Introduction
Hepatocellular carcinoma (HCC, also called malignant
hepatoma) is the most common among the various forms of liver
cancer. This tumor also has a variant type that consists of both
HCC and cholangiocarcinoma components. The risk of hepatocellular
carcinoma in type 2 diabetics is increased by 2.5 to 7.1 times
(1) that of the non-diabetic risk
depending on the duration of diabetes and treatment protocol, and
despite the efforts of scientists and clinicians, the mechanism is
not clear.
The novel cancer suppressor, resveratrol is a
stilbenoid, which is a type of natural phenol and phytoalexin
produced naturally by several plants. Resveratrol is currently the
topic of numerous animal and human studies, and although its
anticancer effects have been reported, the underlying mechanism
remains to be elucidated (2).
During recent research, we found that resveratrol inhibits multiple
myeloma cell proliferation and induces apoptosis through
downregulation of STAT3 (3), which
is a member of the STAT signaling protein family (4). In response to cytokines and growth
factors, STAT family members are phosphorylated by
receptor-associated kinases and then form homodimers or
heterodimers that translocate to the cell nucleus, where they act
as transcription activators. STAT3 is activated through
phosphorylation of tyrosine 705 in response to various cytokines
and growth factors including interferons, epidermal growth factor,
interleukin (IL)-5, IL-6, hepatocyte growth factor, leukemia
inhibitory factor (LIF), bone morphogenetic protein 2 and also the
hormone leptin. STAT3 regulates the expression of a variety of
genes in response to cell stimuli, and thus plays a key role in
many cellular processes such as cell growth and apoptosis (5). The downregulation of STAT3 in
resveratrol-treated medulloblastoma cells confirmed the growth
inhibition effects of this drug (6).
Resveratrol can induce the expression of SIRT1,
which is a longevity protein. SIRT1 can inhibit the proliferation
of pancreatic cancer cells (7), and
the downregulation of SIRT1 in hAT MSCs induces senescence and
inhibits cell proliferation (8).
STAT3 phosphorylation and function in the liver were found to be
tightly regulated by the nutritional status of an animal, through
SIRT1-mediated deacetylation of key STAT3 lysine sites (9). Yet, the relationship among
resveratrol, SIRT1 and STAT3 in HepG2 cells is not clear.
In the present study, we found that resveratrol
suppressed proliferation and cell viability of high glucose-exposed
HepG2 cells. STAT3 was constitutively expressed in human HepG2
cells under a high glucose condition; therefore, we investigated
the effects of resveratrol on the proliferation and invasive
activity of HepG2 cells. The results showed that high glucose
induced increased expression of STAT3 and that resveratrol
suppressed the proliferation and invasive activity of high
glucose-exposed HepG2 cells. Furthermore, resveratrol suppressed
the expression of MMP-9, VEGF and cyclin B1, but not cyclin D1.
These data demonstrate that resveratrol suppresses the
proliferation of high glucose-exposed HepG2 cells and indicate that
this effect is mediated via the STAT3 signaling pathway. The
present study also demonstrated that the effect of resveratrol on
the STAT3 and AKT signaling pathways was SIRT1-dependent.
Materials and methods
Materials
Resveratrol (0.1% DMSO) was purchased from Sigma
(St. Louis, MO, USA), and the MTT cell proliferation and
cytotoxicity assay kit was purchased from Sangon Biotech (Shanghai,
China). The Matrigel invasion chamber was purchased from
Becton-Dickinson Labware (Bedford, MA, USA). Antibodies against
STAT3 and STAT3 phosphorylated at tyrosine 705 and ser727, AKT and
AKT phosphorylated at tyrosine 473, SIRT1 and the SIRT1 suppressor
EX527 were purchased from Cell Signaling Technology (Danvers, MA,
USA); secondary antibodies were purchased from Beijing Biosynthesis
Biotechnology (Beijing, China). Penicillin, streptomycin, DMEM, and
fetal bovine serum (FBS) were obtained from Gibco (Grand Island,
NY, USA).
Cell culture
HepG2 cells, which belong to the classic subclass of
human HCC, were obtained from the American Type Culture Collection
(ATCC, Manassas, VA, USA). The HepG2 cells were maintained in DMEM
containing glucose (2.8, 5.5 and 25 mM) supplemented with 10% FBS,
L-glutamine and antibiotics (100 U/ml penicillin and 100 μg/ml
streptomycin) at 37°C in the presence of 5% CO2. The
effect of resveratrol and the role of SIRT1 in HepG2 cells cultured
in high glucose DMEM were investigated by the addition of
resveratrol (100 μM) and EX527 (40 μM).
Western blot analysis
Whole cell extracts of resveratrol-treated cells
were examined for expression of phospho-STAT3, STAT3, phospho-AKT,
AKT, SIRT1 and β-actin by western blot analysis as previously
described (3). The whole cell
extracts were prepared by lysing resveratrol-treated cells in lysis
buffer [20 mM Tris (pH 7.4), 250 mM NaCl, 2 mM EDTA (pH 8.0), 0.1%
Triton X-100, 0.01 mg/ml aprotinin, 0.005 mg/ml leupeptin, 0.4 mM
phenylmethylsulphonylfluoride and 4 mM sodium orthovanadate). The
protein concentration was quantified using BSA protein assay kits.
Equal amounts of protein (40 μg) were separated by 8–10%
Tris-glycine gel (Novex, Invitrogen, USA) electrophoresis. The
proteins were then electro-transferred to a nitrocellulose membrane
(Novex), blocked with 5% non-fat milk and probed with various
primary antibodies overnight at 4°C. The blots were then washed
three times (5 min per wash), and then exposed to horseradish
peroxidase-conjugated secondary antibodies for 2 h.
Immunoreactivity was finally examined by an enhanced
chemiluminescence (ECL) reagent (Thermo Fisher Scientific, Waltham,
MA, USA). The band for each protein was then quantified by
densitometry using ImageJ software (version 1.41; NIH, Bethesda,
MD, USA) and normalized to the expression of β-actin for protein
loading.
Reverse transcription-PCR analysis
Total RNA was isolated from HepG2 cells after 24 h
of treatment with resveratrol using TRIzol® reagent
(Life Technologies, Rockville, MD, USA) following the
manufacturer’s protocol. cDNA was synthesized from 1 μg of total
RNA using a cDNA synthesis kit (Takara Biotechnology Co., Ltd.,
Dalian, China) following the manufacturer’s instructions. Primer
sequences (Genencor, Shanghai, China) specific for MMP-9, STAT3,
cyclin B1, cyclin D1 and VEGF are shown in Table I. β-actin was used to normalize the
cDNA input levels. After cDNA synthesis, the PCR reaction consisted
of 32 cycles of denaturation at 95°C for 30 sec, annealing at 56°C
for 30 sec, extension at 72°C for 30 sec, and a further 5 min at
72°C in the last cycle. PCR products were separated by
electrophoresis on a 2% agarose gel and visualized by staining with
ethidium bromide. Each sample was analyzed in three biological
replicates, and at least three reactions were used to calculate the
expression. The relative expression was quantified
densitometrically using the Gel Image Ver. 3.74 System (Tianon,
Shanghai, China).
 | Table IOligonucleotides used in the PCR
assay. |
Table I
Oligonucleotides used in the PCR
assay.
| Name | Sequence (5′-3′) | Product (bp) |
|---|
| homo_MMP9_F |
ACACCGACGACCGGTTTGGC | |
| homo_MMP9_R |
TCGAGTCAGCTCGGGTCGGG | 219 |
| homo_actin_F |
AGCCTCGCCTTTGCCGATCC | |
| homo_actin_R |
ACATGCCGGAGCCGTTGTCG | 100 |
| homo_cyclin B1_F |
GCAGCACCTGGCTAAGAATGT | |
| homo_cyclin B1_R |
GCCTTGGCTAAATCTTGAACT | 147 |
| homo_cycllin
D1_F |
GCGAGGAACAGAAGTGCG | |
| homo_cycllin
D1_R |
AGGCGGTAGTAGGACAGGAA | 484 |
| home_VEGF_F1 |
ACGGACAGACAGACAGACACC | |
| home_VEGF_R1 |
CCCAGAAGTTGGACGAAAAGT | 176 |
Migration assay in vitro
Cell migration assays were performed in a Matrigel
chamber (Becton-Dickinson Labware). Briefly, 5×104 cells
in 800 μl of serum-free culture medium were added to the upper
chamber, while medium supplemented with 10% FBS and resveratrol was
added to the lower chamber. After incubation for 24 h, the cells
remaining on the upper side of the filters were removed with a
cotton swab. The cells that had migrated to the bottom surface of
the filter were fixed with 4% paraformaldehyde, and stained with
(0.1% H2O) crystal violet. Stained cells were visualized
using an Olympus IX71 microscope (Olympus, Tokyo, Japan).
Transient RNA interference and
transfections
SIRT1 was knocked down by using small siRNAs, with a
non-targeting siRNA used in parallel as a negative control
(GenePharma Co., Shanghai, China). Primary cultures were
transfected with siRNA SIRT1 or irrelevant siRNA using X-tremeGENE
HP DNA transfection reagent (Roche Diagnostics GmbH/Roche Applied
Science, Mannheim, Germany). After 2 days of mRNA silencing, HepG2
cells were analyzed for protein expression of SIRT1, phospho-STAT3,
STAT3, phospho-AKT, AKT and β-actin.
MTT assays in vitro
The antiproliferative effects of resveratrol on
HepG2 cells under a high glucose condition were determined by the
nicotinamide, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium
bromide (MTT) dye uptake method as previously described (10). Briefly, HepG2 were counted using a
hemocytometer and equally distributed in 96-well plates at a
density of 1×104 cells/well. Cells were treated with
resveratrol at the indicated concentrations (0, 10 and 100 μM) for
6, 12, 24 and 48 h. To determine cell viability, the DMEM (serum
10%) was removed, and cells were incubated with MTT (Sigma) at a
final concentration of 5 mg/ml in DMEM for 6 h in the dark at 37°C.
The supernatant was then removed from each well. The colored
formazan crystals produced from MTT were dissolved in 150 μl DMSO,
and cell viability was determined spectrophotometrically at 490 nm
(BioTek, Winooski, VT, USA).
Statistical analysis
All results are expressed as means ± SD relative to
the control value. Experiments were performed on three independent
occasions. Statistical comparisons between groups were analyzed by
t-test or ANOVA followed by Tukey’s post-hoc test as appropriate.
P<0.05 was considered to indicate a statistically significant
result. All data analysis was performed with the use of SPSS 13.0
statistical software.
Results
Resveratrol reduces the cell viability of
high glucose-exposed HepG2 cells
In the present study, we showed that a high glucose
concentration (25 mM) increased the proliferation of HepG2 cells
when compared to that of the control group (5.5 mM) (Fig. 1A). Furthermore, under a high glucose
condition, resveratrol reduced the viability of HepG2 cells at a
dose of 100 μM but not at 10 μM (Fig.
1B).
 | Figure 1(A) Effect of high glucose on the cell
viability of HepG2 cells. HepG2 cells were pretreated with DMEM
containing glucose [2.8 (LG), 5.5 (NG) and 25 mM (HG)]. After
incubation for 12, 24 and 48 h at 37°C, cell viability was
determined by the MTT reduction assay as described in Materials and
methods. (B) Effect of resveratrol on cell viability in high
glucose-exposed HepG2 cells. After pretreatment with high glucose
DMEM (25 mM) for 12 h, and then resveratrol (10 and 100 μM, 0.1%
DMSO) for 6, 12, 24 and 48 h at 37°C, cell viability was determined
by the MTT reduction assay. Data are the mean ± SD values for three
independent experiments (*P<0.05 compared with the
control). LG, low glucose; NG, normal glucose; HG, high
glucose. |
Resveratrol suppresses the proliferation
and invasive capacity of HepG2 cells
In order to elucidate the relationship between
resveratrol and STAT3 in HepG2 cells under a high glucose
condition, we analyzed the effect of resveratrol on the
proliferation and invasive capacity of high glucose-exposed HepG2
cells. Resveratrol (100 and 200 μM) suppressed the proliferation of
high glucose-exposed HepG2 cells in a dose-dependent manner
(Fig. 2A and B). Compared with the
control DMSO group, resveratrol (100 and 200 μM) also suppressed
the invasive capacity of the HepG2 cells under a high glucose
condition (Fig. 2C and D).
Resveratrol suppresses the expression of
p-STAT3 in high glucose-exposed HepG2 cells
STAT3 can promote oncogenesis by being
constitutively active in various signaling pathways (11,12).
Furthermore, STAT3 suppression inhibits human HepG2 cell
proliferation in vitro(13).
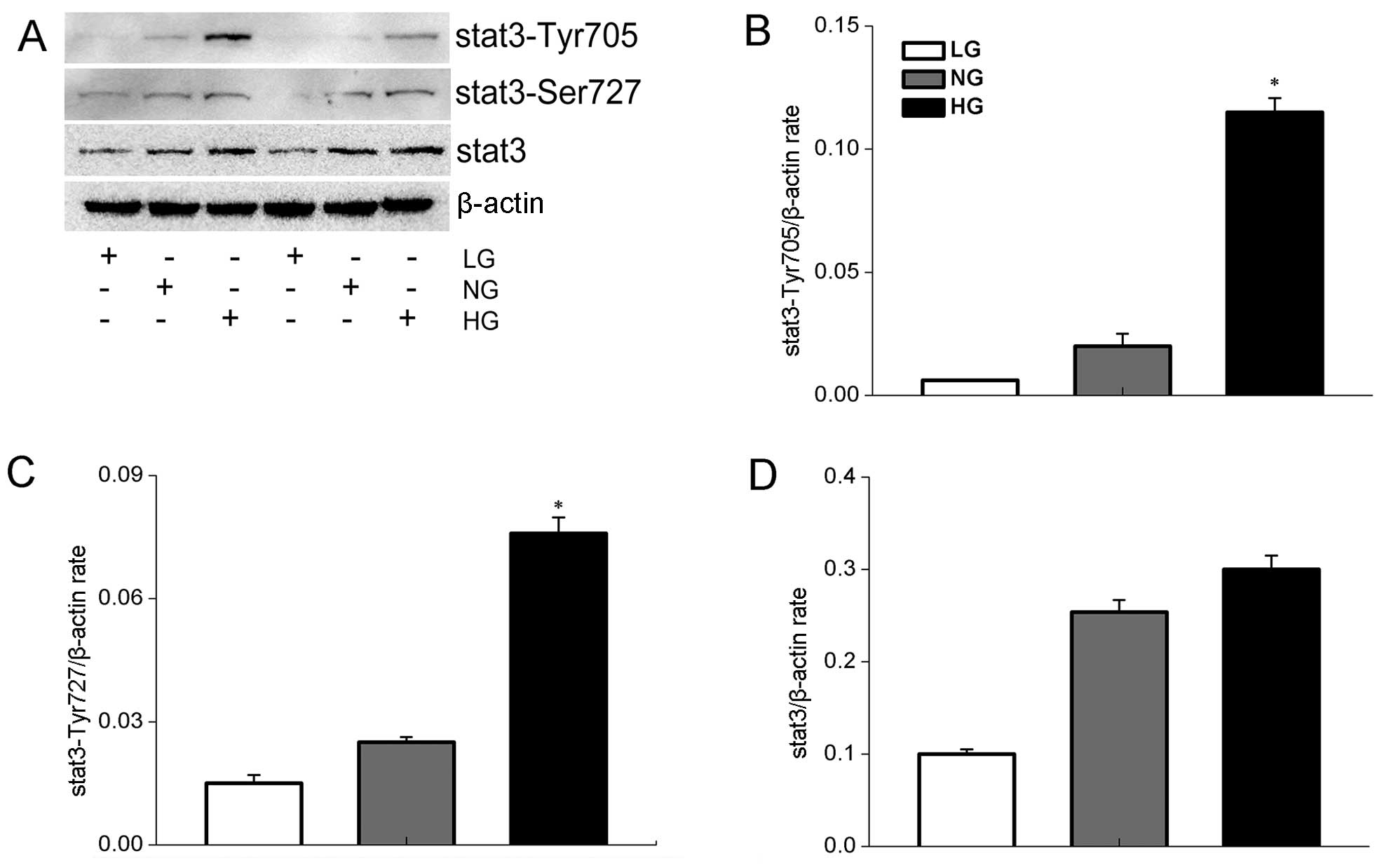
In the present study, we showed that expression of STAT3 and
phospho-STAT3 was induced in high glucose-exposed HepG2 cells, with
expression of the phosphorylated form increasing with glucose
levels. Thus, phospho-STAT3 expression was induced by a high
glucose condition (Fig. 3A).
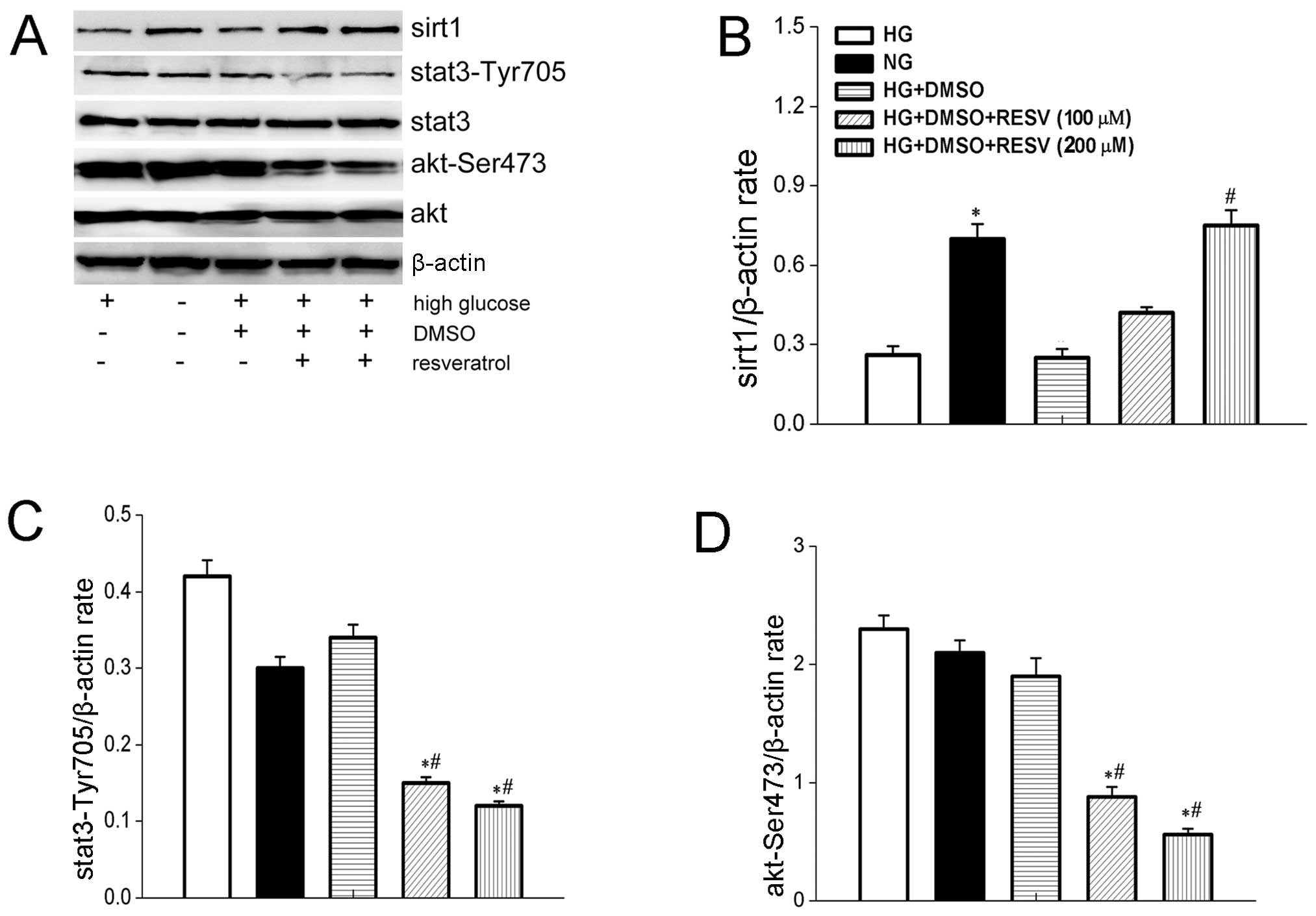
Investigation of the effect of resveratrol on the
STAT3 signaling pathway revealed that resveratrol downregulated the
expression of p-STAT3-Tyr705 in a dose-dependent manner, but had no
effect on the total level of STAT3 in HepG2 cells (Fig. 4A).
Resveratrol suppresses the expression of
p-STAT3-related genes associated with cell proliferation and
migration
The effect of resveratrol (100 and 200 μM) on
p-STAT3-related genes associated with cell proliferation and
migration, MMP-9, cyclin B1, cyclin D1 and VEGF, was investigated
by reverse transcription-PCR. Analysis of HepG2 cells cultured in
the presence of resveratrol (100 and 200 μM) for 24 h showed that
the expression of MMP-9, cyclin B1 and VEGF was downregulated,
although cyclin B1 expression was unaffected (Fig. 5A).
Resveratrol induces the expression of
SIRT1 and suppresses the AKT signaling pathway
The expression of SIRT1 and p-AKT by HepG2 cells
under the influence of difference concentrations of resveratrol was
investigated by western blot analysis. Resveratrol induced the
overexpression of SIRT1 (Fig. 4A),
while the expression of p-AKT was downregulated (Fig. 4A). Both effects of resveratrol were
dose-dependent.
Role of EX527 in STAT3 and AKT signaling
and cell viability
We analyzed the role of SIRT1 in STAT3 and AKT
signaling by using the SIRT1-specific inhibitor, EX527. SIRT1
expression in HepG2 cells was inhibited in a dose-dependent manner
by pretreatment with EX527 (10, 20 and 40 μM) for 30 min (Fig. 6A). Furthermore, EX527 (40 μM)
suppressed the effects of resveratrol on STAT3 and AKT signaling;
it weakened the effect of resveratrol function on p-STAT3 and p-AKT
(Fig. 7A). EX527 also partially
abrogated the suppression of resveratrol on HepG2 cell viability
under a high glucose condition (Fig.
7D).
Effect of siRNA-SIRT1 on the STAT3 and
AKT signaling pathways and cell viability
To investigate the role of SIRT1 in the effects of
resveratrol on the STAT3 and AKT signaling pathways we used
siRNA-SIRT1 (10 μM) to downregulate the expression of SIRT1 at the
level of transcription (Fig. 8A).
We found that siRNA-SIRT1 weakened the suppressive function of
resveratrol on the STAT3 and AKT signaling pathways. Furthermore,
siRNA-SIRT1 exhibited an effect on the cell viability of HepG2
cells under the function of resveratrol. These observations
indicated that the effects of resveratrol on the STAT3 and AKT
signaling pathways and cell viability in HepG2 cells under
conditions of high glucose were partially SIRT1-dependent (Fig. 8A and E).
Discussion
The risk of hepatocellular carcinoma, which is the
most common type of liver cancer, is increased in type 2 diabetic
patients, although the reason for this is not clear (1). Resveratrol exerts important effect on
cancer cell proliferation; therefore, elucidation of the mechanism
of the effects of resveratrol in HepG2 cells under high glucose
conditions may provide important insights into the reasons why the
risk of hepatocellular carcinoma is greater in type 2 diabetic
patients.
In the present study, we showed that a high glucose
condition upregulated the expression of phospho-STAT3, and enhanced
the viability of HepG2 cells (Fig.
1). Resveratrol suppressed the proliferation and invasive
capacity of HepG2 cells and also suppressed phospho-STAT3 and
phospho-AKT expression. Furthermore, our data indicated that these
effects were partly mediated through SIRT1. Thus, the present study
indicated that resveratrol plays an important role in controlling
the proliferation and invasion of high glucose-exposed HepG2
cells.
Resveratrol is a stilbenoid that is naturally
produced by several plants. Although the capacity of resveratrol to
prevent cancer development has been studied for many years
(14), the mechanism underlying the
efficacy of resveratrol, even at massive doses, remains to be fully
elucidated (15). Recent research
has shown that resveratrol suppresses proliferation and induces
apoptosis in medulloblastoma cells by inhibition of STAT3 signaling
and suppression of the expression of a range of cancer-associated
genes (6). The effect of
resveratrol on cancer has been demonstrated in other studies
(16,17) on malignant cells and medulloblastoma
cells. These studies also indicate that the transcription factor
STAT3 is regulated by resveratrol. STAT3, which plays an important
role in cancer development, is a member of the STAT3 protein
family. In response to cytokines and growth factors, STAT family
members are phosphorylated by receptor-associated kinases and then
form homodimers or heterodimers that translocate to the cell
nucleus, where they act as transcription activators. In recent
research, STAT3 was shown to have an oncogenic or tumor suppressor
role depending on the mutational background of tumors. Evidence
indicates that chronic STAT3 activation is a key event in gastric
cancer induction and progression (18). Furthermore, increased STAT3 activity
can upregulate the survival signal in cancer cells (19) and specific inhibition of STAT3 is a
useful cancer treatment approach (20–22).
In the present study, we found that resveratrol suppressed the
expression of STAT3 and inhibited the proliferation of cancer cells
although the function of resveratrol on the STAT3 signaling pathway
in HepG2 cells and the effects of a high glucose concentration on
the expression of STAT3 have not been investigated to date. In the
present study, we found that resveratrol suppressed the STAT3
signaling pathway and inhibited the proliferation of high
glucose-exposed HepG2 cells (Figs.
2, 4 and 5).
In the present study, we found that resveratrol
induced the expression of SIRT1, which was the first sirtuin gene
to be identified. Subsequently, other members of this highly
conserved family have been found in nearly all organisms studied
(23), SIRT2 is the only Class III
histone deacetylase (HDAC) in budding yeast (24). The HDAC activity of SIRT2 results in
tighter packaging of chromatin and a reduction in transcription at
the targeted gene locus. Recently, the STAT3-dependent effects of
IL-22 in human keratinocytes were found to be counter-regulated by
sirtuin SIRT1 through a direct inhibition of STAT3 acetylation
(25). Furthermore, SIRT1 gene
knock-out murine embryonic fibroblast (MEF) cells have been shown
to induce expression of STAT3-tyr705 (26). These observations indicate that the
STAT3 signaling pathway is regulated by SIRT1. The connection
between acetylation and phosphorylation of STAT3 implies that STAT3
may have an important role in other cellular processes that involve
SIRT1 (9). However, the
relationship of SIRT1 and STAT3 in HepG2 cells has not been
reported to date, and whether SIRT1 is involved in the effects of
resveratrol on the suppression of STAT3 has never been proven. In
the present study, we demonstrated that the SIRT1 suppressor EX527
and siRNA-SIRT1 influenced the expression of SIRT1, indicating that
SIRT1 is involved in the effects of resveratrol on the suppression
of p-STAT3 and p-AKT as well as in the proliferation of HepG2 cells
under a high glucose condition.
During our research, we found that resveratrol (100
μM) suppressed the proliferation of HepG2 cells. In recent years,
researchers have also found other factors that affect the
proliferation of HepG2 cells, such as β(2)-AR agonists (R,R′)-fenoterol (Fen) and
(R,R′)-4-methoxy-1-naphthylfenoterol (MNF), BBR and THP which
inhibit growth (25,27). There are many reports concerning the
control of HepG2 cell proliferation, and the mechanism is complex.
We hypothesize that the findings of the present study may help
explain the increased risk of hepatocellular carcinoma in type 2
diabetic patients in whom glucose levels are much higher than
normal and also provide insights into the mechanism by which
resveratrol controls HepG2 cell proliferation. We also provided
evidence clarifying the relationship between resveratrol, STAT3 and
SIRT1 and demonstrated that the effect of resveratrol on the STAT3
signaling pathway is partly mediated via SIRT1 signaling.
In summary, resveratrol is a new cancer suppressor
in high glucose-exposed HepG2 cells, and its mechanism may involve
the STAT3 and AKT signaling pathways. Our observations may explain
the increased risk of hepatocellular carcinoma in type 2 diabetic
patients. However, the mechanism by which resveratrol controls
HepG2 cell proliferation and the relationship of SIRT1 and STAT3 in
this process warrants further research.
Acknowledgements
The present study was supported by grants from the
National Natural Science Foundation of China (#81070110 to M.W) and
Shanghai Science and Tecnology Innovation Research Program
(#11410701900 to M.W). We would like to thank Jianping Tao and
Yanwei Qing for their technical support.
References
|
1
|
El-Serag HB, Hampel H and Javadi F: The
association between diabetes and hepatocellular carcinoma: a
systematic review of epidemiologic evidence. Clin Gastroenterol
Hepatol. 4:369–380. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zheng M, Chen R, Zhong H, et al:
Side-effects of resveratrol in HepG2 cells: reduced pten and
increased bcl-xl mRNA expression. Mol Med Rep. 6:1367–1370.
2012.PubMed/NCBI
|
|
3
|
Bhardwaj A, Sethi G, Vadhan-Raj S, et al:
Resveratrol inhibits proliferation, induces apoptosis, and
overcomes chemoresistance through down-regulation of STAT3 and
nuclear factor-κB-regulated antiapoptotic and cell survival gene
products in human multiple myeloma cells. Blood. 109:2293–2302.
2007.PubMed/NCBI
|
|
4
|
Capiralla H, Vingtdeux V, Venkatesh J, et
al: Identification of potent small-molecule inhibitors of STAT3
with anti-inflammatory properties in RAW 264.7 macrophages. FEBS J.
279:3791–3799. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Haghikia A, Stapel B, Hoch M and
Hilfiker-Kleiner D: STAT3 and cardiac remodeling. Heart Fail Rev.
16:35–47. 2011. View Article : Google Scholar
|
|
6
|
Wen S, Li H, Wu ML, et al: Inhibition of
NF-κB signaling commits resveratrol-treated medulloblastoma cells
to apoptosis without neuronal differentiation. J Neurooncol.
104:169–177. 2011.
|
|
7
|
Cho IR, Koh SS, Malilas W, et al: SIRT1
inhibits proliferation of pancreatic cancer cells expressing
pancreatic adenocarcinoma up-regulated factor (PAUF), a novel
oncogene, by suppression of β-catenin. Biochem Biophys Res Commun.
423:270–275. 2012.PubMed/NCBI
|
|
8
|
Kim YJ, Hwang SH, Lee SY, et al:
miR-486-5p induces replicative senescence of human adipose
tissue-derived mesenchymal stem cells and its expression is
controlled by high glucose. Stem Cells Dev. 21:1749–1760. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nie Y, Erion DM, Yuan Z, et al: STAT3
inhibition of gluconeogenesis is downregulated by SirT1. Nat Cell
Biol. 11:492–500. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li T, Wang W, Chen H, Li T and Ye L:
Evaluation of anti-leukemia effect of resveratrol by modulating
STAT3 signaling. Int Immunopharmacol. 10:18–25. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pawlus MR, Wang L and Hu CJ: STAT3 and
HIF1alpha cooperatively activate HIF1 target genes in MDA-MB-231
and RCC4 cells. Oncogene. Apr 22–2013.(Epub ahead of print).
|
|
12
|
Li J, Cui G, Sun L, et al: STAT3
acetylation-induced promoter methylation is associated with
downregulation of the ARHI tumor-suppressor gene in ovarian cancer.
Oncol Rep. 30:165–170. 2013.PubMed/NCBI
|
|
13
|
Absood A, Hu B, Bassily N and Colletti L:
VIP inhibits human HepG2 cell proliferation in vitro. Regul Pept.
146:285–292. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Baur JA and Sinclair DA: Therapeutic
potential of resveratrol: the in vivo evidence. Nat Rev Drug
Discov. 5:493–506. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Athar M, Back JH, Tang X, et al:
Resveratrol: a review of preclinical studies for human cancer
prevention. Toxicol Appl Pharmacol. 224:274–283. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kotha A, Sekharam M, Cilenti L, et al:
Resveratrol inhibits Src and Stat3 signaling and induces the
apoptosis of malignant cells containing activated Stat3 protein.
Mol Cancer Ther. 5:621–629. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yu LJ, Wu ML, Li H, et al: Inhibition of
STAT3 expression and signaling in resveratrol-differentiated
medulloblastoma cells. Neoplasia. 10:736–744. 2008.PubMed/NCBI
|
|
18
|
Giraud AS, Menheniott TR and Judd LM:
Targeting STAT3 in gastric cancer. Expert Opin Ther Targets.
16:889–901. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Deng J, Liu Y, Lee H, et al: S1PR1-STAT3
signaling is crucial for myeloid cell colonization at future
metastatic sites. Cancer Cell. 21:642–654. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tkach M, Coria L, Rosemblit C, et al:
Targeting Stat3 induces senescence in tumor cells and elicits
prophylactic and therapeutic immune responses against breast cancer
growth mediated by NK cells and CD4+ T cells. J Immunol.
189:1162–1172. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sen M, Joyce S, Panahandeh M, et al:
Targeting Stat3 abrogates EGFR inhibitor resistance in cancer. Clin
Cancer Res. 18:4986–4996. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang CL, Liu YY, Ma YG, et al: Curcumin
blocks small cell lung cancer cell migration, invasion,
angiogenesis, cell cycle and neoplasia through Janus kinase-STAT3
signalling pathway. PLoS One. 7:e379602012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Frye RA: Phylogenetic classification of
prokaryotic and eukaryotic Sir2-like proteins. Biochem Biophys Res
Commun. 273:793–798. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chang KT and Min KT: Regulation of
lifespan by histone deacetylase. Ageing Res Rev. 1:313–326. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sestito R, Madonna S, Scarponi C, et al:
STAT3-dependent effects of IL-22 in human keratinocytes are
counterregulated by sirtuin 1 through a direct inhibition of STAT3
acetylation. FASEB J. 25:916–927. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bernier M, Paul RK, Martin-Montalvo A, et
al: Negative regulation of STAT3 protein-mediated cellular
respiration by SIRT1 protein. J Biol Chem. 286:19270–19279. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen X, Cao Y, Lv D, Zhu Z, Zhang J and
Chai Y: Comprehensive two-dimensional HepG2/cell membrane
chromatography/monolithic column/time-of-flight mass spectrometry
system for screening anti-tumor components from herbal medicines. J
Chromatogr A. 1242:67–74. 2012. View Article : Google Scholar
|