Introduction
Ovarian cancer is the sixth most common cancer in
women (1). A high percentage of
mortality results from late diagnosis and low chemotherapy
effectiveness. In most cases, ovarian cancer is diagnosed when the
disease has progressed to stage III or IV, according to the FIGO
classification (2). These patients
have a poor prognosis with the current therapies. The first-line
treatment includes a combined chemotherapeutic regimen of platinum
and taxane (3). The second-line
chemotherapy generally includes taxane derivatives as well as
cisplatin (Cis), topotecan (Top) and doxorubicin (Dox) (4,5).
The primary reason for low chemotherapy efficacy is
resistance of cancer cells to treatment. Several different
drug-resistance mechanisms have been observed in cancer cells.
These mechanisms include: decreased drug accumulation in the cells,
change in drug cellular localization, faster inactivation of the
drug, faster repair of damage by the drug DNA and cellular
membranes, changes in amino acid sequences of target proteins which
make them insensitive or less sensitive to the drugs actions as
well as changes in regulation of apoptosis. However, the most
important and prevalent mechanism of drug resistance is multiple
drug resistance (MDR). MDR means the ability of cancer cells to
actively remove the cytostatics from the cell via transport
proteins. Drug transporters from the ABC family are responsible for
these phenomena (6). Among them,
P-glycoprotein (P-gp) and breast cancer resistance protein (BCRP)
play the main role in MDR.
Sometimes a drug resistance phenotype is observed in
spite of a lack in drug resistance gene expression. This suggests
that other yet unknown genes may also play a role in cancer drug
resistance. Genome wide expression analysis by oligonucleotide
microarray may provide a new insight into novel candidate genes
that are involved in drug resistance.
Current understanding of drug resistance development
in response to cytostatic drugs is based largely on research on
drug-sensitive and -resistant pairs of cell lines. By analyzing
expression of genes using microarray techniques in six
drug-resistant ovarian cancer cell lines, we observed changes in
the expression of many genes related to extracellular matrix
(ECM).
ECM consists of ground substance and fibers. These
components are a dynamic and interactive system that inform cells
about mechanical and biochemical changes in their extracellular
environment. The ground substance occupies the space between the
cells and fibers and consists of proteoglycans, such as decorin,
lumican and keratocan; multi adhesive glycoproteins including
fibronectin and laminin; and glycosaminoglycans, such as hyaluronan
or keratan sulfate. Among fibers, collagen and elastin fibers can
be distinguished. ECM molecules regulate gene expression, cell
proliferation, differentiation and migration as well as cancer
metastasis (7). In physiological
conditions, expression of ECM proteins is restricted to connective
tissue. In pathological conditions, ECM expression has been
reported in many cancers in vivo (8) and in drug-resistant cancer cell lines
(9).
It has been reported that ECM can affect drug
resistance in two different ways. First, by preventing the drug
penetration in the cancer tissue (10–12).
ECM components such as collagen, elastin and polysaccharides
(hyaluronan and proteoglycan), may limit drug diffusion (13). Some drugs, such as Cis, are readily
distributed within the tumor whereas Dox, methotrexate, vinblastine
and paclitaxel bind to cellular macromolecules (14). Second, by interaction of cancer
cells with components of ECM and with growth factors. These
interactions can affect the apoptosis sensitivity and drug
resistance of cancer cells (15,16).
This type of drug resistance is designated as cell
adhesion-mediated drug resistance (CAM-DR) (17). An example of this is resistance of
small cell lung cancer (SCLC) cells to Dox and melphalan, mediated
by interaction of β1-integrins with ECM (8). Alteration in expression of ECM
components, matrix metalloproteinases (MMPs) and other enzymes can
lead to ECM remodelling and increased cancer metastasis (18,19).
The present study shows alterations in the gene
expression levels of ECM and related proteins in the methotrexate
(W1MR), cisplatin (W1CR), doxorubicin (W1DR), vincristine (W1VR),
topotecan (W1TR) and paclitaxel (W1PR)-resistant variant of W1
primary ovarian cancer cell line.
Materials and methods
Reagents
Methotrexate, Cis, Dox, vincristine, Top and
paclitaxel were obtained from Sigma (St. Louis, MO, USA). TRI
reagent, RPMI-1640 medium, fetal bovine serum, penicillin,
streptomycin, amphotericin B (25 μg/ml) and L-glutamine were also
purchased from Sigma. A cell proliferation kit I (MTT) was
purchased from Roche Diagnostics GmbH (Mannheim, Germany).
Affymetrix GeneChip® 3′ IVT Express kit, and Affymetrix
GeneChip Human Genome U219 microarrays were both from Affymetrix
(Santa Clara, CA, USA).
Cell lines and cell culture
Human ovarian cancer cell line W1 was established
from ovarian cancer tissue obtained from an untreated patient.
Sublines resistant to Mtx- W1 methotrexate (W1MR), Cis- W1
cisplatin (W1CR), Dox- W1 doxorubicin (W1DR), Vin- W1 vincristine
(W1VR), Top- W1 topotecan (W1TR) and Pac- W1 paclitaxel (W1PR) were
obtained by exposure of the W1 line to stepwise increasing drug
concentrations. Final concentration of each drug was 2-fold greater
than the concentration in the plasma 2 h after intravenous
administration (20). The cells
were 138-, 8-, 10-, 24-, 20- and 641-fold resistant to their
selective drugs, respectively, as determined by cell proliferation
kit I (MTT). All cell lines were maintained as monolayer in
complete medium [RPMI-1640 medium supplemented with 10% (v/v) fetal
bovine serum, 2 pMl-glutamine, penicillin (100 U/ml), streptomycin
(100 U/ml) and amphotericin B (25 μg/ml)] at 37°C in a 5%
CO2 atmosphere.
RNA isolation and preparation of
microarray and RQ-PCR reactions
RNA isolation from W1 and all resistant sublines was
performed using TRI reagent, according to company protocol. RNA was
quantified using spectrophotometry methods by measuring absorbance
values at 260 and 280 nm wave length as well as ratio of 260/280 nm
to estimate protein contamination level. Each sample ratio was in
(1.8–2.0) interval. RNA degradation level was checked by
electrophoresis method using 1% denaturing agarose gel and was also
measured by estimating RIN factor on Bioanalyzer 2100 (Agilent
Technologies, Inc., Santa Clara, CA, USA). Estimated RIN values
were between 8.5–10 with an average of 9.2. Additionally, each
sample was diluted to final work concentration of 100 ng/μl. All
samples were prepared in triplicate. cDNA for microarray analysis
was synthesized in two steps (separate synthesis for first and
second strand) using Affymetrix GeneChip® 3 ′IVT Express
kit and 100 ng/μl of RNA according to Affymetrix GeneAtlas 3′ IVT
Express kit protocol. Next, in vitro transcription
(resulting in population of cRNA), biotin labelling and cRNA
fragmentation was also performed using the same protocol.
Microarray hybridization and
scanning
Following this procedure, samples were loaded and
hybridized with Affymetrix GeneChip Human Genome U219 microarrays
along with control cRNA and oligo B2. The hybridization process was
conducted on AccuBlock™ Digital Dry Bath (Labnet International,
Inc. New York, NY, USA) hybridization oven at 45°C for 16 h. We
then washed and stained microarrays according to the technical
protocol using Affymetrix GeneAtlas™ Fuidics Station and scanned
chips with Affymetrix GeneAtlas™ Imaging Station (both from
Affymetrix). Scanned microarrays were saved as *.CEL files on hard
disk.
Microarray analysis and gene
screening
Quality control (QC) studies were performed using
Affymetrix GeneAtlas™ (Affymetrix) software according to the
manufacturer’s standards. Secondary quality control studies were
evaluated by Partek® Express™ Software (Partek Inc., St.
Louis, MO, USA). Gene screening analysis with QC results as well as
statistical algorithms (non-parametric Mann-Whitney U test with
α=0.05) were conducted in the same software. As a result, a table
of the most important fold-changes was developed and imported to
Pathway Studio® Explore (Ariadne Genomics, Rockville,
MD, USA) where pathway studies were performed. Genes with most
changes in fold degree between resistant and parental W1 cell lines
are listed.
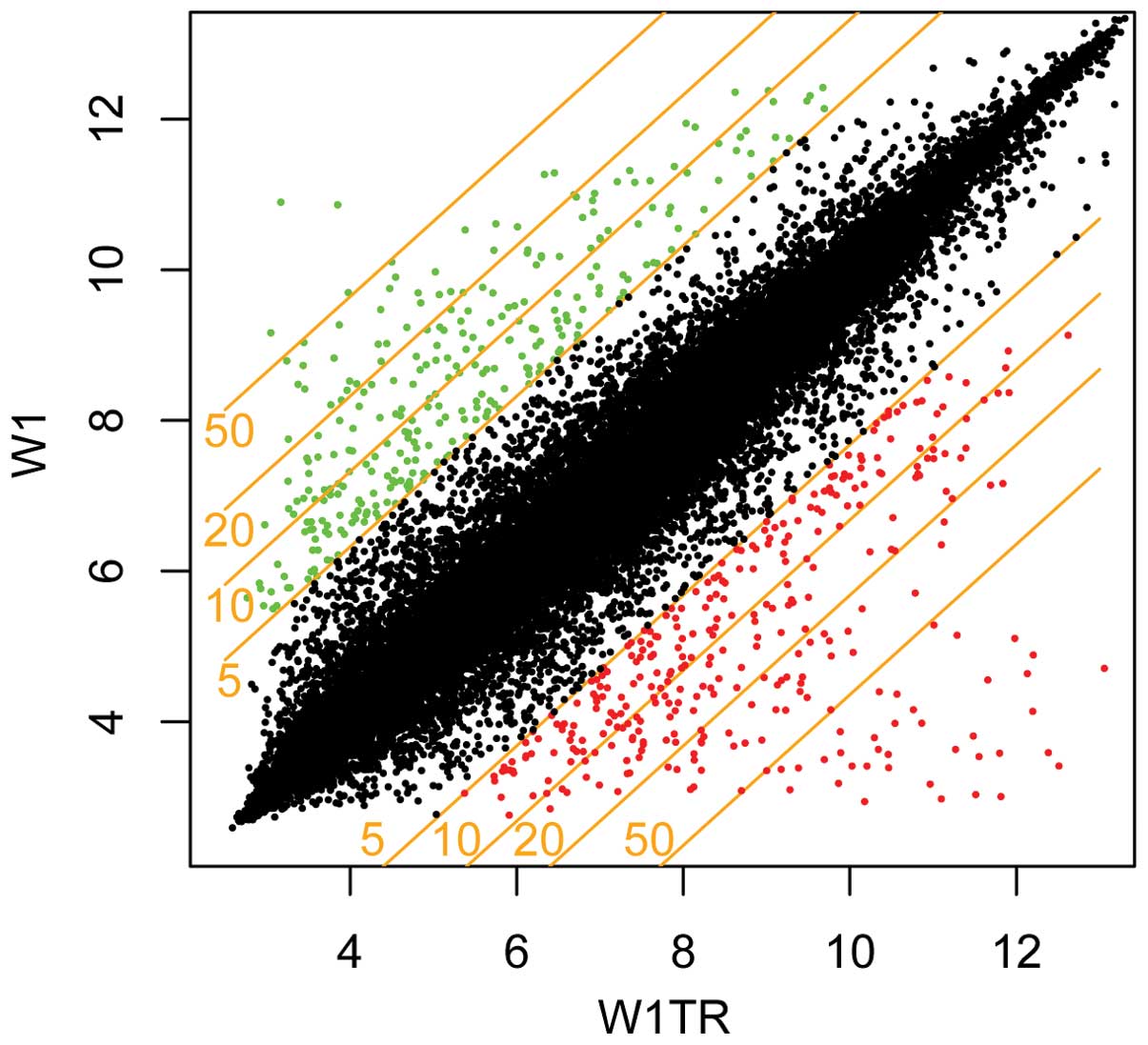
To visualize effects of filtering, we applied gene
list to scatter plot with fold-change level of five as a threshold
(genes upregulated, >5 fold-change and downregulated, <−5
fold-change) (Fig. 1). The same
threshold was applied to the gene table, and in the final step,
genes related to ECM were selected and their participation in drug
resistant phenotype was evaluated. Scatter plot and gene lists were
made using R language (http://www.r-project.org, version 2.14) with
Bioconductor (http://www.bioconductor.org) package installed.
Results
Gene chip quality assessment
In the present study, we used standard factors such
as background to noise signal, internal hybridization controls,
internal poly(A) control RNA as well as GAPDH to β-actin 3′/5′
signal ratios to preliminarily determine the quality of analyzed
samples.
Gene expression evaluation and gene
expression lists
We analyzed changes in transcription level of
ECM-related genes. Analysis of the gene expression in six
drug-resistant ovarian cancer cell lines provided new data
regarding the response of cancer cells to chemotherapeutics
treatment. Tables I and II summarize the changes in the expression
of ECM, matrix metallopeptidases and related gene expression levels
in drug-resistant sublines with respect to W1 drug sensitive cell
line. The genes with statistically significant changes below
p<0.001 and within fold-change intervals (∞, 5] and [0.2, 0)
which correspond to translation >5 and <−5, respectively,
were considered as genes involved in the drug resistance phenomenon
while genes inside interval (0.2, 5) were not. Negative values of
gene expression were rescaled to positive real numbers so that if
-x is negative expression level than equation −1/−x will cast it to
positive real numbers set.
 | Table IThe genes showing the fold-change in
extracellular matrix-related proteins in the drug-resistant
sublines with respect to the parental W1 cell line. |
Table I
The genes showing the fold-change in
extracellular matrix-related proteins in the drug-resistant
sublines with respect to the parental W1 cell line.
| |
Up-/downregulation |
|---|
| |
|
|---|
| Gene symbol | RefSeq transcript
ID | C vs. W1CR | C vs. W1DR | C vs. W1MR | C vs. W1PR | C vs. W1TR | C vs. W1VR |
|---|
| ITGA1 | NM_181501 | NS | NS | −19.67 | −70.9 | −69.34 | −24.78 |
| ITGA6 | NM_000210 | −6.34 | NS | −19.70 | NS | NS | NS |
| ITGB1BP3 | NM_170678 | NS | NS | NS | NS | 73.56 | NS |
| COL1A1 | NM_000088 | NS | NS | NS | NS | 11.70 | NS |
| COL1A2 | NM_000089 | −28.51 | NS | −29.41 | −64.85 | NS | −5.99 |
| COL3A1 | NM_000090 | NS | NS | NS | 85.11 | 548.74 | NS |
| COL4A6 | NM_001847 | NS | NS | NS | −6.84 | −5.26 | NS |
| COL5A2 | NM_000393 | NS | NS | NS | 8.95 | 25.09 | NS |
| COL15A1 | NM_001855 | NS | NS | NS | NS | NS | 66.96 |
| LOX | NM_002317 | NS | NS | NS | NS | 5.72 | NS |
| TGFBI | NM_000358 | NS | NS | NS | NS | 33.78 | NS |
| LAMA1 | NM_005559 | NS | NS | NS | −5.37 | NS | NS |
| LAMA2 | NM_000426 | −27.64 | −38.73 | −6.57 | −33.34 | −14.22 | −13.23 |
| LAMB1 | NM_002291 | NS | NS | −7.28 | NS | NS | −16.67 |
| FN1 | NM_002026 | NS | NS | NS | NS | NS | −6.14 |
| FBN1 | NM_000138 | NS | NS | NS | 8.87 | NS | NS |
| FBN2 | NM_001999 | NS | NS | NS | 12.50 | NS | −6.13 |
| EFEMP1 | NM_001039348 | NS | NS | NS | 5.10 | NS | NS |
| EFEMP2 | NM_016938 | NS | NS | NS | 5.24 | NS | NS |
| GPC3 | NM_004484 | NS | NS | NS | −63.75 | NS | NS |
| GPC6 | NM_005708 | NS | −14.37 | NS | −14.23 | −8.37 | NS |
| DCN | NM_001920 | NS | NS | NS | 35.32 | 151.11 | NS |
| KERA | NM_007035 | NS | NS | NS | NS | 11.79 | NS |
| LUM | NM_002345 | NS | 6.27 | NS | NS | 137.51 | 11.09 |
| BCAN | NM_021948 | NS | NS | NS | NS | 6.66 | NS |
| SPOCK3 | NM_001040159 | NS | 6.19 | NS | NS | NS | NS |
| NID2 | NM_007361 | NS | NS | NS | 7.20 | NS | NS |
| KRT18 | NM_000224 | −6.08 | −5.35 | −9.12 | NS | NS | NS |
| KRT23 | NM_015515 | 9.71 | NS | 10.61 | −37.96 | −12.63 | NS |
| COCH | NM_001135058 | NS | NS | NS | 6.77 | NS | NS |
| VIT | NM_053276 | NS | −7.31 | NS | −85.07 | −52.64 | NS |
| CLDN11 | NM_005602 | −6.73 | NS | NS | NS | NS | NS |
| MATN2 | NM_002380 | NS | NS | NS | 60.78 | 29.10 | NS |
| MATN3 | NM_002381 | NS | NS | NS | NS | 7.44 | −11.24 |
| CD248 | NM_020404 | NS | NS | NS | 12.04 | 11.89 | NS |
| POSTN | NM_001135934 | NS | 70.42 | 80.19 | NS | NS | NS |
| CTHRC1 | NM_138455 | NS | 11.11 | NS | NS | NS | NS |
| HMCN1 | NM_031935 | NS | NS | NS | −61.81 | −13.39 | −29.35 |
| EGFL6 | NM_015507 | NS | NS | NS | NS | 20.90 | NS |
| TGFBR2 | NM_001024847 | NS | NS | NS | NS | 5.12 | NS |
| WNT5A | NM_003392 | NS | NS | NS | 18.47 | NS | NS |
| Table IIThe genes showing the fold-change in
matrix metallopeptidases and related genes in the drug-resistant
sublines with respect to the parental W1 cell line. |
Table II
The genes showing the fold-change in
matrix metallopeptidases and related genes in the drug-resistant
sublines with respect to the parental W1 cell line.
| |
Up-/downregulation |
|---|
| |
|
|---|
| Gene symbol | RefSeq transcript
ID | C vs. W1CR | C vs. W1DR | C vs. W1MR | C vs. W1PR | C vs. W1TR | C vs. W1VR |
|---|
| MMP2 | NM_001127891 | NS | NS | NS | 6.67 | NS | NS |
| MMP11 | NM_005940 | NS | NS | NS | 6.32 | NS | NS |
| MMP14 | NM_004995 | NS | NS | NS | −7.25 | NS | NS |
| ADAMTS1 | NM_133638 | −7.72 | NS | NS | NS | 7.89 | −24.85 |
| ADAMTS4 | NM_005099 | NS | NS | NS | NS | NS | NS |
| ADAMTS5 | NM_007038 | NS | NS | NS | NS | 5.01 | NS |
| ADAMTS19 | NM_133638 | NS | −40.24 | NS | −97.87 | −38.77 | −79.69 |
| ADAMTSL1 | NM_001040272 | 5.13 | NS | NS | NS | NS | NS |
| TIMP1 | NM_003254 | NS | NS | NS | 5.04 | 5.36 | NS |
| TIMP3 | NM_000362 | NS | NS | NS | −5.37 | NS | NS |
Expression of 41 genes encoding ECM proteins,
integrin receptors and related genes were altered in drug-resistant
cell lines (Tables I and II, Figs.
2 and 3). Twenty-four genes
were upregulated in at least one drug-resistant cell line. Fourteen
genes were downregulated in at least one drug-resistant cell line
and three genes were downregulated or upregulated depending on the
cell line. The most variable cell lines were the Pac- and
Top-resistant cell lines. In these cell lines we observed changes
in expression of 28 and 26 genes, respectively. The most stable
cell lines were the Cis- and Mtx-resistant cell lines. In these
cell lines, we observed changes in expression of only eight
genes.
From these 41 genes, expression of 10 was
upregulated very significantly, >20-fold. These genes included:
ITGB1BP3, COL3A1, COL5A2, COL15A1, TGFBI, DCN, LUM, MATN2, POSTN
and EGFL6. Seven genes were very significantly downregulated:
ITGA1, COL1A2, LAMA2, GPC3, KRT23, VIT and HMCN1. Changes in
expression levels of all genes are summarized in Tables I and II and Figs.
2 and 3.
High similarity between the Pac- and Top-resistant
cell lines has been observed. In both cell lines, we observed
upregulation and downregulation of six identical genes. Of the six
upregulated genes, three were upregulated very significantly in
both cell lines: COL3A1, DCN and MATN2. Among the six downregulated
genes, three were downregulated very significantly in both cell
lines: ITGA1, COL1A2 and VIT (Table
I and Fig. 2).
Among MMPs, ADAMT and TIMP (Table II and Fig. 3) expression of nine genes was
altered in drug-resistant cell lines. Five genes were upregulated,
three genes were downregulated and one gene was upregulated or
downregulated depending on the cell line. We did not observe any
notable changes in upregulated genes. Expression of all genes
increased between 5–10-fold. One gene, ADAMTS19, was significantly
downregulated (>20-fold) in four of six investigated cell
lines.
Discussion
The present study demonstrated a correlation between
resistance to cytostatic drugs and expression of genes encoding ECM
and related proteins. Genes with a fold-change situated in (0.2, 5)
interval were considered as not significantly altered in expression
and are not taken into consideration.
Expression of drug transporters from the ABC family
is the main reason for the MDR phenotype of cancer in vivo
as well as cancer cell lines in vitro. The most important
are P-gp and BCRP (21). Expression
of these two proteins has also been reported by us in investigated
cell lines (22,23). Using western blotting, we confirmed
microarray data at the protein level of many other proteins and
among them for COL3A1 (data not shown). In all cases, we observed a
correlation between microarray results and expression at the
protein level.
Previous data indicated that ECM proteins and
related molecules may also be involved in cancer drug resistance
(17). ECM may increase the drug
resistance of a solid tumor by blocking the penetration of
therapeutic agents (10–13) or by increasing resistance to
apoptosis (15,16). ECM expression has been reported in
tumors in vivo as well as in vitro in drug resistant
cell lines (9,24). Expression of ECM molecules together
with drug transporters from the ABC family in vitro indicate
that ECM components can play a significant role in drug
resistance.
Integrins are transmembrane receptors that mediate
the attachment between a cell and the ECM. They are involved in
cell signalling and the regulation of cell cycle, shape and
motility. We observed decreased expression of two integrin genes
ITGA1 and ITGA6. This is in contrast to other results, where an
increase rather than a decrease in the expression of integrin genes
was observed (9,25,26).
ITGA1 form with the β1 subunit cell-surface receptor for collagen
and laminin. Notably, we observed strong downregulation of ITGA1 in
cell lines with the highest expression of COL genes. This is in
contrast to the study of Varma et al, who observed
overexpression of ITGA1 along with COL6A1 in A2780/C10 cell line
resistant to oxaliplatin (27). Our
result is difficult to explain in the context of studies showing
anti-apoptotic effects of cell adhesion to ECM via integrins. In
Top-resistant cell line we observed upregulation of the ITGB1BP3
gene. This gene code nicotinamide riboside kinase 2 is also known
as muscle integrin-binding protein (MIBP). We did not find any data
regarding the role of this protein in cancer drug resistance in the
literature. Elucidation of its role in drug resistance requires
further investigation.
Upregulation of four collagen genes in investigated
cell lines have been observed: COL1A1, COL3A1, COL5A2 and COL15A1.
COL3A1 was upregulated in Pac- and Top-resistant cell lines.
Expression in Top-resistant cell line increased >500-fold. This
suggests that in can play a very important role in the resistance
to Top. COL1A1 and COL5A2 were also upregulated in Top-resistant
cell lines and COL15A1 was upregulated in a Vin-resistant cell
line. These results indicate that COL gene upregulation can be a
specific cellular response to Pac, Top and Vin treatment. Similar
results were observed by us in other drug-resistant ovarian cancer
cell lines resistant to Pac and Top (24). Increased expression of COL genes has
also been observed by others in vitro and in vivo. In
drug-resistant variants of MCF-7 breast cancer cell line,
overexpression of six COL genes was observed (9). In Cis-resistant variant of A2780
ovarian cancer cell line, overexpression of COL6A3 and COL11A1 was
detected (28). Collagen genes were
also upregulated in advanced ovarian cancer (29). These data suggest that collagens
indeed play an important role in drug resistance. It appears that
COL overexpression can protect cells against cytostatics by two
ways. First, by interaction of collagen with cellular receptors. It
has been observed that cultivation of Cis-sensitive A2780 ovarian
cancer cells in the presence of collagen VI protein led to
resistance to Cis in vitro. This effect can result from
interaction of collagen with cellular receptors that lead to
apoptosis resistance (27). The
other possibility is that collagens specifically interact with
cytostatic drugs decreasing the amount of drug that can target
cells. It has been observed that in tumor resistance to
penetration, collagen network is lengthened and this can contribute
to drug resistance by preventing the penetration of therapeutic
agents (12). Dense and tortuous
ECM can be an important barrier for drugs (18). Diffusion rates for larger molecules
inversely correlate with level of fibrillar collagen as well as its
organization and orientation (21,30,31).
We observed considerable expression of fibrillar COL3A1 in
Top-resistant cell lines and increased expression in Pac-resistant
cell line. Very high expression of COL3A1 in Top-resistant cell
lines suggests that it can play a specific role in resistance to
this drug. In contrast, expression of COL1A2 and COL4A6 was
downregulated in four and two drug-resistant cell lines,
respectively, with high downregulation of COL1A2 in W1PR, W1MR and
W1CR lines. In the context of other results, it is difficult to
elucidate downregulation of these genes in investigated cell
lines.
A lysine oxidase (LOX) is important for the
crosslinking of collagens and elastin (32). Increase in expression of LOX in W1TR
cell lines may be related to expression of many COL genes.
Expression of this gene has been reported by us in a Pac-resistant
cell line that also expressed many COL genes (24). Increased LOX expression seems to
play a critical role in promoting tumor growth and metastasis in
many types of cancer, including breast (33), colorectal (34) and lung cancer (35,36).
Expression of LOX in W1TR cell line can lead to a more invasive
character of this cell line. However, this needs to be confirmed by
further study.
Transforming growth factor-β-induced protein
(TGFBI), functions in physiological and pathological conditions
including carcinogenesis (37). It
can play a tumor suppressor (38)
or promoter (39) role depending on
the tumor environment. We observed upregulation of TGFBI in
Top-resistant cell line. Upregulation of this gene has also been
observed by us in other Top-resistant cell lines (24). This indicates that TGFBI can play a
role in Top resistance. We did not find any information in the
literature regarding the role of TGFBI in Top resistance. TGFBI
binds to type I, II and IV collagens. This suggest that it may be
important in cell-collagen interactions. It appears that TGFBI
expression in Top-resistant cell lines may be related to its
interaction with COL1A1 also overexpressed in these cell lines.
Laminins belongs to multiadhesive proteins and play
an important role in linking ECM to cell surface. They are trimeric
proteins that contain an α-, a β- and a γ-chain. The α-chain is
responsible for cell surface receptor, heparin and heparin sulfate
binding. β- and γ-chains are responsible for collagen IV binding.
Laminins influence cell differentiation, migration and adhesion
(40) and play a role in invasive
behavior of tumor cells. In investigated cell lines, we observed
decreased expression of three laminin genes with downregulation of
LAMA2 in all drug-resistant cell lines. This suggests that
downregulation of LAMA2 gene is rather general than specific cell
response for cytostatic-induced stress. These results are in
contrast to results of Işeri et al, who observed strong
upregulation of laminin gene expression in MCF-7 drug resistant
sublines (9). It appears that
changes in laminin gene expression after cytostatic treatment are
cell line-dependent.
FN1 gene encoding fibronectin was slightly
downregulated in W1VR cell lines. This is in contrast to other
studies that showed increased expression of this gene in
drug-resistant cell lines (9).
Shibata et al suggested that fibronectin can increase
invasiveness of ovarian cancer cells (41).
Fibrillins are microfibrils of ECM and play a role
in elastic fiber assembly. FBN1 was upregulated in W1PR cell line.
FBN2 was upregulated in W1PR cell line but downregulated in W1VR
cell line. Işeri et al also observed upregulation of FBN1
and FBN2 in Pac and other drug-resistant cell lines (9). Thus, it appears that FBN genes can
play a role in drug resistance; however, this requires further
study.
We observed increased expression of EFEMP1 (also
known as fibulin-3) and EFEMP2 (also known as fibulin-4) genes in
Pac-resistant cell line. They are members of the fibulin family of
ECM glycoproteins. In pancreatic adenocarcinoma, EFEMP1 expression
promoted tumor growth in vivo and inhibited apoptosis
induced by 5-fluorouracil, gemcitabine and irinotecan (42). We previously observed very high
expression of EFEMP1 in a Dox-resistant cell line (24). Thus, it seems that this gene can
indeed play a role in drug resistance. EFEMP2 expression in
relation to drug resistance has yet to be fully described.
Glypican (GPC), decorin (DCN), keratocan (KERA),
lumican (LUM), brevican (BCAN) and testican encoded by SPOCK3 gene
are ECM proteoglycans. It has been reported that some glypicans
play a role in cell proliferation and survival. We observed
downregulation of two GPC genes, GPC3 and GPC6, with strong
downregulation of GPC3 in W1PR cell line. This is consistent with
observations of Varma et al, who observed GPC3
downregulation in oxaliplatin-resistant ovarian carcinoma cell line
A2780/C10 (27). GPC3 expression is
frequently silenced in ovarian cancer cell lines and it seems that
GPC3 plays a tumor suppressor role in ovarian (43) and lung cancer (44). In both cases, GPC3 expression was
silenced by promoter hypermethylation. In contrast, it is
upregulated in hepatocellular carcinoma (HCC) and regulates cell
proliferation (45). Increased
expression of GPC6 has been reported in drug-resistant MCF-7 cell
lines (9). We recently observed
downregulation as well as upregulation of GPC genes in ovarian
cancer cell lines (24). Hence, the
role of glypicans in drug resistance remains unclear.
DCN belongs to the small leucine-rich proteoglycan
(SLRP) family. It is a component of connective tissue, binds to
type I collagen fibrils and plays a role in collagen
fibrillogenesis when helps to orient fibers. It also can interact
with collagen type I, II, III and IV (46). We observed overexpression of DCN
gene in W1TR and W1PR cell lines. High expression in these cell
lines can be related to high expression level of type I and III
collagen. It has been reported that DCN is a TGF-β1 inhibitor and
reverses TGF-β1-mediated drug resistance (47); it also regulates proliferation and
migration of A549 lung cancer cells (47) and increases angiogenesis and tumor
cell invasiveness in bladder cancer (49). On the one hand, DCN plays an
important function in cancer development and metastasis, on the
other, it is a potential target in cancer therapy (46).
KERA and LUM are also members of the SLRP family. We
observed upregulation of KERA in W1TR cell line and LUM in W1TR as
well as in W1VR cell lines. To our knowledge, the relationship
between KERA expression and drug resistance has not been described.
Increased LUM expression has been reported in Cis-resistant head
and neck squamous cell carcinoma cell lines as well as in patients
not responding to treatment with Cis-based chemotherapy (50). It has been reported that LUM
expression correlates with poor prognosis in advanced colorectal
cancer (51). LUM overexpression
appears to be a marker of drug resistance, yet its role in this
process requires further investigation.
Cytokeratins are intermediate filaments found in the
intracytoplasmic cytoskeleton of epithelial tissue. We observed
small downregulation of KRT18 in Cis-, Dox- and Mtx-resistant cell
lines. Expression of KRT23 was increased in Mtx-resistant cell line
but decreased in Pac- and Top-resistant cell lines. To our
knowledge, changes in expression of these genes have not been
reported in drug-resistant cancer cell lines. Thus, the role of KER
gene expression in drug resistance requires further
investigation.
Cochlin (COCH) is expressed by fibrocytes in the
inner ear and localized to extracellular spaces (52). COCH expression was slightly
increased in Pac-resistant cell line. To our knowledge, to date,
its expression in cancer has not been described.
We observed strong downregulation in VIT gene in
W1TR and W1PR cell lines. Vitrin was first isolated from the
vitreous of the bovine (53). Its
role in drug resistance and cancer has not been reported.
Matrilin-2 (MATN2) is a member of the von Willebrand
factor A domain containing protein family. It may be involved in
the formation of filamentous networks in the ECM, however its
specific function has not yet been determined. Its expression has
been reported in HCC (54). We
observed strong upregulation of MATN2 gene in Pac- and
Top-resistant cell lines. This suggests that it may be related to
COL gene upregulation in these cell lines. The role of MATN2 in
drug resistance has not been described by other researchers. Thus,
the exact role of this protein in resistance to cytostatic drugs
requires further investigation.
We observed increased expression of CD248,
endosialin/tumor endothelial marker 1 (TEM-1) in Pac- and
Top-resistant cell lines. Endosialin is a member of C-type lectin
transmembrane receptors which play a role in cell-cell adhesion
processes. Its role in cancer neoangiogenesis and tumor growth has
been reported (55,56). Thus, it is possible that CD248
expression in vivo can lead to more invasive phenotype of
Pac- and Top-resistant cell lines. This, however, requires animal
studies.
Periostin (POSTN) is a secreted protein and is a
ligand for α-V/β-3 and α-V/β-5 integrins. It is expressed in
epithelial ovarian tumors (29) and
plays a role in migration of ovarian epithelial cells (29,63).
We observed very high expression level of POSTN in W1DR and W1MR
cell line. This is the most highly expressed gene in these cell
lines. Xiao et al reported that oxaliplatin or
5-FU-treatment increase POSTN expression in SW480 and HT-29 colon
cancer cell lines and lead to chemoresistance through activation of
the PI3K/Akt/survivin pathway (58). It is also possible that POSTN
induced drug resistance in our investigated cell lines. However,
this requires further and more detailed study.
Collagen triple helix repeat containing 1 (CTHRC1)
may reduce collagen deposition by inhibition of Smad2/3 activation
(59). Its upregulation was
reported in gastric cancer as well as in colon cancer cell lines
and clinical specimens contributed to cell invasion (60,61).
Thus, increased expression of this gene in W1DR cell line can lead
to a more invasive character of this cell line.
Hemicentin-1 is a large extracellular member of the
immunoglobulin superfamily and is associated with age-related
macular degeneration (62). To our
knowledge, its role in drug resistance or even cancer has not been
described to date. In the present study, we observed its
downregulation in W1TR, W1PR and W1VR cell lines. Its role in drug
resistance requires further investigation.
We observed EGFL6 overexpression in W1TR cell line.
This protein is a member of the epidermal growth factor (EGF)
repeat superfamily and is a secreted protein that promotes
endothelial cell migration and angiogenesis via ERK activation
(63). Its increased expression has
been reported in benign meningioma (64), however its role in drug resistance
has yet to be described.
Small increase in TGFBR2 encoding TGF, β receptor II
has been observed in Top-resistant cell line. Overexpression of
this protein has also been reported by others in Pac-, Doc- and
Dox-resistant variants of MCF-7 breast cancer cell line (65). The authors suggested that this
protein stimulated epithelial-mesenchymal transition (EMT) in these
cell lines. Whether our Top-resistant cell line has characteristics
of cells in EMT requires further investigation.
We observed increased expression of WNT5A in W1PR
cell line. WNT5A is a secreted protein implicated in oncogenic and
several developmental processes. Its expression has been reported
in oxaliplatin-resistant ovarian carcinoma cell line A2780/C10 and
in pancreatic cancer where it play an important role in drug
resistance by increasing resistance to apoptosis (27,66).
Thus, its expression in W1PR cell line may be one of the factors
that increases its drug resistance.
Matrix metalloproteinases (MMPs) are zinc proteases
responsible for ECM degradation. They are a main player in cleaving
a number of bioactive molecules and are important in cell
proliferation, differentiation and migration. They also play a role
in pathological conditions, such as arthritis and cancer
metastasis. We observed small changes in MMP expression. MMP2 and
MMP11 were increased in W1PR cell line and MMP14 was decreased in
this cell line. MMP2 gene encodes an enzyme which degrades type IV
collagen. However, we observed downregulation of type IV collagen
in W1PR cell line. In this context, increased expression of MMP2 in
W1PR cell line is difficult to explain. It has been also reported
that MMP2 plays a role in invasion and induces drug resistance of
oral squamous cell carcinoma in vitro (67). It is possible that MMP2 plays a
similar role in the W1PR cell line. A second MMP overexpressed in
W1PR cell line was MMP11. MMP11 is devoid of enzymatic activity
against the matrix components (68). It has been reported that MMP11
suppresses cancer cell apoptosis and necrosis and promotes tumor
development in animal models (68,69).
This enzyme is also overexpressed in primary breast cancers, their
metastasis and correlates with poor clinical outcome (70). Thus, its expression in the W1PR cell
line can play a role in the resistance to apoptosis and may be
related to a more invasive character of this cell line. This,
however, require further, more detailed study. MMP14 was slightly
decreased in the W1PR cell line. It has been described that
overexpression of MMP14 in prostate carcinoma induces resistance to
gemcitabine by mechanisms involving increases in ERK1/2
phosphorylation (71). The role of
this enzyme in resistance to Pac has not been described to
date.
ADMST belongs to a disinterring and
metalloproteinase with thrombospondin motif family of proteinases.
These molecules are involved in various biological events, such as
cell adhesion, cell fusion, cell migration, membrane protein
shedding and proteolysis. It has been shown that overexpression of
ADAMTS-1 promotes metastasis of mammary and lung carcinoma cells
(72). We observed increased
expression of ADAMTS-1 in a Top-resistant cell line and decreased
expression in Cis- and Vin-resistant cell lines. In the study by
Işeri et al (9), expression
of ADAMST-1 was increased in Vin- and Pac-resistant cell lines. In
the context of these results, the role of this enzyme in drug
resistance appears to be cell line-dependent.
ADAMTS-5 is known to cleave aggrecan and brevican.
Its expression has been reported in human glioblastomas where it
cleaves brevican and promotes invasion (73). We observed increased expression of
ADAMTS-5 in a Top-resistant cell line. This cell line also
overexpresses brevican. Thus, increased expression of ADAMS5 in
this cell line may be related to brevican overexpression.
The function of ADAMST-19 remains unclear. The
relationship between this gene expression and cancer has also not
been described. In the present study, we observed its
downregulation in four of six drug-resistant cell lines. Thus, its
role in drug resistance and cancer needs to be further
investigated.
Tissue inhibitors of metalloproteinases (TIMPs) form
inhibitory complexes with MMPs as well as ADAMs and block the ECM
degradation. We observed increased expression of TIMP1 in W1PR and
W1TR cell lines. In addition to its inhibitory function, TIMP1 can
be implicated in the promotion of cell proliferation, inhibition of
apoptosis as well as drug resistance. In breast cancer, high levels
of TIMP1 have also been associated with a reduced response to
chemotherapy (74,75). Thus, its increased expression in
investigated cell lines can also be related to chemotherapy
resistance.
TIMP3 expression was slightly downregulated in W1PR
cell line. This is consistent with results of Işeri et al,
who also observed strong downregulation of TIMP3 in MCF-7 cell
lines resistant to Doc and Dox (9).
Decreased expression of TIMP3 was also reported in ovarian tumors
after adjuvant chemotherapy (76).
Thus, decreased expression of TIMP3 appears to be an unspecific
response of cancer cells to cytostatic drug treatment.
In summary, our results present alteration in
expression of many genes encoding ECM-related proteins in six
ovarian cancer cell lines resistant to different cytostatic drugs.
These results indicate that ECM proteins may also be implicated in
drug resistance. Any general response to cytostatic treatment has
not been observed. Alterations in gene expression appear to be
rather cytostatic-dependent; however, we observed some similarity
in gene expression profile between Pac- and Top-resistant cell
lines. The importance of the investigated gene expression in drug
resistance requires further investigation and should be confirmed
in other ovarian cancer cell lines.
Acknowledgements
This study was funded by grant no. N N401 204139
from the National Science Centre.
References
|
1
|
Garcia M, Jemal A, Ward EM, et al: Global
cancer facts & figures 2007. American Cancer Society; Atlanta,
GA: 2007
|
|
2
|
Kurman RJ: Blaustein’s Pathology of the
Female Genital Tract. 5th edition. Springer; New York, NY: 2002
|
|
3
|
Parmar MK, Ledermann JA, Colombo N, du
Bois A, Delaloye JF, Kristensen GB, et al: Paclitaxel plus
platinum-based chemotherapy versus conventional platinum-based
chemotherapy in women with relapsed ovarian cancer: the
ICON4/AGO-OVAR-2.2 trial. Lancet. 361:2099–2106. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pujade-Lauraine E, Wagner U,
Aavall-Lundqvist E, et al: Pegylated liposomal doxorubicin and
carboplatin compared with paclitaxel and carboplatin for patients
with platinum-sensitive ovarian cancer in late relapse. J Clin
Oncol. 28:3323–3329. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sehouli J, Stengel D, Oskay-Oezcelik G,
Zeimet AG, Sommer H, Klare P, et al: Non platinum topotecan
combinations versus topotecan alone for recurrent ovarian cancer:
results of a phase III study of the North-Eastern German Society of
Gynecological Oncology Ovarian Cancer Study Group. J Clin Oncol.
26:3176–3182. 2008. View Article : Google Scholar
|
|
6
|
Choi CH: ABC transporters as multidrug
resistance mechanisms and the development of chemosensitizers for
their reversal. Cancer Cell Int. 5:302005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Stupack DG and Cheresh DA: Get a ligand,
get a life: integrins, signaling and cell survival. J Cell Sci.
115:3729–3738. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sethi T, Rintoul RC, Moore SM, MacKinnon
AC, Salter D, Choo C, et al: Extracellular matrix proteins protect
small cell lung cancer cells against apoptosis: a mechanism for
small cell lung cancer growth and drug resistance in vivo. Nat Med.
5:662–668. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Işeri OD, Kars MD, Arpaci F and Gündüz U:
Gene expression analysis of drug-resistant MCF-7 cells:
implications for relation to extracellular matrix proteins. Cancer
Chemother Pharmacol. 65:447–455. 2010.PubMed/NCBI
|
|
10
|
Jain RK: The next frontier of molecular
medicine: delivery of therapeutics. Nat Med. 4:655–657. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tannock IF, Lee CM, Tunggal JK, Cowan DS
and Egorin MJ: Limited penetration of anticancer drugs through
tumor tissue: a potential cause of resistance of solid tumors to
chemotherapy. Clin Cancer Res. 8:878–884. 2002.PubMed/NCBI
|
|
12
|
Netti PA, Berk DA, Swartz MA, Grodzinsky
AJ and Jain RK: Role of extracellular matrix assembly in
interstitial transport in solid tumors. Cancer Res. 60:2497–2503.
2000.PubMed/NCBI
|
|
13
|
Jain RK: Transport of molecules in the
tumor interstitium: a review. Cancer Res. 47:3039–3051.
1987.PubMed/NCBI
|
|
14
|
Di Paolo A and Bocci G: Drug distribution
in tumors: mechanisms, role in drug resistance, and methods for
modification. Curr Oncol Rep. 9:109–114. 2007.PubMed/NCBI
|
|
15
|
St Croix B, Flørenes VA, Rak JW, Flanagan
M, Bhattacharya N, Slingerland JM and Kerbel RS: Impact of the
cyclin-dependent kinase inhibitor p27Kip1 on resistance of tumor
cells to anticancer agents. Nat Med. 2:1204–1210. 1996.PubMed/NCBI
|
|
16
|
St Croix B and Kerbel RS: Cell adhesion
and drug resistance in cancer. Curr Opin Oncol. 9:549–556.
1997.
|
|
17
|
Dalton WS: The tumor microenvironment as a
determinant of drug response and resistance. Drug Resist Updat.
2:285–288. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chauhan VP, Stylianopoulos T, Boucher Y
and Jain R: Delivery of molecular and nanoscale medicine to tumors:
transport barriers and strategies. Annu Rev Chem Biomol Eng.
2:281–298. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ramanujan S, Pluen A, McKee TD, Brown EB,
Boucher Y and Jain RK: Diffusion and convection in collagen gels:
implications for transport in the tumor interstitium. Biophys J.
83:1650–1660. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dietel M, Bals U, Schaefer B, Herzig I,
Arps H and Zabel M: In vitro prediction of cytostatic drug
resistance in primary cell cultures of solid malignant tumours. Eur
J Cancer. 29A:416–420. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Leonard GD, Fojo T and Bates SE: The role
of ABC transporters in clinical practice. Oncologist. 8:411–424.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Januchowski R, Zawierucha P, Andrzejewska
M, Ruciński M and Zabel M: Microarray-based detection and
expression analysis of ABC and SLC transporters in drug-resistant
ovarian cancer cell lines. Biomed Pharmacother. 67:240–245. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Januchowski R, Wojtowicz K,
Sujka-Kordowska P, Andrzejewska M and Zabel M: MDR gene expression
analysis of six drug-resistant ovarian cancer cell lines. Biomed
Res Int. 2013:2417632013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Januchowski R, Zawierucha P, Ruciński M,
Nowicki M and Zabel Maciej: Extracellular matrix proteins
expression profiling in chemoresistant variants of the A2780
ovarian cancer cell line. Biomed Res Int. 2014:3658672014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liang Y, Meleady P, Cleary I, McDonnell S,
Connolly L and Clynes M: Selection with melphalan or paclitaxel
(Taxol) yields variants with different patterns of multidrug
resistance, integrin expression and in vitro invasiveness. Eur J
Cancer. 37:1041–1052. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Correia AL and Bissell MJ: The tumor
microenvironment is a dominant force in multidrug resistance. Drug
Resist Updat. 15:39–49. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Varma RR, Hector SM, Clark K, Greco WR,
Hawthorn L and Pendyala L: Gene expression profiling of a clonal
isolate of oxaliplatin-resistant ovarian carcinoma cell line
A2780/C10. Oncol Rep. 14:925–932. 2005.PubMed/NCBI
|
|
28
|
Sherman-Baust CA, Weeraratna AT, Rangel
LB, Pizer ES, Cho KR, Schwartz DR, Shock T and Morin PJ: Remodeling
of the extracellular matrix through overexpression of collagen VI
contributes to cisplatin resistance in ovarian cancer cells. Cancer
Cell. 3:377–386. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ismail RS, Baldwin RL, Fang J, Browning D,
Karlan BY, Gasson JC and Chang DD: Differential gene expression
between normal and tumor-derived ovarian epithelial cells. Cancer
Res. 60:6744–6749. 2000.PubMed/NCBI
|
|
30
|
Brown E, McKee T, diTomaso E, Pluen A,
Seed B, et al: Dynamic imaging of collagen and its modulation in
tumors in vivo using second-harmonic generation. Nat Med.
9:796–800. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Stylianopoulos T, Diop-Frimpong B, Munn LL
and Jain RK: Diffusion anisotropy in collagen gels and tumors: the
effect of fiber network orientation. Biophys J. 99:3119–3128. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Csiszar K: Lysyl oxidases: a novel
multifunctional amine oxidase family. Prog Nucleic Acid Res Mol
Biol. 70:1–32. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
El-Haibi CP, Bell GW, Zhang J, Collmann
AY, Wood D, Scherber CM, Csizmadia E, Mariani O, Zhu C, Campagne A,
Toner M, Bhatia SN, Irimia D, Vincent-Salomon A and Karnoub AE:
Critical role for lysyl oxidase in mesenchymal stem cell-driven
breast cancer malignancy. Proc Natl Acad Sci USA. 109:17460–17465.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Baker AM, Cox TR, Bird D, Lang G, Murray
GI, Sun XF, Southall SM, Wilson JR and Erler JT: The role of lysyl
oxidase in SRC-dependent proliferation and metastasis of colorectal
cancer. J Natl Cancer Inst. 103:407–424. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shi W, Yang B, Li X, Sun S, Wang L and
Jiao S: The effect of lysyl oxidase polymorphism on susceptibility
and prognosis of nonsmall cell lung cancer. Tumour Biol.
33:2379–2383. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wilgus ML, Borczuk AC, Stoopler M,
Ginsburg M, Gorenstein L, Sonett JR and Powell CA: Lysyl oxidase: a
lung adenocarcinoma biomarker of invasion and survival. Cancer.
117:2186–2191. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ween MP, Oehler MK and Ricciardelli C:
Transforming growth factor-beta-induced protein (TGFBI)/(βig-H3): a
matrix protein with dual functions in ovarian cancer. Int J Mol
Sci. 13:10461–10477. 2012.
|
|
38
|
Li B, Wen G, Zhao Y, Tong J and Hei TK:
The role of TGFBI in mesothelioma and breast cancer: association
with tumor suppression. BMC Cancer. 12:2392012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ma C, Rong Y, Radiloff DR, Datto MB,
Centeno B, Bao S, Cheng W, Lin F, Jiang S, Yeatman TJ and Wang XF:
Extracellular matrix protein βig-h3/TGFBI promotes metastasis of
colon cancer by enhancing cell extravasation. Genes Dev.
22:308–321. 2008.
|
|
40
|
Timpl R, Rohde H, Robey PG, Rennard SI,
Foidart JM and Martin GR: Laminin - a glycoprotein from basement
membranes. J Biol Chem. 254:9933–9937. 1979.PubMed/NCBI
|
|
41
|
Shibata K, Kikkawa F, Nawa A, Suganuma N
and Hamaguchi M: Fibronectin secretion from human peritoneal tissue
induces Mr 92,000 type IV collagenase expression
and invasion in ovarian cancer cell lines. Cancer Res.
57:5416–5420. 1997.PubMed/NCBI
|
|
42
|
Seeliger H, Camaj P, Ischenko I, Kleespies
A, De Toni EN, Thieme SE, Blum H, Assmann G, Jauch KW and Bruns CJ:
EFEMP1 expression promotes in vivo tumor growth in human
pancreatic adenocarcinoma. Mol Cancer Res. 7:189–198. 2009.
View Article : Google Scholar
|
|
43
|
Lin H, Huber R, Schlessinger D and Morin
PJ: Frequent silencing of the GPC3 gene in ovarian cancer
cell lines. Cancer Res. 59:807–810. 1999.
|
|
44
|
Kim H, Xu GL, Borczuk AC, Busch S, Filmus
J, Capurro M, Brody JS, Lange J, D’Armiento JM, Rothman PB and
Powell CA: The heparan sulfate proteoglycan GPC3 is a
potential lung tumor suppressor. Am J Respir Cell Mol Biol.
29:694–701. 2003.PubMed/NCBI
|
|
45
|
Miao HL, Pan ZJ, Lei CJ, Wen JY, Li MY,
Liu ZK, Qiu ZD, Lin MZ, Chen NP and Chen M: Knockdown of GPC3
inhibits the proliferation of Huh7 hepatocellular carcinoma cells
through down-regulation of YAP. J Cell Biochem. 114:625–631. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Sofeu Feugaing DD, Götte M and Viola M:
More than matrix: the multifaceted role of decorin in cancer. Eur J
Cell Biol. 92:1–11. 2013.PubMed/NCBI
|
|
47
|
Teicher BA1, Ikebe M, Ara G, Keyes SR and
Herbst RS: Transforming growth factor-β 1 overexpression produces
drug resistance in vivo: reversal by decorin. In Vivo. 11:463–472.
1997.
|
|
48
|
Liang S, Xu JF, Cao WJ, Li HP and Hu CP:
Human decorin regulates proliferation and migration of human lung
cancer A549 cells. Chin Med J. 126:4736–4741. 2013.PubMed/NCBI
|
|
49
|
El Behi M, Krumeich S, Lodillinsky C,
Kamoun A, Tibaldi L, Sugano G, De Reynies A, Chapeaublanc E,
Laplanche A, Lebret T, Allory Y, Radvanyi F, Lantz O, Eiján AM,
Bernard-Pierrot I and Théry C: An essential role for decorin in
bladder cancer invasiveness. EMBO Mol Med. 5:1835–1851.
2013.PubMed/NCBI
|
|
50
|
Yamano Y, Uzawa K, Saito K, Nakashima D,
Kasamatsu A, Koike H, et al: Identification of cisplatin-resistance
related genes in head and neck squamous cell carcinoma. Int J
Cancer. 126:437–449. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Seya T, Tanaka N, Shinji S, Yokoi K,
Koizumi M, Teranishi N, Yamashita K, Tajiri T, Ishiwata T and Naito
Z: Lumican expression in advanced colorectal cancer with nodal
metastasis correlates with poor prognosis. Oncol Rep. 16:1225–1230.
2006.PubMed/NCBI
|
|
52
|
Robertson NG, Resendes BL, Lin JS, Lee C,
Aster JC, Adams JC and Morton CC: Inner ear localization of mRNA
and protein products of COCH, mutated in the sensorineural
deafness and vestibular disorder, DFNA9. Hum Mol Genet.
10:2493–2500. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Mayne R, Ren ZX, Liu J, Cook T, Carson M
and Narayana S: VIT-1: the second member of a new branch of the von
Willebrand factor A domain superfamily. Biochem Soc Trans.
27:832–835. 1999.PubMed/NCBI
|
|
54
|
Szabó E, Korpos E, Batmunkh E, Lotz G,
Holczbauer A, Kovalszky I, Deák F, Kiss I, Schaff Z and Kiss A:
Expression of matrilin-2 in liver cirrhosis and hepatocellular
carcinoma. Pathol Oncol Res. 14:15–22. 2008.
|
|
55
|
Rupp C, Dolznig H, Puri C, Sommergruber W,
Kerjaschki D, Rettig WJ and Garin-Chesa P: Mouse endosialin, a
C-type lectin-like cell surface receptor: expression during
embryonic development and induction in experimental cancer
neoangiogenesis. Cancer Immun. 6:102006.
|
|
56
|
Valdez Y, Maia M and Conway EM: CD248:
reviewing its role in health and disease. Curr Drug Targets.
13:432–439. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Gillan L, Matei D, Fishman DA, Gerbin CS,
Karlan BY and Chang DD: Periostin secreted by epithelial ovarian
carcinoma is a ligand for αVβ3 and
αVβ5 integrins and promotes cell motility.
Cancer Res. 62:5358–5364. 2002.PubMed/NCBI
|
|
58
|
Xiao ZM, Wang XY and Wang AM: Periostin
induces chemoresistance in colon cancer cells through activation of
the PI3K/Akt/survivin pathway. Biotechnol Appl Biochem. Dec
24–2013.(Epub ahead of print). View Article : Google Scholar
|
|
59
|
LeClair R and Lindner V: The role of
collagen triple helix repeat containing 1 in injured arteries,
collagen expression, and transforming growth factor β signaling.
Trends Cardiovasc Med. 17:202–205. 2007.PubMed/NCBI
|
|
60
|
Wang P, Wang YC, Chen XY, Shen ZY, Cao H,
Zhang YJ, Yu J, Zhu JD, Lu YY and Fang JY: CTHRC1 is
upregulated by promoter demethylation and transforming growth
factor-β1 and may be associated with metastasis in human gastric
cancer. Cancer Sci. 103:1327–1333. 2012. View Article : Google Scholar
|
|
61
|
Kim HC, Kim YS, Oh HW, Kim K, Oh SS, Kim
JT, Kim BY, Lee SJ, Choe YK, Kim DH, Kim SH, Chae SW, Kim KD and
Lee HG: Collagen triple helix repeat containing 1 (CTHRC1) acts via
ERK-dependent induction of MMP9 to promote invasion of colorectal
cancer cells. Oncotarget. 5:519–529. 2014.PubMed/NCBI
|
|
62
|
Thompson CL, Klein BE, Klein R, Xu Z,
Capriotti J, Joshi T, Leontiev D, Lee KE, Elston RC and Iyengar SK:
Complement factor H and hemicentin-1 in age-related macular
degeneration and renal phenotypes. Hum Mol Genet. 16:2135–2148.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Chim SM, Qin A, Tickner J, Pavlos N, Davey
T, Wang H, Guo Y, Zheng MH and Xu J: EGFL6 promotes endothelial
cell migration and angiogenesis through the activation of
extracellular signal-regulated kinase. J Biol Chem.
286:22035–22046. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Wang X, Gong Y, Wang D, Xie Q, Zheng M,
Zhou Y, Li Q, Yang Z, Tang H, Li Y, Hu R, Chen X and Mao Y:
Analysis of gene expression profiling in meningioma: deregulated
signaling pathways associated with meningioma and EGFL6
overexpression in benign meningioma tissue and serum. PLoS One.
7:e527072012. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Işeri OD, Kars MD, Arpaci F, Atalay C, Pak
I and Gündüz U: Drug resistant MCF-7 cells exhibit
epithelial-mesenchymal transition gene expression pattern. Biomed
Pharmacother. 65:40–45. 2011.PubMed/NCBI
|
|
66
|
Griesmann H, Ripka S, Pralle M,
Ellenrieder V, Baumgart S, Buchholz M, Pilarsky C, Aust D, Gress TM
and Michl P: WNT5A-NFAT signaling mediates resistance to apoptosis
in pancreatic cancer. Neoplasia. 15:11–22. 2013.PubMed/NCBI
|
|
67
|
Lu L, Xue X, Lan J, Gao Y, Xiong Z, Zhang
H, Jiang W, Song W and Zhi Q: MicroRNA-29a upregulates MMP2 in oral
squamous cell carcinoma to promote cancer invasion and
anti-apoptosis. Biomed Pharmacother. 68:13–19. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Boulay A, Masson R, Chenard MP, et al:
High cancer cell death in syngeneic tumors developed in host mice
deficient for the stromelysin-3 matrix metalloproteinase. Cancer
Res. 61:2189–2193. 2001.PubMed/NCBI
|
|
69
|
Wu E, Mari BP, Wang F, Anderson IC, Sunday
ME and Shipp MA: Stromelysin-3 suppresses tumor cell apoptosis in a
murine model. J Cell Biochem. 82:549–555. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Noël A, Boulay A, Kebers F, et al:
Demonstration in vivo that stromelysin-3 functions through its
proteolytic activity. Oncogene. 19:1605–1612. 2000.PubMed/NCBI
|
|
71
|
Dangi-Garimella S, Krantz SB, Barron MR,
Shields MA, Heiferman MJ, Grippo PJ, Bentrem DJ and Munshi HG:
Three-dimensional collagen I promotes gemcitabine resistance in
pancreatic cancer through MT1-MMP-mediated expression of HMGA2.
Cancer Res. 71:1019–1028. 2011. View Article : Google Scholar
|
|
72
|
Liu YJ, Xu Y and Yu Q: Full-length
ADAMTS-1 and the ADAMTS-1 fragments display pro- and antimetastatic
activity, respectively. Oncogene. 25:2452–2467. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Nakada M, Miyamori H, Kita D, et al: Human
glioblastomas overexpress ADAMTS-5 that degrades brevican. Acta
Neuropathol. 110:239–246. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Schrohl AS, Meijer-van Gelder ME,
Holten-Andersen MN, Christensen IJ, Look MP, Mouridsen HT, et al:
Primary tumor levels of tissue inhibitor of metalloproteinases-1
are predictive of resistance to chemotherapy in patients with
metastatic breast cancer. Clin Cancer Res. 12:7054–7058. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Klintman M, Ørnbjerg Würtz S, Christensen
IJ, Braemer Hertel P, Fernö M, Malmberg M, et al: Association
between tumor tissue TIMP-1 levels and objective response to
first-line chemotherapy in metastatic breast cancer. Breast Cancer
Res Treat. 121:365–371. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
L’Espérance S, Popa I, Bachvarova M,
Plante M, Patten N, Wu L, Têtu B and Bachvarov D: Gene expression
profiling of paired ovarian tumors obtained prior to and following
adjuvant chemotherapy: Molecular signatures of chemoresistant
tumors. Int J Oncol. 29:5–24. 2006.PubMed/NCBI
|