Introduction
Umbilical cord blood has gradually become an
important source for hematopoietic stem cell transplantation (HSCT)
in the human (particularly in pediatric patients) and has been
applied in the radical treatment of hematologic malignancies, due
to its richness in CD34+ cells, higher transplant
survival rate, lower risk of graft-versus-host disease (GVHD) and
convenient collection (1). Adoptive
cell immunotherapy (AIT) of patients with hematologic malignancies
after umbilical cord blood transplant has become a research hotspot
in recent years, and is important to improve the long-term survival
of children with hematologic malignancies after umbilical cord
blood transplant. In recent years, cytokine-induced killer (CIK)
cells as a new type of antitumor immune cells in tumor prevention
and treatment have increasingly attracted the attention of
researchers.
AIT involves the transfer into patients of immune
cells that have been expanded and activated ex vivo to
eliminate cancer cells. This approach has become an important and
effective method of cancer therapy (2). The main limitation preventing the
successful clinical translation of AIT strategies is the obtainment
of sufficient numbers of antitumor immune effector cells and their
persistence in vivo (3). In
recent years, the application of CIK cells has evolved into an
extensive clinical research topic. CIK cells exhibit a higher
proliferation rate and cytotoxicity in contrast with
lymphokine-activated killer (LAK) cells and tumor infiltrating
lymphocytes (TILs) (4).
CIK cells are obtained ex vivo by stimulating
mononuclear cells (MNCs) with interferon-γ (IFN-γ), interleukin-2
(IL-2) and an anti-CD3 monoclonal antibody for a few weeks as
initially described by Schmidt-Wolf et al (5). Bulk CIK cells can be described as a
heterogeneous population consisting of a majority of cells with a
CD3+CD56+ phenotype (NKT cells) and a minor
fraction of cells with a CD3−CD56+ phenotype
(NK cells) and a CD3+CD56− phenotype
(CD3+ T lymphocytes). The major effector cells are the
CD3+CD56+ subset, termed a mixed T-NK
phenotype and with major histocompatibility complex
(MHC)-unrestricted cytolytic activities against tumor cells
(6). CIK cells derived from
peripheral blood (PB) have demonstrated an effective antitumor
potential both in vitro and in vivo against solid
tumors, hematological malignancies and virus-associated tumors
(7–10). In recent years, a large amount of
clinical studies have proven the effectiveness, safety and
feasibility of CIK cells derived from PB (11,12).
However, PB is not always a viable cell source, particularly for
allogeneic transplantation, due to the risk of acute GVHD (13). Moreover, the application of
autologous CIK cells is limited due to the short survival period
in vivo and the lower anti-apoptotic ability. Furthermore,
older patients who have poor health cannot tolerate repeated
bleeding and cell infusion. Umbilical cord blood has become a
viable alternative source for transplantation due to its several
advantages over PB, such as widespread availability, absence of
donor and attrition, low risk of transmissible infectious diseases
in the cells, decreased GVHD and more importantly, a higher
frequency of immune accessory and effector cell precursors such as
dendritic cell precursors, natural killer cell precursors and
T-cell precursors (14). Hence,
they have attracted extensive concern.
CIK cells derived from cord blood (CB-CIK) have been
used in the clinic, and the safety, feasibility and availability
have been proven in clinical studies (15). However, CIK cells used in
preclinical and clinical studies are mainly derived from term
infant cord blood, from which an adequate and high quantity of
cells can be harvested for transfer. In recent years, the birth
rate of preterm infants has increased significantly. A
retrospective study was conducted on neonates born in 2005 in the
maternity departments of 72 urban hospitals from 22 provinces in
China (16). A total of 45,722
infants born between January 1, 2005 and December 31, 2005 were
enrolled. Preterm births accounted for 8.1%, and the incidence of
very low birth weight infants was 0.7%. Whether the phenotypic and
functional characterization of CIK cells derived from preterm and
term infant cord blood is consistent remains unclear. Moreover, the
biological characterization of CIK cells obtained from preterm
infant cord blood has rarely been reported. In the present study,
we cultured CIK cells from preterm and term infant cord blood and
analyzed the proliferation, phenotype, the secretion of cytokines
and the cytotoxicity in the two cell groups.
Materials and methods
Human materials
Between June 2013 and December 2013, umbilical cord
blood was obtained from 20 Cesarean deliveries at the Obstetrical
Department of the First Affiliated Hospital of Zhengzhou University
(Zhengzhou, China). The samples were classified into two groups
according to gestational age (GA): preterm infant (n=10, GA <37
weeks and mean 34+1 weeks+day) and term infant (n=10, GA ≥37 weeks
and mean 38+4 weeks+day). The maternal age range was 18 to 35
years. During pregnancy, there was no presence of hematological
system diseases, communicable diseases and premature rupture of
membranes. This study was conducted with written informed consent
obtained from the parturients and was approved by the Ethics
Committee of the First Affiliated Hospital of Zhengzhou
University.
Isolation and culture of CIK cells
CIK cells were generated as previously described
(15). In brief, cord blood MNCs
(CBMCs) were separated by Ficoll density gradient (Tianjin, China)
centrifugation, washed and resuspended at 2×106 cells/ml
in RPMI-1640 (Gibco, USA), consisting of 10% fetal calf serum (FCS;
Sigma, USA), 100 U/ml penicillin and 100 U/ml streptomycin (both
from Shijiazhuang, China). Recombinant human IFN-γ (1,000 U/ml)
(Beijing, China) was added on day 0. After 24 h of incubation,
1,000 U/ml recombinant human IL-2 (Beijing, China) and 25 ng/ml
anti-CD3 monoclonal antibody (Boehringer, Mannheim, Germany) were
added. Cells were incubated at 37°C in a humidified atmosphere of
5% CO2 and subcultured every 2–3 days in fresh complete
medium and IL-2 at 2×106 cells/ml.
K562 cell line
The K562 cell line (a chronic myeloid leukemia cell
line; Shanghai, China) was stored at −80°C. The recovered cells
were maintained in RPMI-1640 medium supplemented with 10% FCS, 100
U/ml penicillin and 100 U/ml streptomycin. Fresh complete medium
was replenished every 2 days.
Proliferation assay
CIK cells derived from preterm and term infant cord
blood were collected on day 5, washed and resuspended in RPMI-1640
at a final concentration of 1×106 cells/ml. In dark
conditions, 5 mmol/l 5,6-carboxyfluorescein succinimidyl ester
(CFSE; Invitrogen) was added, and cells were labeled for 10 min at
37°C. Staining was then stopped with 200 μl FCS for 8 min at 4°C.
CIK cells were washed twice with phosphate-buffered saline (PBS). A
total number of 1×105 labeled CIK cells were detected by
FACSCanto II flow cytometry (BD, USA), while the other CIK cells
were cultured on 24-well plates. IL-2 and fresh complete medium
were replenished every 2–3 days. A volume of 1×105 CIK
cells was collected daily from day 5 to 10, and the proliferation
index (PI) of the CIK cells was determined using FlowJo software
(BD).
Flow cytometry
The immunophenotype of CIK cells derived from
preterm and term infant cord blood was monitored by flow cytometry
at day 0, 7 and 14 using the following anti-human antibodies:
CD3-PE-Cy7, CD4-APC-Cy7, CD8-PerCP and CD56-PE. The activated or
inhibitory cell surface markers of CIK cells derived from preterm
and term infant cord blood were monitored at day 0 and 14 using the
following anti-human antibodies: CD27-FITC, CD28-APC and PD-1-PE.
The antibodies and isotype-matched monoclonal antibodies were
purchased from BD Biosciences (BD). A total of 5×105
cells was collected and washed twice, and then the supernatant was
discarded. A volume of 10 μl appropriate monoclonal antibody was
added and then the cells were incubated for 30 min at 37°C. After
incubation, the cells were washed twice and resuspended in 500 μl
PBS. Data acquisition was performed by flow cytometry and analyzed
using Diva software (both from BD). When detecting the
immunophenotype, we gated the population of CD3+ T
cells, and when detecting the expression of CD27, CD28 and PD-1, we
gated the population of CD3+CD8+ or
CD3+CD4+ T cells.
Determination of cytokine secretion
On day 12, CIK cells were adjusted to a final
concentration of 2×106 cells/ml and cultured in
RPMI-1640 medium consisting of 1,000 U/ml IL-2. After 48 h, the
supernatants were collected and stored at −80°C. To detect the
secretion of IL-10, IFN-γ and tumor necrosis factor (TNF)-α, an
enzyme-linked immunosorbent assay (ELISA) development with matched
antibody pairs (BioLegend, USA) was performed according to the
manufacturer’s instructions.
Cytotoxicity assay
We selected the logarithmic phase of K562 cells as
the target cells in this cytotoxicity assay. CIK and K562 cells
were co-cultured at an effector-target ratio of 20:1 with 5 μg/ml
1X Brefeldin A (BFA; BioLegend) for 5 h. For the positive control,
CIK cells (2×106) were stimulated 5 h with 1 mg/ml
phorbol myristate acetate (PMA), 1 mg/ml ionomycin (both from
Sigma) and 2.5 μg/ml 1X BFA. Cells were collect and resuspended in
PBS, mixed with 10 μl of CD107a-FITC (BD), CD3-PE-Cy7 and CD56-PE.
Incubation was carried out for 30 min at 37°C and then cells were
suspended in PBS. We also detected the expression of granzyme B and
perforin in the two groups. CIK cells (2×106) were
stimulated for 5 h with 1 mg/ml PMA, 1 mg/ml ionomycin and 2.5
μg/ml 1X BFA. After 5 h of incubation, CD3-PE-Cy7, CD4-APC-Cy7 and
CD8-PerCP were added, and then the cells were incubated for 15 min
at 37°C. We then sequentially added 4% paraformaldehyde (Tianjin,
China) and permeabilization wash buffer (1X; Caltag, USA) to the
system. The cells were washed twice with permeabilization wash
buffer (1X) and mixed with granzyme B-PE and perforin-FITC. The
expression levels of CD107a, granzyme B and perforin were detected
by flow cytometry. The antibodies and isotype-matched monoclonal
antibodies were purchased from BD Biosciences.
Statistical analysis
All data are presented as means ± SD. SPSS 16.0
(SPSS Inc., Chicago, IL, USA) was used for statistical analysis,
and GraphPad Prism version 5.0 (GraphPad Software Inc., USA) was
used for data plotting. Statistical differences between the groups
were analyzed using t-test. A P-value of <0.05 was considered to
indicate a statistically significant result.
Results
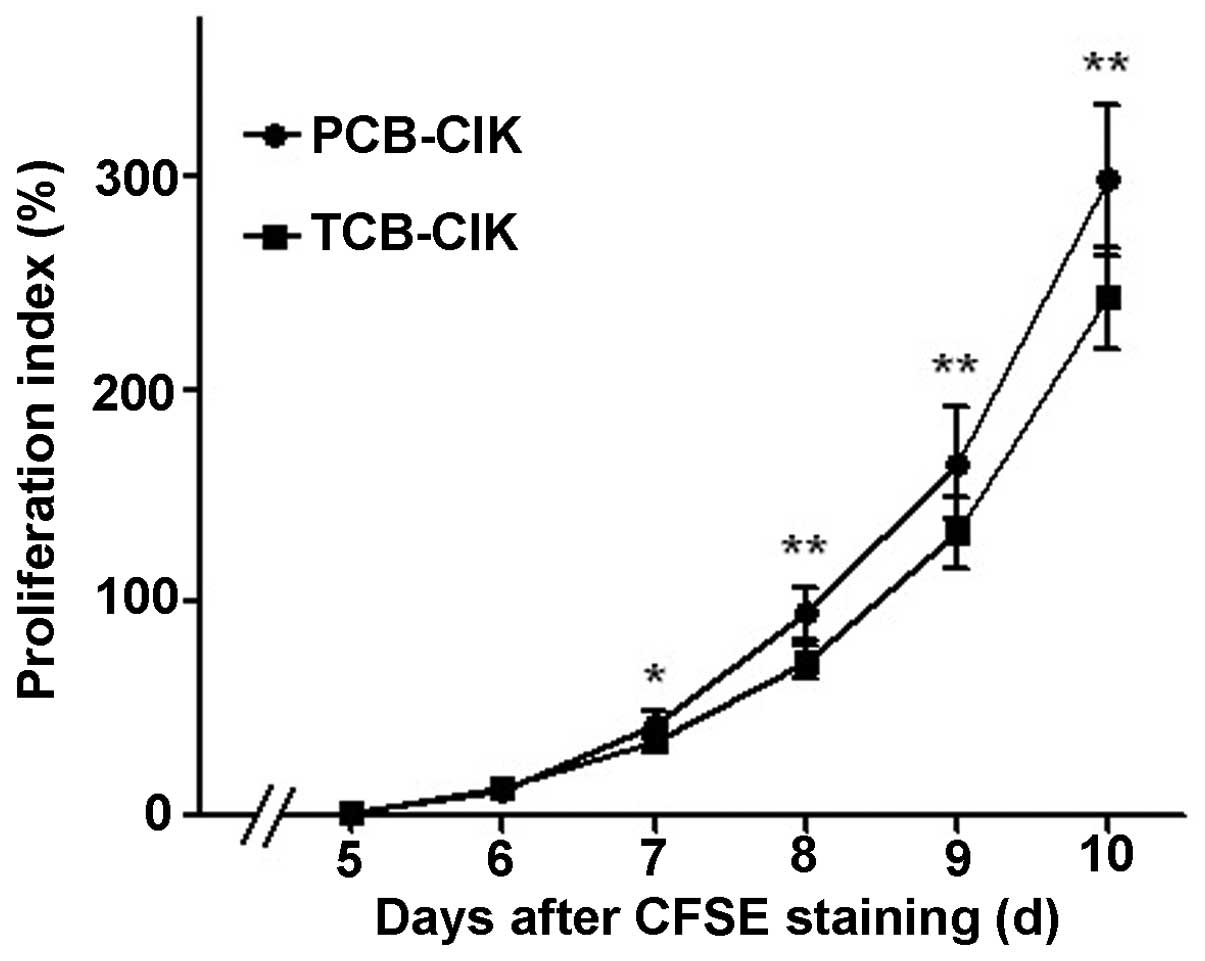
Cell proliferation of CIK cells
To detect the proliferation of PCB-CIK and TCB-CIK
cells, we used the CFSE assay to examine the change in the
proliferation of the two groups. The PI of PCB-CIK and TCB-CIK
cells had no difference on days 5 and 6, yet PCB-CIK showed a
significantly elevated PI compared with TCB-CIK on days 8, 9 and 10
(95.0±11.8 vs. 72.3±7.2%, 165.5±25.8 vs. 132.6±17.2%, 298.4±35.2
vs. 243.0±23.7%, respectively, all P<0.01, Fig. 1).
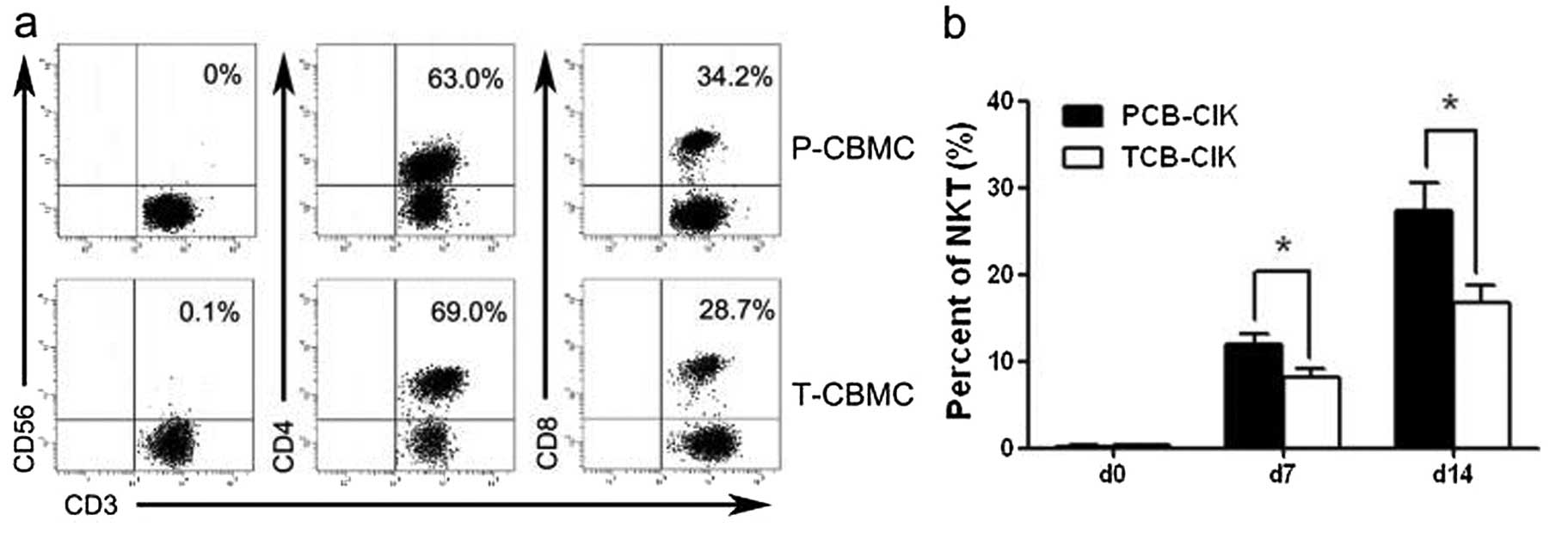
Phenotype of CIK cells
MNCs derived from preterm and term infant cord blood
(P-CBMCs and T-CBMCs) were isolated and induced to CIK cells. We
detected the phenotype by flow cytometry at days 0, 7 and 14
(Fig. 2a, c and e). Along with an
increase in the culture days, the proportion of
CD3−CD56+ (NK) cells gradually increased, and
the proportion of NK cells was higher in the group of preterm
infants (P<0.05, Table I). The
percentage of CD3+CD56+ T (NKT) cells was
higher in the PCB-CIK than that in the TCB-CIK cells on days 7 and
14 (P<0.05, Fig. 2b). The
percentage of CD3+CD4+ T cells was lower in
the PCB-CIK than that in the TCB-CIK cells on days 7 and 14
(P<0.05, Fig. 2d). Yet, the
percentages of NKT and CD3+CD4+ T cells had
no difference in the MNCs derived from preterm and term infants on
day 0 (P>0.05). The percentage of CD3+CD8+
T cells was higher in the PCB-CIK than that in the TCB-CIK cells on
day 14 (P<0.05, Fig. 2f).
However, the percentage of CD3+CD8+ T cells
did not differ in the CIK cells derived from preterm and term
infant cord blood on days 0 and 7 (P>0.05).
 | Table IPhenotypic characteristics of the
cells from preterm and term infant cord blood. |
Table I
Phenotypic characteristics of the
cells from preterm and term infant cord blood.
| Preterm infant | Term infant |
|---|
|
|
|
|---|
| Days |
CD3+CD56+ |
CD3+CD4+ |
CD3+CD8+ |
CD3+CD56+ |
CD3+CD4+ |
CD3+CD8+ |
|---|
| 0 | 0.2±0.2 | 71.1±9.3 | 26.9±9.0 | 0.4±0.4 | 72.1±7.0 | 26.2±7.4 |
| 7 |
11.9±4.2a |
32.9±9.9a | 67.7±9.9 | 8.1±3.5 | 43.1±9.4 | 59.9±10.0 |
| 14 |
27.2±11.0a |
16.9±7.7a |
84.1±7.1a | 16.7±7.1 | 23.8±5.5 | 78.0±5.1 |
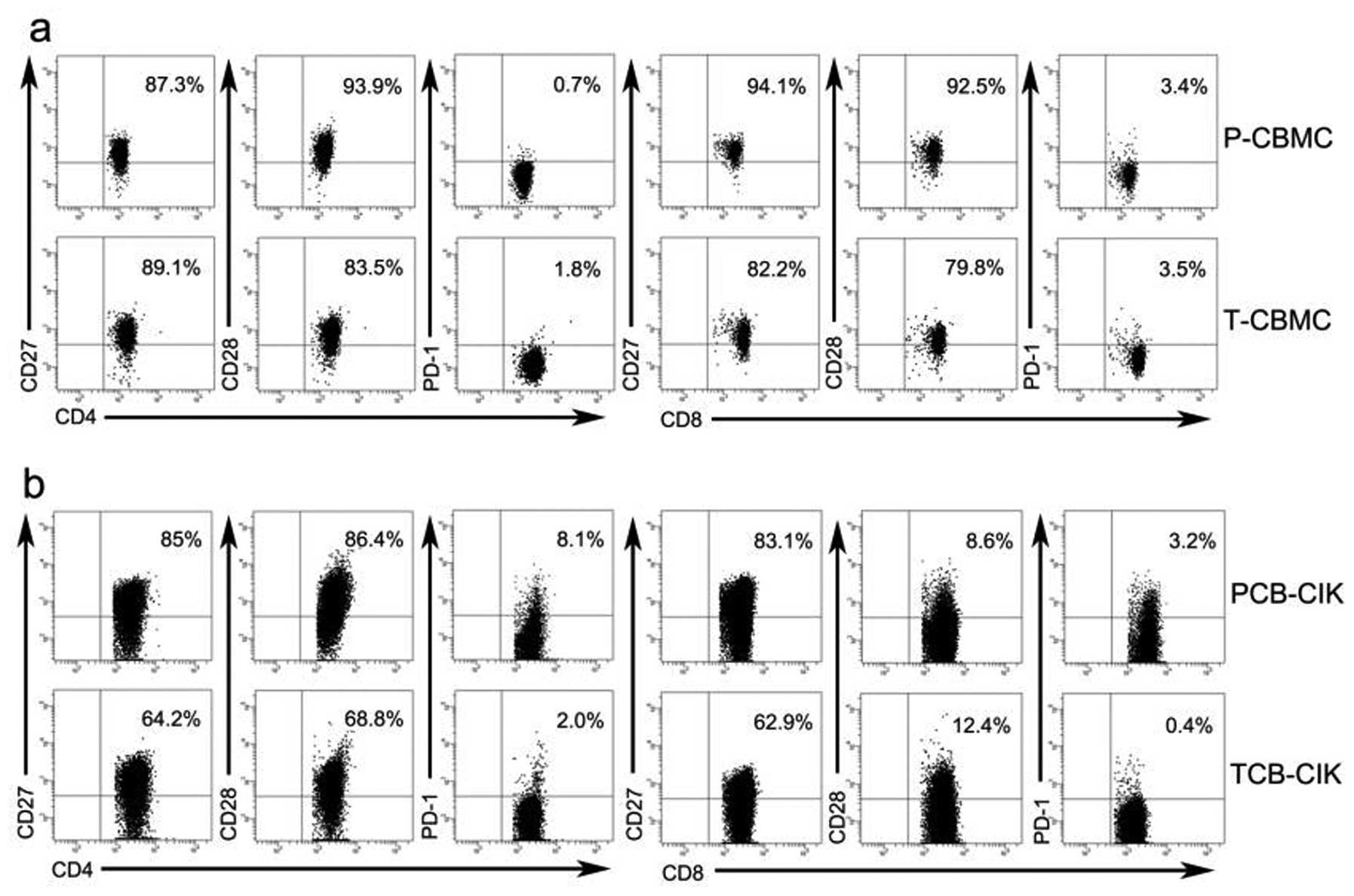
Activated and inhibitory cell surface
markers of CIK cells
We detected the cell surface markers by flow
cytometry on days 0 and 14. We gated the population of
CD3+CD8+ or CD3+CD4+ T
cells, and analyzed the expression of CD27, CD28 and PD-1 in CBMCs
and CIK cells (Fig. 3a, b and e).
On day 0, the proportions of activated
CD8+CD27+ (94.4±5.2 vs. 86.5±6.8%, P=0.004)
and CD8+CD28+ (92.2±5.4 vs. 82.5±8.9%,
P<0.05) T cells were higher in the P-CBMCs than the proportions
in the T-CBMCs, while the proportions of
CD8+PD-1+, CD4+CD27+,
CD4+CD28+ and CD4+PD-1+
T cells showed no significant differences between the P-CBMCs and
the T-CBMCs (P>0.05, Fig. 3c).
On day 14, the proportion of activated
CD8+CD27+ T cells was higher in the PCB-CIK
than that in the TCB-CIK cells (82.5±9.6 vs. 71.5±9.2%, P<0.05),
while the proportions of CD8+PD-1+,
CD8+CD28+, CD4+CD27+,
CD4+CD28+ and CD4+PD-1+
T cells showed no significant differences between the PCB-CIK and
TCB-CIK cells (P>0.05, Fig.
3d).
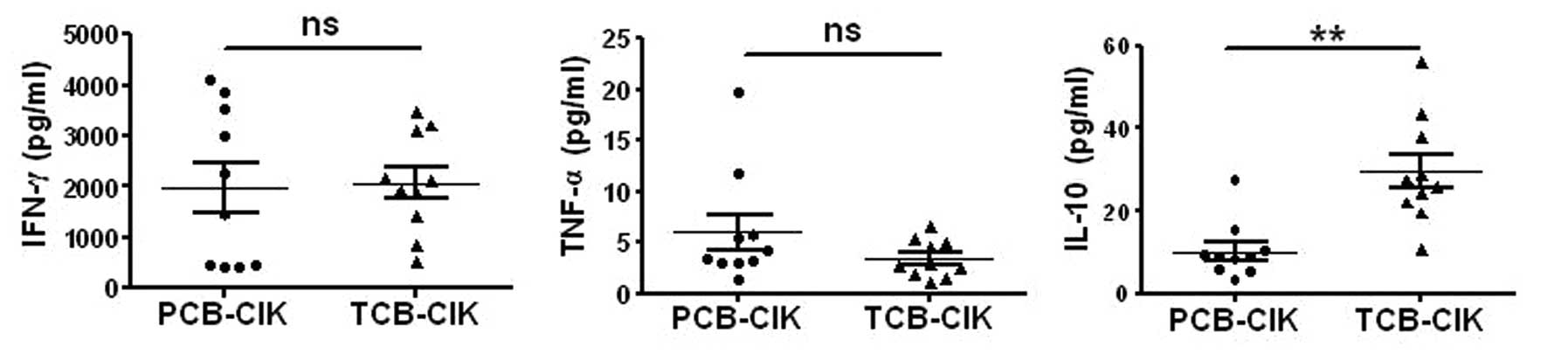
Cytokine production of CIK cells
We examined the production of IFN-γ, IL-10 and TNF-α
in the two groups. On day 14, the supernatants were collected and
assayed by ELISA to quantify the three cytokines. PCB-CIK cells
produced more TNF-α (6±5.5 pg/ml), yet the two groups had no
statistical significance (P=0.19). However, the secretion of IL-10
was lower in the PCB-CIK cells (10.2±6.8 vs. 29.6±13.0 pg/ml,
P<0.01, Fig. 4), while there was
no difference in the secretion level of IFN-γ (P>0.05).
Expression of granzyme B and perforin in
the CIK cells
To investigate the expression of granzyme B and
perforin, CIK cells were harvested on day 14. We analyzed the
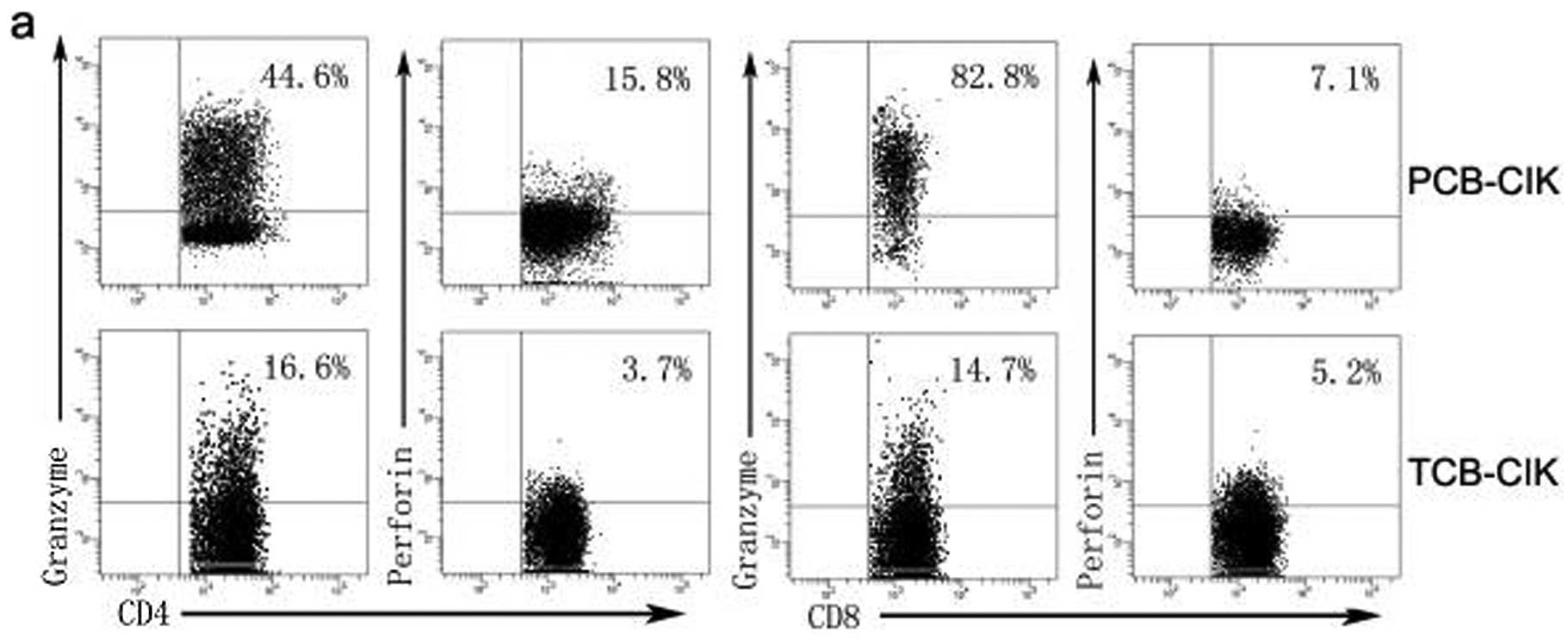
expression of granzyme B and perforin by flow cytometry (Fig. 5a). The percentage of
CD3+CD8+granzyme B+ was higher in
the PCB-CIK than that in the TCB-CIK cells (59.5±14.3 vs.
25.2±20.5%, P<0.05, Fig. 5b).
However, the percentages of
CD3+CD8+perforin+,
CD3+CD4+perforin+ and
CD3+CD4+granzyme+ had no
difference between the PCB-CIK and TCB-CIK cells (P>0.05).
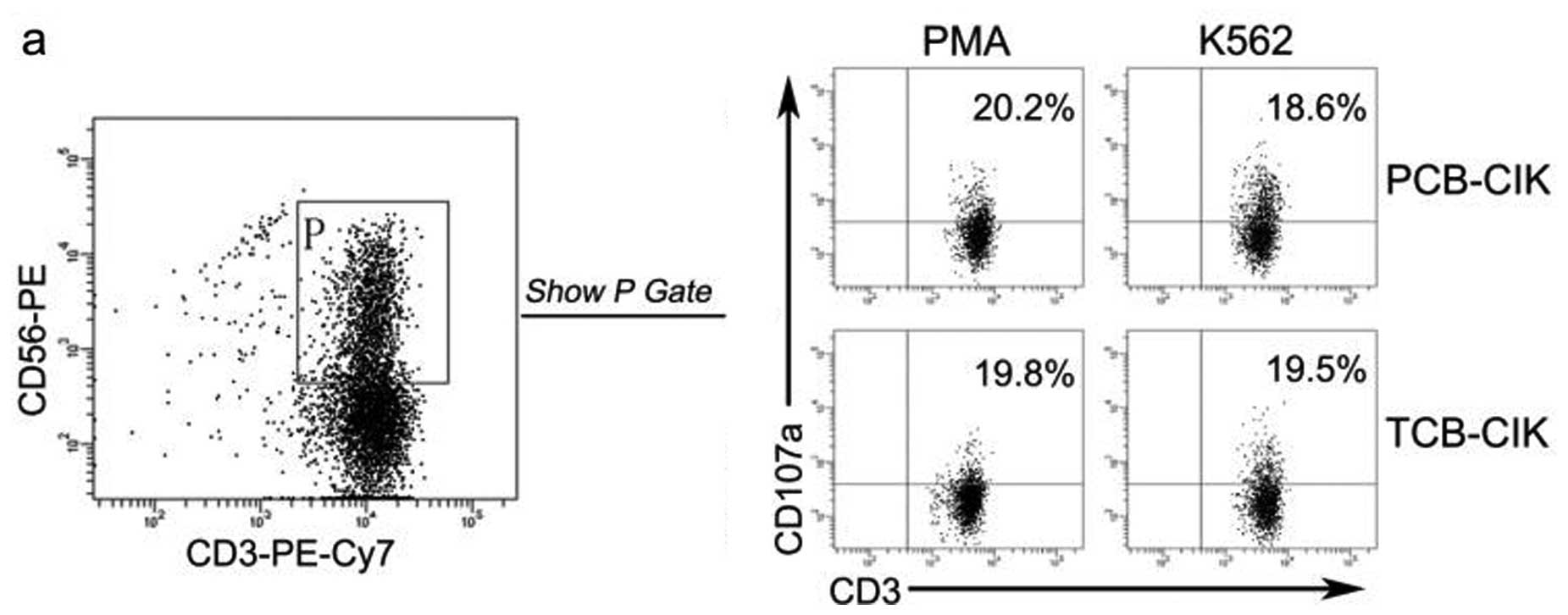
Cytotoxicity activity of CIK cells
CIK cells were co-cultured with logarithmic phase
K562 cells at an effector-target ratio of 20:1. The proportion of
activated CIK cells was identified by expression of CD107a, which
is located in the cytoplasm and transported to the cell surface
after activation-induced granule exocytosis (17). The percentage of CD107a+
cells exhibited no difference between the PCB-CIK and TCB-CIK cells
(Fig. 6a and b, P>0.05).
Discussion
Adoptive cellular immunotherapy has been available
for treating tumors for nearly 30 years. One of the first
prototypes used was lymphokine-activated killer (LAK) cells.
Subsequently, tumor-infiltrating lymphocytes (TILs), dendritic and
CIK cells appeared in succession (18). CIK cells are a heterogeneous
population with different cell phenotypes that are induced by
incubation of mononuclear cells with various cytokines, such as
IFN-γ, the anti-CD3 monoclonal antibody and IL-2 (4). As a new type of immune cell, CIK cells
have higher proliferation and cytotoxicity than other immune cells.
However, CIK cells that are used in preclinical or clinical study
are mainly derived from peripheral blood and term infant cord
blood. In recent years, the birth rate of preterm infants has
significantly increased. To our knowledge, no other studies have
been published on the comparison of PCB-CIK and TCB-CIK cells.
Whether the biological characteristics and function are different
between PCB-CIK and TCB-CIK cells is still unknown.
In the present study, we cultured PCB-CIK and
TCB-CIK cells for 14 days, and analyzed the proliferation,
phenotype and the expression of activated or inhibitory cell
surface markers. We also detected the production of cytokines, the
expression of granzyme B and perforin, and the cytotoxicity between
the two cell groups. Unlike previously described (19), we cultured the CIK cells for 14
days, since Helms et al (12) revealed that short-term cultured CIK
cells exhibit full cytotoxicity in vitro. Furthermore, many
preclinical and clinical studies have shown that CIK cells require
14 days of culture (20,21).
The main limitation preventing the successful
clinical translation of adoptive cellular immunotherapy strategies
is the obtainment of sufficient numbers of antitumor immune
effector cells. The results showed that PCB-CIK cells had a higher
proliferation rate than the TCB-CIK cells on days 7, 8, 9 and 10.
The possible cause is that preterm infant cord blood has more
immature and undifferentiated cells (22). However, the PCB-CIK cells were
difficult to cultivate. On day 6, PCB-CIK cells had a trend of a
low proliferation index, yet it was similar in the two groups. The
probable cause is that PCB-CIK cells have poor adaptability.
Kornacker et al (23)
reported that CIK cells derived from peripheral blood have a long
survival period and high ability for anti-apoptosis. Whether there
exist differences in PCB-CIK and TCB-CIK cells is unknown. Thus, we
must further explore the apoptosis of CIK cells from preterm and
term infant cord blood.
CIK cells consist of CD3+CD56+
double positive and NK cells, and CD3+CD56− T
lymphocytes. The antitumor activity of CIK cells seems to be
associated with the CD3+CD56+ subset, which
has an in vitro fold expansion that varies from a few to
>1,000-fold (24). The reason
for this variability is unclear, and additional strategies are
currently under investigation to improve the expansion rates. In
the present study, the content of CD3+CD56+
subsets of P-CBMCs and T-CBMCs was low, and there was no
statistical difference between the two groups. However, the
proportion of CD3+CD56+ was higher in the
PCB-CIK cells than that in the TCB-CIK cells on days 7 and 14. The
percentage of NKT cells in the present study was different from the
percentage found in previous research (25). It may be associated with the
activity of cytokines and individual differences. The results
demonstrate that PCB-CIK cells may be superior to TCB-CIK cells in
cytotoxicity.
In recent years, a number of studies have reported
the proliferation rate and immune phenotype of CB-CIK cells, yet
few studies have investigated activation or inhibitory cell surface
markers. In the present study, we detected the expression of CD27,
CD28 and PD-1. CD27 is a transmembrane homodimer with subunits of
55 kDa, which was first detected and named in 1987 (26). Importantly, CD27 is a member of the
TNF-receptor superfamily expressed on T cells that provide a
costimulatory signal, which is required for generation, activation
and differentiation of T cell immunity (27). CD28, which is expressed on the
surface of T cells, is required for T cell activation. CD28
improves the in vivo expansion and persistence of
CAR-modified T cells by providing costimulatory signals (28). The expression of CD28 and CD27 was
higher in the PCB-CIK than that in the TCB-CIK cells. PD-1 is a
member of the CD28/CTLA-4 superfamily. Studies have reported that
PD-1 and its ligands negatively regulate immune responses (29). In the present study, the expression
of PD-1 showed no significant difference between the PCB-CIK and
TCB-CIK cells. The results preliminary demonstrated that PCB-CIK
cells had more potential to promote the proliferation and
differentiation of T cells.
IFN-γ is an important factor for antitumor
activities, including the activation of CTLs and NK cells, the
induction of chemokines that mediate T-cell infiltration into
tumors, and the upregulation of MHC class I expression in tumor
cells (30). TNF-α, which consists
of 157 amino acids, is secreted by activated lymophocytes and
monocytes. TNF-α can enhance the antitumor ability of CIK cells
(31). In the present study, the
levels of IFN-γ and TNF-α in PCB-CIK and TCB-CIK cells had no
statistical significance. Yet there was a trend that PCB-CIK cells
could produce more TNF-α than CB-CIK. The reason may be that the
study had a small sample size. IL-10 is an inhibitory cytokine
(32), which can suppress the
secretion of IL-2 and IFN-γ. In this study, the secretion of IL-10
was lower in the PCB-CIK cells. The results indicate that the
activity of PCB-CIK cells may not be lower than TCB-CIK cells.
We evaluated the expression of granzyme B and
perforin in CIK cells. Granzyme B and perforin are required for the
cytotoxicity of CIK cells (33).
Perforin perforates the target cell membrane and forms a channel,
and then water and electrolytes move inside the cells and
subsequently wreak havoc. Granzyme B can induce the cellular
apoptosis of target cells by activating the apoptosis-related
enzyme system (34). The results
suggest that the expression of granzyme B was higher in the PCB-CIK
cells than that in the TCB-CIK cells. This directly demonstrate
that PCB-CIK cells have more potential to kill tumors. We also
detected the expression of CD107a, also known as
lysosome-associated membrane protein 1 (LAMP1), which is a high
glycosylation protein and a functional marker for the
identification of natural killer cell activity (18). The expression of CD107a had no
statistical significance between the two cell groups. The
expression of CD107a was in line with the secretion of granzyme B
and perforin, which is in accordance with previous research
(35). The results demonstrate that
PCB-CIK cells have the same antitumor function as TCB-CIK
cells.
In conclusion, the present study suggests that
PCB-CIK cells have a high proliferation rate and expression of
activated cell surface markers in vitro. Furthermore, the
cytotoxicity did not differ between the PCB-CIK and TCB-CIK cells.
All of the results suggest that PCB-CIK cells have the potential
for clinical application. However, it is difficult to culture
PCB-CIK cells, therefore further study is needed to explore the
culture optimization conditions.
Acknowledgements
The authors gratefully thank the Key Laboratory of
Clinical Medicine, First Affiliated Hospital of Zhengzhou
University for providing facilities. The present study was
supported by the Fund for Project of Newborn Disease, Key
Laboratory of the Ministry of Health (1213).
References
|
1
|
Bear AS, Hanley PJ, Bosque DM, et al: Low
rate of infusional toxicity after expanded cord blood
transplantation. Cytotherapy. 16:1153–1157. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Humphries C: Adoptive cell therapy: honing
that killer instinct. Nature. 504:S13–S15. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yee C: Adoptive T-cell therapy for cancer:
boutique therapy or treatment modality? Clin Cancer Res.
19:4550–4552. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Restifo NP, Dudley ME and Rosenberg SA:
Adoptive immunotherapy for cancer: harnessing the T cell response.
Nat Rev Immunol. 12:269–281. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Schmidt-Wolf IG, Negrin RS, Kiem HP, et
al: Use of a SCID mouse/human lymphoma model to evaluate
cytokine-induced killer cells with potent antitumor cell activity.
J Exp Med. 174:139–149. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pievani A, Borleri G, Pende D, et al:
Dual-functional capability of CD3+CD56+ CIK
cells, a T-cell subset that acquires NK function and retains
TCR-mediated specific cytotoxicity. Blood. 118:3301–3310.
2011.PubMed/NCBI
|
|
7
|
Liu L, Zhang W, Qi X, et al: Randomized
study of autologous cytokine-induced killer cell immunotherapy in
metastatic renal carcinoma. Clin Cancer Res. 18:1751–1759. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gammaitoni L, Giraudo L, Leuci V, et al:
Effective activity of cytokine-induced killer cells against
autologous metastatic melanoma including cells with stemness
features. Clin Cancer Res. 19:4347–4358. 2013. View Article : Google Scholar
|
|
9
|
Laport GG, Sheehan K, Baker J, et al:
Adoptive immunotherapy with cytokine-induced killer cells for
patients with relapsed hematologic malignancies after allogeneic
hematopoietic cell transplantation. Biol Blood Marrow Transplant.
17:1679–1687. 2011. View Article : Google Scholar
|
|
10
|
Ma YJ, He M, Han JA, et al: A clinical
study of HBsAg-activated dendritic cells and cytokine-induced
killer cells during the treatment for chronic hepatitis B. Scand J
Immunol. 78:387–393. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mesiano G, Todorovic M, Gammaitoni L, et
al: Cytokine-induced killer (CIK) cells as feasible and effective
adoptive immunotherapy for the treatment of solid tumors. Expert
Opin Biol Ther. 12:673–684. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Helms MW, Prescher JA, Cao YA, et al:
IL-12 enhances efficacy and shortens enrichment time in
cytokine-induced killer cell immunotherapy. Cancer Immunol
Immunother. 59:1325–1334. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chan JK, Hamilton CA, Cheung MK, et al:
Enhanced killing of primary ovarian cancer by retargeting
autologous cytokine-induced killer cells with bispecific
antibodies: a preclinical study. Clin Cancer Res. 12:1859–1867.
2006. View Article : Google Scholar
|
|
14
|
Frumento G, Zheng Y, Aubert G, et al: Cord
blood T cells retain early differentiation phenotype suitable for
immunotherapy after TCR gene transfer to confer EBV specificity. Am
J Transplant. 13:45–55. 2013. View Article : Google Scholar
|
|
15
|
Introna M, Pievani A, Borleri G, et al:
Feasibility and safety of adoptive immunotherapy with CIK cells
after cord blood transplantation. Biol Blood Marrow Transplant.
16:1603–1607. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li J, Wang QH, Wu HM, et al: A survey of
neonatal births in maternity departments in urban China in 2005.
Zhongguo Dang Dai Er Ke Za Zhi. 14:7–10. 2012.(In Chinese).
|
|
17
|
Krzewski K, Gil-Krzewska A, Nguyen V, et
al: LAMP1/CD107a is required for efficient perforin delivery to
lytic granules and NK-cell cytotoxicity. Blood. 121:4672–4683.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jiang J, Wu C and Lu B: Cytokine-induced
killer cells promote antitumor immunity. J Transl Med. 11:832013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Durrieu L, Gregoire-Gauthier J, Dieng MM,
et al: Human interferon-α increases the cytotoxic effect of
CD56+ cord blood-derived cytokine-induced killer cells
on human B-acute lymphoblastic leukemia cell lines. Cytotherapy.
14:1245–1257. 2012.
|
|
20
|
Li R, Wang C, Liu L, et al: Autologous
cytokine-induced killer cell immunotherapy in lung cancer: a phase
II clinical study. Cancer Immunol Immunother. 61:2125–2133. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang L, Ren B, Li H, et al: Enhanced
antitumor effects of DC-activated CIKs to chemotherapy treatment in
a single cohort of advanced non-small-cell lung cancer patients.
Cancer Immunol Immunother. 62:65–73. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
D’Alessio F, Mirabelli P, Gorrese M, et
al: Polychromatic flow cytometry analysis of CD34+
hematopoietic stem cells in cryopreserved early preterm human cord
blood samples. Cytometry A. 79:14–24. 2011.
|
|
23
|
Kornacker M, Moldenhauer G, Herbst M, et
al: Cytokine-induced killer cells against autologous CLL: direct
cytotoxic effects and induction of immune accessory molecules by
interferon-γ. Int J Cancer. 119:1377–1382. 2006.PubMed/NCBI
|
|
24
|
Zanon C, Stocchero M, Albiero E, et al:
Multivariate statistical data analysis as a tool to analyze ex vivo
expansion dynamics of cytokine-induced killer cells. Cytometry B
Clin Cytom. 86:257–262. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Niu Q, Wang W, Li Y, et al: Cord
blood-derived cytokine-induced killer cells biotherapy combined
with second-line chemotherapy in the treatment of advanced solid
malignancies. Int Immunopharmacol. 11:449–456. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
van Lier RA, Borst J, Vroom TM, et al:
Tissue distribution and biochemical and functional properties of
Tp55 (CD27), a novel T cell differentiation antigen. J Immunol.
139:1589–1596. 1987.PubMed/NCBI
|
|
27
|
Taraban VY, Rowley TF, Kerr JP, et al:
CD27 costimulation contributes substantially to the expansion of
functional memory CD8+ T cells after peptide
immunization. Eur J Immunol. 43:3314–3323. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pufnock JS, Cigal M, Rolczynski LS, et al:
Priming CD8+ T cells with dendritic cells matured using
TLR4 and TLR7/8 ligands together enhances generation of
CD8+ T cells retaining CD28. Blood. 117:6542–6551.
2011.
|
|
29
|
Prosser ME, Brown CE, Shami AF, et al:
Tumor PD-L1 co-stimulates primary human CD8+ cytotoxic T
cells modified to express a PD1:CD28 chimeric receptor. Mol
Immunol. 51:263–272. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Saga K, Tamai K, Yamazaki T, et al:
Systemic administration of a novel immune-stimulatory pseudovirion
suppresses lung metastatic melanoma by regionally enhancing IFN-γ
production. Clin Cancer Res. 19:668–679. 2013.PubMed/NCBI
|
|
31
|
Wang Y, Bo J, Dai HR, et al: CIK cells
from recurrent or refractory AML patients can be efficiently
expanded in vitro and used for reduction of leukemic blasts in
vivo. Exp Hematol. 41:241–252. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang YF, Kunda PE, Lin JW, et al:
Cytokine-induced killer cells co-cultured with complete tumor
antigen-loaded dendritic cells, have enhanced selective
cytotoxicity on carboplatin-resistant retinoblastoma cells. Oncol
Rep. 29:1841–1850. 2013.
|
|
33
|
Introna M, Franceschetti M, Ciocca A, et
al: Rapid and massive expansion of cord blood-derived
cytokine-induced killer cells: an innovative proposal for the
treatment of leukemia relapse after cord blood transplantation.
Bone Marrow Transplant. 38:621–627. 2006. View Article : Google Scholar
|
|
34
|
Kim TD, Lee SU, Yun S, et al: Human
microRNA-27a* targets Prf1 and GzmB expression to regulate NK-cell
cytotoxicity. Blood. 118:5476–5486. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Alter G, Malenfant JM and Altfeld M:
CD107a as a functional marker for the identification of natural
killer cell activity. J Immunol Methods. 294:15–22. 2004.
View Article : Google Scholar : PubMed/NCBI
|