elias A and Skarin AT: Role of serum tumor markers CA 125 and
CEA in non-small cell lung cancer. Anticancer Res. 21:1241–1246.
2001.PubMed/NCBI
|
|
25
|
Iggo R, Gatter K, Bartek J, Lane D and
Harris AL: Increased expression of mutant forms of p53 oncogene in
primary lung cancer. Lancet. 335:675–679. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mulshine JL and Sullivan DC: Clinical
practice. Lung cancer screening. N Engl J Med. 352:2714–2720. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ishibashi H, Suzuki T, Suzuki S, Niikawa
H, Lu L, Miki Y, Moriya T, Hayashi S, Handa M, Kondo T and Sasano
H: Progesterone receptor in non-small cell lung cancer - A potent
prognostic factor and possible target for endocrine therapy. Cancer
Res. 65:6450–6458. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Beattie CW, Hansen NW and Thomas PA:
Steroid receptors in human lung cancer. Cancer Res. 45:4206–4214.
1985.PubMed/NCBI
|
|
29
|
Stabie L and Siegfried J: Estrogen
receptor pathways in lung cancer. Curr Oncol Rep. 6:259–267. 2004.
View Article : Google Scholar
|
|
30
|
Villa E, Colantoni A, Grottola A, Ferretti
I, Buttafoco P, Bertani H, De Maria ND and Manenti F: Variant
estrogen receptors and their role in liver disease. Mol Cell
Endocrinol. 193:65–69. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hershberger P, Vasquez A, Kanterewicz B,
Land S, Siegfried J and Nichols M: Regulation of endogenous gene
expression in human non-small cell lung cancer cells by estrogen
receptor ligands. Cancer Res. 65:1598–1605. 2005. View Article : Google Scholar : PubMed/NCBI
|