Introduction
Epithelial ovarian cancer is the leading cause of
cancer death among the gynecological malignancies and its incidence
rate is highest in Western industrialized countries. A lack of
early symptoms or screening methods of ovarian cancer often leads
to late diagnosis in advanced stage and subsequent high mortality
rate (1). Moreover, no highly
effective therapeutics and strategies against metastatic and
recurrent ovarian cancers have been established.
Receptor tyrosine kinases (RTKs) play pivotal roles
in cellular process including proliferation, migration and
survival. Mutations in RTKs and dysregulated activation of their
intracellular signaling pathways have been reported to be closely
involved in a variety of diseases such as cancers, angiogenesis,
and inflammation (2). RTKs such as
epidermal growth factor receptor (EGFR), vascular endothelial
growth factor receptor (VEGFR) and fibroblast growth factor
receptor (FGFR) are highly expressed or activated in ovarian cancer
(3–8). Selective inhibition of RTK activation
or downstream signaling pathways appears to be a potent strategy
with little or no side effects, compared to conventional therapy.
Epithelial-to-mesenchymal transition (EMT), commonly occurs in
epithelial cancers, it is a cellular process which involves changes
in cellular morphology, the loss and remodeling of cell-cell and
cell-matrix adhesions, and subsequent acquisition of cell migration
and invasion (9). During EMT,
expression of the epithelial markers such as epithelial cadherin
(E-cadherin) and claudins is reduced, whereas that of mesenchymal
markers including neural-cadherin (N-cadherin), integrin α5β1, and
matrix metalloproteinases (MMPs) is increased (10–12).
EMT-associated cancer cell invasion and metastasis are mediated by
the tissue remodeling due to changes in expression levels of cell
adhesion molecules and/or elevated levels of secreted proteolytic
enzymes including MMPs (13).
Therefore, unraveling the molecular mechanisms and targets of
ovarian cancer progression within the context of extracellular
matrix (ECM) and the function of the tumor microenvironment may
help to improve current concepts and therapies for the treatment of
ovarian cancer.
Siegesbeckia glabrescens (SG) Makino
(Compositae) has been used as a traditional medicine for the
treatment of allergic diseases, paralysis, acute hepatitis, and
hypertension. Many investigations have demonstrated that the
extracts and biologically active components of SG have
anti-allergic and anti-inflammatory activities (14–17).
In addition, the water extract of SG has been reported to have
antitumor activity against breast cancer cells by inducing
apoptosis (18), however, the
effects and molecular mechanisms of SG on other cancer cells remain
unexplored. In the present study, we evaluated the regulatory
effects and signaling pathways of SG on proliferation, adhesion,
migration and invasion in ovarian cancer cells.
Materials and methods
Cell culture conditions
Human ovarian carcinoma cells (SKOV-3) from American
Type Culture Collection (Manassas, VA, USA) were grown in 10% fetal
bovine serum-Dulbecco’s modified Eagle’s medium (FBS-DMEM) (HyClone
Laboratories, Logan, UT, USA).
Reagents
The following antibodies were purchased from
commercial sources: anti-integrin β1, anti-N-cadherin,
anti-phospho-FAK (Y397), anti-FAK, (BD Biosciences, Bedford, MA,
USA); anti-phospho-Src (Y416), anti-Src, anti-phospho-ERK
(T202/Y204), anti-phospho-Akt (S473),
anti-phospho-p70S6K (T421/S424), anti-phospho-pRb
(S780), anti-phospho-pRb (S807/S811), anti-pRb (Cell Signaling,
Beverly, MA, USA); anti-EGFR, anti-VEGFR-2, anti-FGFR-1, anti-ILK,
anti-ERK, anti-Akt, anti-p70S6K, anti-MMP-2, anti-Cdk4,
anti-Cdk2, anti-cyclin D, anti-cyclin E, anti-actin antibodies, and
mouse and rabbit IgG-horseradish peroxidase conjugates (Santa Cruz
Biotechnology, Santa Cruz, CA, USA).
Preparation of SG extract
The ethanol extract was prepared by mixing 100 g SG
with 1 liter of ethanol and stirring for 90 min. The extract was
then filtered through a filter paper (Advantec no. 5C, Toyo Roshi
Kaisha, Tokyo, Japan), and the filtrate was concentrated using a
rotary evaporator (Heidolph Instruments GmbH & Co., Schwabach,
Germany) at 46°C under vacuum. The yield of the dried extract was
approximately 6.5 g/l.
Cell growth assay
SKOV-3 cells, plated on 6-well plates
(2×104 cells/well, BD Biosciences), were serum-starved
for 24 h to synchronize cells in G1/G0 phase
of the cell cycle, and further incubated with 10% FBS-DMEM for 24 h
in the presence of SG at different concentrations (0.1–10 μg/ml).
The cells were then washed in ice-cold phosphate buffered saline
(PBS, pH 7.4), detached with trypsin, and counted using trypan blue
exclusion method (19, 20). The results from triplicate
determinations (mean ± standard deviation) are presented as the
fold-increase of untreated controls.
Western blot analysis
Subconfluent cells in 100-mm dishes (BD Biosciences)
were serum-starved for 24 h in DMEM and replaced with fresh media,
followed by treatments for different time points, as indicated, at
37°C. Cells were rinsed twice with ice-cold PBS and lysed by
incubation in 50 mM Tris-HCl (pH 7.4), 150 mM NaCl, 10% glycerol,
1% Triton X-100, 1 mM EDTA, 100 μg/ml
4-(2-aminoethyl)benzenesulfonyl fluoride, 10 μg/ml aprotinin, 1
μg/ml pepstatin A, 0.5 μg/ml leupeptin, 80 mM β-glycerophosphate,
25 mM sodium fluoride and 1 mM sodium orthovanadate for 30 min at
4°C. Cell lysates were clarified at 13,000 × g for 20 min at 4°C,
and the supernatants were subjected to western blot analysis as
previously described (21).
Adhesion assay
Subconfluent cells were detached with trypsin and
allowed to recover in 10% FBS-DMEM for 1 h at 37°C with gentle
rocking. After recovery, the cells were collected by low-speed
centrifugation and resuspended in serum-free DMEM. The cell
suspension was pretreated with or without SG for 30 min, and
followed by serum treatment. The cells were plated on 96-well
plates (1.5×104 cells/well), and further incubated for 2
h at 37°C. Following incubation unattached cells were removed by
washing the wells three times with PBS. Attached cells were fixed
with methanol, and then stained with 0.04% Giemsa staining solution
(Sigma-Aldrich Co., St. Louis, MO, USA). The cells were
photographed and counted. The results (mean ± standard deviation)
are presented as the numbers of adherent cells (22).
Migration assay
Cell migration was quantified in the in vitro
wound-healing assay as previously described (23,24).
After cells were plated on 48-well plates, and grown to confluence,
a single wound was created in the center of the cell monolayer by
the gentle removal of the attached cells with a sterile plastic
pipette tip. Following incubation with serum-free DMEM for 2 h,
cells were pretreated with or without SG for 30 min, followed by
serum stimulation for 14 h. Cells were fixed with methanol, and
then stained with 0.04% Giemsa solution. The migration of the cells
into the wound was observed with still images taken at the
indicated time points.
Invasion assay
The upper side of the transwell insert (Costar, 6.5
mm diameter insert, 8 μm pore size) (Corning Inc., Corning, NY,
USA) was coated with 50 μl of 1 mg/ml Matrigel (BD Biosciences)
diluted in serum-free DMEM at 37°C. Aliquots (100 μl) of cells
(8×105 cells/ml) resuspended in serum-free DMEM were
added to the upper compartment of the Matrigel-coated transwell and
600 μl of serum-free DMEM was added to the lower compartment. After
serum starvation for 2 h, cells were pretreated with or without SG
for 30 min, followed by serum stimulation for 15 h. The inserts
were fixed with methanol and using a cotton-tipped swab the
non-invasive cells were removed from the top of the membrane. After
staining with 0.04% Giemsa solution, the number of invasive cells
was determined from six different fields using ×200 objective
magnification (25).
Zymogram analysis
Activities of matrix metalloproteinases (MMPs) were
measured by zymography (21,26).
Aliquots of conditioned medium were diluted in sample buffer,
applied to 10% polyacrylamide gels containing 1 mg/ml gelatin
(Sigma-Aldrich Co.) as a substrate. After electrophoresis, the gels
were incubated in 2.5% Triton X-100 for 1 h to remove SDS and allow
re-naturalization of MMPs, and further incubated in developing
buffer containing 50 mM Tris-HCl (pH 7.5), 10 mM CaCl2,
and 150 mM NaCl for 15 h at 37°C. The gels were stained with 0.5%
Coomassie brilliant blue R-250 in 30% methanol-10% acetic acid for
2 h and followed by destaining with 30% methanol-10% acetic acid.
Gelatinolytic activities were detected as unstained bands against
the background of the Coomassie blue-stained gelatin.
Statistical analysis
Statistical analysis was performed using Student’s
t-test, and was based on at least three different experiments. The
results were considered to be statistically significant when
p<0.05.
Results
SG suppresses cell proliferation through
modulating the expression of cell cycle-related proteins
We first examined the effect of SG on cell
proliferation in SKOV-3 cells. SG treatment marginally, but
significantly, suppressed cell proliferation in a dose-dependent
manner (Fig. 1A). Based on these
findings, we next analyzed the changes of cell cycle-related
proteins. It has been reported that cell cycle progression requires
activation of cyclin-dependent kinases (Cdks) through formation
with cyclins and subsequent phosphorylation of retinoblastoma
protein (pRb), resulting in transition from G1 to S phase of the
cell cycle. The kinase activity of these complexes is regulated by
the Cip/Kip family of Cdk inhibitors such as p27Kip1 and
p21WAF1/Cip1 as well as INK-4 family that specifically
inhibit Cdk4/6-cyclin D complexes (27). SG treatment dramatically
downregulated the expression of cyclin E and upregulated the
p27Kip1 levels to levels observed in untreated controls
(Fig. 1B). However, SG treatment
did not significantly affect the levels of Cdk4, Cdk2, and cyclin D
but decreased those of Cdk4 and cyclin D reproducibly (Fig. 1B). In addition, we analyzed the
phosphorylation of pRb on residues Ser 780 and Ser 807/811, sites
specific for Cdk4 and Cdk2 phosphorylation, respectively. As shown
in Fig. 1C, SG treatment
dose-dependently suppressed pRb phosphorylation at both sites, in
good agreement with the expression levels of Cdks and cyclins
(Fig. 1B). Collectively, these
findings demonstrate that SG regulates the expression of cell
cycle-related proteins and subsequent phosphorylation of pRb,
resulting in inhibition of cell cycle progression and
proliferation.
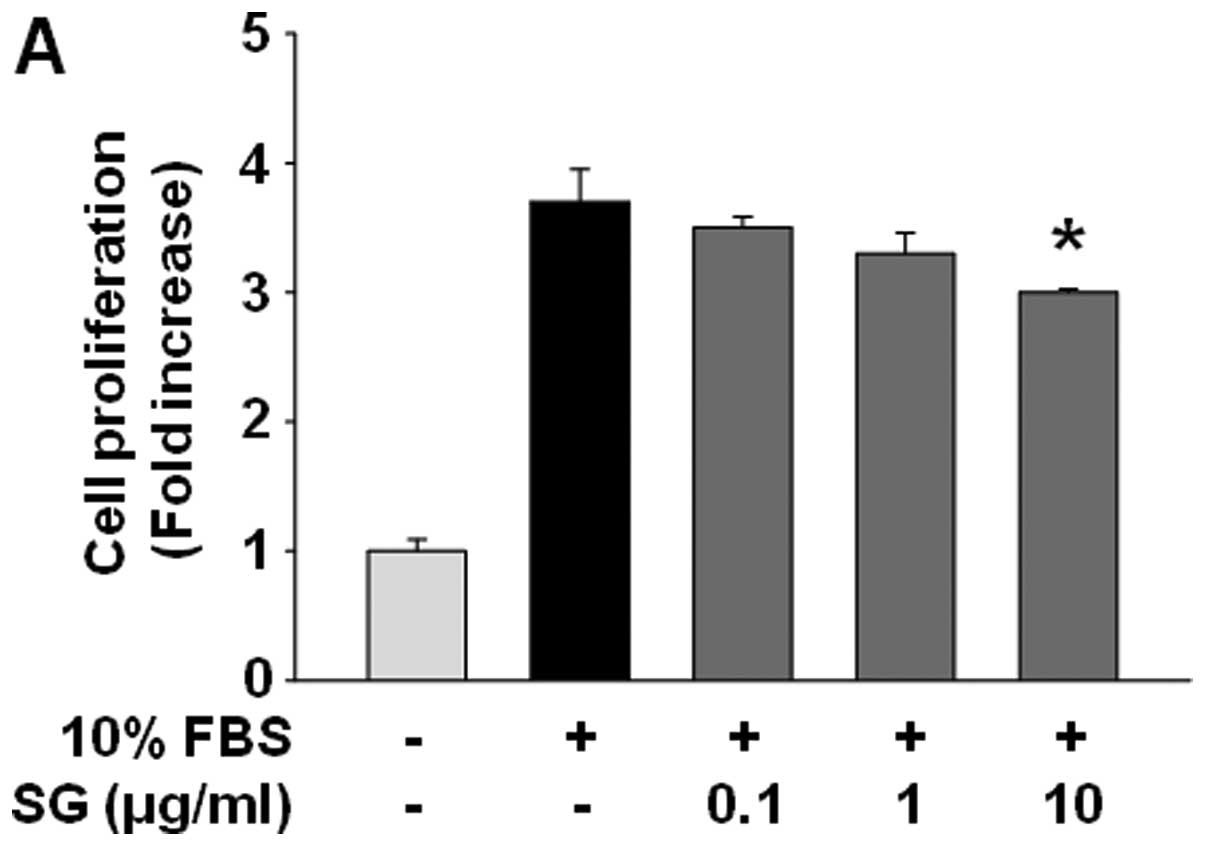 | Figure 1SG treatment suppresses proliferation
by downregulation of cyclin E and upregulation of
p27Kip1 in SKOV-3 cells. (A) Quiescent cells were
pretreated with or without SG at different concentrations (0.1–10
μg/ml) for 30 min, followed by 10% serum stimulation for 24 h. Cell
proliferation results from three independent experiments (mean ±
SD) are presented as the fold-increase of untreated controls.
Statistical significance is indicated (*p<0.05,
compared with 10% serum-treated cells). (B and C) Cell lysates were
western blotted with anti-Cdk4, anti-cyclin D, anti-Cdk2,
anti-cyclin E, anti-p27Kip1, anti-actin,
anti-phospho-pRb (S780), anti-phospho-pRb (S807/S811), or anti-pRb
antibodies. Results shown are representative of at least three
independent experiments. |
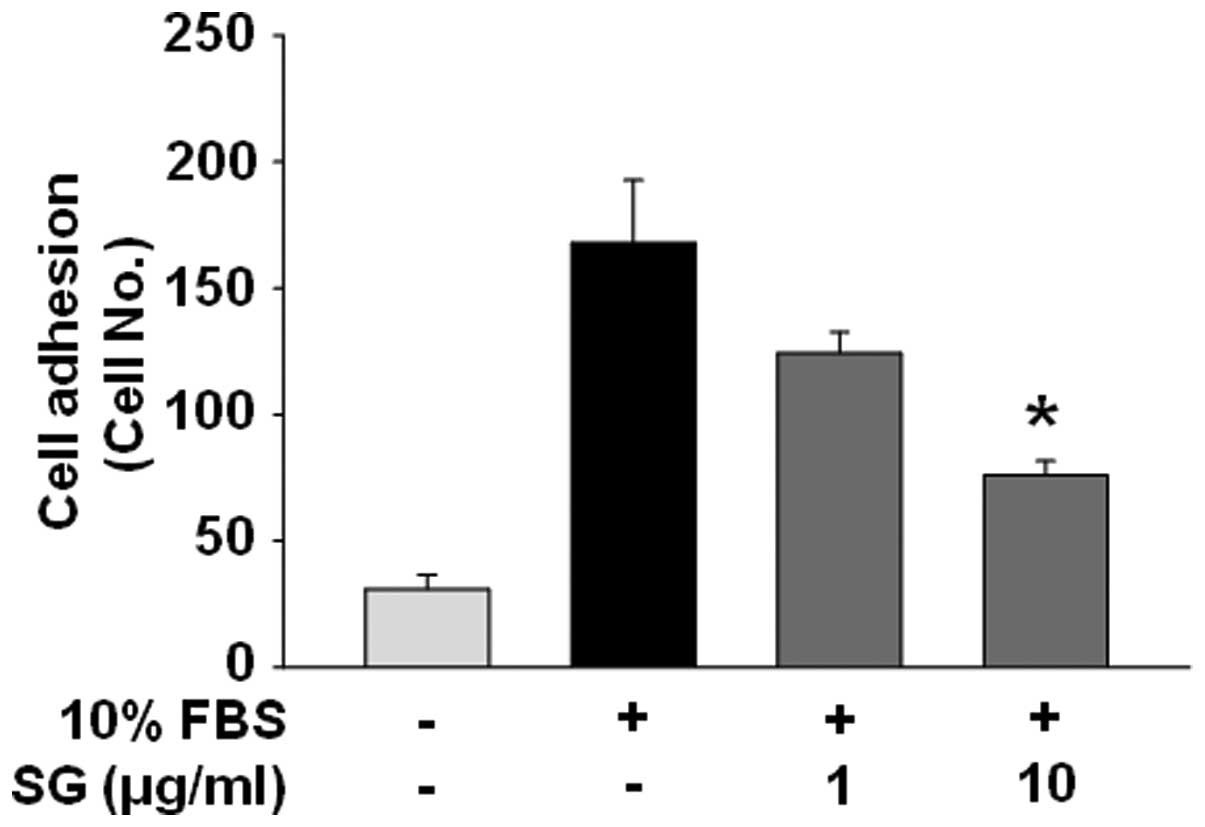
SG inhibits ovarian cancer cell adhesion,
motility and invasion
Cell adhesion and migration associated with EMT and
tumor microenvironment are controlled by coordinated processes
through the interactions with ECMs as well as intercellular
components (9–12). We next examined the effects of SG on
adhesion, migration and invasion of SKOV-3 cells in response to
mitogenic stimulation. SG treatment markedly inhibited cell
adhesion, migration, invasion in a dose-dependent manner (Figs. 2 and 3A
and B). Expression and activation of MMPs have been reported to
promote cell migration and invasion by selective proteolytic
degradation of ECM components (13). Based on SG-mediated inhibition of
cell migration and invasion, we analyzed the levels of MMPs in
SKOV-3 cells. SG treatment showed little or no change of expression
and activity of MMP-2 and MMP-9 (Fig.
3C and D), suggesting the existence of alternative mechanism to
inhibit cell migration and invasion by SG treatment, and this
mechanism appears to be independent of regulation of MMP
activity.
In vitro antitumor effects of SG on
ovarian cancer are mediated through inhibition of mitogenic
signaling pathways, and downregulation of receptor tyrosine kinases
and N-cadherin
To further investigate the molecular mechanisms by
which SG regulates cell fates, we examined the changes in
activation of intracellular signaling components including focal
adhesion kinase (FAK), Src, extracellular signal-regulated kinase
(ERK), phosphatidylinositol 3-kinase (PI3-K)/Akt, and mammalian
target of rapamycin (mTOR)/p70S6K and expression of RTKs
including EGFR, VEGFR-2, and FGFR-1 which play the pivotal roles in
cell proliferation, migration, and invasion (2,28). As
expected, mitogenic stimulation for 15 min markedly increased the
phosphorylation/activation of FAK, ERK, Akt, and p70S6K,
but not that of Src, when compared with unstimulated controls
(Fig. 4A). SG treatment
significantly inhibited the phosphorylation of FAK, ERK, Akt and
p70S6K in a dose-dependent manner. This suppression of
downstream signaling pathways of integrins and RTKs appears to
mediate anti-proliferative, anti-adhesive, anti-migratory and
anti-invasive properties of SG in ovarian cancer cells.
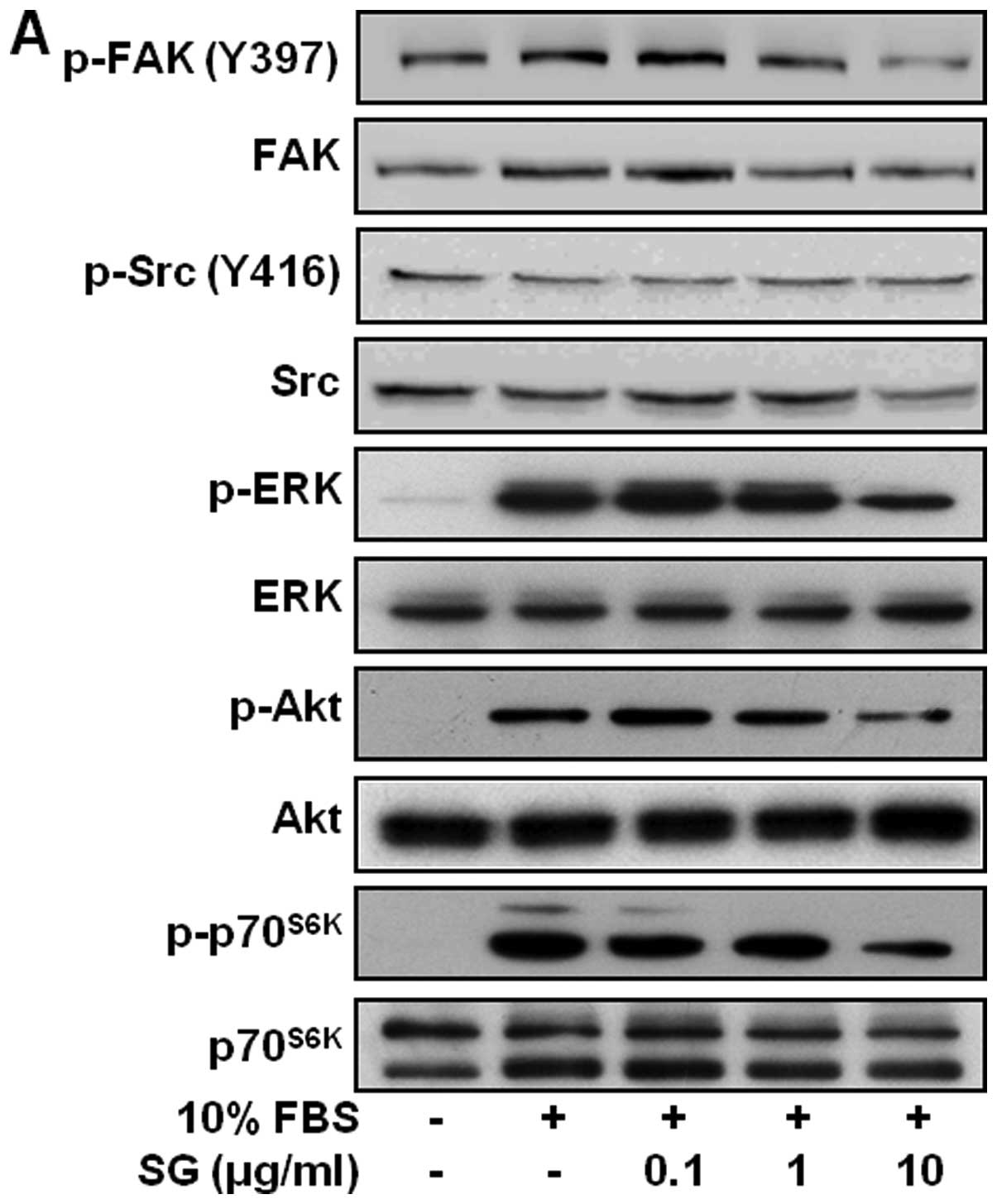 | Figure 4Changes of signaling pathways and RTK
expression in SG-treated cells. (A) Quiescent cells were pretreated
with SG at different concentrations (0.1–10 μg/ml) for 30 min,
followed by 10% serum stimulation for 15 min. Cell lysates were
western blotted with anti-phospho-FAK, anti-FAK, anti-phospho-Src,
anti-Src, anti-phospho-ERK, anti-ERK, anti-phospho-Akt, anti-Akt,
anti-phsopho-p70S6K, or anti-p70S6K
antibodies. (B) Cells were pretreated with SG for 30 min, followed
by serum stimulation for 24 h. Cell lysates were western blotted
for EGFR, VEGFR-2, FGFR-1, integrin β1 and ILK. (C) Cells were
pretreated with PD98059 (25 μM), LY294002 (10 μM), or rapamycin (50
nM) for 30 min, and then stimulated with 10% serum for 24 h. Cell
lysates were western blotted for EGFR, VEGFR-2 and FGFR-1. Results
shown are representative of three independent experiments. |
We next analyzed the changes in the expression of
RTKs and integrin β1, which are overexpressed and correlate with
ovarian cancer progression and metastatic potential (4,6–8,29–31).
As shown in Fig. 4B, SG treatment
markedly inhibited mitogen-induced expression of EGFR, VEGFR-2 and
FGFR-1, but not that of integrin β1, to levels observed in
unstimulated controls. In addition, SG treatment suppressed the
expression of integrin-linked kinase (ILK), which can interact with
the cytoplasmic domain of integrin β subunit and is activated by
both integrin activation and growth factors (32). Finally, pretreatment of cells with
PD98059, an inhibitor of ERK pathway, LY294002, an inhibitor of
PI3-K/Akt pathway, or rapamycin, an inhibitor of
mTOR/p70S6K pathway, mimicked the suppressive effects of
SG on RTK expression (Fig. 4C),
cell proliferation, adhesion, and migration (data not shown),
indicating that the ethanol extract of SG contains the
pharmacologically effective components, similar to these
inhibitors, and may share the roles and mechanisms of action in
regulating cell fates.
Loss of E-cadherin from the cell surface, commonly
occurs in cancer progression, resulting in disruption of cell-cell
contacts, cell detachment, cell shape change, cell migration and
invasion. On the other hand, elevation of N-cadherin, one of the
mesenchymal markers, induces EMT and cancer progression (9,13).
Alteration in cadherin function can be assessed by the levels of
cadherin detectable in the Triton X-100 insoluble cell extracts. As
shown in Fig. 5, SG treatment
completely inhibited mitogen-stimulated distribution of N-cadherin
in detergent-insoluble fraction to levels observed in unstimulated
controls, however, did not alter the expression of N-cadherin in
detergent-soluble fraction. In contrast, the expression of
E-cadherin in SKOV-3 cells was not clearly detectable (data not
shown). Taken together, these findings demonstrate that suppression
of cell proliferation, adhesion, migration and invasion by SG
ethanol extracts might be mediated through inactivation of
mitogen-stimulated signaling pathways and subsequent downregulation
of RTKs and N-cadherin.
Discussion
Siegesbeckia glabrescens (SG) has mainly been
used in traditional medicine for the treatment of allergic and
inflammatory diseases. This application is well supported by recent
studies that SG possesses pharmacologically active components such
as flavones and sesquiterpene lactone to reduce inflammation
(16,17). In addition to anti-inflammatory
activity, it has been reported to have anti-proliferative effect on
breast cancer cell lines by inducing apoptosis (18). However, detailed molecular
mechanisms of SG responsible for regulation of cancer growth and
progression have been clearly reported to date. In the present
study, we demonstrate for the first time that the ethanol extract
of SG inhibits proliferation, adhesion, migration and invasion of
ovarian cancer SKOV-3 cells. These antitumor activities of SG were
found to be mediated through inactivation of RTK signaling pathways
and downregulation of RTK and N-cadherin, as evidenced by using
pharmacological inhibitors such as PD98059, LY294002 and
rapamycin.
Overexpressed RTKs and dysregulation of RTK
signaling pathways contribute to ovarian cancer growth and
progression (4–8). These events are very complex and
closely associated with EMT, a process which is controlled by a
variety of signals from tumor microenvironment as well as up- and
down-regulation of EMT-related cellular markers (9–11).
Our results clearly show that SG treatment inhibits
mitogen-stimulated signaling pathways and expression of EGFR,
VEGFR-2, FGFR-1, and ILK as well as N-cadherin, which plays the
pivotal roles in the formation and stability of blood vessels
associated with cancer growth and progression (9), similar to previous reports
demonstrating that suppression of human EGFR-2 expression inhibits
cell proliferation, migration and invasion through upregulation of
epithelial marker E-cadherin and downregulation of mesenchymal
markers N-cadherin and vimentin in SKOV-3 cells (33). These findings suggest that
cross-talk between RTKs and integrins, and interactions of cellular
and ECM molecules coordinately function to regulate cancer
progression within tumor microenvironment. Therefore, further
understanding of the molecular mechanisms and identification of
targets in ovarian cancer progression are warranted to improve
therapeutic efficacy and reduce the incidence of drug resistance to
cancer therapy.
In conclusion, our findings provide important
insights into the roles and pharmacological efficacy of SG in
regulation of ovarian cancer growth and progression, and support
the development as a potent antitumor agent for the treatment of
ovarian cancer.
Acknowledgements
This study was supported by the research fund of
Dankook University in 2012.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar
|
|
2
|
Lemmon MA and Schlessinger J: Cell
signaling by receptor tyrosine kinases. Cell. 141:1117–1134. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hynes NE and Lane HA: ERBB receptors and
cancer: the complexity of targeted inhibitors. Nat Rev Cancer.
5:341–354. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Alper Ö, Bergmann-Leitner ES, Bennett TA,
Hacker NF, Stromberg K and Stetler-Stevenson WG: Epidermal growth
factor receptor signaling and the invasive phenotype of ovarian
carcinoma cells. J Natl Cancer Inst. 93:1375–1384. 2001.PubMed/NCBI
|
|
5
|
van der Bilt ARM, de Vries EGE, de Jong S,
Timmer-Bosscha H, van der Zee AGJ and Reyners AKL: Turning promise
into progress for antiangiogenic agents in epithelial ovarian
cancer. Crit Rev Oncol Hematol. 84:224–242. 2012.
|
|
6
|
Masoumi Moghaddam S, Amini A, Morris D and
Pourgholami M: Significance of vascular endothelial growth factor
in growth and peritoneal dissemination of ovarian cancer. Cancer
Metastasis Rev. 31:143–162. 2012.PubMed/NCBI
|
|
7
|
Cole C, Lau S, Backen A, et al: Inhibition
of FGFR2 and FGFR1 increases cisplatin sensitivity in ovarian
cancer. Cancer Biol Ther. 10:495–504. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gui T and Shen K: The epidermal growth
factor receptor as a therapeutic target in epithelial ovarian
cancer. Cancer Epidemiol. 36:490–496. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tiwari N, Gheldof A, Tatari M and
Christofori G: EMT as the ultimate survival mechanism of cancer
cells. Semin Cancer Biol. 22:194–207. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Thiery JP and Sleeman JP: Complex networks
orchestrate epithelial-mesenchymal transitions. Nat Rev Mol Cell
Biol. 7:131–142. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lee JM, Dedhar S, Kalluri R and Thompson
EW: The epithelial-mesenchymal transition: new insights in
signaling, development, and disease. J Cell Biol. 172:973–981.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bourboulia D and Stetler-Stevenson WG:
Matrix metalloproteinases (MMPs) and tissue inhibitors of
metalloproteinases (TIMPs): Positive and negative regulators in
tumor cell adhesion. Semin Cancer Biol. 20:161–168. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kang BK, Lee EH and Kim HM: Inhibitory
effects of Korean folk medicine ‘Hi-Chum’ on histamine release from
mast cells in vivo and in vitro. J Ethnopharmacol.
57:73–79. 1997.
|
|
15
|
Kim H-M, Lee J-H, Won J-H, et al:
Inhibitory effect on immunoglobulin E production in vivo and
in vitro by Siegesbeckia glabrescens. Phytother Res.
15:572–576. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim JY, Lim HJ and Ryu J-H: In
vitro anti-inflammatory activity of 3-O-methyl-flavones
isolated from Siegesbeckia glabrescens. Bioorg Med Chem
Lett. 18:1511–1514. 2008. View Article : Google Scholar
|
|
17
|
Li H, Kim JY, Hyeon J, Lee HJ and Ryu J-H:
In vitro antiinflammatory activity of a new Sesquiterpene
lactone isolated from Siegesbeckia glabrescens. Phytother
Res. 25:1323–1327. 2011.
|
|
18
|
Jun SY, Choi YH and Shin HM:
Siegesbeckia glabrescens induces apoptosis with different
pathways in human MCF-7 and MDA-MB-231 breast carcinoma cells.
Oncol Rep. 15:1461–1467. 2006.
|
|
19
|
Cho Y-R, Kim SH, Ko HY, Kim M-D, Choi SW
and Seo D-W: Sepiapterin inhibits cell proliferation and migration
of ovarian cancer cells via down-regulation of
p70S6K-dependent VEGFR-2 expression. Oncol Rep.
26:861–867. 2011.PubMed/NCBI
|
|
20
|
Seo D-W, Saxinger WC, Guedez L, Cantelmo
AR, Albini A and Stetler-Stevenson WG: An integrin-binding
N-terminal peptide region of TIMP-2 retains potent angio-inhibitory
and anti-tumorigenic activity in vivo. Peptides.
32:1840–1848. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim SH, Cho Y-R, Kim H-J, et al:
Antagonism of VEGF-A-induced increase in vascular permeability by
an integrin α3β1-Shp-1-cAMP/PKA pathway. Blood. 120:4892–4902.
2012.PubMed/NCBI
|
|
22
|
Yoon HJ, Cho Y-R, Joo J-H and Seo D-W:
Knockdown of integrin α3β1 expression induces proliferation and
migration of non-small cell lung cancer cells. Oncol Rep.
29:662–668. 2013.
|
|
23
|
Kim SH, Cho Y-R, Kim M-D, Kim HJ, Choi SW
and Seo D-W: Inhibitory effects of sepiapterin on vascular
endothelial growth factor-A-induced proliferation and adhesion in
human umbilical vein endothelial cells. Arch Pharm Res.
34:1571–1577. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cho Y-R, Choi S and Seo D-W: Sepiapterin
regulates cell proliferation and migration: its association with
integrin α3β1 and p53 in human lung cancer cells. Genes Genom.
33:577–582. 2011.
|
|
25
|
Bourboulia D, Jensen-Taubman S, Rittler
MR, et al: Endogenous angiogenesis inhibitor blocks tumor growth
via direct and indirect effects on tumor microenvironment. Am J
Pathol. 179:2589–2600. 2011. View Article : Google Scholar
|
|
26
|
Hong SY, Cho JY and Seo D-W: Ginsenoside
Rp1 inhibits proliferation and migration of human lung cancer
cells. Biomol Ther. 19:411–418. 2011. View Article : Google Scholar
|
|
27
|
Malumbres M and Barbacid M: Cell cycle,
CDKs and cancer: a changing paradigm. Nat Rev Cancer. 9:153–166.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mitra SK, Hanson DA and Schlaepfer DD:
Focal adhesion kinase: in command and control of cell motility. Nat
Rev Mol Cell Biol. 6:56–68. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Desgrosellier JS and Cheresh DA: Integrins
in cancer: biological implications and therapeutic opportunities.
Nat Rev Cancer. 10:9–22. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Slack-Davis JK, Atkins KA, Harrer C,
Hershey ED and Conaway M: Vascular cell adhesion molecule-1 is a
regulator of ovarian cancer peritoneal metastasis. Cancer Res.
69:1469–1476. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Eliceiri BP: Integrin and growth factor
receptor crosstalk. Circ Res. 89:1104–1110. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hannigan GE, McDonald PC, Walsh MP and
Dedhar S: Integrin-linked kinase: Not so ‘pseudo’ after all.
Oncogene. 30:4375–4385. 2011.
|
|
33
|
Lu Y-M, Rong M-L, Shang C, et al:
Suppression of HER-2 via siRNA interference promotes apoptosis and
decreases metastatic potential of SKOV-3 human ovarian carcinoma
cells. Oncol Rep. 29:1133–1139. 2013.PubMed/NCBI
|