Introduction
Tumor formation, evolution and metastasis are
affected by oncogene, tumor-suppressor gene and cell cycle-related
molecular. The regulation of transcription factors on potential
downstream target genes plays an important role in the occurrence
and progression of tumor (1).
Hypoxia is the main factor for tumor migration and invasion, and
hypoxia-inducible factor-1 (HIF-1) is involved in the processes
(2,3). In a previous study, we showed that
hypoxia can induce gastric cancer invasion and migration (4), and HIF-1 is a key molecular regulator
for the hypoxic signaling pathway (5).
HIF-1 upregulation is an early event of cells in
hypoxia that triggers hypoxia-related gene transcription (6). HIF-1 can promote tumor angiogenesis,
drug resistance and cell migration by binding with downstream HRE
(7,8). The exact mechanism involved in HIF-1
promotion of gastric cancer metastasis under hypoxic conditions
remains to be determined.
Under hypoxic conditions, a tumor microenvironment
is generated and tumor cells undergo epithelial-to-mesenchymal
transition (EMT) (9). During this
process, tumor cells can adjust to the newly formed
microenvironment and gain stem-cell characteristics that promote
differentiation and proliferation of tumor cells. The continuous
proliferation of tumor cells results in the center portion of
hypoxia, which greatly promotes EMT (10,11).
The results of a previous study demonstrated that KLF8 was highly
expressed in gastric cancer tissues and exhibited a poor prognosis
(12). KLF8 was initially
identified as a transcription repressor of Krüppel-like C2H2
zinc-finger transcription factor family proteins (13). KLF8 influenced other tumor
metastasis by EMT process (14) and
was found to play an active role in renal cell carcinoma in which
HIF-1 was highly expressed (15).
However, the exact mechanism of KLF8 in hypoxia-induced EMT in
gastric cancer remains to be elucidated.
In the present study, we confirmed that KLF8 is
important in gastric cancer invasion and metastasis. KLF8
siRNA-transfected SGC7901 cells showed less invasion and
metastasis. Subsequently, HIF-1 siRNA was used to examine the
effect of KLF8 expression and the mechanism of KLF8-induced EMT
under hypoxia. We also confirmed that the HIF-1 binding site was
located in the KLF8 promoter.
Materials and methods
Cell lines, cell culture conditions and
hypoxic treatment
The SGC7901 human gastric adenocarcinoma cell line
was obtained from the Academy of Military Medical Science (Beijing,
China). The MKN45 and AGS human gastric adenocarcinoma cell lines
were obtained from the Shanghai Cell Bank (Shanghai, China). The
cell lines were cultured in RPMI-1640 medium containing 10% fetal
bovine serum (FBS) plus L-glutamine, vimentin, sodium pyruvate,
non-essential amino acids, 100 IU/ml penicillin and 100 μg/ml
streptomycin (Sigma-Aldrich, St. Louis, MO, USA). For hypoxic
culture, the tumor cells were incubated in a hypoxic incubator
(Precision Scientific, Winchester, VA, USA) with 1% atmospheric
oxygen balanced by nitrogen and CO2 for 0–24 h.
HIF-1α siRNA plasmid constructs and
transfection
The PSilencerTM neo U6 2.1 vector, containing a
HIF-1α-specific targeting sequence (5′-AAAGAGGTGGATATGTCTGGG-3′ and
5′-TTTCTCCACCTATACAGACCC-3′) and a scramble sequence were
introduced into SGC7901 cells. To observe the change in HIF-1
target genes induced by HIF-1 in hypoxia, we introduced the HIF-1
siRNA vector pSilenser3.1/HIF-si and control scramble siRNA vector
pSilenser into SGC7901 cells, designated as SGC7901/HIF-si and
SGC7901/Scr-si, respectively. The cells were transfected by using
Lipofectamine 2000 (Invitrogen Life Technologies, Grand Island, NY,
USA) according to the manufacturer’s instructions and then treated
with 0.4 mg/ml of G418 to select the transfected cells. Stably
transfected cell clones were obtained by limiting the dilution
culture under the pressure of G418. To avoid the effects of clonal
variety, three random clones for each group were used in all the
experiments.
KLF8 cDNA vector and RNAi lentivirus
generation
KLF8-targeting oligonucleotides for generating cDNA
were designed from full-length KLF8 by Shanghai GeneChem Co., Ltd.
(Shanghai, China): forward, TCTGCAGGGACTACA GCAAG and reverse,
TCACATTGGTGAATCCGTCT; KLF8 siRNA1 forward: ACUUGGAGGUCCAACUUAATT
and reverse, UUAAGUUACCUCCAAGGTG; siRNA2 forward,
CGAUAUGGAUAACUCAUATT and reverse, UAU GAGUUUAUCCAUAUCGAC; siRNA 3
forward, CACUGG UUAAUGACAUCAATT and reverse, UUGAUGUCAUUA
AACAGUGCTA. After testing the overexpression and knockdown
efficiencies, stem-loop oligonucleotides were synthesized and
cloned into the lentivirus-based vector PsicoR (Addgene). A
non-targeting stem-loop DNA PsicoR vector was generated as a
negative control. Lentiviral particles were prepared as previously
described (12). Gastric cancer
cells were infected with KLF8 cDNA, siRNA-lentivirus or negative
control virus at 7 days and examined at 10 days. The SGC7901
gastric cancer cells were transfected with KLF8 si1, KLF8 si2, KLF8
si3 or empty vector and designated as SGC7901/KLF-si1,
SGC7901/KLF-si2, SGC7901/KLF-si3 and SGC7901/con. SGC7901 gastric
cancer cells transfected with KLF8 vector or empty vector were
designated as SGC7901/KLF and SGC7901/con.
Cell migration assay
Transwell chamber polycarbonate membranes were used
to determine the cell migration. SGC7901 cells
(5×104/well) were seeded on the top surface of the
24-well membrane with an 8-μm pore culture insert (Millipore,
Billerica, MA, USA). RPMI-1640 medium (0.5 ml) plus 10% FBS was
added to the bottom chamber as a chemoattractant. After 24 h
incubation, the filter membrane was fixed with 4% paraformaldehyde
and stained with hematoxylin and eosin. Non-invading cells on the
surface inside the chamber were wiped off using a cotton swab. The
number of migrated cells was counted by a microscope at 10 random
fields of vision for each chamber. Experiments were performed in
triplicate.
Cell invasion assay
The invasive activity of the SGC7901 cells was
measured using 24-well BioCoat Matrigel Invasion Chambers
(Millipore). In brief, the chamber containing the 8-μm pore size
filter was coated with Matrigel protein (BD Biosciences, Franklin
Lakes, NJ, USA). The filters were subsequently inserted into a
24-well culture plate. The SGC7901 cells were transfected with KLF
siRNA or scramble siRNA for ~48 h and then the cell density was
adjusted to ~5×104. The SGC7901 cell suspension was
inoculated into each insert well and incubated with RPMI-1640
medium containing 10% FBS for 24 h in 5% CO2 at 37°C.
After 24 h of incubation, the non-invading cells on the upper
surface inside the chamber were wiped off using a cotton swab. The
invading cells on the lower surface of the chamber were stained
with 0.2% crystal violet for 2 h. The number of cells adhering to
the lower surface of the filter was counted using a microcope at 10
random fields of vision for each chamber. Experiments were
performed in triplicate.
Metastasis in nude mice
Mice were handled using best humane practices, and
cared for in accordance with the NIH Animal Care and Use Committee
guidelines. The cells were harvested from tissue culture flasks
using trypsin and washed three times with PBS. The mice were
injected through the tail vein with 1×106 cells in 0.1
ml PBS, and monitored for overall health, and total body weight.
Mice-bearing luciferase-positive tumors were imaged 4–6 weeks after
injection with an IVIS Imaging System (Caliper Life Sciences, MA,
USA). The mice were subsequently sacrificed and each experimental
group comprised 5 mice.
Western blotting
The expression of KLF8, E-cadherin and fibronectin
in SGC7901 cells transfected with KLF-siRNA or Scr-siRNA under
normoxia conditions, as well as that of HIF-1, KLF8, E-cadherin,
keratin, fibronectin and vimentin in SGC7901 cells transfected with
KLF-siRNA, Scr-siRNA or HIF-siRNA under hypoxic conditions (1%
atmospheric oxygen) between 0 and 24 h was examined. Following
pre-treatment, whole cells (SGC7901, SGC7901/KLF-si,
SGC7901/Scr-si, SGC7901/HIF-si) were collected, respectively, and
lysed on ice for 30 min in lysis buffer [10 mM Tris (pH 8.0), 1 mM
ethylenediaminetetraacetic acid (EDTA), 400 mM NaCl, 10% glycerol,
0.5% NP-40, 5 mM sodium fluoride, 0.1 mM phenylmethylsulfonyl
fluoride, 1 mM dithiothreitol]. Equal amounts of protein (25 μg)
were loaded. For western blotting, mouse monoclonal antibodies
against KLF8 (1:200; Santa Cruz Biotechnology Inc., Santa Cruz, CA,
USA), E-cadherin (1:2,500; Abcam, Cambridge, UK), fibronectin
(1:2,500; Abcam), keratin (1:2,500; Abcam), vimentin (1:1,000;
Abcam), anti-HIF-1 (mouse mAb; 1:200; Chemicon Corporation Ltd.,
Bangladesh), anti-β-actin (1:5,000; Sigma-Aldrich), anti-mouse
secondary antibody (Ab; 1:2,000; Sigma-Aldrich), anti-rabbit
secondary antibody (Ab; 1:2,000; Sigma-Aldrich). The relative
content of each protein was detected by enhanced chemiluminescence
(ECL; Amersham, Arlington Heights, IL, USA).
Real-time RT-PCR
The whole cells (SGC7901, SGC7901/Scr-si and
SGC7901/HIF-si) were seeded after 0, 4, 8, 12 and 24 h exposure to
hypoxia (1% atmospheric oxygen), respectively. RNA was isolated
using RNAzol (Invitrogen, Carlsbad, CA, USA) according to the
manufacturer’s instructions. RNA (1 μg) was subjected to reverse
transcription, followed by cDNA strand synthesis at a dilution of
1:20. cDNA (10 μl) was used for TaqMan real-time RT-PCR to detect
mRNA expression. The ABI Prism 7700 Sequence Detection System (PE
Applied Biosystems) was subsequently applied. The primers used
were: KLF8 forward, 5′-TCTGCAGGGACTACAGCAAG and reverse,
5′-TCACATTGGTGAATCCGTCT; and GAPDH forward, 5′-TGGTATCGTGGAAGGACTCA
and reverse, 5′-CCAGTAGAGGCAGGGATGAT. Experiments were performed in
triplicate.
Immunofluorescence
SGC7901 cells were grown on 10-well glass microscope
slides (VWR International, Fontenay-sous-Bois, France) under
normoxic conditions, while SGC7901/Scr-si, SGC7901/KLF-si cells
were grown on 10-well glass microscope slides under hypoxic
conditions (1% atmospheric oxygen). The cells were fixed with 4%
neutral formalin for 20 min and then permeabilized with 0.2% Triton
X-100 in PBS for 10 min. After blocking with 3% bovine serum
albumin, the cells were incubated with E-cadherin and fibronectin
primary antibody (BD Biosciences) overnight at 4°C. After rinsing
three times in PBS, the cells were incubated with a secondary
antibody at room temperature. The nuclei were stained by DAPI
(Roche Applied Science, Basel, Switzerland). Immunostaining signal
and DAPI-stained nuclei were analyzed using a fluorescence
microscope.
HIF-1 binding site search
A genomic region of 2,000 bp upstream of the KLF8
transcriptional initiation site was determined using the NCBI
genomic BLAST program. The DNA strider 1.0 program was then applied
to identify putative HRE. The HIF-1-binding consensus sequence
BDCGTV (B is C/T/G; D is A/G/T; and V is G/C/A) was used as the key
criterion for searching functional HRE. It was established using
definitions of the consensus sequences, as previously provided
(16).
Dual luciferase reporter gene assay
To assay the transcriptional activity of KLF8 under
hypoxic conditions, SGC7901 cells in 24-well plates (50,000
cells/well) were co-transfected with or without HIF-1 siRNA1 or
KLF-Luc using Lipofectamine 2000 (Invitrogen). PRL-TK was the
control for transfection efficiency in DMEM without serum. The
construct KLF-Luc contained HRE-like or mutated sites from the KLF8
promoter nucleotides −133 to −128, as 520 bp containing CTCGTG to
CTTTTG, was synthesized and inserted into the pGL3 promoter vector
(Promega, Madison, WI, USA). After 48 h, luciferase activity was
measured and quantified in a luminometer using the Dual-Luciferase
Reporter Assay System (Promega). Experiments were performed in
triplicate. Results are expressed as a means of the ratio between
the firefly and Renilla luciferase activity.
Chromatin immunoprecipitation assay
HIF-1 binding to KLF8 promoter was analyzed by ChIP
in gastric cancer cells, using methodologies previously described
(5). Twenty-six SGC7901 cells were
fixed with 1% paraformaldehyde, and chromatin derived from isolated
nuclei was sheared by using a F550 microtip cell sonicator (Fisher
Scientific, Morris Plains, NJ, USA). Following centrifugation,
supernatants containing sheared chromatin were incubated with an
anti-HIF-1α antibody or control IgG. Protein A-Sepharose was added,
the incubation was continued overnight, and immune complexes were
subsequently eluted. The complexes were treated with RNase and
proteinase K and extracted with phenol/chloroform and then with
chloroform. DNA was precipitated, washed, dried, resuspended in
water and analyzed by PCR. The primers used in this analysis
spanned 216 bp around the first possible HIF-1α binding site
located −133 bp from the translation start site (sense,
5′-GCAGGTACAAGGGCTGGGTA-3′ and antisense,
5′-TGCCGGGACTGGGCTTTT-3′).
Statistical analysis
Results were presented as means ± SEM of at least
three independent experiments. Bands from western blotting or
RT-PCR analysis were quantified using Quantity One software
(Bio-Rad, Hercules, CA, USA). Relative protein or mRNA levels were
calculated relative to the amount of β-actin or GAPDH,
respectively. The difference between means was performed with
one-way analysis of variance or an unpaired t-test. All statistical
analyses were performed using SPSS 13.0 software (SPSS, Chicago,
IL, USA). P<0.05 was considered to indicate a statistically
significant result.
Results
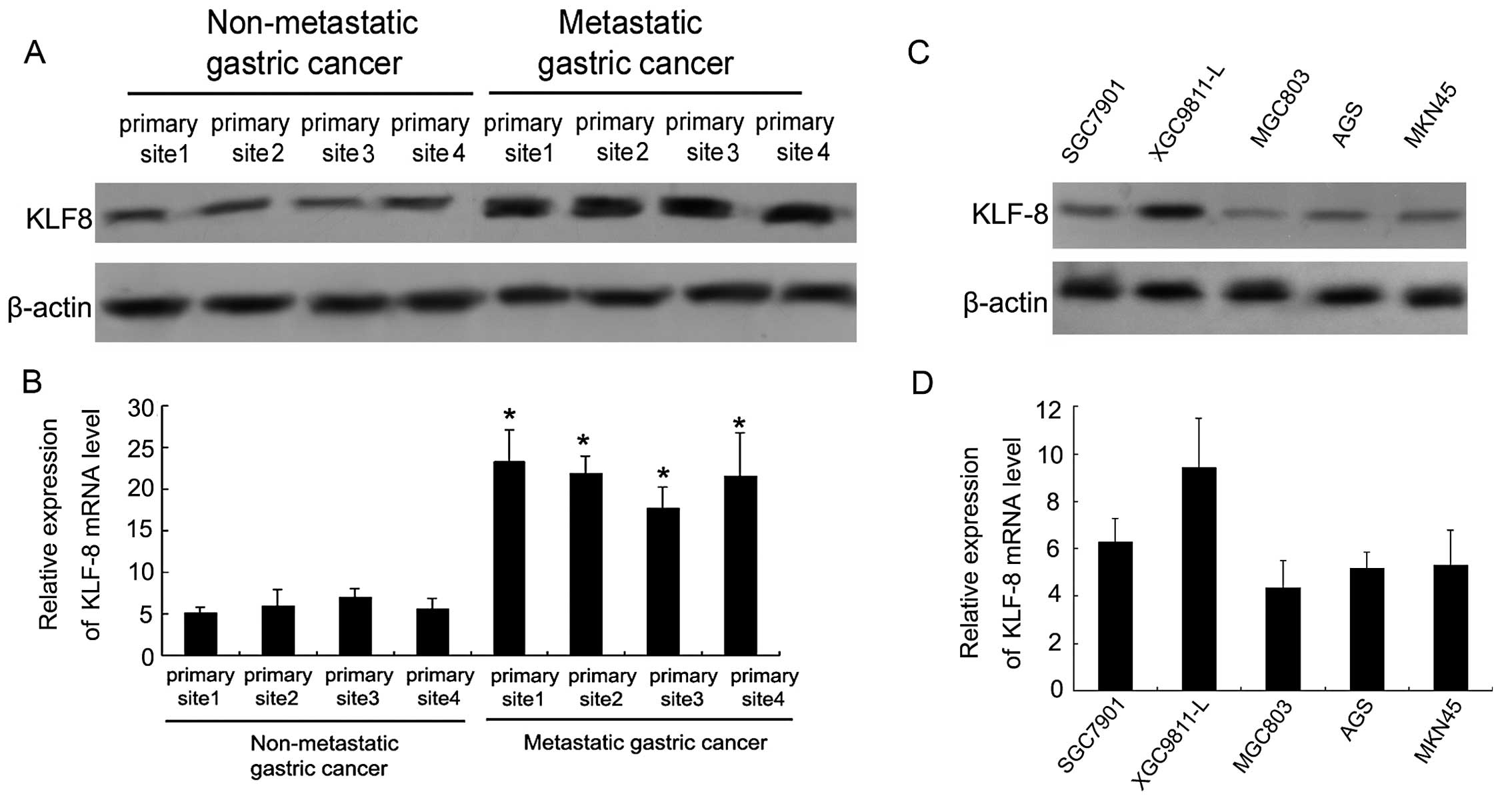
KLF8 is highly expressed in metastatic
gastric cancer tissues and cells
The expression of KLF8 protein in non-metastatic and
metastatic gastric cancer tissues was analyzed by western blotting.
Four tissue samples were obtained from different primary sites. As
shown in Fig. 1A, KLF8 protein
expression was significantly upregulated in four primary sites of
metastatic gastric cancer tissue compared with the non-metastatic
gastric cancer tissue. RT-PCR was also performed to examine the
KLF8 mRNA level of four different primary site tissues in
non-metastatic gastric cancer and metastatic gastric cancer.
Fig. 1B shows that KLF8 mRNA levels
in metastatic gastric cancer tissue samples were significantly
higher than the non-metastatic gastric cancer samples
(P<0.05).
KLF8 protein and mRNA were examined in five types of
gastric tumor cells of high and low metastatic potential. XGC9811-L
gastric tumor cells were found to have higher metastatic potential
than the remaining four gastric tumor cells. A significantly higher
expression level of KLF8 protein and mRNA in XGC9811-L cells is
shown in Fig. 1C and D. The low
metastatic potential cancer cells such as SGC7901, MGC803, AGS and
MKN45 exhibited a lower expression of KLF8 protein and mRNA
compared with XGC9811-L cells.
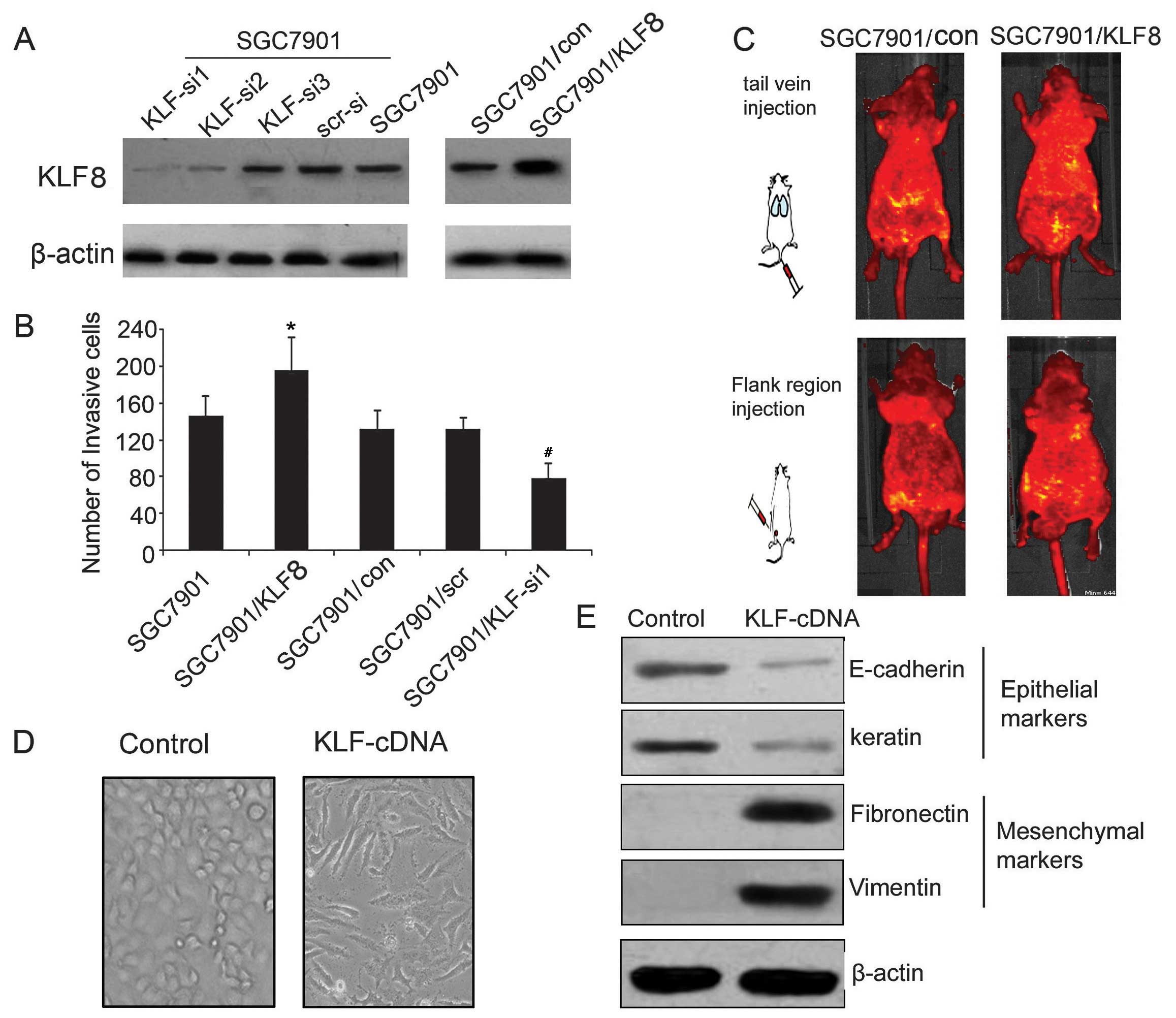
Forced overexpression of KLF8 promotes
gastric tumor cell metastasis and induces EMT
To evaluate the role of KLF8 in the promotion of the
invasion and migration in gastric cancer cells, a lentivirus-based
delivery system was used to transfer the KLF8 sense vector and
three siRNAs plasmids targeting KLF8 into SGC7901 cells. As shown
in Fig. 2A, SGC7901 cells infected
with KLF8 sense vector upregulated KLF8 expression. KLF8 siRNA1
markedly decreased KLF8 expression, while the effect of KLF-si3 was
minimal. As shown in Fig. 2B, for
each ×200 field under inverted light microscope, the invasion cells
were 148±13, 186±25, 127±14, 125±10, 86±7 of SGC7901, SGC7901/KLF,
SGC7901/con, SGC7901/KLF-scr and SGC7901/KLF-si1 cells,
respectively, suggesting that KLF8 siRNA1 significantly decreased
the invasion ability of gastric cancer.
To determine the role of KLF8 in tumor growth and
metastasis in vivo, SGC7901/KLF and SGC7901/con cells were
administered into nude mice by the tail vein and flank region
injection. Compared with the SGC7901/con group, the injection of
SGC7901/KLF led to a significant increase in metastatic lesions
(Fig. 2C). These results indicated
that KLF8 overexpression had the potential to promote the invasion,
migration and metastatic ability of SGC7901 cells in vitro
and in vivo.
Morphological assessment of SGC7901 cells following
infection with KLF8 cDNA indicated altered cell structure (Fig. 2D). Normal SGC7901 cells as control
were gathered and polyhedral-shaped, whereas SGC7901-KLF8 appeared
more scattered and spindle-shaped.
The protein expression of epithelial markers
E-cadherin and keratin, and mesenchymal markers fibronectin and
vimentin in SGC7901 cells transfected with KLF8 or negative control
vector were analyzed by western blotting. As shown in Fig. 2E, SGC7901 cells transfected with
KLF8 cDNA vector significantly downregulated E-cadherin and keratin
expression and upregulated fibronectin and vimentin. These results
suggested that KLF8 overexpression increased the invasion ability
and induced EMT in gastric cancer.
Hypoxia induces KLF8 expression and KLF8
siRNA reversed hypoxia-induced EMT
SGC7901 gastric tumor cells were cultured in
RPMI-1640 medium containing 10% FBS for 0–24 h under hypoxia and
the KLF8 and HIF-1 protein level were subsequently determined by
western blot analysis. The KLF8 and HIF-1 expression increased
under hypoxic condition in a time-dependent manner (Fig. 3A). RT-PCR was performed to examine
the mechanism by which the KLF8 mRNA was upregulated by hypoxia.
Fig. 3B shows that SGC7901 gastric
tumor cells exposed to 1% oxygen condition induced a gradually
increased expression of KLF8 mRNA in a time-dependent manner (means
± SEM, n=3). These results led to investigation of the possibility
of hypoxia-mediated transcriptional activation.
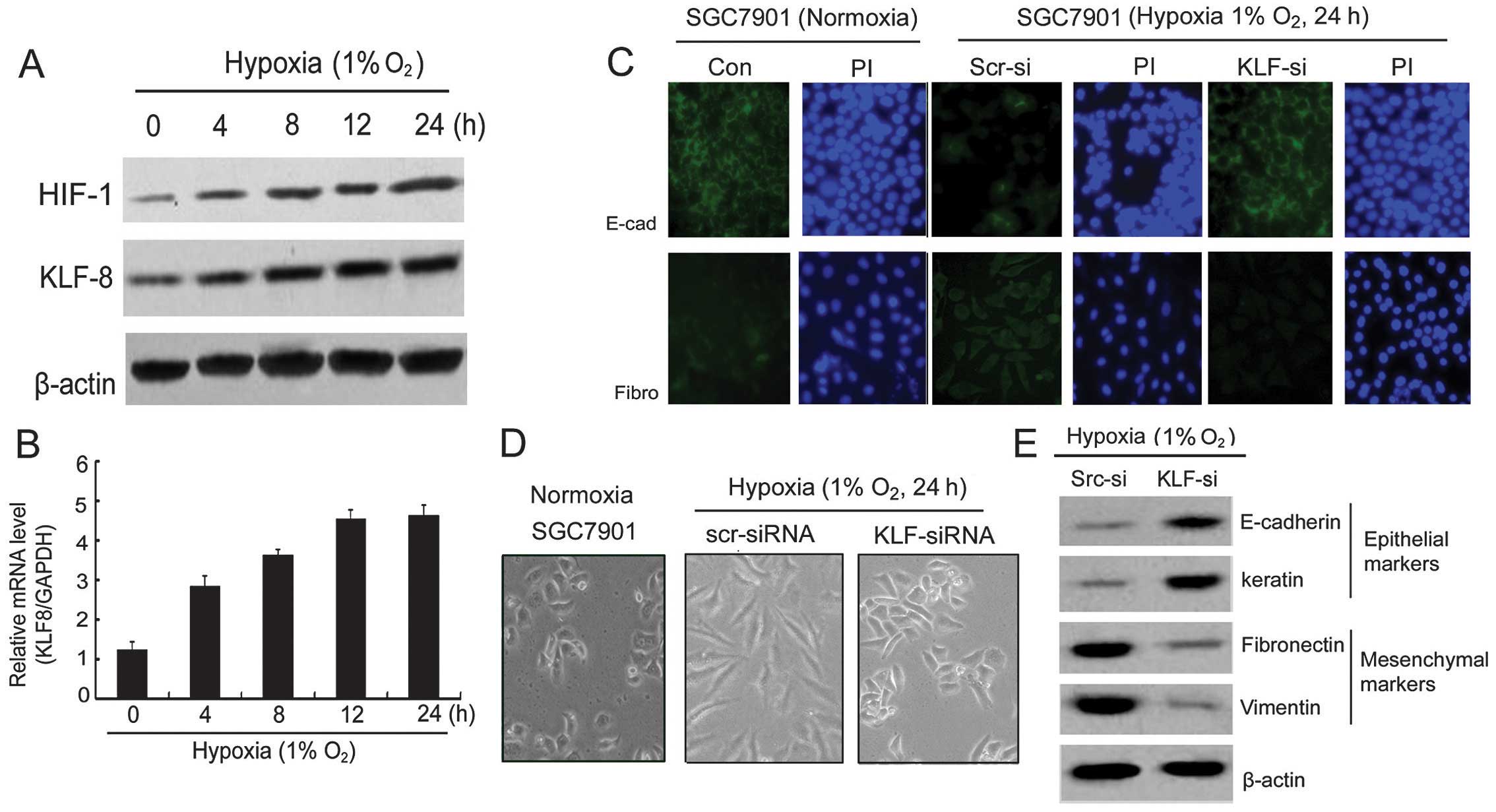 | Figure 3Hypoxia increases KLF8 expression and
inhibition of KLF8-reversed hypoxia induces EMT. (A) Western blot
analysis of HIF-1 and KLF8 expression in hypoxia at 0, 4, 8, 12 and
24 h. (B) Analysis of KLF8 mRNA level normalized by GAPDH of
SGC7901 cells in hypoxia at 0, 4, 8, 12 and 24 h. (C)
Immunofluorescence for E-cadherin and fibronectin expression in
SGC7901 cells in normoxia, SGC7901/KLF-si and SGC7901/Scr-si cells
in hypoxia for 24 h. (D) Morphological changes of SGC7901 cells in
normoxia or transfected with KLF-siRNA or Scr-siRNA in hypoxia.
Representative images are shown; original magnification, ×200. (E)
Western blot analysis of epithelial markers (i.e. E-cadherin and
keratin) and mesenchymal markers (i.e. fibronectin and vimentin) in
SGC7901 cells transfected with the HIF-siRNA or Scr-siRNA as a
control in hypoxia. KLF8, Krüppel-like factor 8; EMT,
epithelial-mesenchymal transition. |
The morphological assessment of SGC7901 cells after
24 h transfection with siRNA against KLF8 under hypoxia indicated
altered cell structure (Fig. 3D).
Normal SGC7901 cells in normoxia culture appreared
polyhedral-shaped. However, 24 h after KLF8 siRNA transfection,
these cells appeared polyhedral and more gathered under hypoxia for
24 h. The scramble siRNA produced a profound morphological change
from polyhedral to fusiform. To investigate the mechanism of
KLF8-induced EMT, we analyzed the expression of E-cadherin,
keratin, fibronectin and vimentin EMT markers. Hypoxia and
KLF8-siRNA affected the protein level of E-cadherin and
fibronectin, as confirmed by immunofluorescence staining (Fig. 3C). E-cadherin exhibited a strong
expression, while fibronectin exhibited a weak expression in the
SGC7901 cell line under normoxia. However, under hypoxic conditions
(1% O2, 24 h), immunofluorescence staining showed that
E-cadherin expression had almost disappeared in SGC7901/Scr-si
cells but markedly increased in SGC7901/KLF-si cells. By contrast,
fibronectin was increased significantly in SGC7901/Scr-si and
hardly observed in SGC7901/KLF-si cells. These findings suggested
one of the hallmarks of EMT. To confirm whether EMT can be reversed
by KLF-si RNA under hypoxia, we examined several epithelial and
mesenchymal markers. As shown in Fig.
3E, SGC7901/Scr-si cells exhibited an enhanced expression of
the fibronectin and vimentin mesenchymal markers, which had almost
disappeared for E-cadherin and keratin epithelial marker
expression. However, in SGC7901/KLF-si cells, the results were
completely reversed compared with SGC7901/Scr-si cells.
Consequently, the morphological and protein changes evident in the
SGC7901/Scr-si cells in hypoxia indicated that these cells had
undergone EMT, a process that can be reversed by KLF8-siRNA.
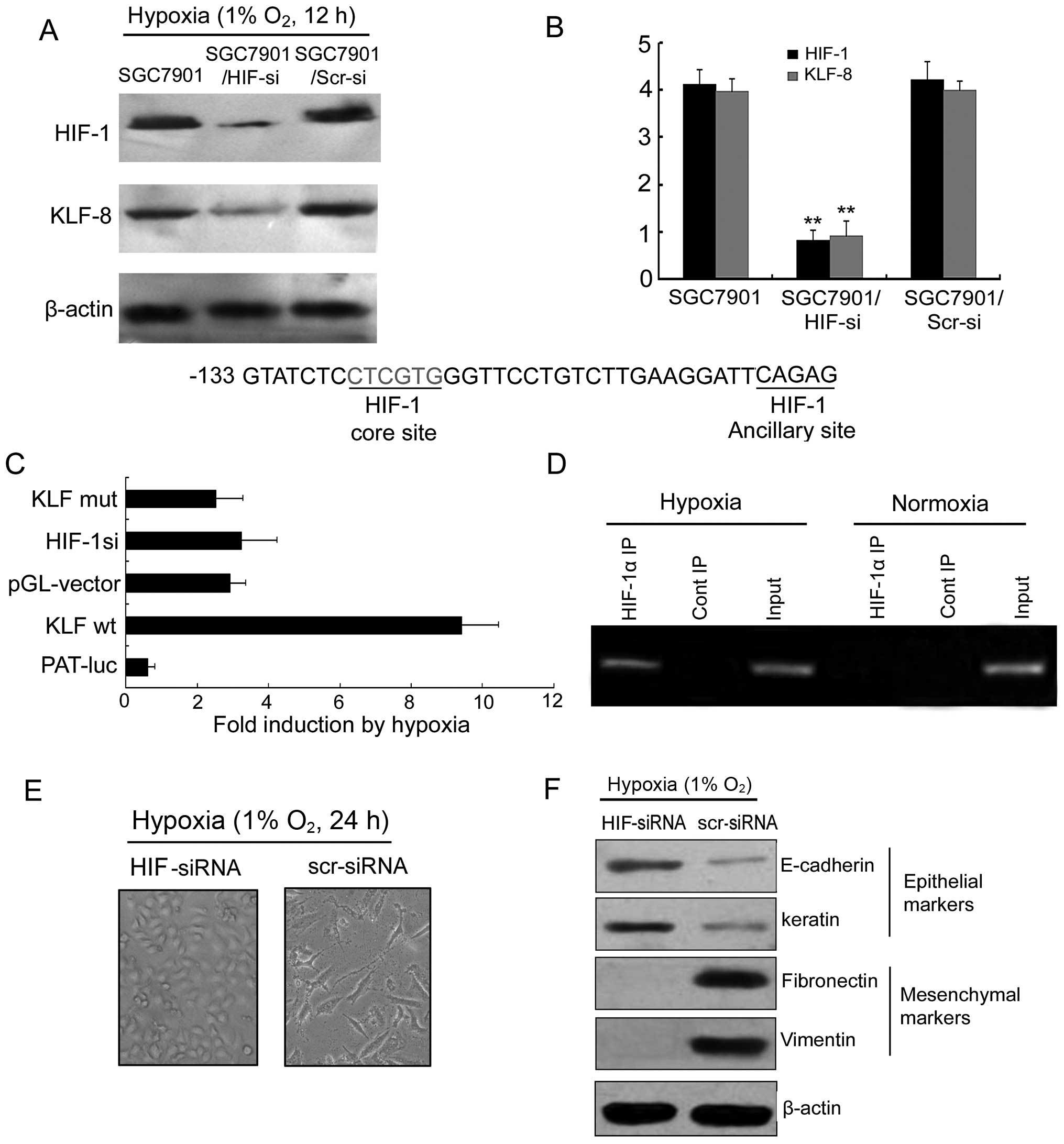
HIF-1 increases KLF8 expression and
transcriptional activity involved in hypoxia-induced EMT
To determine the role of HIF-1 in hypoxia-induced
KLF8 expression, HIF-1 siRNA vector was constructed and transfected
into the SGC7901 gastric cancer cell line. Western blotting and
RT-PCR assays revealed that inhibition of HIF-1 with targeted siRNA
could block hypoxia-induced KLF8 expression (Fig. 4A and B). Dual luciferase reporter
gene assay showed that luciferase activity transfected with
pGL3-KLF8wt was markedly increased compared with the pGL-vector
under hypoxia. However, luciferase activity transfected with
pGL3-KLF8mut had no significant difference compared with the
pGL-vector. Thus, it is possible that the HIF-1 binding site is
located −140 to −101 from the translation start site of the human
KLF8 gene. The possible site contains the HIF-1 core sequence
5′-CTCGTG-3′ between −126 to −121 and HIF-1 ancillary sequence
5′-CAG AG-3′ (Fig. 4C). Chromatin
immunoprecipitation (ChIP) was used to analyze the 1 HRE of KLF8
promoters in SGC7901 tumor cells exposed to hypoxia or normoxia. An
obvious band containing the possible binding site in the hypoxic
environment but not the normoxic one was observed (Fig. 3D). There was no band in control IgG
immunoprecipitates in the hypoxic and normoxic conditions. These
results show that the proximal HRE at −133 is the main HIF-1
binding site in the KLF8 promoter.
As shown in Fig. 4E and
F, in SGC7901/HIF-si cells, the EMT phenotype was almost
completely reversed compared with SGC7901/Scr-si cells.
Consequently, protein changes evident in the SGC7901/HIF-si cells
in hypoxia suggested that the EMT process can also be reversed by
HIF-siRNA.
Discussion
Gastric cancer is the second leading cause of
cancer-related mortalities worldwide (17,18).
As symptoms are often absent in the early stages of disease,
gastric cancer is usually diagnosed at an advanced stage and there
is <20% 5-year survival for patients. Gastric cancer deaths
results from cancer cell migration and invasion. Therefore,
exploring the mechanisms involved is essential in finding a cure.
Human KLF8 (ZNF741) was first cloned by PCR from K562 cells, a
human hematopoietic cell line (19). KLF8 is broadly expressed in human
tissues, with the greatest expression in kidney, heart and
placenta. Various KLF8 transcripts have been identified, and the
relative levels of the transcript expression appear to be similar
to that in various tissues. In a previous study, we confirmed that
KLF8 is important in SGC7901 gastric cell progression in
vitro and in vivo (12).
However, its underlying role in SGC7901 cells with invasion and
migration under normoxic or hypoxic conditions have yet to be
adequately elucidated. In the present study, we revealed a novel
role and mechanisms of KLF8 in the mediation of EMT in gastric
cancer and firstly confirmed a novel HRE binding site of HIF-1 in
the KLF8 promoter.
We confirmed KLF8 is important in gastric cancer
invasion and that the KLF8 siRNA-transfected SGC7901 cell line may
decrease gastric cancer cell invasion in vitro and in
vivo. In addition, the E-cadherin protein was found to be
downregulated, whereas fibronectin was upregulated following
infection of the KLF cDNA vector (20). Findings of recent studies have shown
that the epithelial cell adhesion molecule E-cadherin is often
downregulated during carcinoma progression and metastatic tumor
spread (21–24). These observations suggest that KLF8
plays a vital role in EMT in gastric cancer cells. It is well known
that the lack of E-cadherin plays an important role in the
development of gastric cancer and EMT. Three mechanisms of gene
deletion include mutation (25),
methylation (26) and E-box in the
promoter region (27,28). Results of a previous study have
shown that KLF17 is a negative regulation of E-cadherin by binding
the E-Box promoter in breast cancer. The association of KLF8 and
E-cadherin promoter remains to be investigated (29).
Hypoxia is an important characteristic in the tumor
tissue microenvironment and is associated with several
pathophysiological processes in tumor progression (30,31).
Malignant cancer cells form the tumor chords by colonic
proliferation at a diameter of 200–300 μm. When the radius of the
chords exceeds 300 μm, the cells in the middle of the chords may
undergo hypoxia. However, hypoxia leads to cell progression with a
more invasive phenotype. HIF is a transcription factor involved in
several biological processes. HIF-1 is able to generate the Warburg
effect through enhanced glucose uptake and the downregulation of
mitochondrial activities (32,33).
Previously, it was suggested that the hypoxic signal can be
triggered through HIF, resulting in the occurrence of EMT (34). Tumor growth factor-β was initially
confirmed as an inducer of EMT in normal mammary epithelial cells
and has since been shown to mediate the EMT in tumorigenesis
(35,36). Various transcription factors are
involved in TGFβ signals, including ZEB1 and SNAIL (37). Copple (36) suggested that hypoxia stimulates the
EMT by a HIF-1α- and TGF-β-dependent mechanism during liver
fibrosis in hepatocyte. In addition, Shah et al demonstrated
SNAIL is associated with breast cancer metastatic progression by
inhibiting E-cadherin, which is a hallmark of EMT (23). We employed a KLF8 siRNA to infect
SGC7901 in hypoxia to examine the effect of KLF8 silencing on EMT
markers and cell morphological changes. It was observed that
E-cadherin and keratin epithelial markers were overexpressed while
fibronectin and vimentin mesenchymal markers were downregulated in
the KLF-si group under hypoxia. Immunofluorescence also confirmed
that KLF8 siRNA is able to inhibit the EMT process in hypoxia.
These findings suggest that EMT-regulating transcription inhibitors
may be crucial in the repression of E-cadherin transcription in
gastric tumor. Additionally, KLF8 controls the transcription of
E-cadherin, fibronectin, EMT and invasion of its target gene
promoters.
Results of previous studies have shown that HIF
modulates EMT by regulating the activity of dominant transcription
factors including SNAIL, SLUG and ZEB1. EMT has been induced in
different cell lines under hypoxia or the continuous overexpression
of HIF (38–42). The association between EMT and
hypoxia-related transcription factors to EMT processes serve as a
molecular explanation for cancer metastasis and hypoxia. Our
results have shown that reduced endogenous HIF-1 in SGC7901/HIF-si
cells under hypoxia downregulated the KLF8 level, reversed the
expression of the epithelial markers and inhibited mesenchymal
marker expression. This findings suggests that HIF-siRNA caused the
shift of EMT to MET, and this process is accompanied by the
significant downregulation of KLF8. Our subsequent results revealed
that hypoxia-induced KLF8 was upregulated via a HIF-1-dependent
mechanism. There is a possible binding site of HIF-1 at the KLF8
gene promoter. The dual luciferase reporter gene assay results show
that hypoxia induced KLF8 promoter activity but was blocked by
HIF-1 siRNA. Mutant HRE reporter gene activity decreased. The ChIP
assay showed that the proximal HRE at −133 is the main HIF-1
binding site in the KLF8 promoter.
In summary, we have identified KLF8, which was a
representative transcriptional factor in the regulation of EMT and
SGC7901 gastric cancer cell invasion and migration. The results
confirmed that KLF8 is a new HIF-1 target gene that may be used to
elucidate hypoxia-induced EMT in SGC7901 cells. These results
suggest that HIF-1-dependent induction of KLF8 in hypoxia is a
promising therapeutic target for gastric cancer or other solid
tumors. However, the mechanisms underlying the role of KLF8 in
these processes remains to be determined.
Acknowledgements
This study was supported by the National Nature
Science Foundation of China (nos. 81272349 and 81372608).
References
|
1
|
Chaffer CL and Weinberg RA: A perspective
on cancer cell metastasis. Science. 331:1559–1564. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Carmeliet P, Dor Y, Herbert JM, et al:
Role of HIF-1α in hypoxia-mediated apoptosis, cell proliferation
and tumour angiogenesis. Nature. 394:485–490. 1998.
|
|
3
|
Min JH, Yang H, Ivan M, Gertler F, Kaelin
WG Jr and Pavletich NP: Structure of an HIF-1α-pVHL complex:
hydroxy-proline recognition in signaling. Science. 296:1886–1889.
2002.
|
|
4
|
Liu L, Sun L, Zhao P, et al: Hypoxia
promotes metastasis in human gastric cancer by up-regulating the
67-kDa laminin receptor. Cancer Sci. 101:1653–1660. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu L, Zhang H, Sun L, et al: ERK/MAPK
activation involves hypoxia-induced MGr1-Ag/37LRP expression and
contributes to apoptosis resistance in gastric cancer. Int J
Cancer. 127:820–829. 2010.PubMed/NCBI
|
|
6
|
Wang Y, Li Z, Zhang H, et al: HIF-1α and
HIF-2α correlate with migration and invasion in gastric cancer.
Cancer Biol Ther. 10:376–382. 2010.
|
|
7
|
Liu L, Ning X, Sun L, et al:
Hypoxia-inducible factor-1α contributes to hypoxia-induced
chemoresistance in gastric cancer. Cancer Sci. 99:121–128.
2008.
|
|
8
|
Semenza GL: Targeting HIF-1 for cancer
therapy. Nat Rev Cancer. 3:721–732. 2003. View Article : Google Scholar
|
|
9
|
Lundgren K, Nordenskjöld B and Landberg G:
Hypoxia, Snail and incomplete epithelial-mesenchymal transition in
breast cancer. Br J Cancer. 101:1769–1781. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang MH, Wu MZ, Chiou SH, et al: Direct
regulation of TWIST by HIF-1α promotes metastasis. Nat Cell Biol.
10:295–305. 2008.
|
|
11
|
Yang MH and Wu KJ: TWIST activation by
hypoxia inducible factor-1 (HIF-1): implications in metastasis and
development. Cell Cycle. 7:2090–2096. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu L, Liu N, Xu M, et al:
Lentivirus-delivered Krüppel-like factor 8 small interfering RNA
inhibits gastric cancer cell growth in vitro and in vivo. Tumour
Biol. 33:53–61. 2012.
|
|
13
|
Wang X and Zhao J: KLF8 transcription
factor participates in oncogenic transformation. Oncogene.
26:456–461. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang X, Zheng M, Liu G, et al:
Krüppel-like factor 8 induces epithelial to mesenchymal transition
and epithelial cell invasion. Cancer Res. 67:7184–7193. 2007.
|
|
15
|
Fu WJ, Li JC, Wu XY, et al: Small
interference RNA targeting Krüppel-like factor 8 inhibits the renal
carcinoma 786-0 cells growth in vitro and in vivo. J Cancer Res
Clin Oncol. 136:1255–1265. 2010.
|
|
16
|
Fink T, Kazlauskas A, Poellinger L,
Ebbesen P and Zachar V: Identification of a tightly regulated
hypoxia-response element in the promoter of human plasminogen
activator inhibitor-1. Blood. 99:2077–2083. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shah MA and Ajani JA: Gastric cancer - an
enigmatic and heterogeneous disease. JAMA. 303:1753–1754. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar
|
|
19
|
Hu JH, Navas P, Cao H, Stamatoyannopoulos
G and Song CZ: Systematic RNAi studies on the role of Sp/KLF
factors in globin gene expression and erythroid differentiation. J
Mol Biol. 366:1064–1073. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sun Z, Han Q, Zhou N, et al: MicroRNA-9
enhances migration and invasion through KLF17 in hepatocellular
carcinoma. Mol Oncol. 7:884–894. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Deep G, Jain AK, Ramteke A, et al: SNAI1
is critical for the aggressiveness of prostate cancer cells with
low E-cadherin. Mol Cancer. 13:372014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chan SW, Kallarakkal TG and Abraham MT:
Changed expression of E-cadherin and galectin-9 in oral squamous
cell carcinomas but lack of potential as prognostic markers. Asian
Pac J Cancer Prev. 15:2145–2152. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shah P, Gau Y and Sabnis G: Histone
deacetylase inhibitor entinostat reverses epithelial to mesenchymal
transition of breast cancer cells by reversing the repression of
E-cadherin. Breast Cancer Res Treat. 143:99–111. 2014. View Article : Google Scholar
|
|
24
|
Wang Y, Dong H, Xu M, et al: 37-kDa
laminin receptor precursor promotes lung adenocarcinoma cell
invasion and metastasis by epithelial-to-mesenchymal transition.
Cancer Gene Ther. 21:150–157. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Machado JC, Soares P, Carneiro F, et al:
E-cadherin gene mutations provide a genetic basis for the
phenotypic divergence of mixed gastric carcinomas. Lab Invest.
79:459–465. 1999.PubMed/NCBI
|
|
26
|
Leung WK, Yu J, Ng EK, et al: Concurrent
hypermethylation of multiple tumor-related genes in gastric
carcinoma and adjacent normal tissues. Cancer. 91:2294–2301. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
D’souza B and Taylor-Papadimitriou J:
Overexpression of ERBB2 in human mammary epithelial cells signals
inhibition of transcription of the E-cadherin gene. Proc Natl Acad
Sci USA. 91:7202–7206. 1994.PubMed/NCBI
|
|
28
|
Dong C, Wu Y, Wang Y, et al: Interaction
with Suv39H1 is critical for Snail-mediated E-cadherin repression
in breast cancer. Oncogene. 32:1351–1362. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Gumireddy K, Li A, Gimotty PA, et al:
KLF17 is a negative regulator of epithelial-mesenchymal transition
and metastasis in breast cancer. Nat Cell Biol. 11:1297–1304. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Muñoz-Nájar UM, Neurath KM, Vumbaca F and
Claffey KP: Hypoxia stimulates breast carcinoma cell invasion
through MT1-MMP and MMP-2 activation. Oncogene. 25:2379–2392.
2006.PubMed/NCBI
|
|
31
|
McMahon S, Grondin F, McDonald PP, Richard
DE and Dubois CM: Hypoxia-enhanced expression of the proprotein
convertase furin is mediated by hypoxia-inducible factor-1: impact
on the bioactivation of proproteins. J Biol Chem. 280:6561–6569.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Stoeltzing O, McCarty MF, Wey JS, et al:
Role of hypoxia-inducible factor 1α in gastric cancer cell growth,
angiogenesis, and vessel maturation. J Natl Cancer Inst.
96:946–956. 2004.
|
|
33
|
Kong T, Eltzschig HK, Karhausen J, Colgan
SP and Shelley CS: Leukocyte adhesion during hypoxia is mediated by
HIF-1-dependent induction of β2 integrin gene
expression. Proc Natl Acad Sci USA. 101:10440–10445.
2004.PubMed/NCBI
|
|
34
|
Higgins DF, Kimura K, Bernhardt WM, et al:
Hypoxia promotes fibrogenesis in vivo via HIF-1 stimulation of
epithelial-to-mesenchymal transition. J Clin Invest. 117:3810–3820.
2007.PubMed/NCBI
|
|
35
|
Zhou G, Dada LA, Wu M, et al:
Hypoxia-induced alveolar epithelial-mesenchymal transition requires
mitochondrial ROS and hypoxia-inducible factor 1. Am J Physiol Lung
Cell Mol Physiol. 297:L1120–L1130. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Copple BL: Hypoxia stimulates hepatocyte
epithelial to mesenchymal transition by hypoxia-inducible factor
and transforming growth factor-β-dependent mechanisms. Liver Int.
30:669–682. 2010.PubMed/NCBI
|
|
37
|
Leivonen SK and Kähäri VM: Transforming
growth factor-β signaling in cancer invasion and metastasis. Int J
Cancer. 121:2119–2124. 2007.
|
|
38
|
Shields MA, Dangi-Garimella S, Krantz SB,
Bentrem DJ and Munshi HG: Pancreatic cancer cells respond to type I
collagen by inducing snail expression to promote membrane type 1
matrix metalloproteinase-dependent collagen invasion. J Biol Chem.
286:10495–10504. 2011. View Article : Google Scholar
|
|
39
|
Kang Y and Massagué J:
Epithelial-mesenchymal transitions: twist in development and
metastasis. Cell. 118:277–279. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Aigner K, Dampier B, Descovich L, et al:
The transcription factor ZEB1 (δEF1) promotes tumour cell
dedifferentiation by repressing master regulators of epithelial
polarity. Oncogene. 26:6979–6988. 2007.
|
|
41
|
Guaita S, Puig I, Franci C, et al: Snail
induction of epithelial to mesenchymal transition in tumor cells is
accompanied by MUC1 repression and ZEB1 expression. J
Biol Chem. 277:39209–39216. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Peinado H and Cano A: A hypoxic twist in
metastasis. Nat Cell Biol. 10:253–254. 2008. View Article : Google Scholar : PubMed/NCBI
|