Introduction
Cervical cancer is the third most common malignancy
in women of reproductive age, and the incidence just succeeds that
of breast cancer and colorectal cancer (1). In the past few decades, the cervical
cancer-related morbidity and mortality in young adult women have
increased. There are many factors which contribute to cervical
carcinogenesis. The relationship between cervical cancer and
persistent infection with HPV has been well established (2,3). In
recent years, emerging evidence suggests that cancer stem cells
(CSCs) are a rare group of undifferentiated tumorigenic cells which
are considered to be a renewable source of tumor cells and a source
of drug resistance leading to tumor recurrence, metastasis and
tumor progression (4,5). Understanding the molecular mechanisms
governing the initiation, progression and metastasis of cervical
cancer are important for the prevention, detection and treatment of
this prevailing disease.
Hiwi, a human member of the piwi family, maps to
chromosome 12q24.33, and belongs to the piwi-domain proteins, which
are components of ribonucleoprotein complexes. Hiwi plays an
important role in stem cell self-renewal, gametogenesis, RNA
silencing and translational regulation (6,7).
Expression of Hiwi has been found dysregulated in pancreatic
carcinoma, seminomas, gastric carcinomas and soft-tissue sarcoma,
and is described as an indicator of poor prognosis (8–12).
Moreover, recent data suggest that Hiwi is required to maintain the
stemness of hematopoietic stem cells (13). Thus, it has been proposed that Hiwi
is a key regulator in the maintenance of cancer stem cell
populations as well (8,14).
In the present study, we examined the expression of
Hiwi in cervical cancer to investigate the relationship between the
deregulation of Hiwi expresion and cervical carcinogenesis, and
then provide a necessary experimental and theoretical basis for the
diagnosis and therapeutics of cervical cancer.
Materials and methods
Ethics statement
Female BALB/c nude mice (4–6-weeks old) were
supplied by the Experimental Animal Center of Xi’an Jiaotong
University, China. This study was carried out in accordance with
the recommended guidelines for the care and use of laboratory
animals issued by the Chinese Council on Animal Research. The
protocol was approved by the Ethics Committee of Xi’an Jiaotong
University.
Cell lines and cell culture
Human cervical cancer cell lines HeLa, SiHa and C33A
used in this study were purchased from the American Type Culture
Collection (ATCC; Manassas, VA, USA) and were cultured in
Dulbecco’s modified Eagle’s medium (DMEM) (Sigma, St. Louis, MO,
USA) supplemented with 10% fetal bovine serum (FBS) and maintaining
at 37°C in an atmosphere containing 5% CO2.
Tissue collection
All of the archived formalin-fixed,
paraffin-embedded tissue specimens were collected at the Second
Hospital of Xi’an Jiaotong University (15). Before the collection of these
clinical materials, all participating patients provided written
informed consent. Histopathologic diagnosis and malignant
classification were determined by 2 pathologists in a blinded
manner and were based on the International Federation of Gynecology
and Obstetrics classification system.
Immunohistochemistry (IHC) and
immunocytochemistry (ICC)
Slides were prepared from formalin-fixed and
paraffin-embedded tissues and were stained for Hiwi. A standard
immunostaining procedure was performed using the anti-Hiwi antibody
(1:100; Santa Cruz Biotechnology, Santa Cruz, CA, USA). Briefly,
sections were successively deparaffinized and rehydrated, followed
by treatment with 10 mM sodium citrate buffer (pH 6.0) in a steam
pressure cooker to retrieve the endogenous antigen, and treated
with 3% H2O2, and incubated with the primary
antibody overnight at 4°C. Then the sections were incubated with
biotinylated secondary antibody for 30 min at room temperature,
followed by diaminobenzidine development. All slides were examined
under an Olympus-CX31 microscope (Olympus, Tokyo, Japan).
For detection of the expression of Hiwi in cells,
the cells were seeded on coverslips for 48 h, fixed with 4%
paraformaldehyde for 30 min, followed by 0.3% Triton X-100
permeabilization for 20 min at room temperature. Then
immunohistochemistry was carried out as described above.
Immunohistochemical results of Hiwi expression in
the cervical tissues were evaluated by 3 investigators
independently and scoring was determined by the percentage of
Hiwi-positive cells (1, 0–25% positive cells; 2, 26–50% positive
cells; 3, >50% positive cells) and the staining intensity (0, no
staining; 1, weak staining; 2, moderate staining; 3, strong
staining).
Western blot analysis
Cells were lysed with ice-cold lysis buffer with a
cocktail of protease and phosphatase inhibitors (Complete Mini;
Roche Diagnostics, Branchburg, NJ, USA). Protein samples (20 μg)
underwent electrophoresis on 10% sodium dodecyl sulfate
polyacrylamide gel electrophoresis (SDS-PAGE) and were transferred
onto polyvinylidene fluoride (PVDF) membranes. The membranes were
blocked with 5% fat-free milk in Tris-buffered saline and then
incubated with anti-Hiwi or anti-β-actin (both from Santa Cruz
Biotechnology, at 1:1,000) at 4°C overnight. After washing, the
membranes were incubated with their associated horseradish
peroxidase (HRP)-conjugated secondary antibody at the appropriate
dilution and then visualized on X-ray film using an enhanced
chemiluminescence reagent (Millipore, Billerica, MA, USA).
Array dataset
The ‘Cervical cancer response to chemoradiotherapy’
dataset (GDS3017) was downloaded from http://www.ncbi.nlm.nih.gov/sites/GDSbrowser?acc=GDS3017.
This array is an analysis of 156 cervical cancer biopsy samples
from patients receiving radiotherapy alone or radiotherapy plus
concomitant chemotherapy with cisplatin (DDP). Results provide
insight into the molecular mechanisms underlying the therapeutic
response to DDP (16).
Plasmid construction and
transfection
The human Hiwi CDS fragment was amplified by PCR and
inserted into pIRSE2-eGFP to create the Hiwi overexpression vector
(pIRSE2-eGFP-Hiwi) and the pIRES2-eGFP vector was used as a
negative control. The plasmids expressing Hiwi-specific short
hairpin RNA (pGPU6/GFP-shHiwi) were designed and purchased from
GenePharm Company (Shanghai, China).
According to the manufacturer’s instructions, the
overexpression plasmids were transfected in the SiHa cells and
silencing vectors were transfected in HeLa cells using
Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA). After a 24-h
transfection, cells were passaged into DMEM with 10% FBS in the
presence of 1,000 μg/ml of G418 for 3 weeks. Individual
drug-resistant clones were selected, pooled, expanded, and
identified by western blotting.
Drug resistance and MTT assay
For drug resistance assays, cells were plated in
96-well plates at a density of 104 cells/well and
allowed to recover overnight before initiating drug treatments. The
cells were exposed to various concentrations of cisplatin (0, 3, 6,
12, 24 or 48 μg/ml for SiHa and HeLa cells) for 24 h, and cell
viability was measured by MTT. In separate experiments, the cells
were exposed to a constant concentration of cisplatin (3 μg/ml for
SiHa and HeLa cells) for 24, 48 or 72 h, and cell viability was
measured by MTT.
Following the manufacturer’s instructions, 20 μl of
MTT (Sigma) solution (5 mg/ml) was added to 200 μl of the culture
medium. The plates were then incubated for 4 h at 37°C. Following
the incubation, 150 μl dimethyl sulfoxide (DMSO; Sigma) was added
to each well for dye extraction. The dark-blue crystals of
MTT-formazan were thoroughly dissolved by shaking the plates at
room temperature for 10 min. Spectrometric absorbance at 490 nm was
measured by a microplate reader (Bio-Rad, Hercules, CA, USA).
Cell cycle analysis
Cell cycle distribution was analyzed by propidium
iodide (PI; Sigma) according to the manufacturer’s instructions.
Cells (1×106) were resuspended in PBS with 50 μg/ml PI
and 10 μg/ml RNase A, following fixation with 70% ethanol. Cells
were then detected for DNA content by flow cytometry and analyzed
using ModFit® LT Software (Verity Software House Inc.,
Topsham, ME, USA).
Real-time PCR
Total RNA from the cells was extracted with TRIzol
reagent (Invitrogen, Carlsbad, CA, USA) in accordance with the
manufacturer’s protocol. RNA concentration was determined for
reverse transcription-PCR. Then total cDNA was used as a template
for real-time PCR amplification using the SYBR-Green PCR kit
(Takara, Japan). The cycle threshold value was determined as the
point at which the fluorescence exceeded a preset limit determined
by the instrument’s software.
Tumorsphere formation assay
Cells were seeded in 6-well plates in 1 ml
serum-free DMEM/F12 medium supplemented with 20 ng/ml basic
fibroblastic growth factor (bFGF; PeproTech Inc., Rocky Hill, NJ,
USA), 20 ng/ml human recombinant epidermal growth factor (EGF;
PeproTech), N2 and B27 (Invitrogen). Fresh medium (0.5 ml) was
added to each well every 3 days. The spheres were counted by two
individuals in a blinded manner after 2–3-weeks of culture
(17,18).
Tumor xenograft experiment
Cells (105) were injected into the
subcutaneous tissue in the dorsum of BALB/c nude mice. Three
animals/group were used in each experiment. Engrafted mice were
monitored twice per week by visual observation and palpation for
the appearance of tumors over 12 weeks. The tumor volume (V) was
determined by the length (a) and width (b) as V = ab2/2
(19).
Statistical analysis
All experiments were repeated at least 3 separate
times. Data from all experiments were pooled, and the results were
expressed as mean ± SD. The t-test and log-rank test were performed
with the Statistical Package for Social Sciences (SPSS) 16.0
statistical software (SPSS Inc., Chicago, IL, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
Expression of Hiwi in the cervical
tissues
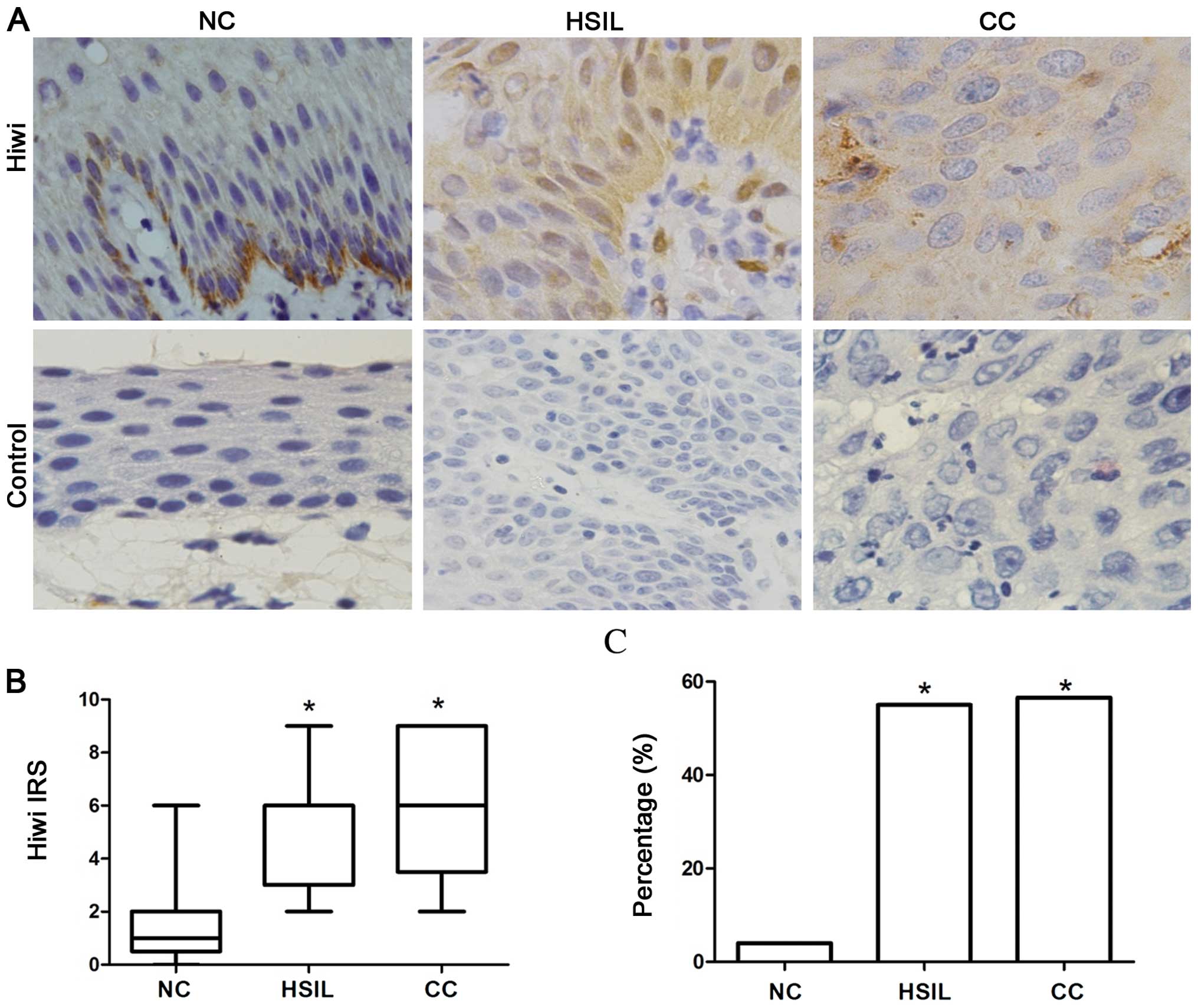
We performed immunohistochemistry to detect the
expression of Hiwi in 25 normal cervical tissues (NCs), 20
high-grade squamous intraepithelial lesions (HSILs) and 53 cervical
cancer (CC) tissues. Hiwi was expressed in most cells of the CC and
HSIL tissues but only in basal cells of the normal cervical
epithelia (Fig. 1A). In addition,
the IRS scores and the percentage of Hiwi-positive cases were
markedly elevated in the HSILs and CCs when compared to these
parameters in the NCs (Fig. 1B and
C, P<0.05), showing a positive correlation with tumor
progression.
To further investigate the relationship between Hiwi
expression and clinical disease progression, the results showed
that the percentage of cases with positive Hiwi expression was
higher in the advanced stage II–III cancers (25/37, 67.57%) than
the frequency of cases with positive expression in stage I cancers
(5/16, 31.25%) (Table I,
P<0.05). Thus, we speculated that Hiwi expression is
significantly correlated with the stage of malignancy; however, no
significant correlations were found between Hiwi expression and
other clinical characteristics, including age and grade (Table I, P>0.05).
 | Table ICorrelation between Hiwi expression
and clinicopathologic parameters in patients with cervical
cancer. |
Table I
Correlation between Hiwi expression
and clinicopathologic parameters in patients with cervical
cancer.
| Variables | No. of patients | Hiwi IRS | P-value |
|---|
|
|---|
| Positive | % |
|---|
| Age, years | | | | >0.05 |
| <45 | 18 | 12 | 66.67 | |
| ≥45 | 35 | 18 | 51.43 | |
| Grade | | | | >0.05 |
| I | 7 | 4 | 57.14 | |
| II | 19 | 12 | 63.16 | |
| III | 27 | 14 | 51.85 | |
| Stage | | | | <0.05 |
| I | 16 | 5 | 31.25 | |
| II–III | 37 | 25 | 67.57 | |
Expression of Hiwi in cervical cancer
cell lines
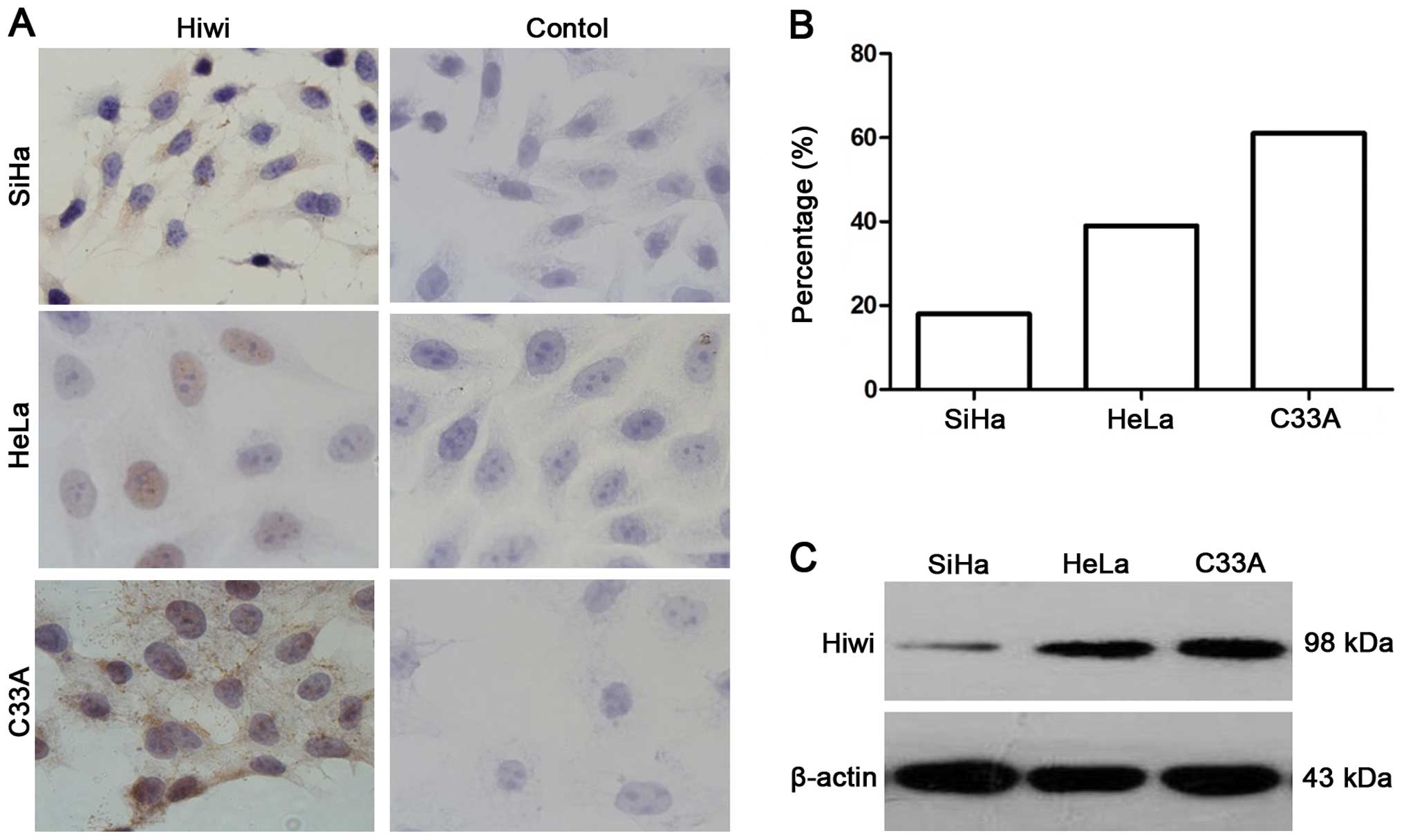
Endogenous Hiwi protein levels were examined by
western blotting and ICC in the three cervical cancer cell lines.
Immunocytochemical staining results showed that Hiwi was expressed
in the three cervical cancer cell lines and no immunostaining was
detected in the negative controls (Fig.
2A). The positive percentage of Hiwi expression in the SiHa,
HeLa and C33A cells was 16, 39 and 61%, respectively (Fig. 2B). Compared to the SiHa cells, the
expression of Hiwi in the C33A and HeLa cells was higher as
determined by western blot analysis (Fig. 2C).
Hiwi faciliates resistance to cisplatin
in cervical cancer
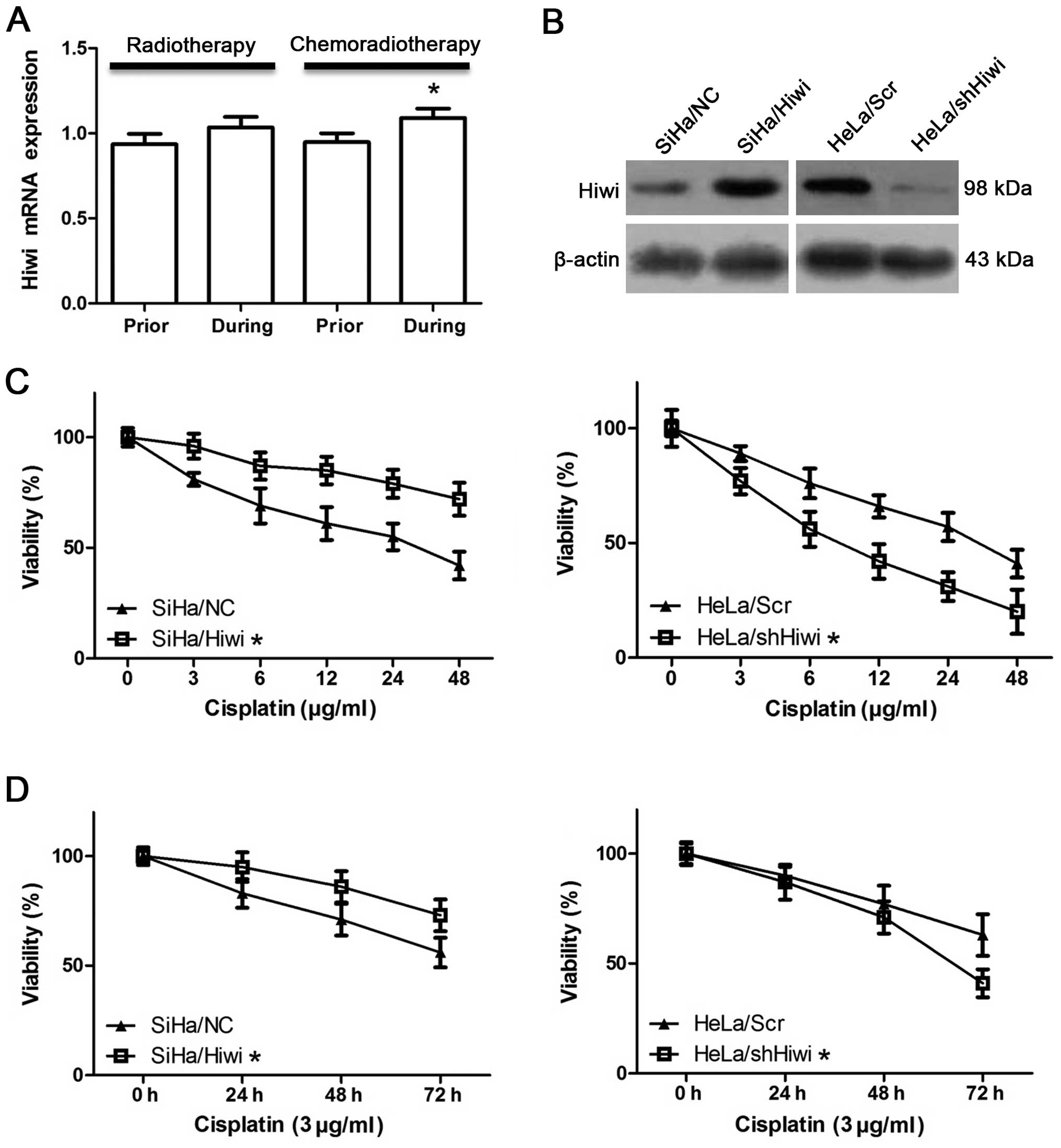
To investigate the role of Hiwi in cervical
carcinogenesis, we analyzed the expression of Hiwi in the response
to radiotherapy and chemoradiotherapy of cervical cancers. The
arrays showed that the mRNA expression of Hiwi was significantly
upregulated after chemoradiotherapy when compared to the level
before therapy (Fig. 3A,
P<0.05), while radiotherapy did not affect Hiwi expression
(Fig. 3A, P>0.05). This
indicated that Hiwi may induce resistance to chemotherapy drugs. In
order to investigate whether Hiwi influences the effects of
chemotherapy drugs, the Hiwi-overexpression vector or a negative
control vector was stably transfected in SiHa cells, and the
Hiwi-specific short hairpin interference RNA vector or a scramble
control vector was stably transfected in HeLa cells, respectively.
The efficiencies of transfection were validated by western blot
analysis (Fig. 3B). We then treated
the SiHa/Hiwi and HeLa/shHiwi cells with cisplatin (DDP), which is
one of the most commonly used chemotherapeutic drugs for the
treatment of cervical cancer. The effect of cisplatin (DDP) was
determined by MTT assay. After a 24-h treatment at different
concentrations, the viability of the SiHa/Hiwi cells was
significantly higher than that of the SiHa/NC cells, while it was
obviously lower in the HeLa/shHiwi cells when compared with that of
the HeLa/Scr cells (Fig. 3C,
P<0.05). Moreover, cell viability was also determined by MTT
assay after exposure to 3 μg/ml cisplatin for 24, 48, or 72 h. The
viability of the SiHa/Hiwi cells was significantly higher than that
of the SiHa/NC cells, while it was lower in the HeLa/shHiwi cells
than that in the HeLa/Scr cells (Fig.
3D, P<0.05). These results demonstrated that cisplatin
caused inhibition of viability in a dose-dependent and
time-dependent manner in both of the SiHa and HeLa cells suggesting
that Hiwi induces resistance to chemotherapy.
Hiwi induces stem cell characteristics in
cervical cancer
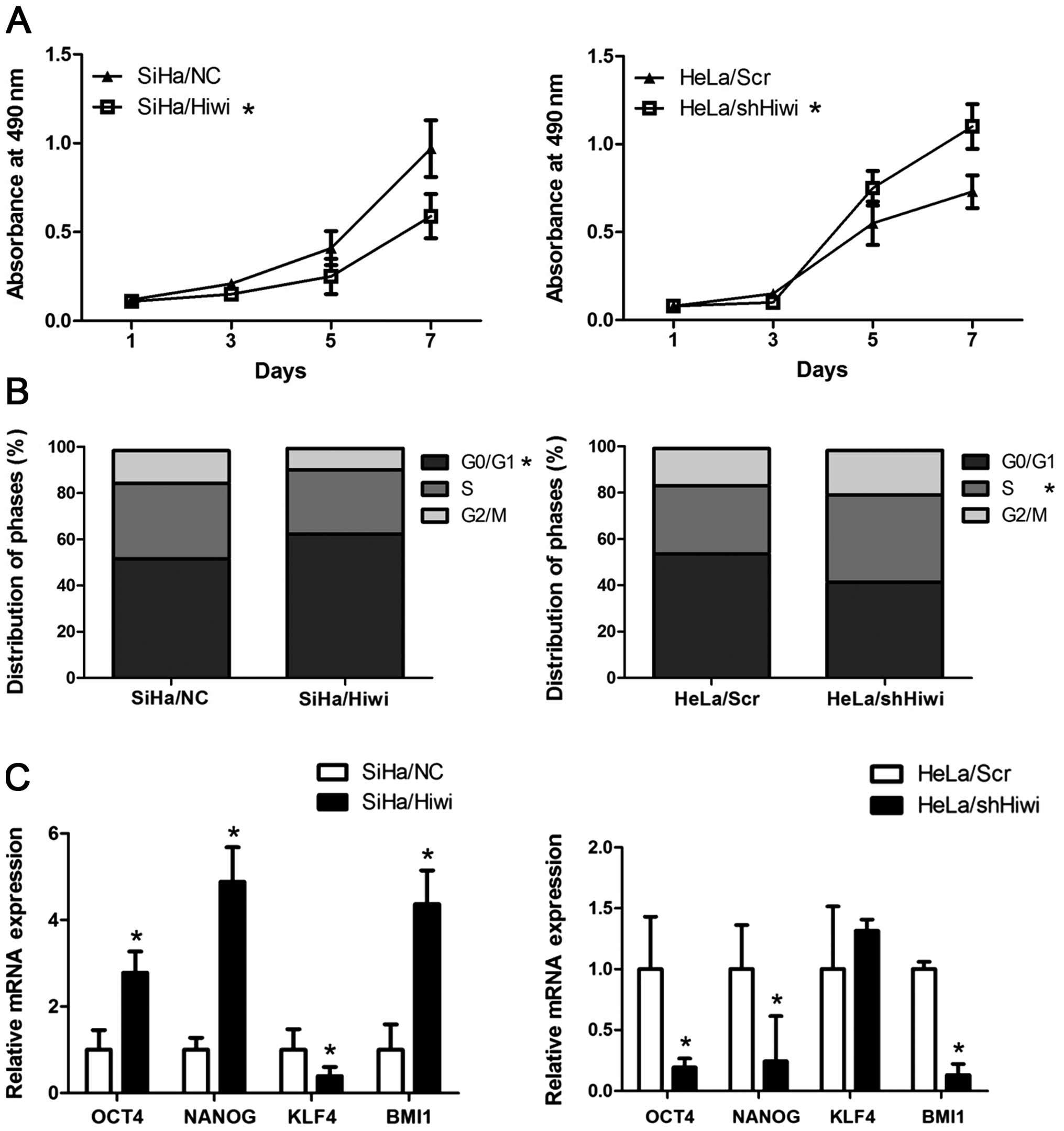
We further investigated the effect of Hiwi on the
growth and proliferation of cervical cancer. As a stem
cell-associated protein, ectopic Hiwi inhibited the cell viability
in SiHa cells, while silencing of Hiwi in HeLa cells significantly
promoted the cell viability as determined by MTT assay (Fig. 4A, P<0.05). In addition, ectopic
Hiwi arrested the cell cycle at the G0/G1
phase in SiHa/Hiwi cells (62.3±7.36%) when compared to SiHa/NC
cells (51.6±4.98%), while silencing of Hiwi accelerated the cell
cycle into the S phase in the HeLa/shHiwi cells (37.7±5.10) when
compared to the HeLa/Scr cells (29.5±3.45) (Fig. 4B, P<0.05). Moreover, we detected
the expression of stem cell-related transcription factors, OCT4,
NANOG, KLF4 and BMI1, which are important for maintaining the
self-renewal of embryonic stem cells. Real-time PCR analysis showed
that SiHa/Hiwi cells expressed higher levels of OCT4, NANOG and
BMI1 than the SiHa/NC cells while HeLa/shHiwi cells dispayed lower
levels when compared with the HeLa/Scr cells (Fig. 4C, P<0.05). These data indicate
that Hiwi displays a stemness signature in cervical cancer
cells.
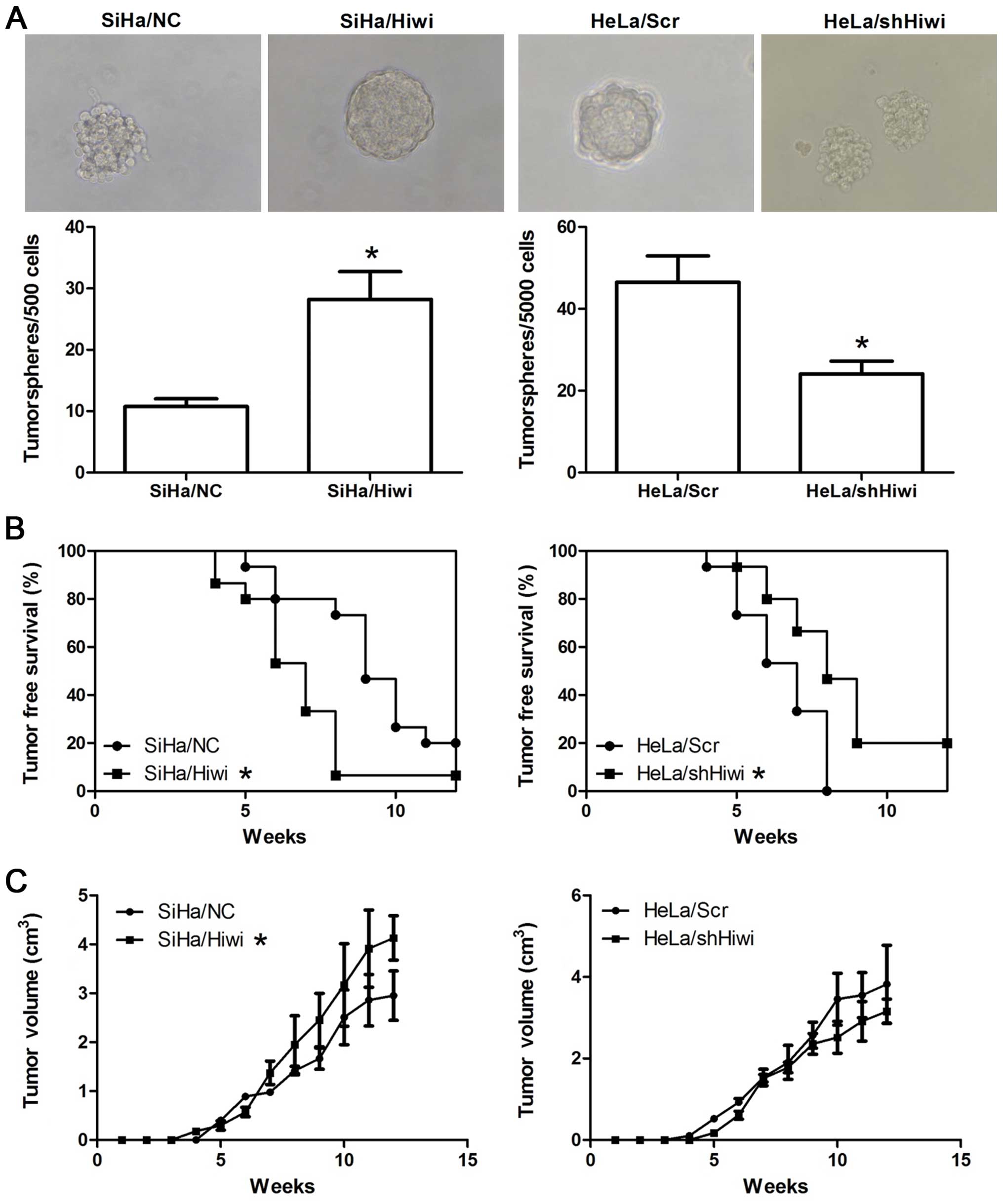
Hiwi promotes tumorigenicity in vitro and
in vivo
Tumor stem cells resemble stem cells in their
ability to grow as ‘spheres’ when cultured in conditions where they
cannot attach to a solid substratum (19,20).
To verify the role of Hiwi in tumorigenicity in vitro, a
tumorsphere formation assay was performed. Notably, ectopic Hiwi
promoted tumorsphere formation in SiHa cells, while silencing of
Hiwi inhibited this ability in HeLa cells (Fig. 5A, P<0.05). In SiHa cells, ~5.6%
of SiHa/Hiwi cells and 2% of SiHa/NC cells formed tumorspheres,
while in the HeLa cells, ~0.9% of HeLa/Scr cells formed
tumorspheres after a 2-week culture, but only ~0.5% of HeLa/shHiwi
cells did, even when the latter were cultured up to 3 weeks.
To further confirm the potential of Hiwi in
tumorigenesis in vivo, a tumor formation assay was conducted
with BALB/c nude mice. Cells began to form palpable tumors in the
majority of injected mice after 4–5 weeks. Importantly, ectopic
Hiwi promoted the frequency of tumor formation and inhibited the
latency period of tumors derived from the SiHa cells, and silencing
of Hiwi had the opposite effect (Fig.
5B, P<0.05). After tumor initiation, Hiwi significantly
promoted the tumor growth (Fig. 5C,
P<0.05). However, silencing of Hiwi did not significantly affect
the xenograft growth rates (Fig.
5C, P>0.05).
Discussion
Hiwi, encoding a highly basic 861-amino acid
protein, is a member of the Piwi family that represents the only
known class of evolutionarily conserved genes that are required for
stem cell function in diverse organisms (21,22).
Recently, increasing evidence suggests that dysregulation of Hiwi
is associated with the pathogenesis of several human cancers
(14,23–25).
In this study, we determined the expression of Hiwi
in normal and pathologic cervical tissues, as well as in cervical
cancer cell lines. IHC showed a higher level of Hiwi expression in
high-grade squamous intraepithelial lesions (HSILs) and cervical
cancers (CCs) than that in normal cervix (NC) (Fig. 1), and a positive relationship with
pathological stage (Table I), which
indicates that Hiwi expression may be associated with cervical
carcinogenesis. This finding is of great importance as Hiwi
expression was found in the basal cells of normal cervical tissues
where epithelial reserve cells are located. Reserve cells are
likely candidates for cervical stem cells (26). These results are similar to previous
studies that found that Hiwi has functions in the maintenance of
stem cells and cancer stem cells (8,13,27),
suggesting that Hiwi participates in the physiological function of
the cervix and may be involved in cervical carcinogenesis as a
cancer stem marker.
On the basis of this hypothesis, we analyzed Hiwi
expression in the response of cervical cancer to radiotherapy alone
or radiotherapy plus concomitant chemotherapy with cisplatin, since
chemical resistance is one of the characteristics of CSCs. The
array dataset showed that Hiwi was upregulated after
chemoradiotherapy, suggesting that Hiwi is associated with
cisplatin resistance. Thus, overexpression and silencing of Hiwi
were induced in SiHa and HeLa cells, respectively, for further
investigation. The MTT assays demonstrated that cisplatin caused
dose-dependent and time-dependent inhibition of viability in both
of the SiHa and HeLa cells suggesting that Hiwi induced resistance
to chemotherapy (Fig. 3).
In addition, we observed that ectopic Hiwi inhibited
the growth and proliferation of SiHa cells while the silencing of
Hiwi promoted these abilities in HeLa cells (Fig. 4). This result suggests that Hiwi
induces a stemness characteristic in cervical cancer as stem cells
exhibit a quiescent state. Furthermore, the mechanism of cisplatin
resistance has been previously proposed. Cisplatin is a nonspecific
drug which acts on the cell cycle in several types of cancers,
inhibiting DNA synthesis by binding to and causing crosslinking of
DNA, which ultimately triggers apoptosis (28). Hiwi-induced cisplatin resistance may
be due to cell cycle arrest.
Apart from chemical resistance, cancer stem cells
are characterized by two other common properties: self-renewal and
tumorigenesis (29,30). Thus, we detected several
self-renewal-related transcription factors and found that Hiwi
facilitates expression of these markers, indicating a stemness
signature. Moreover, the tumorsphere formation assay showed larger
tumorspheres and a higher percentage of formation in the SiHa/Hiwi
and HeLa/Scr cells than these parameters in the SiHa/NC or
HeLa/shHiwi cells, while the tumor xenograft experiment showed a
reduced tumor-free period and a lower tumor-free rate in mice
injected with the SiHa/Hiwi and HeLa/Scr cells than in mice
injected with the SiHa/NC or HeLa/shHiwi cells (Fig. 5), demonstrating that Hiwi promotes
cells to possess a higher tumor-initiating capacity in vitro
and in vivo.
In summary, the present study elucidated the role of
Hiwi expression in cervical cancers. Cells expressing Hiwi
exhibited resistance to chemotherapy drugs and increased
tumorigenesis in vitro and in vivo. Additionally,
these cells possessed the ability for self-renewal and expressed
high levels of stem cell-related transcription factors. Based on
this study, Hiwi may be considered as a marker for cervical CSCs,
and a target with which to explore novel strategies for the
diagnosis, prognosis and therapy of cervical cancer.
Acknowledgements
This project was supported by the National Natural
Science Foundation of China (no. 81270435).
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar
|
|
2
|
Reid R: Genital warts and cervical cancer.
II Is human papillomavirus infection the trigger to cervical
carcinogenesis? Gynecol Oncol. 15:239–252. 1983.PubMed/NCBI
|
|
3
|
Burd EM: Human papillomavirus and cervical
cancer. Clin Microbiol Rev. 16:1–17. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Clarke MF, Dick JE, Dirks PB, et al:
Cancer stem cells - perspectives on current status and future
directions: AACR Workshop on cancer stem cells. Cancer Res.
66:9339–9344. 2006. View Article : Google Scholar
|
|
5
|
Visvader JE and Lindeman GJ: Cancer stem
cells in solid tumours: accumulating evidence and unresolved
questions. Nat Rev Cancer. 8:755–768. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lingel A and Sattler M: Novel modes of
protein - RNA recognition in the RNAi pathway. Curr Opin Struct
Biol. 15:107–115. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cox DN, Chao A, Baker J, Chang L, Qiao D
and Lin H: A novel class of evolutionarily conserved genes defined
by piwi are essential for stem cell self-renewal. Genes Dev.
12:3715–3727. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Grochola LF, Greither T, Taubert H, et al:
The stem cell-associated Hiwi gene in human adenocarcinoma of the
pancreas: expression and risk of tumour-related death. Br J Cancer.
99:1083–1088. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Qiao D, Zeeman AM, Deng W, Looijenga LH
and Lin H: Molecular characterization of hiwi, a human
member of the piwi gene family whose overexpression is
correlated to seminomas. Oncogene. 21:3988–3999. 2002.
|
|
10
|
Liu X, Sun Y, Guo J, et al: Expression of
hiwi gene in human gastric cancer was associated with
proliferation of cancer cells. Int J Cancer. 118:1922–1929.
2006.
|
|
11
|
Taubert H, Greither T, Kaushal D, et al:
Expression of the stem cell self-renewal gene Hiwi and risk
of tumour-related death in patients with soft-tissue sarcoma.
Oncogene. 26:1098–1100. 2007.PubMed/NCBI
|
|
12
|
Taubert H, Würl P, Greither T, et al: Stem
cell-associated genes are extremely poor prognostic factors for
soft-tissue sarcoma patients. Oncogene. 26:7170–7174. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sharma AK, Nelson MC, Brandt JE, et al:
Human CD34(+) stem cells express the hiwi gene, a human
homologue of the Drosophila gene piwi. Blood.
97:426–434. 2001.PubMed/NCBI
|
|
14
|
Siddiqi S, Terry M and Matushansky I: Hiwi
mediated tumorigenesis is associated with DNA hypermethylation.
PLoS One. 7:e337112012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gao Q, Liu W, Cai J, et al: EphB2 promotes
cervical cancer progression by inducing epithelial-mesenchymal
transition. Hum Pathol. 45:372–381. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Iwakawa M, Ohno T, Imadome K, et al: The
radiation-induced cell-death signaling pathway is activated by
concurrent use of cisplatin in sequential biopsy specimens from
patients with cervical cancer. Cancer Biol Ther. 6:905–911. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ji J and Zheng PS: Expression of Sox2 in
human cervical carcinogenesis. Hum Pathol. 41:1438–1447. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang Y, Li B, Ji ZZ and Zheng PS: Notch1
regulates the growth of human colon cancers. Cancer. 15:5207–5218.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Singh SK, Clarke ID, Terasaki M, et al:
Identification of a cancer stem cell in human brain tumors. Cancer
Res. 63:5821–5828. 2003.PubMed/NCBI
|
|
20
|
Ponti D, Costa A, Zaffaroni N, et al:
Isolation and in vitro propagation of tumorigenic breast cancer
cells with stem/progenitor cell properties. Cancer Res.
65:5506–5511. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Benfey PN: Stem cells: A tale of two
kingdoms. Curr Biol. 9:R171–R172. 1999.PubMed/NCBI
|
|
22
|
Cox DN, Chao A and Lin H: piwi
encodes a nucleoplasmic factor whose activity modulates the number
and division rate of germline stem cells. Development. 127:503–514.
2000.
|
|
23
|
Sun G, Wang Y, Sun L, et al: Clinical
significance of Hiwi gene expression in gliomas. Brain Res.
1373:183–188. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu C, Qu L, Dong B, et al: Combined
phenotype of 4 markers improves prognostic value of patients with
colon cancer. Am J Med Sci. 343:295–302. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu WK, Jiang XY and Zhang ZX: Expression
of PSCA, PIWIL1 and TBX2 and its correlation with HPV16 infection
in formalin-fixed, paraffin-embedded cervical squamous cell
carcinoma specimens. Arch Virol. 155:657–663. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Peters WM: Nature of ‘basal’ and ‘reserve’
cells in oviductal and cervical epithelium in man. J Clin Pathol.
39:306–312. 1986.
|
|
27
|
Liang D, Yang Y and Liu Y: The role Hiwi
gene in the maintenance of lung cancer stem cell populations.
Neoplasma. Dec 4–2013.(Epub ahead of print). View Article : Google Scholar
|
|
28
|
Stordal B, Pavlakis N and Davey R: A
systematic review of platinum and taxane resistance from bench to
clinic: an inverse relationship. Cancer Treat Rev. 33:688–703.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lin H: The tao of stem cells in the
germline. Annu Rev Genet. 31:455–491. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Podberezin M, Wen J and Chang CC: Cancer
stem cells: a review of potential clinical applications. Arch
Pathol Lab Med. 137:1111–1116. 2013. View Article : Google Scholar : PubMed/NCBI
|