Introduction
Heat shock protein 90 (Hsp90) is a molecular
chaperone with a key role in the preservation of the conformation,
stability and function of its client proteins (1). The largest groups of client proteins
associated with Hsp90 (e.g., IKK, GSK3, CHK1 and Raf-1) are protein
kinases and transcription factors that are important in cellular
carcinogenesis (2). Due to the
critical roles of Hsp90 client proteins in cancer cell growth
arrest, apoptosis and other cancer hallmarks, the inhibition of
Hsp90 has received considerable attention as a potential target for
cancer treatment (2–4). 17-AAG is the first Hsp90 inhibitor to
undergo clinical research, but has several issues related to poor
solubility, unacceptable hepatotoxicity and limited bioavailability
(5–7). These drawbacks have catalyzed efforts
to explore novel scaffolds with improved pharmacological properties
for clinical applications.
SNX-2112, a novel Hsp90 inhibitor with the
2-aminobenza-mide scaffold, has been widely used in cancer research
(8–12). SNX-2112 exerts its growth inhibitory
and apoptosis-inducing activity on many human cancer cells
(1,8,11,13).
However, subsequent development has been discontinued due to ocular
toxicity observed in animal models and in a separate phase I study
(14). Developing alternatives of
SNX-2112 based on the 2-aminobenzamide scaffold may be a feasible
method for meeting clinical needs. SNX-7081 is another Hsp90
inhibitor derived from 2-aminobenzamide and optimized by SAR
explorations (15), with a
side-chain of indole instead of indazole at SNX-2112. To the best
of our knowledge, SNX-7081 has not attracted enough attention in
cancer therapy, and is only limited to investigations on its
potential in chronic lymphocytic leukemia and inflammatory diseases
(16–19).
In the present study, we tested the anticancer
effects and examined the mechanisms of SNX-7081 using several human
cancer cells in vitro, with SNX-2112 as a reference. To
demonstrate more details involved in the antitumor effects of the
two compounds, we investigated Hsp90 affinity, cell growth, cell
cycle, apoptosis and the expression of Hsp90 clients in various
cancer cells.
Materials and methods
Reagents and antibodies and cells
SNX-7081 and SNX-2112 were prepared in our
laboratory with purities of >98.0% according to a previously
described procedure (20,21), dissolved in 10 mM dimethyl sulfoxide
(DMSO) as a stock solution and stored at −20°C. Antibodies against
IKKα, Raf-1, CHK1 and GSK3β were purchased from Epitomics
(Burlingame, CA, USA), and the glyceraldehyde-3-phosphate
dehydrogenase (GAPDH) antibody was obtained from Millipore
(Billerica, MA, USA).
Cell culture
Eleven cell lines, including eight human cancer cell
lines and three human normal cell lines, were used in the present
study (Table I). Primary HDF cells
were isolated from the foreskins of newborns using a previously
reported procedure (22,23). All other cell lines were purchased
from the Cell Bank of the China Science Academy (Shanghai, China).
CML K562 cells, A375 melanoma cells, the normal human diploid
fibroblast (HDF) and the normal human liver L-02 cells were
cultured in RPMI-1640 medium. MCF-7 breast cancer cells, Hep-2
laryngeal epidermoid carcinoma, HepG2 liver carcinoma, A549 lung
adenocarcinoma, HeLa cervical carcinoma and SW-620 colon carcinoma
cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM).
The normal human fibroblastic cell line (MRC-5) was cultured in
Minimum Essential Medium (MEM). Cells were maintained at 37°C in a
5% CO2 atmosphere. All the media were supplemented with
10% heat-inactivated fetal bovine serum (FBS) supplemented with 50
U/ml penicillin and streptomycin.
 | Table IIC50 values of SNX-7081
and SNX-2112. |
Table I
IC50 values of SNX-7081
and SNX-2112.
| Drug:
IC50 (μM) |
|---|
|
|
|---|
| Cell lines | SNX-7081 | SNX-2112 |
|---|
| Human cancer
cells |
| K562
(leukemia) | <0.01 | <0.01 |
| A375
(melanoma) |
0.26±0.12a | 0.55±0.22 |
| MCF-7 (breast
cancer) | 1.09±0.74 | 2.58±1.30 |
| Hep-2 (laryngeal
cancer) |
1.13±0.64a | 3.58±0.95 |
| HepG2 (liver
cancer) | 1.29±0.51 | 2.52±1.35 |
| A549 (lung
cancer) |
3.61±1.50a | 5.86±2.24 |
| SW-620 (colon
carcinoma) | 0.05±0.03 | <0.01 |
| HeLa (cervical
cancer) | 9.16±4.50 | 6.31±3.34 |
| Normal human
cells |
| L-02 (liver
cells) | >100.00 | >100.00 |
| HDF (dermal
fibroblasts) | >100.00 | >100.00 |
| MRC5 (fetal lung
fibroblasts) | 70.16±27.85 | >100.00 |
MTT assay
Exponentially growing cells were seeded in 96-well
culture plates (5–10×103 cells/well) and allowed to
adhere overnight. Cancer cells were incubated with SNX-7081 or
SNX-2112 at various concentrations (0–10 μM) for 72 h. HDF, MRC5
and L-02 normal human cells were incubated with drugs at various
concentrations (0–50 μM) for 72 h, along with an equal volume of
DMSO as the solvent control. MTT solution (10 μl) was added to each
well (0.5 mg/ml) for an additional 4-h incubation (37°C, 5%
CO2). The precipitated formazan was dissolved in 100 μl
of DMSO. A 96-well multi-scanner autoreader (M450; Bio-Rad
Laboratories, Hercules, CA, USA) was used to measure the absorbance
of each well at 570 nm, with a reference wavelength of 630 nm. The
IC50 values, defined as the drug concentration that
caused 50% inhibition of absorbance compared with the control cells
treated with DMSO only, were calculated using the PrismPad
program.
Cell cycle analysis
Cell cycle distribution was determined by DNA
staining with propidium iodide (PI) (13). Briefly, cancer cell lines were
cultured and treated in 6-well culture plates (6×105
cells/well) with SNX-7081 or SNX-2112 (1.0 μM) for 48 h. The cells
were then washed in phosphate-buffered saline (PBS) and fixed in
70% ethanol overnight. The cells were collected and resuspended in
PBS containing 50 μg/ml PI, 0.1 mg/ml RNase, and 5% Triton X-100
and incubated at 37°C for 30 min. Subsequently, the cells were
analyzed on a flow cytometer (Becton-Dickinson, San Jose, CA, USA),
and the percentages of cells present in different phases of the
cell cycle were analyzed using the CellQuest software
(Becton-Dickinson).
Apoptosis assay
Apoptosis was measured by flow cytometry after
staining with Annexin V-FITC and PI according to the instructions
of the Annexin V-FITC/PI staining kit (Nanjing KeyGen Biotech.,
Co., Ltd., Nanjing, China). Briefly, cancer cells were cultured in
the presence of the indicated concentrations of SNX-7081 or
SNX-2112 (1.0 μM) for 48 h, harvested, washed twice and resuspended
in 500 μl of 1X binding buffer containing Annexin V-FITC and PI.
Samples were incubated at room temperature for 10 min and analyzed
by FACS.
Immunoblotting
K562 cells were incubated with 1.0 μM SNX-7081 or
SNX-2112 for 0, 6, 12, 24 or 48 h. Whole-cell lysates were prepared
by washing the cells with PBS and subjecting them to lysis with
RIPA buffer for 30 min on ice. Total protein concentrations of
whole-cell lysates were determined using the BCA protein assay kit.
Equal amounts of protein samples were loaded onto 8–12% sodium
dodecyl sulfate (SDS) polyacrylamide gel electrophoresis (PAGE)
gels. After electrophoresis, the proteins were transferred to
polyvinylidene fluoride (PVDF) membranes (Millipore), examined with
primary antibodies and then incubated with horseradish peroxidase
(HRP)-conjugated secondary antibodies. Specific protein bands were
visualized using the chemiluminescence method and imaged by
autoradiography. Any differences in protein loading were normalized
to the corresponding levels of the GAPDH control.
Docking assay
The affinity of SNX-7081 and SNX-2112 against Hsp90
was determined by the MOE docking assay. The crystal structure of
Hsp90 was taken from the Protein Data Bank (PDB code: 3R92). The
two compounds were converted to 3D structures, and energy was
minimized in MOE. The binding site of Hsp90 was minimized using the
AMBER 99 force field in MOE with the default parameter. The two
compounds were docked, employing Triangle Matcher as the placement
method and the function London dG as the first scoring function.
The refinement was set to force field (AMBER 99), and the docked
poses were energy minimized in the receptor pocket (24). The conformations of lowest energy
were given, and a lower scoring value indicated a more favorable
binding.
Statistical analysis
Data were evaluated by the Welch t-test when only
two value sets were compared. One-way ANOVA followed by the
Dunnett’s test was used for more group comparisons if ≥3
experiments were involved. Results were expressed as means ± SD
with a significance at *P<0.05 or
**P<0.01.
Results
Anti-proliferative effects of SNX-7081
and SNX-2112 on hunman cancer cells
The growth inhibitory effects of SNX-7081 and
SNX-2112 on human cancer lines originating from bone marrow, colon,
skin, larynx, breast, liver, cervix and lung were comopared. Cells
were exposed to various concentrations (0–10 μM) for 72 h and cell
viability was quantified by the MTT assay. SNX-7081 and SNX-2112
significantly inhibited the growth of eight human cancer cells in a
dose-dependent manner. The cell viability of SNX-7081 was
significantly lower than that of SNX-2112 at the low concentration
(<1 μM) in K562, A375, MCF-7, Hep-2, HepG2 and A549 cells
(Fig. 1A). In addition, SNX-7081
exhibited similar or weaker effects to SNX-2112 in SW-620 and HeLa
cells (Fig. 1B). The
IC50 values of SNX-7081 and SNX-2112 for these cells
were always ~1 μM (Table I). For
the remaining experiments, 1 μM was applied as the optimal
concentration.
Cytotoxic effects of SNX-7081 and
SNX-2112 on human normal cells
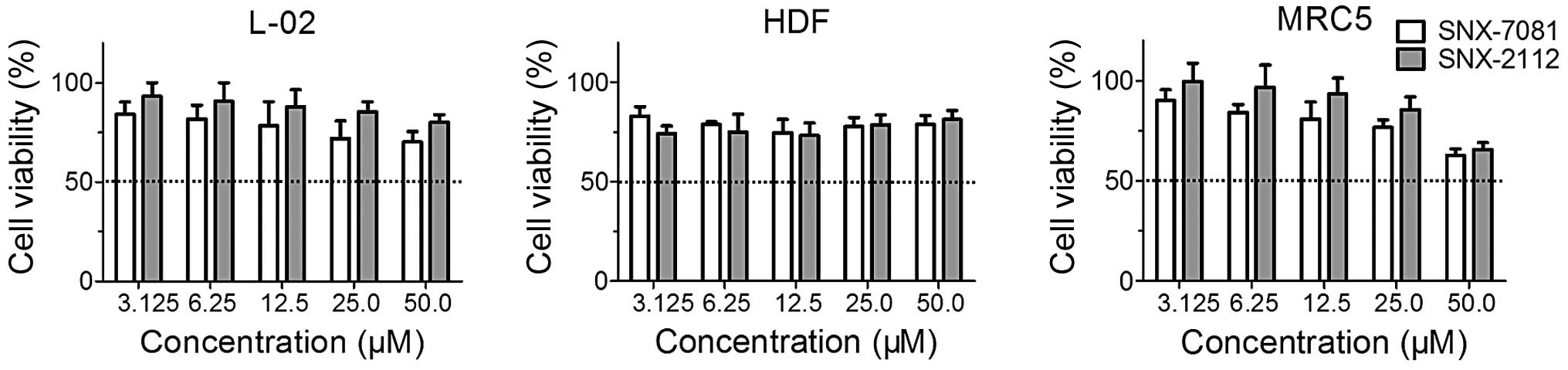
The cytotoxic effects of two agents on three human
normal cell lines (L-02, HDF and MRC5 cells) were determined by the
MTT assay. The cells were exposed to SNX-7081 or SNX-2112 at
concentrations of 3.13–50.0 μM for 72 h. The IC50 value
could not be determined even at the high concentration of 50.0 μM
(Table I and Fig. 2), suggesting that SNX-7081
cytotoxicity was lower as compared to SNX-2112 in L-02, MRC5 and
HDF cells. Therefore, SNX-7081 and SNX-2112 were provided with
acceptable cytotoxicity toward normal cells and high selectivity in
cancer cells.
Induction of cell cycle arrest at G2/M
phase by SNX-7081 and SNX-2112
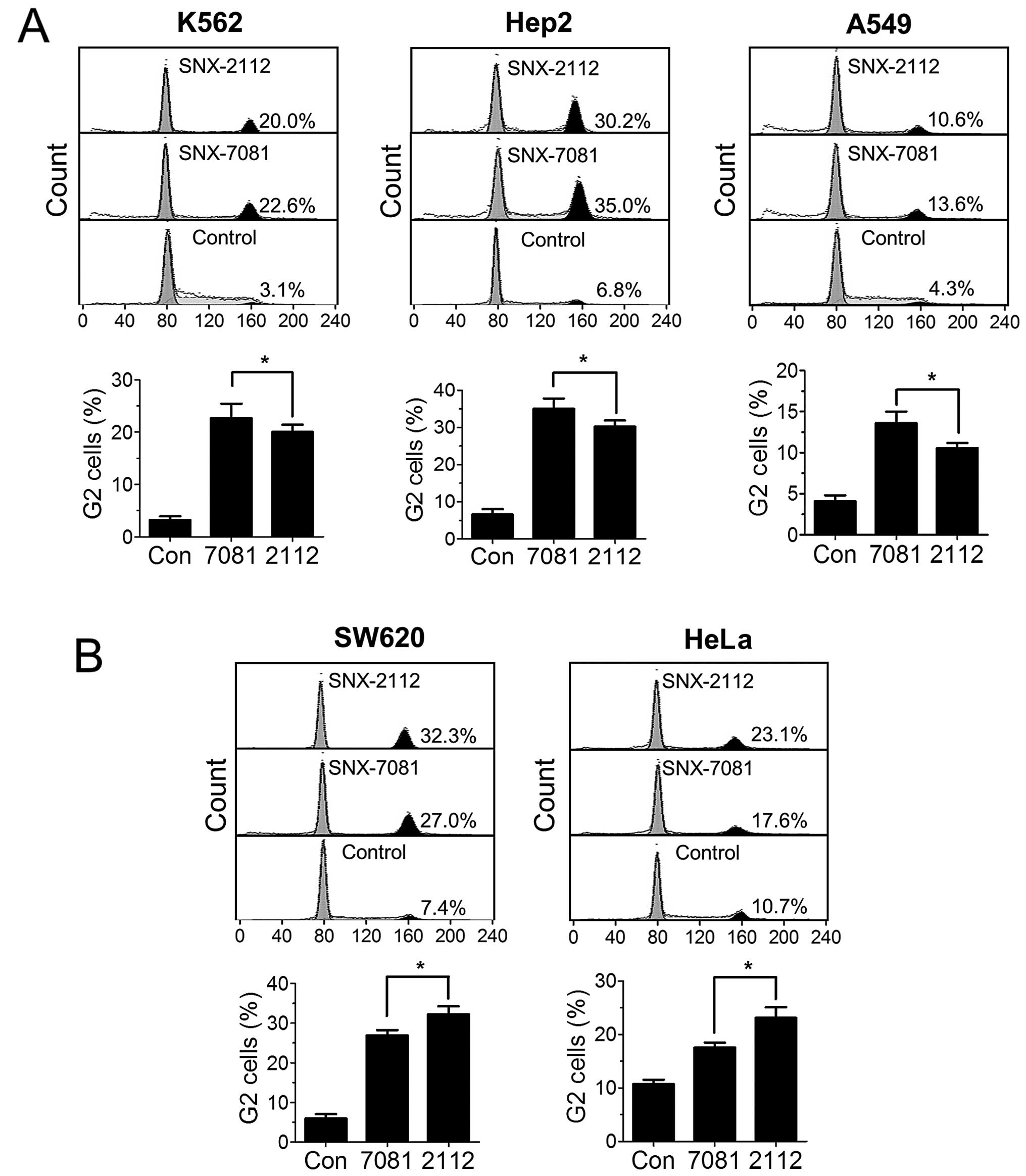
To explore the anti-proliferative mechanism of
SNX-7081 and SNX-2112, we assessed their effects on the cell cycle
distribution in the K562, Hep-2, A549, SW620 and HeLa cancer cell
lines using flow cytometry. Cells treated with SNX-7081 and
SNX-2112 (1 μM) for 48 h, were subjected to flow cytometric
analysis after PI staining. SNX-7081 more effectively increase the
cell amount of G2/M phase in K562, Hep-2 and A549 cells (Fig. 3A), while SNX-2112 induces a more
significant G2/M arrest in SW-620 and HeLa cells (Fig. 3B). The finding suggested that
SNX-7081-induced inhibitory effects may be mediated by cell cycle
arrest.
Effects of SNX-7081 and SNX-2112 on
cancer cell apoptosis
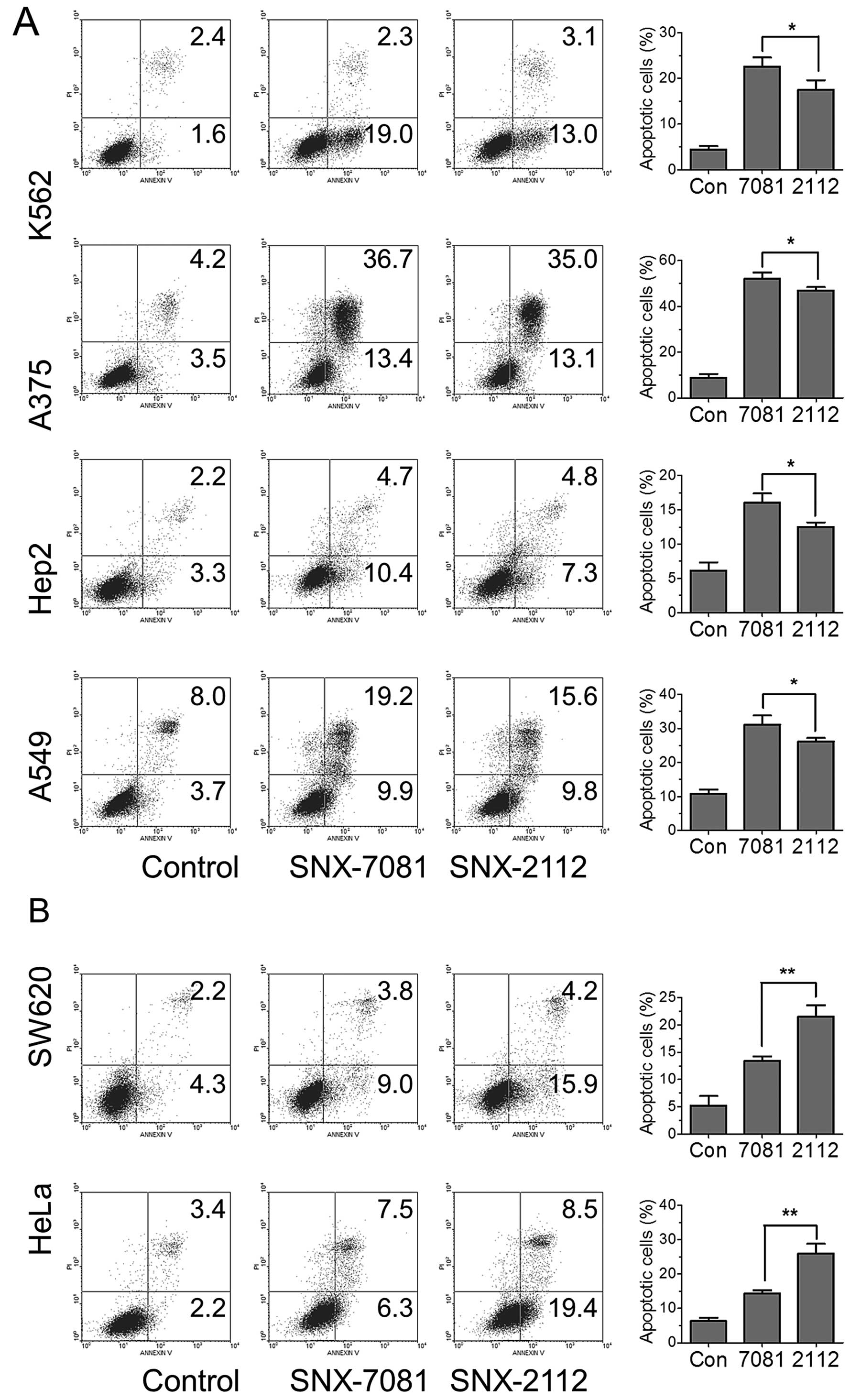
To examine whether apoptosis is involved in the
inhibition of cancer cell proliferation by SNX-2112 or SNX-7081,
cancer cells were treated with 1 μM SNX-2112 or SNX-7081 for 48 h
and subjected to Annexin V- and PI-double staining for flow
cytometric analysis. The degree of apoptosis was calculated as the
sum of the percentages of cells in the lower right and upper right
quadrants. SNX-7081 and SNX-2112 clearly induced apoptosis in all
six cell lines. SNX-7081 induced more apoptosis in K562, Hep-2 and
A549 cells (Fig. 4A), but less
apoptosis in SW-620 and HeLa cells as compared to that of SNX-2112
(Fig. 4B). The data indicated that
SNX-7801-induced apoptosis in some cancer cells was more potent
than SNX-2112.
Effects of SNX-7081 and SNX-2112 on the
expression of Hsp90 client proteins
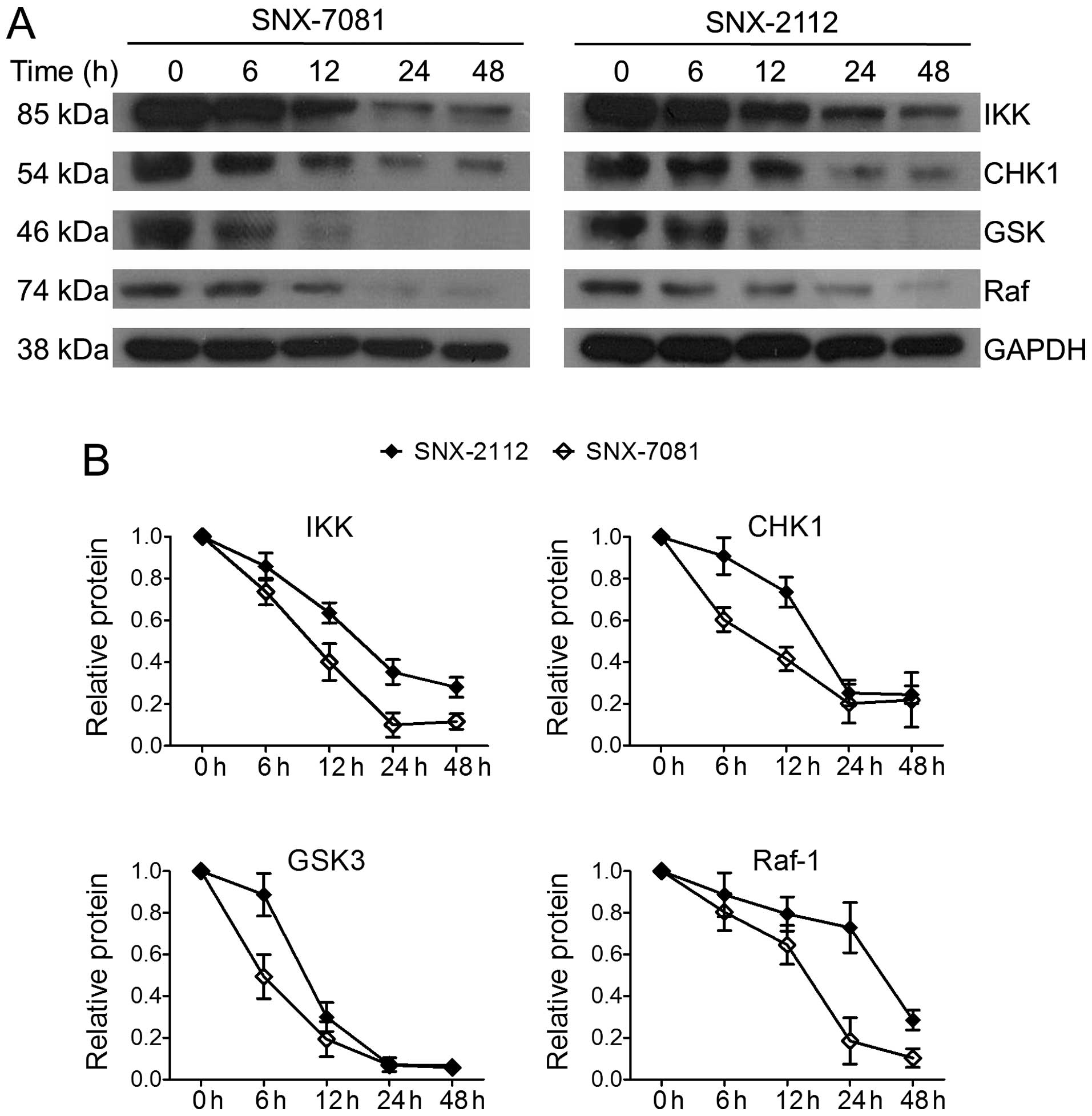
The basic mechanism of Hsp90 inhibitors involves
binding to the ATP pocket and suppressing the ATPase activity of
Hsp90, resulting in the degradation of its client proteins
(25). It was previously
demonstrated that SNX-2112 led to downregulation of the Hsp90
client proteins Bcr-abl and Akt in K562 cells (13). In the present study, we compared the
effect of SNX-7081 and SNX-2112 on the expression of other
essential Hsp90 client proteins, including IKKα, GSK3, CHK1 and
Raf-1, which are crucial for the growth of cancer cells. Western
blot analysis showed that the levels of these proteins were reduced
in a time-dependent manner following exposure to the compounds
(Fig. 5). After treatment with 1 μM
of SNX-7081 for 48 h, the expression of IKKα, CHK1, GSK3 and Raf-1
was significantly decreased to 11.7, 21.8, 5.8 and 10.4%,
respectively. SNX-7081 exerted stronger inhibitory effects on the
expression levels of Hsp90 client proteins than SNX-7081. These
data support the principle that SNX-7081 and SNX-2112 act via the
inhibition of Hsp90 chaperone function, and the SNX-7081 was more
potent in suppressing Hsp90 client proteins.
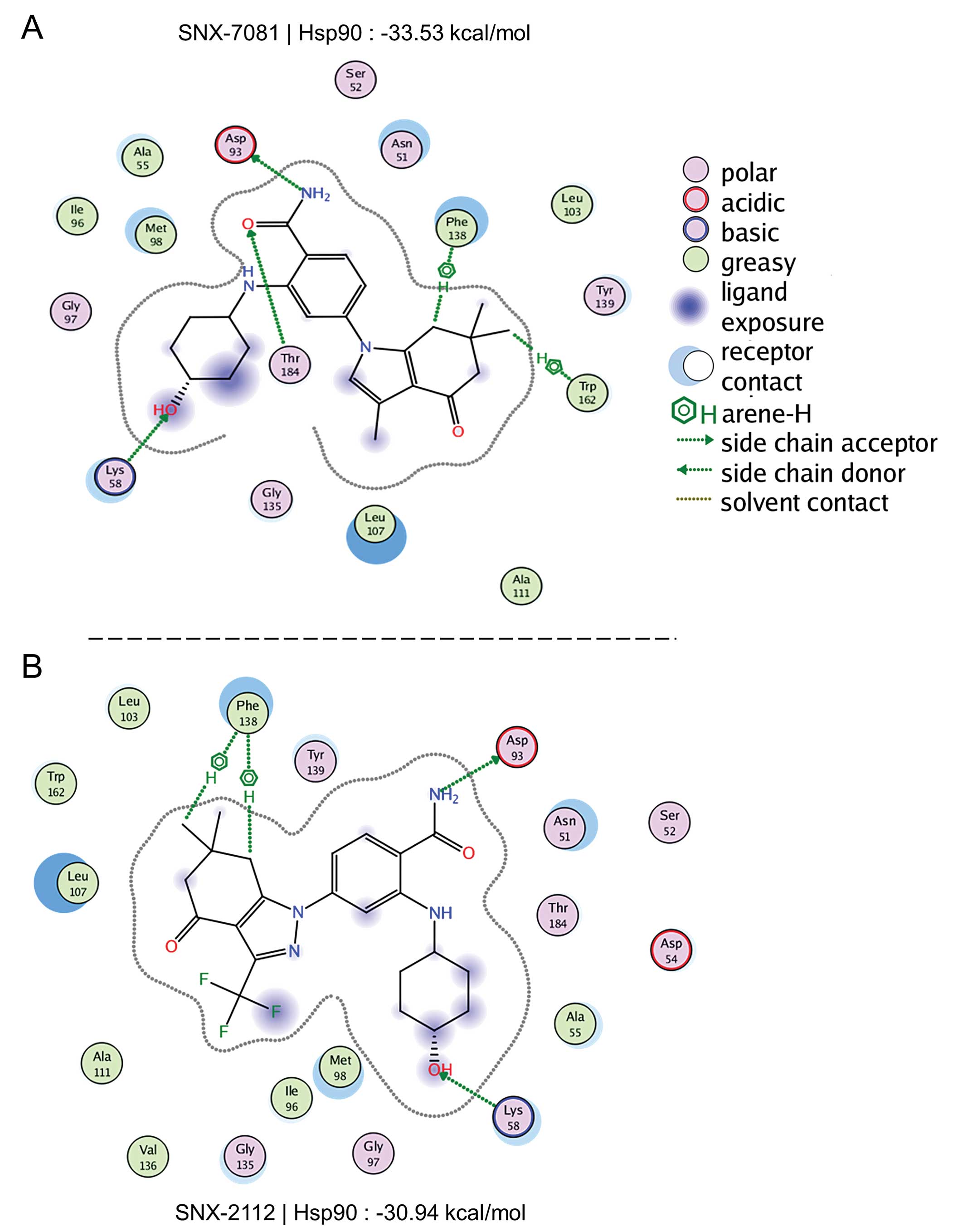
Binding affinity of SNX-7081 and SNX-2112
to Hsp90
To determine the binding affinity of SNX-7081 and
SNX-2112 to Hsp90, the inhibitors were docked into the ATP site of
Hsp90, using MOE. A lower scoring value suggests a more favorable
binding. The ligand-receptor interaction maps for SNX-7081 and
SNX-2112 are shown in Fig. 6. We
found two hydrogen bond residues (Phe-138 and Trp-163), two side
chain donor molecules (Lys-58 and Thr-184), and one side chain
acceptor molecule (Asp-93) that bound to SNX-7081. SNX-7081
interacted with five residues in the pocket, while SNX-2112 only
interacted with three residues. The scoring value of SNX-7081 was
−33.53 kcal/mol, which was lower than the −30.94 kcal/mol on
SNX-2112. It was evident that SNX-7081 was the highly-scored
compound, suggesting that SNX-7081 had a stronger binding affinity
to Hsp90 than SNX-2112.
Discussion
The discontinuity in clinical trials of SNX-2112
results in opportunities and challenges for other 2-aminobenzamide
inhibitors of Hsp90. In the present study, we found that SNX-7081,
an analogue of SNX-2112, exhibited higher affinity for Hsp90 than
SNX-2112 in molecular docking experiments. The structure of
SNX-7081 is different from SNX-2112, with a side chain of indole
instead of indazole. In addition, the methyl substituent in the
indole ring instead the of 5-fluoro substituent in indazole may
also contribute to the higher affinity of SNX-7081. A similar
binding model suggested that SNX-7081 may be a feasible alternative
of SNX-2112, leading to a comparison of the anticancer activity of
the two compounds.
SNX-7081 had better activity than SNX-2112 in most
cancer cells. One interpretation of this finding is that the
expressions and functions of various Hsp90 isoforms are not uniform
in different cell types. These Hsp90 isoforms include Hsp90α and
Hsp90β in the cytoplasm and nucleus, GRP94 in the endoplasmic
reticulum (ER) and TRAP1 in the mitochondria (25). The functions of Hsp90α include
stress-induced cytoprotection and cell cycle regulation, whereas
Hsp90β plays an important role in early embryonic development,
signal transduction and long-term cell adaptation (26,27).
GRP94 is specialized for protein folding and plays a role in the
degradation of misfolded proteins through the ER-associated
degradation pathway (28).
Additionally, TRAP1 is involved in signal transduction, protein
folding, protein degradation and morphologic evolution (29). Thus, the development of Hsp90
inhibitors against specific Hsp90 isoforms is a promising strategy
for cancer therapy.
In addition, SNX-7081 and SNX-2112 exhibited strong
selectivity in cell growth inhibition between cancer cells and
normal cells. This is probably because the expression and molecular
conformation of Hsp90 are greatly different in cancer and normal
cells. In tumor cells, Hsp90 may be exclusively complexed with
co-chaperones, forming a super-chaperone complex in a state of high
affinity for ATP/ADP or ligands (e.g., Hsp90 inhibitors) of this
regulatory pocket. However, in normal cells, Hsp90 is predominantly
in a latent, uncomplexed and low affinity state (30,31).
Therefore, compared with Hsp90 in normal cells, tumor Hsp90 is
hypersensitive to Hsp90 inhibitors.
Results of this study show that SNX-7081 exhibits an
anti-tumor profile of the natural product Hsp90 inhibitors: growth
inhibition, G2 cell cycle arrest, cell apoptosis induction and
degradation of Hsp90 clients (32).
Previously, it was found that SNX-2112 can induce the degradation
of Bcr-abl and Akt in K562 cells (21). Although there are many other Hsp90
client proteins, the focus of the present study was on the levels
of IKKα, GSK3, CHK1 and Raf-1, due to their critical roles in the
proliferation of human leukemia cells. For instance, IKKα is an
NF-κB inhibitory protein that contributes to the de-regulated
expression of various cellular genes in T-cell leukemia cells
(33,34). GSK3 controls cell survival, and its
inactivation induces cell apoptosis in leukemia cells (35,36).
Inhibition of CHK1 kinase has been shown to induce cell cycle
arrest in K562 cells (37), while
targeting Raf-1 gene expression by a DNA enzyme inhibits the growth
of leukemia cells (38). Therefore,
the downregulation of Hsp90 client proteins in the upstream pathway
may be the mechanism of the induction of growth inhibition, cell
cycle arrest and apoptosis pertaining to SNX-7081 in cancer
cells.
In conclusion, we have demonstrated that SNX-7081
was significantly more potent than SNX-2112 in a majority of human
cancer cells. Consequently, this superiority effect merits further
confirmation in xenograft experiments in vivo. The mode of
action of antitumor activity may be associated with the induction
of cell cycle arrest, apoptosis and Hsp90 client proteins
degradation. Our results suggest that the novel Hsp90 inhibitor
SNX-7081 is a promising alternative to SNX-2112, although more
fundamental investigations are required to confirm the results.
Acknowledgements
We would like to thank Professor Bao-Jian Wu from
Jinan University (Guangzhou, China) for proofreading the
manuscript. The present study was supported by grants from the
National Natural Science Foundation of China (grant no. 81201727),
the China Postdoctoral Science Foundation (grant nos. 2012M511882
and 2013T60827), the open project of State Key Laboratory of
Molecular Oncology (SKL-KF-2013–14), the Guangdong Province and
Ministry of Education Ministry of Science and Technology Products
Research Combined Platform Project (grant no. 2010B091000013), and
the Natural Science Foundation of Guangdong Province (grant no.
S2012040006873).
References
|
1
|
Taipale M, Jarosz DF and Lindquist S:
HSP90 at the hub of protein homeostasis: emerging mechanistic
insights. Nat Rev Mol Cell Biol. 11:515–528. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Trepel J, Mollapour M, Giaccone G and
Neckers L: Targeting the dynamic HSP90 complex in cancer. Nat Rev
Cancer. 10:537–549. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: the next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mahalingam D, Swords R, Carew JS, Nawrocki
ST, Bhalla K and Giles FJ: Targeting HSP90 for cancer therapy. Br J
Cancer. 100:1523–1529. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Banerji U, O’Donnell A, Scurr M, et al:
Phase I pharmacokinetic and pharmacodynamic study of 17-allylamino,
17-demethoxy-geldanamycin in patients with advanced malignancies. J
Clin Oncol. 23:4152–4161. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Taldone T, Gozman A, Maharaj R and Chiosis
G: Targeting Hsp90: small-molecule inhibitors and their clinical
development. Curr Opin Pharmacol. 8:370–374. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Goetz MP, Toft D, Reid J, et al: Phase I
trial of 17-allylamino-17-demethoxygeldanamycin in patients with
advanced cancer. J Clin Oncol. 23:1078–1087. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chinn DC, Holland WS, Yoon JM, Zwerdling T
and Mack PC: Anti-tumor activity of the HSP90 inhibitor SNX-2112 in
pediatric cancer cell lines. Pediatr Blood Cancer. 58:885–890.
2012. View Article : Google Scholar
|
|
9
|
Wang R, Shao F, Liu Z, et al: The Hsp90
inhibitor SNX-2112, induces apoptosis in multidrug resistant
K562/ADR cells through suppression of Akt/NF-kappaB and disruption
of mitochondria-dependent pathways. Chem Biol Interact. 205:1–10.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang SX, Ju HQ, Liu KS, et al: SNX-2112, a
novel Hsp90 inhibitor, induces G2/M cell cycle arrest and apoptosis
in MCF-7 cells. Biosci Biotechnol Biochem. 75:1540–1545. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bachleitner-Hofmann T, Sun MY, Chen CT, et
al: Antitumor activity of SNX-2112, a synthetic heat shock
protein-90 inhibitor, in MET-amplified tumor cells with or without
resistance to selective MET inhibition. Clin Cancer Res.
17:122–133. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu KS, Liu H, Qi JH, et al: SNX-2112, an
Hsp90 inhibitor, induces apoptosis and autophagy via degradation of
Hsp90 client proteins in human melanoma A-375 cells. Cancer Lett.
318:180–188. 2012. View Article : Google Scholar
|
|
13
|
Crissman HA and Steinkamp JA: Rapid,
simultaneous measurement of DNA, protein, and cell volume in single
cells from large mammalian cell populations. J Cell Biol.
59:766–771. 1973. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rajan A, Kelly RJ, Trepel JB, et al: A
phase I study of PF-04929113 (SNX-5422), an orally bioavailable
heat shock protein 90 inhibitor, in patients with refractory solid
tumor malignancies and lymphomas. Clin Cancer Res. 17:6831–6839.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Huang KH, Veal JM, Fadden RP, et al:
Discovery of novel 2-aminobenzamide inhibitors of heat shock
protein 90 as potent, selective and orally active antitumor agents.
J Med Chem. 52:4288–4305. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Best OG, Che Y, Singh N, Forsyth C,
Christopherson RI and Mulligan SP: The Hsp90 inhibitor SNX-7081
synergizes with and restores sensitivity to fludarabine in chronic
lymphocytic leukemia cells with lesions in the TP53 pathway: a
potential treatment strategy for fludarabine refractory disease.
Leuk Lymphoma. 53:1367–1375. 2012. View Article : Google Scholar
|
|
17
|
Che Y, Best OG, Zhong L, et al: Hsp90
inhibitor SNX-7081 dysregulates proteins involved with DNA repair
and replication and the cell cycle in human chronic lymphocytic
leukemia (CLL) cells. J Proteome Res. 12:1710–1722. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rice JW, Veal JM, Fadden RP, et al: Small
molecule inhibitors of Hsp90 potently affect inflammatory disease
pathways and exhibit activity in models of rheumatoid arthritis.
Arthritis Rheum. 58:3765–3775. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Best OG, Singh N, Forsyth C and Mulligan
SP: The novel Hsp-90 inhibitor SNX7081 is significantly more potent
than 17-AAG against primary CLL cells and a range of haematological
cell lines, irrespective of lesions in the TP53 pathway. Br J
Haematol. 151:185–188. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Barta TE, Veal JM, Rice JW, et al:
Discovery of benzamide tetrahydro-4H-carbazol-4-ones as novel small
molecule inhibitors of Hsp90. Bioorg Med Chem Lett. 18:3517–3521.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jin L, Xiao CL, Lu CH, et al:
Transcriptomic and proteomic approach to studying SNX-2112-induced
K562 cells apoptosis and anti-leukemia activity in K562-NOD/SCID
mice. FEBS Lett. 583:1859–1866. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee YH, Lee NH, Bhattarai G, et al:
PPARgamma inhibits inflammatory reaction in oxidative stress
induced human diploid fibloblast. Cell Biochem Funct. 28:490–496.
2010. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bettger WJ, Boyce ST, Walthall BJ and Ham
RG: Rapid clonal growth and serial passage of human diploid
fibroblasts in a lipid-enriched synthetic medium supplemented with
epidermal growth factor, insulin, and dexamethasone. Proc Natl Acad
Sci USA. 78:5588–5592. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang D, Gu Q, Ge H, et al: On the value
of homology models for virtual screening: discovering hCXCR3
antagonists by pharmacophore-based and structure-based approaches.
J Chem Inf Model. 52:1356–1366. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kawano T, Kobayakawa T, Fukuma Y, et al: A
comprehensive study on the immunological reactivity of the Hsp90
molecular chaperone. J Biochem. 136:711–722. 2004. View Article : Google Scholar
|
|
26
|
Sreedhar AS, Kalmar E, Csermely P and Shen
YF: Hsp90 isoforms: functions, expression and clinical importance.
FEBS Lett. 562:11–15. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hong DS, Banerji U, Tavana B, George GC,
Aaron J and Kurzrock R: Targeting the molecular chaperone heat
shock protein 90 (HSP90): lessons learned and future directions.
Cancer Treat Rev. 39:375–387. 2013. View Article : Google Scholar
|
|
28
|
Marzec M, Eletto D and Argon Y: GRP94: an
HSP90-like protein specialized for protein folding and quality
control in the endoplasmic reticulum. Biochim Biophys Acta.
1823:774–787. 2012. View Article : Google Scholar :
|
|
29
|
Matassa DS, Amoroso MR, Maddalena F,
Landriscina M and Esposito F: New insights into TRAP1 pathway. Am J
Cancer Res. 2:235–248. 2012.
|
|
30
|
Kamal A, Thao L, Sensintaffar J, et al: A
high-affinity conformation of Hsp90 confers tumour selectivity on
Hsp90 inhibitors. Nature. 425:407–410. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vilenchik M, Solit D, Basso A, et al:
Targeting wide-range oncogenic transformation via PU24FCl, a
specific inhibitor of tumor Hsp90. Chem Biol. 11:787–797. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Miyata Y: Hsp90 inhibitor geldanamycin and
its derivatives as novel cancer chemotherapeutic agents. Curr Pharm
Des. 11:1131–1138. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Harhaj EW and Sun SC: IKKgamma serves as a
docking subunit of the IkappaB kinase (IKK) and mediates
interaction of IKK with the human T-cell leukemia virus Tax
protein. J Biol Chem. 274:22911–22914. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mori N, Yamada Y, Ikeda S, et al: Bay
11–7082 inhibits transcription factor NF-kappaB and induces
apoptosis of HTLV-I-infected T-cell lines and primary adult T-cell
leukemia cells. Blood. 100:1828–1834. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhou F, Zhang L, van Laar T, van Dam H and
Ten Dijke P: GSK3β inactivation induces apoptosis of leukemia cells
by repressing the function of c-Myb. Mol Biol Cell. 22:3533–3540.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jin ZH, Kurosu T, Yamaguchi M, Arai A and
Miura O: Hematopoietic cytokines enhance Chk1-dependent G2/M
checkpoint activation by etoposide through the Akt/GSK3 pathway to
inhibit apoptosis. Oncogene. 24:1973–1981. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jia XZ, Yang SY, Zhou J, et al: Inhibition
of CHK1 kinase by Go6976 converts 8-chloro-adenosine-induced G2/M
arrest into S arrest in human myelocytic leukemia K562 cells.
Biochem Pharmacol. 77:770–780. 2009. View Article : Google Scholar
|
|
38
|
Davis JM, Navolanic PM,
Weinstein-Oppenheimer CR, et al: Raf-1 and Bcl-2 induce distinct
and common pathways that contribute to breast cancer drug
resistance. Clin Cancer Res. 9:1161–1170. 2003.PubMed/NCBI
|