Introduction
Aside from tumor cell proliferation, expansion and
metastasis, angiogenesis is one of the essential steps in tumor
progression. When the diameter is over 0.5 mm, the tumor stimulates
blood vessel growth to cover the need for nutrition and oxygen,
known as the process of angiogenesis (1). The correlation between angiogenesis
and tumor metastasis has been described in two points indicating
that angiogenesis plays a key role in tumor metastasis: first, the
increased vascularity of the primary tumor always leads to
increased number of metastatic colonies; second, increased vascular
density is indicative of increased metastasis colony size (2). Thus, anti-angiogenesis is one of the
crucial targets of tumor therapy and anti-angiogenic therapy has
been used to inhibit cancer growth and metastasis in the clinic
(3). For instance, bevacizumab, the
first anti-angiogenic drug approved by the FDA, is a monoclonal
antibody against vascular endothelial growth factor (VEGF)-A,
inhibiting tumor growth and metastasis (4,5) via
anti-angiogenesis (6). Various
factors induce angiogenesis, including hypoxia. In tumor, due to
the accelerated proliferation of tumor resulting in an abnormal and
chaotic blood supply, blood vessels cannot provide oxygen
adequately or consistently to the whole tumor. Moreover, the
temporal changes in blood flow cause endothelium to suffer hypoxia
in certain regions (7).
Dwarf lilyturf tuber, a traditional Chinese
medicine, is generally used for cardiovascular disease treatment
(8). It has been proved that the
saponin monomer of dwarf lilyturf tuber, DT-13, exerts anticancer
activity (9,10). It has also been reported that some
saponin, such as ginseng saponin, could inhibit angiogenesis
(11,12). Previous studies on cancer cell lines
indicated that DT-13 inhibits MDA-MB-435 cell migration and
adhesion. DT-13 downregulates early growth response gene-1
expression resulting in the reduced excretion of tissue factor
under hypoxia (13). In addition,
DT-13 decreases the excretion and expression of matrix
metallopeptidase 2/9 (MMP-2/9) and reduces the phosphorylation of
p38 (14). However, the effect of
DT-13 on angiogenesis remains unclear. In the present study, we
investigated if DT-13 impacts angiogenesis under normoxia or
hypoxia and we examined the potential mechanisms.
Materials and methods
Cell culture
Human umbilical vein endothelial cells (HUVECs) were
isolated as previously described (15). HUVECs were cultured in complete
medium consisting of M199 medium (Gibco), 20% FBS, 50 ng/ml of
endothelial cell growth supplement (ECGS, Sigma) and 100 ng/ml
heparin. Endothelial cells from third to sixth passages were used
for experiments. The cells were incubated in a humidified
atmosphere of 95% air, 5% CO2 at 37°C. For hypoxic
treatment, we placed 70–80% confluent cells in a tissue culture
incubator in an atmosphere of 1% O2 and 5%
CO2 at 37°C (16).
HUVEC proliferation assay
HUVECs were harvested, washed with phosphate buffer
solution (PBS), re-suspended in complete medium and plated in
96-well plates at the density of 5×103 cells/well,
followed by a 12-h incubation. Cells were then treated with
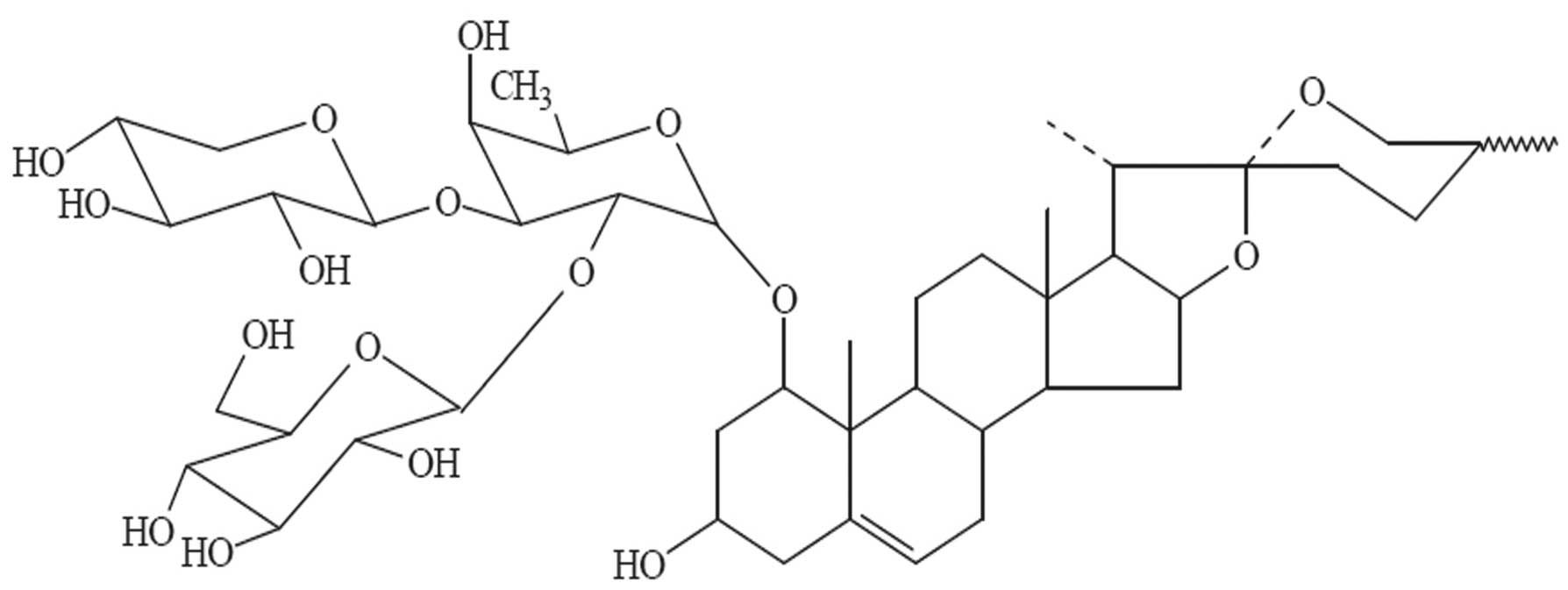
10−8, 10−7 and 10−6 M DT-13
(provided by Professor Bo-Yang Yu) (Fig. 1) for 24 and 48 h under normoxia. The
cell viability was evaluated by the Cell Counting kit (CCK-8,
Dojindo Laboratories), or the medium was changed to ECGS-free
medium (M199 medium with 1% FBS without ECGS). HUVECs were
pretreated with 10−8, 10−7 and
10−6 M DT-13 for 1 h, and were then treated with 10
ng/ml VEGF for 48 h. Following treatment, the cell viability was
evaluated by the Cell Counting kit.
Cell migration assay
The cell migration assay was performed by using a
Transwell chamber migration system with polycarbonate membranes
(8.0 μM pore size), as previously described (17). Briefly, HUVECs were harvested,
re-suspended in serum-free culture medium and (2×104
cells/well) seeded with various concentrations of DT-13 into the
upper chamber. Complete culture medium was added into the lower
chamber. The cell migration system was incubated at 37°C for 8 h,
or, HUVECs were harvested, re-suspended with ECGS-free medium and
plated (2×104 cell/well) in the presence of various
concentrations of DT-13 into the upper chamber. VEGF (10 ng/ml) in
ECGS-free medium was added into the lower chamber. The cell
migration system was incubated at 37°C for 4 h. Following
treatment, cells were stained and counted in three randomly
selected fields.
Tube formation assay
Tube formation assay was performed as previously
described (18). Briefly, 96-well
plates were coated with Matrigel (50 μl/well), and incubated at
37°C for 1 h to solidify Matrigel. HUVECs (1.5×104
cells/well) were incubated with various concentrations of DT-13 at
37°C. After 6 h, tubular structures were quantified by manual
counting of the number of tubes in three randomly selected fields.
When HUVECs were treated with 10 ng/ml of VEGF, the growth factors
reduced Matrigel and ECGS-free medium was preferred, and HUVECs
were incubated under normoxia. If not, basement Matrigel and
complete medium was preferred, and HUVECs were incubated under
hypoxia and normoxia.
Chicken embryo chorioallantoic membrane
(CAM) assay
A small window was created on the broad side of the
5-day-old egg. The filter paper with different doses of DT-13
(final concentration 0.001, 0.01 and 0.1 nmol/egg) was dropped on
the window. The eggs were incubated at 37°C for another 2 days. We
then observed the density and length of vessels toward the CAM face
and then the CAMs were photographed. Eight eggs were used per
group, and the newly formed vessels were counted.
Western blot analysis
Cellular protein extraction and western blot
analysis were performed as previously described (16). Briefly, 40 μg total protein was
fractionated using 10% sodium dodecyl sulfate polyacrylamide gel
electrophoresis (SDS-PAGE) and transferred onto nitrocellulose
membranes under semi-dry conditions. Membranes were probed with
rabbit anti-human p-extracellular signal-regulated kinase (ERK) 1/2
(CST), ERK1/2 (CST), p-AKT (CST), AKT (CST), HIF-1α (Santa Cruz),
and murine anti-human β-actin (Sigma). Horseradish peroxidase (HRP)
linked anti-mouse immunoglobulin G (Sigma) and anti-rabbit
immunoglobulin G (CST) were used as the secondary antibodies.
Protein bands were visualized by enhanced chemiluminescence
reagents (Amersham Pharmacia Biotech).
Enzyme-linked immunosorbent assay
(ELISA)
HUVECs were plated into 60-mm dishes with completed
medium overnight. HUVECs were changed medium to serum and growth
factor supplement-free medium. After 2 h, cells were treated with
various concentrations of DT-13 for 12 h under hypoxia or normoxia.
After treatment, the medium was harvested to detect the
concentration of VEGF by ELISA. The level of VEGF in HUVEC medium
was measured with a VEGF ELISA kit (4A Biotech) according to the
manufacturer’s protocol.
Statistical analysis
The data represent at least three independent
experiments and are expressed as the means ± SEM. For statistical
analysis, the Student’s t-test was used as appropriate.
Results
Effect of DT-13 on HUVEC
proliferation
To identify the cytotoxicity of DT-13, we treated
HUVECs with 0.01, 0.1 and 1 μM DT-13 for 24 and 48 h. DT-13 at 1 μM
inhibited cell proliferation to 71 and 86% of control at 24 and 48
h of treatment, respectively (Fig. 2A
and B). However, 0.01 μM of DT-13 slightly improved HUVEC
proliferation after 48 h of treatment. These results indicate that
DT-13 inhibits HUVEC proliferation at high concentrations but
promotes proliferation at low concentrations.
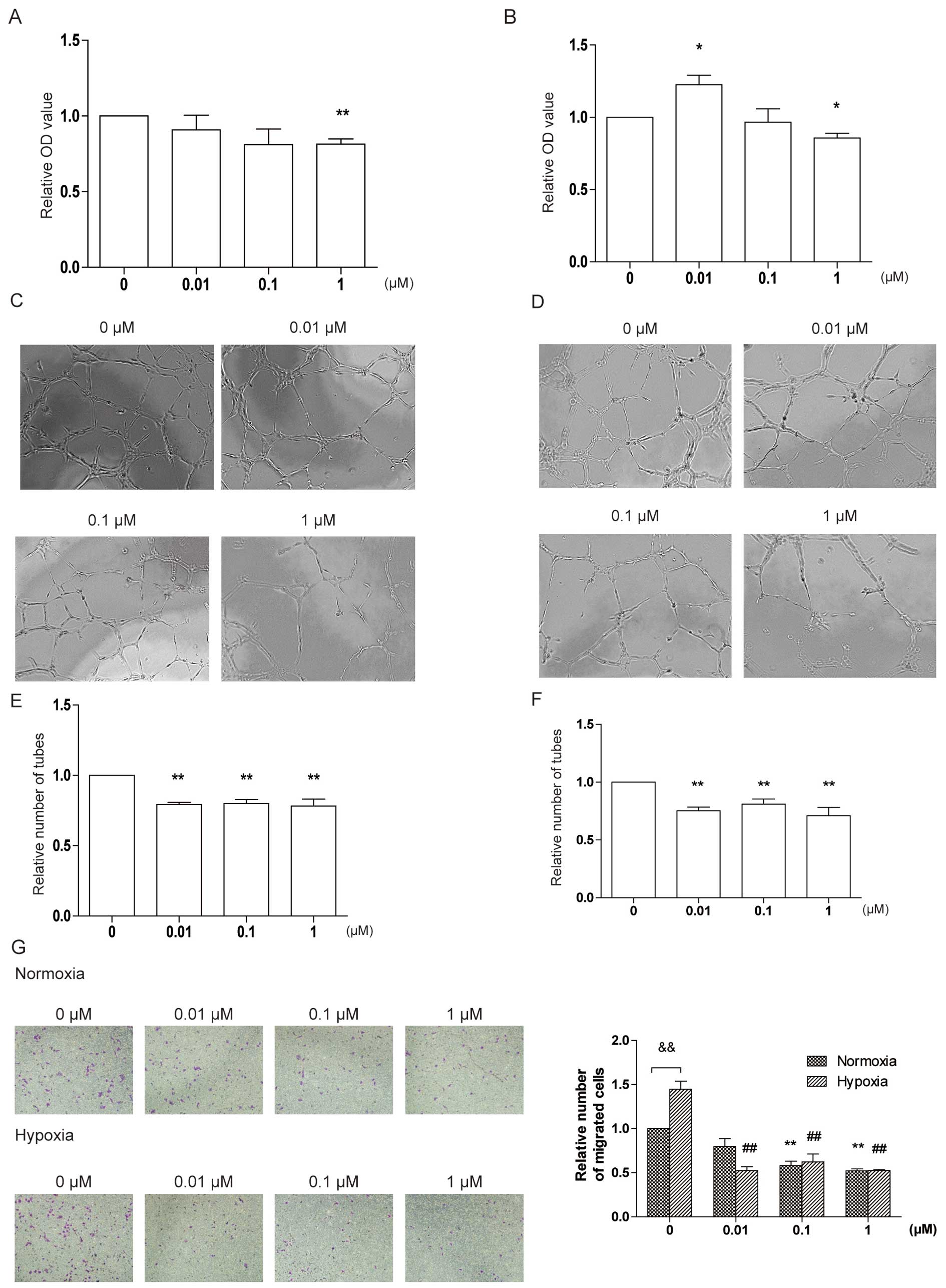 | Figure 2DT-13 inhibits angiogenesis under
normoxia and hypoxia in vitro. (A) The effect of DT-13 on
HUVEC proliferation for 24 h under normoxia. HUVECs were treated
with DT-13 (0.01, 0.1 and 1 μM) or PBS for 24 h under normoxia.
Following treatment, the number of cells was counted by CCK-8.
**p<0.01, statistical significance relative to
vehicle control. (B) The effect of DT-13 on HUVEC proliferation for
48 h under normoxia. HUVECs were treated with DT-13 (0.01, 0.1 and
1 μM) or PBS for 48 h under normoxia. Following treatment, the
number of cells was counted by CCK-8. *p<0.05,
statistical significance relative to vehicle control. (C and E) The
effect of DT-13 on tube formation under normoxia. HUVECs were
plated into 96-well plates pre-coated by Matrigel and treated with
DT-13 (0.01, 0.1 and 1 μM) or PBS. HUVECS were incubated under
normoxia for 6 h. After treatment, the tubes were captured and
counted. **p<0.01, statistical significance relative
to vehicle control. (D and F) The effect of DT-13 on tube formation
under hypoxia and normoxia. HUVECs were plated into 96-well plates
pre-coated by Matrigel and treated with DT-13 (0.01, 0.1 and 1 μM)
or PBS. HUVECS were incubated under hypoxia for 6 h. After
treatment, the tubes were captured and counted.
**p<0.01, statistical significance relative to
vehicle control. (G) The effect of DT-13 on HUVEC migration under
normoxia and hypoxia. HUVECs in serum-free medium with DT-13 (0.01,
0.1 and 1 μM) or PBS were plated into the inner chamber of
Transwell and complete medium was added into the outer chamber. The
Transwell system was incubated under normoxia or hypoxia for 8 h.
After treatment, the cells on the outer side membrane were fixed
and stained by crystal violet. These cells were captured and
counted. **p<0.01, statistical significance relative
to vehicle control under normoxia. ##p<0.01,
statistical significance relative to vehicle control under hypoxia.
&p<0.01, statistical significance relative to
vehicle control under hypoxia. |
DT-13 inhibits tube formation and HUVEC
migration under hypoxia and normoxia
To further investigate the effect of DT-13 on
angiogenesis, we performed tube formation and migration assays
under normoxic and hypoxic conditions, since DT-13 displayed marked
anticancer activity under hypoxia (13). In the tube formation study, HUVECs
were treated with 0.01, 0.1 and 1 μM DT-13 and incubated under
normoxia or hypoxia for 6 h. Under normoxia, different
concentrations of DT-13 showed similar inhibitory effects on tube
formation by 20% compared with control (21, 20 and 21% by 0.01, 0.1
and 1 μM of DT-13 respectively) (Fig.
2C and E). Under hypoxia, the inhibitory activity was 25, 19
and 29% compared with control, respectively (Fig. 2D and F).
In the migration assay, we planted HUVECs into the
upper chamber of Transwell with serum-free medium in the presence
or absence of DT-13, and added complete medium into the lower
chamber. Under normoxia, DT-13 reduced the number of migrated cells
with the inhibitory rate of 20, 40 and 48% at the concentration of
0.01, 0.1 and 1 μM, respectively (Fig.
2G). Under hypoxia, DT-13 performed stronger activity on
inhibiting migration; different concentrations of DT-13 showed
similar inhibitory effects on migration by 40% compared with
control (36, 43 and 36% by 0.01, 0.1 and 1 μM of DT-13
respectively) (Fig. 2G). These
results indicate that DT-13 inhibits tube formation and migration
under normoxia and this effect is stronger under hypoxia.
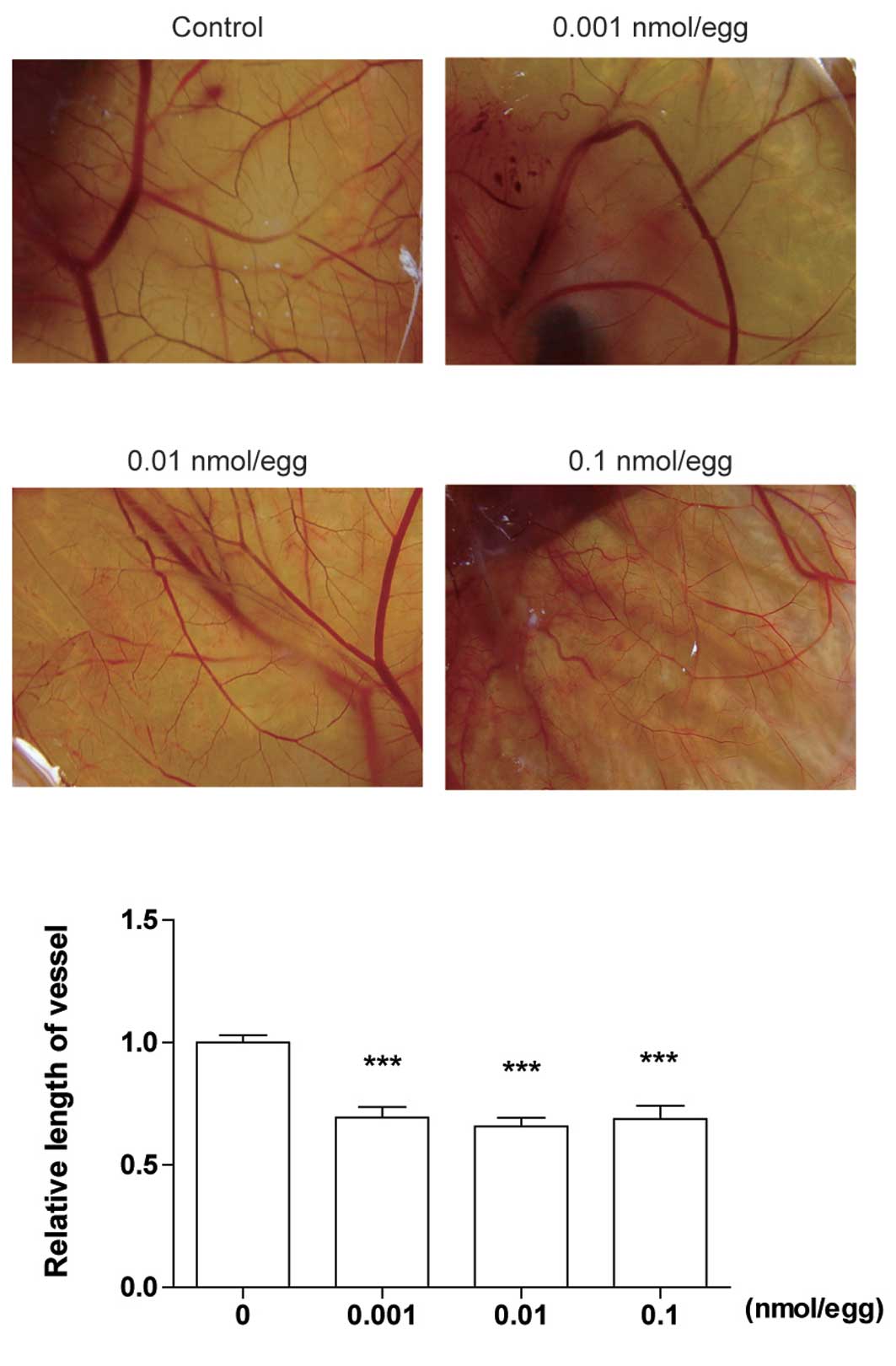
DT-13 inhibits angiogenesis in vivo
To confirm the anti-angiogenesis activity of DT-13
in vivo, we used a CAM model. Five-day-old eggs were treated
with PBS or DT-13 (0.1, 0.01 and 0.001 nmol/egg) for 48 h under
normoxia. The normal branching pattern of blood vessels formed
after 2 days of incubation. As shown in Fig. 3, DT-13 significantly reduced the
relative length of vessels. Different concentrations of DT-13
showed similar inhibitory rates approximately 35% compared with
PBS. These data suggest that DT-13 inhibits angiogenesis in
vivo with a large spectrum of dose.
DT-13 downregulates the level of HIF-1α,
p-ERK, and p-Akt under hypoxia
Hypoxia-inducible factor 1α (HIF-1α) accumulates
under hypoxia and is a crucial factor for angiogenesis (19). Thus, we investigated if DT-13
affects the level of HIF-1α. Hypoxia strongly upregulated HIF-1α
expression compared with normoxia (Fig.
4A). However, DT-13 decreased the amount of HIF-1α both under
normoxia and hypoxia. Under hypoxia, DT-13 at the concentrations of
0.01 and 1 μM significantly downregulated increased HIF-1α induced
by hypoxia. Moreover, DT-13 significantly downregulated ERK1/2 and
Akt phosphorylation in a dose-dependent manner under hypoxia
(Fig. 4A). Under normoxia, DT-13
only slightly reduced the level of p-Akt at 1 μM (Fig. 4A). These data suggest that DT-13
exerts anti-angiogenic effects under hypoxia by inhibiting ERK1/2
and Akt phosphorylation.
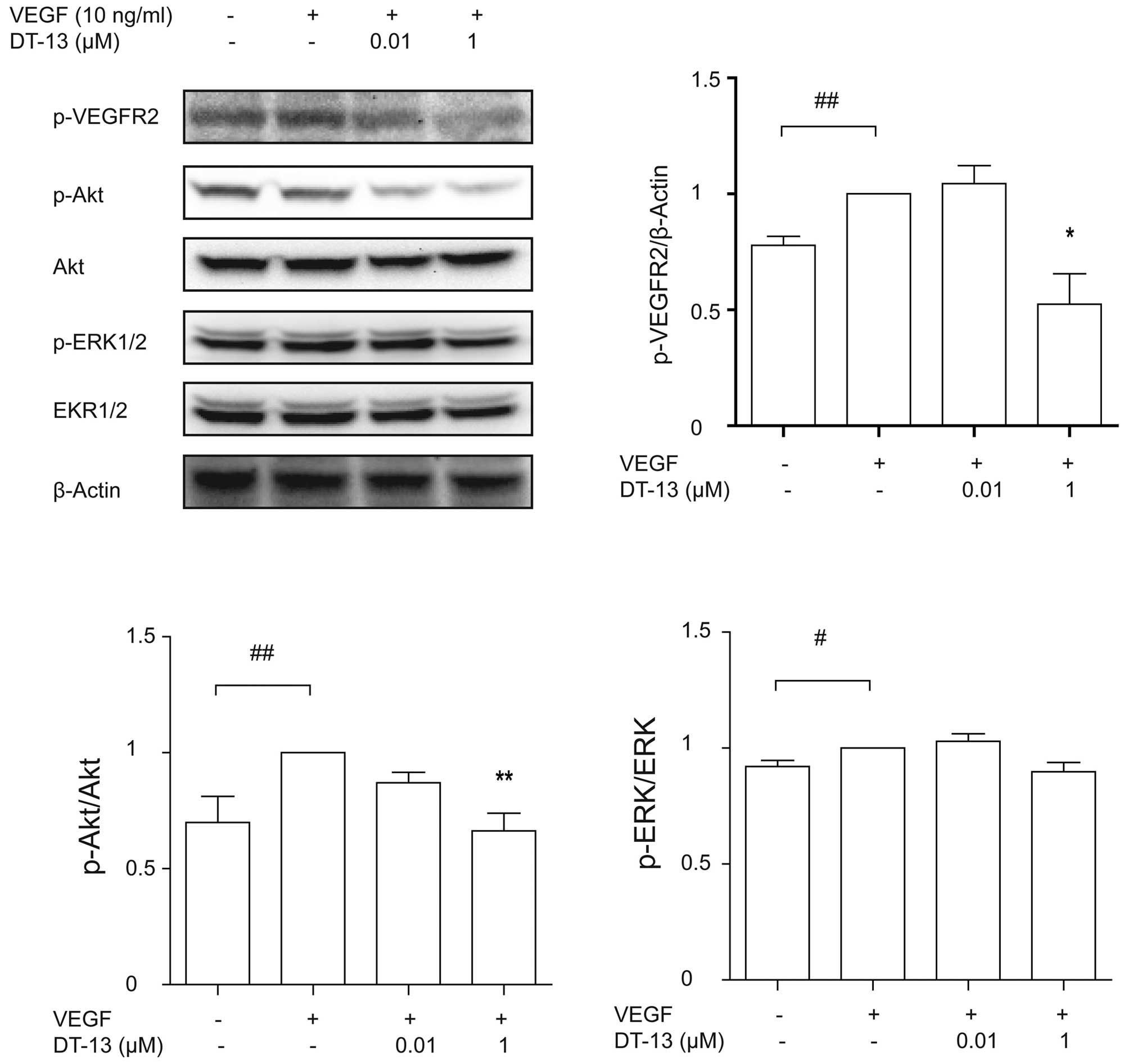
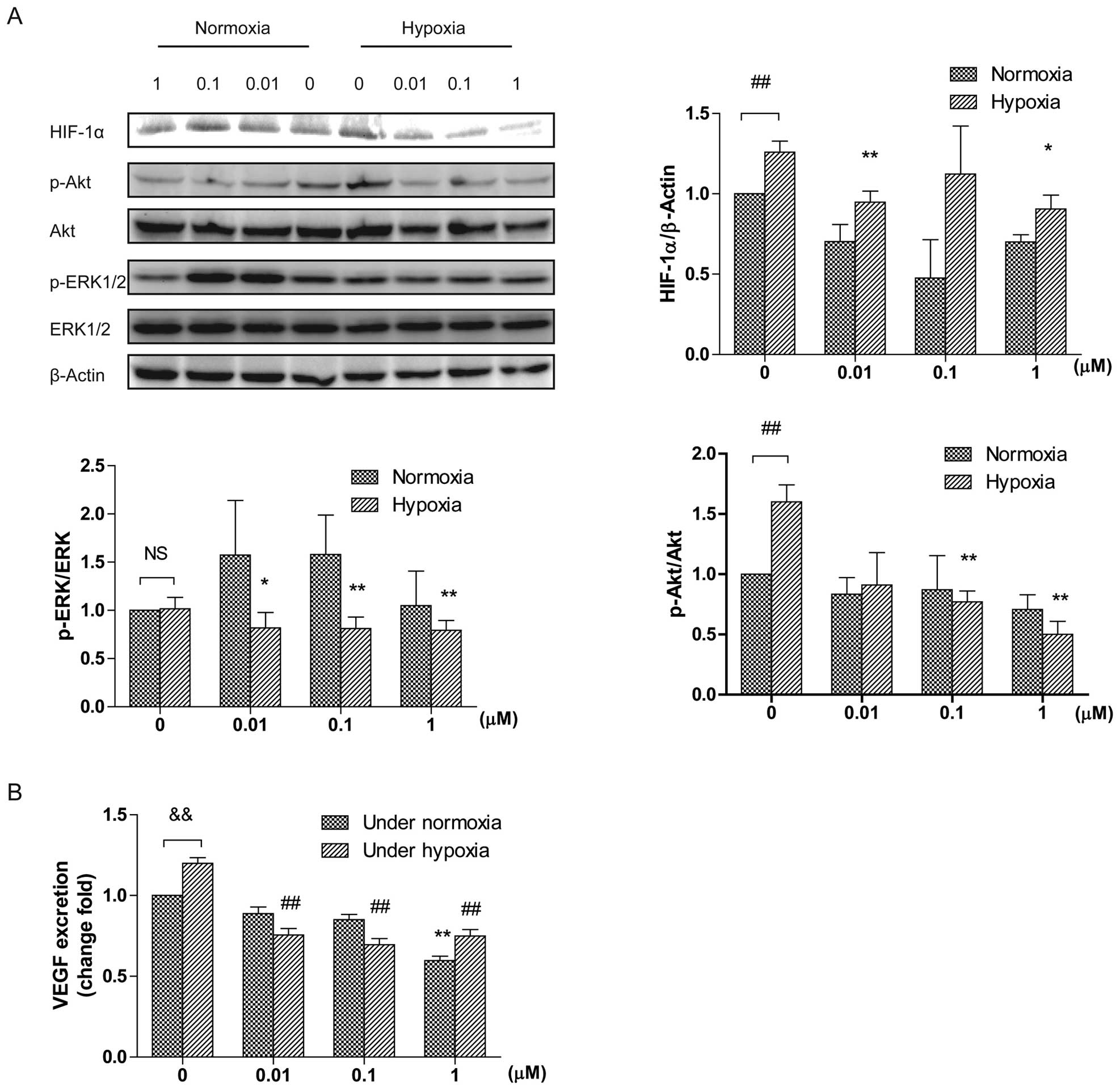 | Figure 4(A) The effect of DT-13 on ERK1/2 and
Akt activation and the level of HIF-1α under normoxia and hypoxia.
Western blot analysis demonstrated that DT-13 inhibits
phosphorylation of ERK1/2, Akt and HIF-1α. Total cell lysate was
mixed with SDS loading buffer and separated by SDS-PAGE, blotted
onto nitrocellulose membranes. Membranes were immunoblotted with
the respective primary antibodies: p-ERK1/2, ERK1/2, p-Akt, Akt,
HIF-1α and β-actin. *p<0.05; **p<0.01,
statistical significance relative to vehicle control under hypoxia.
##p<0.01, statistical significance relative to
vehicle control under normoxia. (B) The effect of DT-13 on VEGF
secretion under normoxia and hypoxia. HUVECs were plate into 60-mm
dishes with completed medium overnight. HUVECs were changed medium
to serum and growth factor supplement-free medium. After 2 h, cells
were treated with various concentrations of DT-13 for 12 h under
hypoxia or normoxia. After treatment, the medium was harvested to
detect the concentration of VEGF by ELISA. **p<0.01,
statistical significance relative to vehicle control under
normoxia. ##p<0.01, statistical significance relative
to vehicle control under hypoxia. &p<0.01,
statistical significance relative to vehicle control under
hypoxia. |
DT-13 inhibits VEGF excretion under
hypoxia
We also detected whether DT-13 impacts VEGF
secretion under normoxia and hypoxia with the ELISA assay. The
medium of HUVECs was changed to serum-free medium and incubated
with or without DT-13 under normoxia or hypoxia for 12 h. The
conditioned medium was collected and examined. As shown in Fig. 4B, under normoxia DT-13 decreased
VEGF secretion in a dose-dependent manner with the inhibition rate
of 11, 15 and 40% by 0.01, 0.1 and 1 μM of DT-13. Under hypoxia,
DT-13 always showed similar inhibitory rates by 40% compared with
control (37, 42 and 38% by 0.01, 0.1 and 1 μM of DT-13
respectively) (Fig. 4B).
DT-13 inhibits angiogenesis induced by
VEGF
As shown above, DT-13 exhibited anti-angiogenic
ability and inhibited VEGF excretion under normoxia and hypoxia,
thus we investigated whether DT-13 inhibits angiogenesis induced by
VEGF. Initially, DT-13 did not inhibit cell proliferation at the
presence of VEGF within 48 h (Fig.
5A). However, DT-13 exhibited anti-angiogenic effects observed
from cell migration and tube formation assays. DT-13 reduced the
number of migrated cells induced by VEGF with the inhibitory rate
of 61.5, 40.1 and 38.9% at the concentration of 0.01, 0.1 and 1 μM
respectively (Fig. 5B), and
decreased tube formation to 63.4, 49.3 and 35% of control at the
concentration of 0.01, 0.1 and 1 μM (Fig. 5C). The results indicate that DT-13
attenuates the effect of VEGF on angiogenesis by inhibiting HUVEC
migration and tube formation, but does not affect HUVEC
proliferation induced by VEGF.
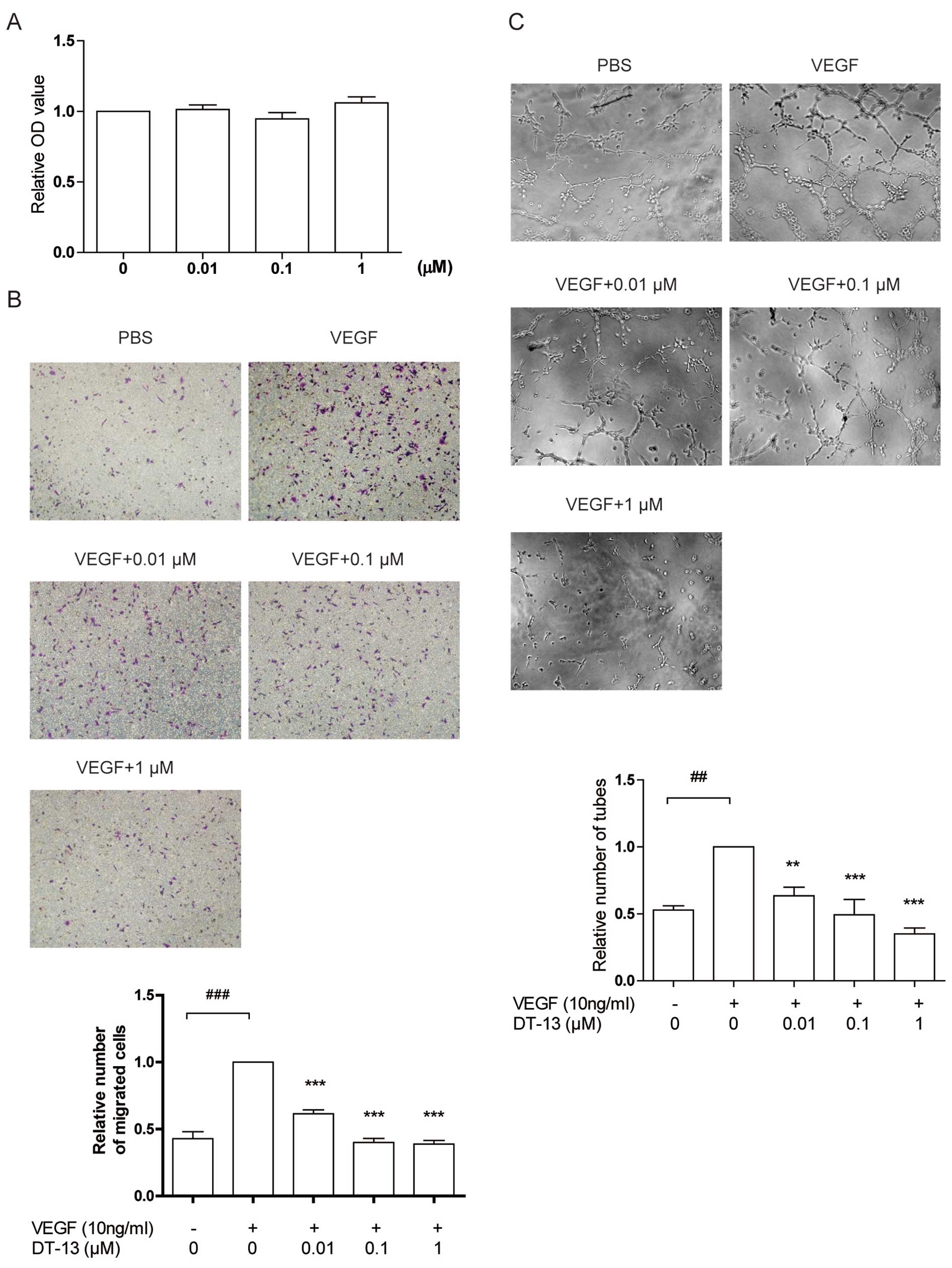 | Figure 5DT-13 inhibits angiogenesis stimulated
by VEGF in vitro. (A) The effect of DT-13 on HUVEC
proliferation stimulated by VEGF for 48 h under normoxia. HUVECs
were pre-treated with DT-13 (0.01, 0.1 and 1 μM) or PBS for 1 h,
then treated with 10 ng/ml VEGF for 48 h. After treatment, the
number of cells was counted by CCK-8. (B) The effect of DT-13 on
HUVEC migration stimulated by VEGF. HUVECs in 1% FBS and ECGS-free
medium with DT-13 (0.01, 0.1 and 1 μM) or PBS were plated into the
inner chamber of Transwell and 1% FBS and ECGS-free medium plus 10
ng/ml VEGF was added into the outer chamber. The Transwell system
was incubated for 4 h. After treatment, the cells on the outer side
membrane were fixed and stained by crystal violet. These cells were
captured and counted. ***p<0.001, statistical
significance relative to the VEGF plus group.
###p<0.001, statistical significance relative to the
VEGF-free group. (C) The effect of DT-13 on tube formation
stimulated by VEGF. HUVECs in 1% FBS and ECGS-free medium were
plated into 96-well plates pre-coated by growth factor reduced
Matrigel and treated with VEGF (10 ng/ml), DT-13 (0.01, 0.1 and 1
μM) or PBS. HUVECS were incubated for 6 h. After treatment, the
tubes were captured and counted. **p<0.01;
***p<0.001, statistical significance relative to the
VEGF plus group. ##p<0.01, statistical significance
relative to the VEGF-free group. |
DT-13 exhibits anti-angiogenic activity
by inhibiting the VEGF pathway
As shown above, angiogenesis induced by VEGF is
weakened by DT-13. Therefore, we evaluated if DT-13 affects the
downstream of VEGF pathway. Firstly, we detected the
phosphorylation of VEGFR2, the receptor of VEGF. We pretreated
HUVECs with or without DT-13 for 1 h and then treated with 10 ng/ml
VEGF for 15 min. VEGF significantly promoted the phosphorylation of
VEGFR2 compared with PBS, while DT-13 decreased the level of
p-VEGFR2 induced by VEGF, with a significant difference at the
concentration of 1 μM (Fig. 6).
Furthermore, we examined the phosphorylation of Akt and ERK1/2
after VEGF and DT-13 treatment. VEGF increased the level of p-Akt,
which was markedly attenuated by 1 μM DT-13 (Fig. 6). However, increased p-ERK1/2 was
slightly decreased by DT-13 without statistical difference
(Fig. 6). These results suggest
that under hypoxia, DT-13 inhibits angiogenesis by inhibiting the
VEGF pathway.
Discussion
In this study, we demonstrated that DT-13 exhibits
anti-angiogenic activity under hypoxia and normoxia, and inhibits
angiogenesis induced by VEGF under normoxia in vitro. DT-13
administration resulted in a significant reduction of HUVEC
proliferation, migration and tube formation under hypoxia and
normoxia. Further evaluation with CAM likely results in reduced
relative vessel length in DT-13-treated eggs. To address the
potential mechanism of the anti-angiogenic effects of DT-13, we
detected the elements involved in angiogenesis, such as HIF-1α,
p-Akt, and ERK1/2 which showed significantly decreased levels after
DT-13 treatment under hypoxia but not under normoxia. Additionally,
DT-13 reduced VEGF secretion and VEGF induced p-VEGFR2 and p-Akt.
These findings indicate a role for DT-13 in the inhibition of
angiogenesis under hypoxia and normoxia or even of angiogenesis
induced by VEGF.
At present, there are three types of
anti-angiogenesis drugs: antibody, synthetic small molecules, and
natural products (20). Although
the effects of the anti-VEGF-A antibody, bevacizumab, have been
confirmed in clinical anti-angiogenic therapy combined with a
cytotoxic agent (21), there are
still adverse effects in the cardiovascular and renal systems
(22,23). Multiple-target small molecules and
natural products performed anti-angiogenic activity, such as
sorafenib and curcumin (20).
Previous studies on cancer cells suggested that DT-13 exhibits
anticancer activity via inhibiting cancer cell migration and
adhesion in vitro(13,14).
Our data showed that DT-13 displays anti-angiogenic activity by
inhibiting tube formation and HUVEC migration. Endothelial cells in
tumor are under two main oxygen conditions, normoxia and hypoxia.
Endothelial cells are under hypoxia during the pathological state,
such as atherosclerosis and cancer (19,24–26).
Hypoxic conditions stimulate the expression of pro-angiogenic
protein, such as VEGF and its receptor VEGFR2, in endothelial cells
(27–29). VEGFR-2 plays a crucial role in
angiogenesis and activation of VEGFR-2 promotes endothelial cell
growth, migration and tube formation (30). In VEGFR-2-deficient mice, failed
formation of blood islands and blood vessels led to embryonic
lethality (31), which indicates
that VEGFR-2 is essential in angiogenesis. Stimulation of VEGFR-2
phosphorylates downstreams, such as ERK1/2 (32) and Akt (33). DT-13 reduces the level of p-Akt and
ERK1/2 under hypoxia and p-VEGFR-2 and p-Akt induced by VEGF under
normoxia, but displays little effect on p-ERK1/2 induced by VEGF.
DT-13 displays higher activity of inhibition on p-Akt and p-ERK1/2
under hypoxia than under normoxia. Furthermore, DT-13 reduces the
increased HIF-1α induced by hypoxia, which may also reduce
angiogenesis considering the role of HIF-1α in angiogenesis
(19). Thus, DT-13 could be a new
multi-targeting anticancer agent by inhibiting angiogenesis, tumor
growth and metastasis.
In conclusion, we provided evidence for the first
time that DT-13 exhibits anti-angiogenic activity. Further animal
studies are required to confirm the effect of DT-13 on angiogenesis
and to explore the side-effects of DT-13.
Acknowledgements
This study was financially supported by the National
Natural Science Fund Nos. 81102853 and 81071841, and 2011’ Program
for Excellent Scientific and Technological Innovation Team of
Jiangsu Higher Education.
References
|
1
|
Bamias A and Dimopoulos MA: Angiogenesis
in human cancer: implications in cancer therapy. Eur J Intern Med.
14:459–469. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zetter BR: Angiogenesis and tumor
metastasis. Annu Rev Med. 49:407–424. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
He K, Jin K, Wang H and Teng L:
Anti-angiogenic therapy for colorectal cancer: on the way to
getting better! Hepatogastroenterology. 59:1113–1117.
2012.PubMed/NCBI
|
|
4
|
Das M and Wakelee H: Targeting VEGF in
lung cancer. Expert Opin Ther Targets. 16:395–406. 2012. View Article : Google Scholar
|
|
5
|
Teghom C, Giraud P, Menei P, et al: Renal
carcinoma: point on treatment of brain metastasis. Bull Cancer.
99:627–634. 2012.(In French).
|
|
6
|
Wicki A and Rochlitz C: Targeted therapies
in breast cancer. Swiss Med Wkly. 142:w135502012.
|
|
7
|
Dachs GU and Tozer GM: Hypoxia modulated
gene expression: angiogenesis, metastasis and therapeutic
exploitation. Eur J Cancer. 36:1649–1660. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fang XC and Yu BY: Application of
pyrolysis-high-resolution gas chromatography-pattern recognition to
the identification of the Chinese traditional medicine mai dong. J
Chromatogr. 514:287–292. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang PMAX: The initial research on Hupeh
Liriope Root Tuber inducing HL60 cell differentiation. Dalian Med
College J. 14:37–41. 1992.
|
|
10
|
Mimaki Y, Takaashi Y, Kuroda M, Sashida Y
and Nikaido T: Steroidal saponins from Nolina recurvata
stems and their inhibitory activity on cyclic AMP
phosphodiesterase. Phytochemistry. 42:1609–1615. 1996.
|
|
11
|
Jeong A, Lee HJ, Jeong SJ, Lee EO, Bae H
and Kim SH: Compound K inhibits basic fibroblast growth
factor-induced angiogenesis via regulation of p38 mitogen activated
protein kinase and AKT in human umbilical vein endothelial cells.
Biol Pharm Bull. 33:945–950. 2010. View Article : Google Scholar
|
|
12
|
Tong Y, Zhang X, Tian F, et al:
Philinopside A, a novel marine-derived compound possessing dual
anti-angiogenic and anti-tumor effects. Int J Cancer. 114:843–853.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sun L, Lin S, Zhao R, Yu B, Yuan S and
Zhang L: The saponin monomer of dwarf lilyturf tuber, DT-13,
reduces human breast cancer cell adhesion and migration during
hypoxia via regulation of tissue factor. Biol Pharm Bull.
33:1192–1198. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang Y, Liu J, Kou J, Yu J and Yu B:
DT-13 suppresses MDA-MB-435 cell adhesion and invasion by
inhibiting MMP-2/9 via the p38 MAPK pathway. Mol Med Rep.
6:1121–1125. 2012.PubMed/NCBI
|
|
15
|
Jaffe EANR, Becker CG and Minick CR:
Culture of human endothelial cells derived from umbilical veins.
Identification by morphologic and immunologic criteria. J Clin
Invest. 52:2745–2756. 1973. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lin S, Sun L, Hu J, et al: Chemokine C-X-C
motif receptor 6 contributes to cell migration during hypoxia.
Cancer Lett. 279:108–117. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee OH, Kim YM, Lee YM, et al: Sphingosine
1-phosphate induces angiogenesis: its angiogenic action and
signaling mechanism in human umbilical vein endothelial cells.
Biochem Biophys Res Commun. 264:743–750. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ashton AW, Yokota R, John G, et al:
Inhibition of endothelial cell migration, intercellular
communication, and vascular tube formation by thromboxane A(2). J
Biol Chem. 274:35562–35570. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen L, Endler A and Shibasaki F: Hypoxia
and angiogenesis: regulation of hypoxia-inducible factors via novel
binding factors. Exp Mol Med. 41:849–857. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wahl O, Oswald M, Tretzel L, Herres E,
Arend J and Efferth T: Inhibition of tumor angiogenesis by
antibodies, synthetic small molecules and natural products. Curr
Med Chem. 18:3136–3155. 2011. View Article : Google Scholar
|
|
21
|
Ma J and Waxman DJ: Combination of
antiangiogenesis with chemotherapy for more effective cancer
treatment. Mol Cancer Ther. 7:3670–3684. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Higa GM and Abraham J: Biological
mechanisms of bevacizumab-associated adverse events. Expert Rev
Anticancer Ther. 9:999–1007. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen HX and Cleck JN: Adverse effects of
anticancer agents that target the VEGF pathway. Nat Rev Clin Oncol.
6:465–477. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chakrabarti S, Rizvi M, Pathak D, Kirber
MT and Freedman JE: Hypoxia influences CD40-CD40L mediated
inflammation in endothelial and monocytic cells. Immunol Lett.
122:170–184. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sluimer JC and Daemen MJ: Novel concepts
in atherogenesis: angiogenesis and hypoxia in atherosclerosis. J
Pathol. 218:7–29. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Izzard AS, Emerson M, Prehar S, et al: The
cardiovascular phenotype of a mouse model of acromegaly. Growth
Horm IGF Res. 19:413–419. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Verlohren S, Stepan H and Dechend R:
Angiogenic growth factors in the diagnosis and prediction of
pre-eclampsia. Clin Sci. 122:43–52. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nagamatsu T, Fujii T, Kusumi M, et al:
Cytotrophoblasts up-regulate soluble fms-like tyrosine kinase-1
expression under reduced oxygen: an implication for the placental
vascular development and the pathophysiology of preeclampsia.
Endocrinology. 145:4838–4845. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Forsythe JA, Jiang BH, Iyer NV, et al:
Activation of vascular endothelial growth factor gene transcription
by hypoxia-inducible factor 1. Mol Cell Biol. 16:4604–4613.
1996.PubMed/NCBI
|
|
30
|
Takahashi S: Vascular endothelial growth
factor (VEGF), VEGF receptors and their inhibitors for
antiangiogenic tumor therapy. Biol Pharm Bull. 34:1785–1788. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shalaby F, Rossant J, Yamaguchi TP, et al:
Failure of blood-island formation and vasculogenesis in
Flk-1-deficient mice. Nature. 376:62–66. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zachary I and Gliki G: Signaling
transduction mechanisms mediating biological actions of the
vascular endothelial growth factor family. Cardiovasc Res.
49:568–581. 2001. View Article : Google Scholar
|
|
33
|
Gerber HP, McMurtrey A, Kowalski J, et al:
Vascular endothelial growth factor regulates endothelial cell
survival through the phosphatidylinositol 3′-kinase/Akt signal
transduction pathway. Requirement for Flk-1/KDR activation. J Biol
Chem. 273:30336–30343. 1998.
|