Introduction
Hepatocellular carcinoma (HCC) is the sixth most
common cancer and the third most common cause of mortality from
cancer worldwide (1). Cisplatin
(DDP) has broad-spectrum antitumor effects used in the chemotherapy
of different malignancies including HCC. However, serious
side-effects were observed with high doses and multiple courses of
treatment (2,3). Nephrotoxicity is frequent and remains
the major limitation in cisplatin-based chemotherapy (4). Previous findings suggested that
cisplatin injury causes endothelial cell dysfunction and
infiltration of circulating leukocytes, particularly neutrophils
(4,5), leading to the excretion of locally
secreted cytokines/chemokines such as tumor necrosis factor (TNF),
which contributes to the initiation and progression of renal
disease (6). However, clinically
alternative strategies such as immunotherapy and their combinations
with chemotherapy have been suggested to reduce the side effects.
Inhibition of proinflammatory mediators in DDP-induced ARF is
associated with reduction in renal neutrophils (7).
The expression of interleukin (IL)-8, an ELR-CXC
chemokine, is involved as a chemotactic attractant of neutrophils,
T cells and other inflammatory cells by binding to its receptors
CXCR1 and CXCR2 (8), found at the
inflammation site of various types of human cancer, including HCC,
and is regulated by many tumor microenvironmental factors, such as
hypoxia and tumor necrosis factor (9).
CXCL8(3-72)K11R/G31P (G31P) is a
recombinant compound produced from human IL-8 using gene
site-directed mutagenesis by introducing a Lys 11 to Arg
substitution and a Gly 31 to Pro substitution to generate a broad
spectrum ELR-CXC chemokine antagonist, human
CXCL8(3-72)K11R/G31P (10,11).
G31P has been shown to bind to CXCR1/2, block signal transduction
and is considered as anti-inflammatory preparation with ‘broad
spectrum’ anti-inflammatory activity (12–14).
G31P exerts inhibitory effects on the growth, metastasis and
angiogenesis of malignant tumors such as prostate cancer and NSCLC
(15). G31P was also previously
found to play an anti-atherosclerosis role (16).
In the present study, the antitumor effects of DDP
and G31P on mouse HCC in vitro and in vivo were
investigated, as well as the effect of high-dose DDP-induced acute
renal failure. The combine use of DDP and G31P resulted in the
inhibition of neutrophil infiltration, cytokine release and
amelioration of ARF.
Materials and methods
Cell lines, animals and reagents
Female Balb/c mice (18–22 g) were provided by the
SPF Animal Center, Dalian Medical University (Dalian, China). H22
mouse HCC cells were cultured in ascites fluid, and after 7 days,
second generation mice were used in experiments. DDP was purchased
from Haosheng Pharmaceutical Corporation (Lianyungang, China). G31P
was produced by the Institute of Biotechnology, National Tsing Hua
University (Hsinchu, Taiwan).
MTT assay
The second generation H22 mouse HCC cells were
collected and maintained in a complete RPMI-1640 medium containing
10% fetal bovine serum (HyClone, Australia). The cells were plated
in 96-well plates (2×104/ml cells/well in 100 μl of
medium) and maintained in a humidified incubator with 5%
CO2 at 37°C. After 12 h of incubation, the cells were
treated with or without DDP (at a doses of 0.25, 5 and 10 μg/ml)
with or without G31P (0.1 μg/ml). After 24 h, 10 μl of MTT (5
mg/ml) was added to each well and incubated for 4 h. The reaction
was stopped by adding 100 μl of dimethyl sulfoxide. Absorbance was
measured at 570 nm and the cell proliferation inhibition rate was
calculated.
DDP tolerance test in mice
To determine the maximum survival dose of DDP,
Balb/c mice were randomly divided into 5 groups on the basis of DDP
dosage. Each group (n=6) was treated with different concentrations
of single-dose DDP starting from 11, 12.5, 14 and 15.5 mg/kg
intraperitoneally (i.p.), and the control group was injected with
same volume of saline.
Establishment of animal model
H22 mouse HCC cells (1×106) in 0.2 ml
suspension were injected subcutaneously into the posterior right
flank of mice for tumor implantation. After 7 days, the mice were
randomly divided into four groups: DDP (n=12), DDP+G31P (n=12),
G31P (n=20) and the control group (n=20). The DDP and DDP+G31P
groups were treated with a single dose of DDP (12.5 mg/kg i.p.).
Mice of the DDP+G31P and G31P groups were treated with G31P (500
μg/kg; subcutaneously) on alternative days, while other groups were
treated with isovolumic saline. Following treatment, the mice were
observed weight and survival rate was recorded. On the 9th day of
drug treatment, the mice in each group were sacrificed. Blood from
eye canthus vein from each mouse was aspirated to measure the serum
urea nitrogen (BUN) and creatinine (SCr) levels and tumor samples
from each mouse were weighed and preserved for subsequent analysis.
The tumor volume was calculated using the formula: Tumor volume =
(length × width2)/2.
Hematoxylin and eosin analysis
Kidney and tumor tissue samples from the DDP,
DDP+G31P, G31P and control groups were fixed in 4% paraformaldehyde
and embedded in paraffin using standard procedures. Sections (6 μm)
were cut and stained with hematoxylin and eosin (H&E). For the
kidney samples, tubular damage was estimated by examining 8–10
high-power fields (HPF; magnification ×200) per section by using a
scoring system based on the percentage of damaged tubules per
field: 1, <25%; 2, 25–50%; 3, 50–75%; and 4, >75%. The mean
score of each animal was compared.
Immunohistochemistry
Tumor samples were embedded in O.C.T. Compound and
cut into slices (8-μm) in cryostat (−22°C). The sections were
warmed at room temperature for 30 min, fixed in ice-cold acetone
for 10 min, and rinsed three times with phosphate-buffered saline
(PBS). The endogenous peroxidase activities were blocked by 3%
H2O2 for 10 min. Sections were then rinsed
three times with PBS, incubated with a protein blocker solution of
10% normal goat serum for 15 min at room temperature, and washed
three times with PBS. Sections were subsequently incubated
overnight at 4°C with primary antibodies: rabbit anti-VEGF-A
antibody (1:200), rabbit anti-NF-κB p65 antibody (1:200) and rabbit
anti-EGFR antibody (1:200) (all from Santa Cruz Biotechnology,
Inc., Santa Cruz, CA, USA). The following day, the slides were
rinsed three times with PBS and incubated in streptavidin
peroxidase-conjugated and biotinylated goat anti-rabbit IgG
(1:5,000; Zhongshan Biotech, Beijing, China) for 1 h at 37°C.
Subsequently, the slides were washed three times with PBS and
stained with 3,3′-diaminobenzidine (DAB) (Zhongshan Biotech) for 6
min and clear DAB in running tape water for 5 min. The sections
were stained with hematoxylin staining solution for 30 sec and
rinsed under running water. Dehydration was performed following
standard procedure in diluted alcohol solutions and the sections
were sealed with neutral resins.
Western blot analysis
After protein extraction from tumor and kidney
tissue samples from DDP, DDP+G31P, G31P and control groups, each
protein sample was separated by 10% SDS-PAGE gel. After
electrophoresis, the proteins were transferred to PVDF membranes
(Amersham, Buckinghamshire, UK). The membranes were incubated with
blocking solution containing 0.3% Tween-20, and 10% skim-dried milk
in TBS for 2 h. The primary antibodies of EGFR (rabbit antibody,
1:200), MMP-2 (goat antibody, 1:200), VEGF-A (rabbit antibody
1:200) (all from Santa Cruz Biotechnology) were incubated with
proteins PVDF of tumor overnight at 4°C. The following day, the
primary antibody was removed by washing in TBST three times. The
membranes were incubated with horseradish peroxidase-conjugated
goat anti-rabbit (or rabbit anti-goat) secondary antibody (1:5,000;
Zhongshan Biotech) for 1 h at room temperature. Following three
washes in TBST, bands were visualized by electrochemical
luminescence (ECL). The amount of each protein was determined from
the intensity of the band. To verify equal protein loading, the
β-actin level in each lane was examined with anti-β-actin antibody.
Bands were semi-quantified with the software Image J2x.
Evaluation of BUN and SCr
Blood urea nitrogen (BUN) and SCr levels were
measured using an automatic biochemical analyzer according to the
manufacturer’s instructions.
Determination of MPO activity of
kidney
The activity of myeloperoxidase (MPO), an enzyme
present in neutrophils, was used as a marker for neutrophil
infiltration. Kidney tissues were homogenized in 10% (w/v) of 20 mM
phosphate buffer (pH 7.4). After centrifugation at 15,000 rpm for
20 min, the pellet was sonicated and resuspended in 2,350 mM
potassium phosphate buffer (pH 6.0) containing 0.5%
hexadecyltrimethylammonium bromide. After being heated at 60°C for
2 h, the samples were centrifuged at 10,000 rpm for 10 min. The
supernatant was added to 2,250 mM phosphate buffer (pH 6.0)
containing 0.167 mg/ml o-dianisidine and 5×10−4 %
hydrogen peroxidase. MPO activity was measured using s
spectrophotometer. The change in absorbance was measured at 460 nm
at 37°C, using a SpectraMax microplate reader (Molecular Devices,
Sunnyvale, CA, USA). Results were expressed as a unit of MPO
activity/g of wet tissue.
RT-PCR detection of inflammatory
cytokines in kidney
TRIzol reagent (Takara, Dalian, China) was used to
extract total RNA from the kidney samples of DDP, DDP+G31P, G31P
and the control group according to the manufacturer’s instructions.
Synthesis of complementary DNA (cDNA) was performed with reverse
transcriptase and oligo(dT) primer (Takara) as per instructions on
the product data sheet. PCR primers used were: β-actin, forward:
(5′-CCAGAGCAAGAGAGGCATCC-3′) and reverse:
(5′-CAACTGTCTCCATGTCGTCC-3′); MIP-2, forward:
(5′-AACATCCAGAGCTTGAGTGTGAC-3′) and reverse:
(5′-GCCTTGCCTTTGTTCAGTATCTT-3′); TNF-α, forward:
(5′-CAGCCGATTTGCTATCTCATACC-3′) and reverse:
(5′-GTACTTGGGCAGATTGACCTCAG-3′). PCR conditions were: 94°C for 2
min, followed by 30 cycles of 94°C for 30 sec, 58°C for 30 sec and
72°C for 1 min. The final extension was at 72°C for 10 min. PCR
products were analyzed on 1.2% (W/V) agarose gel containing 0.5
mg/ml ethidium bromide, visualized under UV light band density, and
analyzed and quantified with the software Image J2x.
Statistical analysis
Data are presented as the means ± standard deviation
(SD). The Student’s t-test and ANOVA were used to determine
significance. Statistical analysis was performed using SPSS 10.0
for windows software. Differences were considered significant at
P<0.05.
Results
Effect of DDP toxicity in vivo
Following the intraperitoneal injection of DDP,
reduced activity and food intake with sparse dull hair and weight
loss were observed for the mice. The 14 and 15.5 mg/kg body weight
DDP-treated mice were in a poor state: they huddled in a group with
retarded activity and began to die on the 2nd day of medication,
with no mice surviving after 4 days. However, the 12.5 mg/kg body
weight DDP-treated mice had maximum survival rate and were used in
subsequent experiments to observe the effects of G31P with a
tolerable high dose of DDP in H22 HCC mice.
G31P and DDP additively inhibit the
proliferation of H22 hepatocarcinoma cells
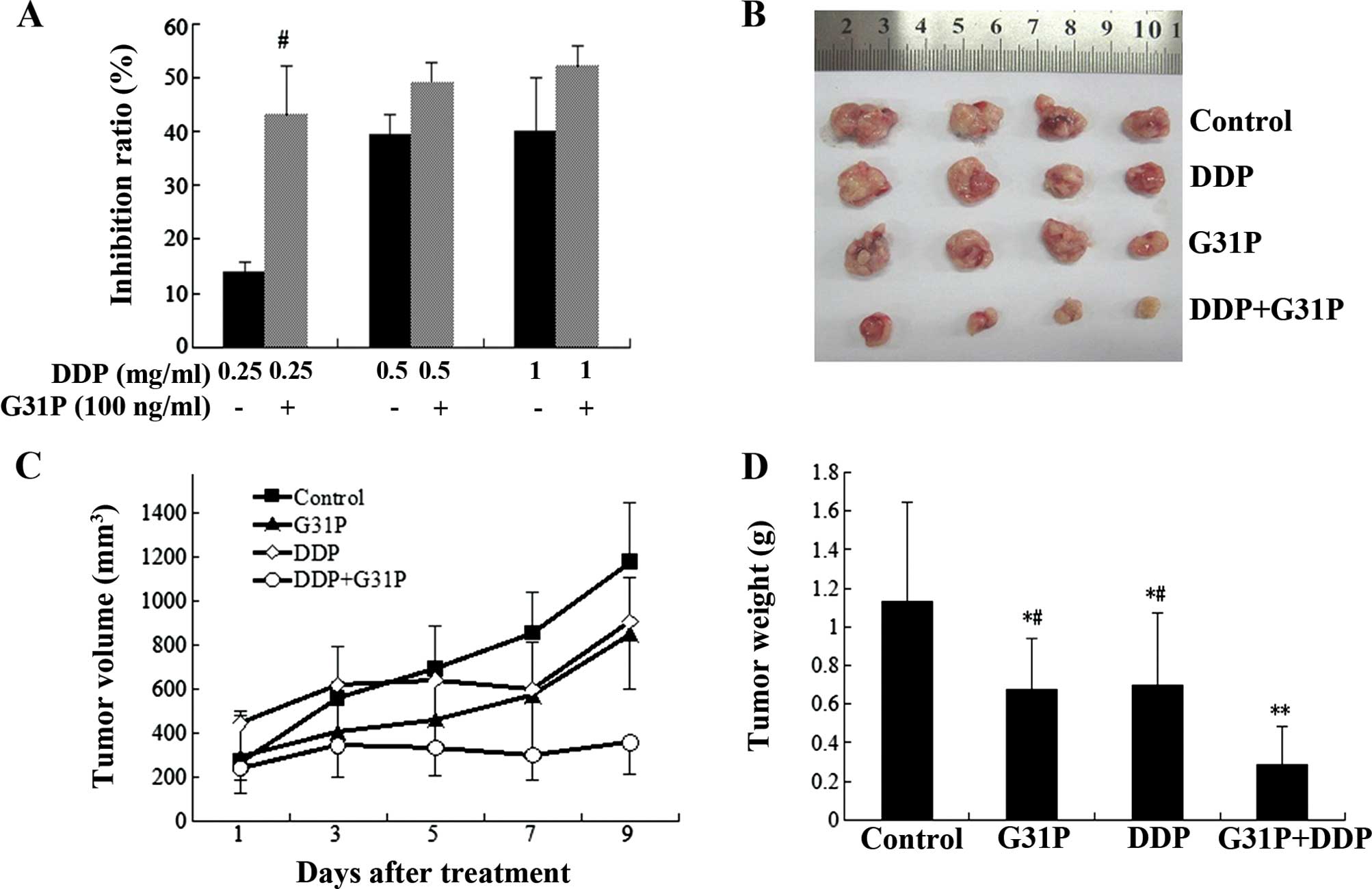
DDP reduced H22 viability in a dose-dependent manner
in vitro. The data also showed that the drug combination
scheme had more effective inhibitory effects on H22 cell viability
than DDP alone (p<0.05; Fig.
1).
To compare tumor growth in vivo among the
different groups, tumor volume and weight were measured. Control
group tumors continued to grow, as compared to those in the G31P
and DDP groups. Following treatment with DDP, tumors in the DDP
group did not grow for a short period, but started growing after 7
days, whereas, tumors in the DDP+G31P group grew extremely slowly
during drug treatment. Tumor mass in the DDP+G31P group was
significantly lower than that in the DDP and G31P groups
(P<0.05; Fig. 1) and the control
group with significant differences (P<0.01; Fig. 1). The inhibition rate of the DDP,
G31P and DDP+G31P groups was 38.40, 40.74 and 74.80%, respectively,
and the general state of mice in the DDP+G31P group was
significantly improved as compared to the DDP group and relatively
agile activity was observed.
Histopathological examination of tumor
tissues
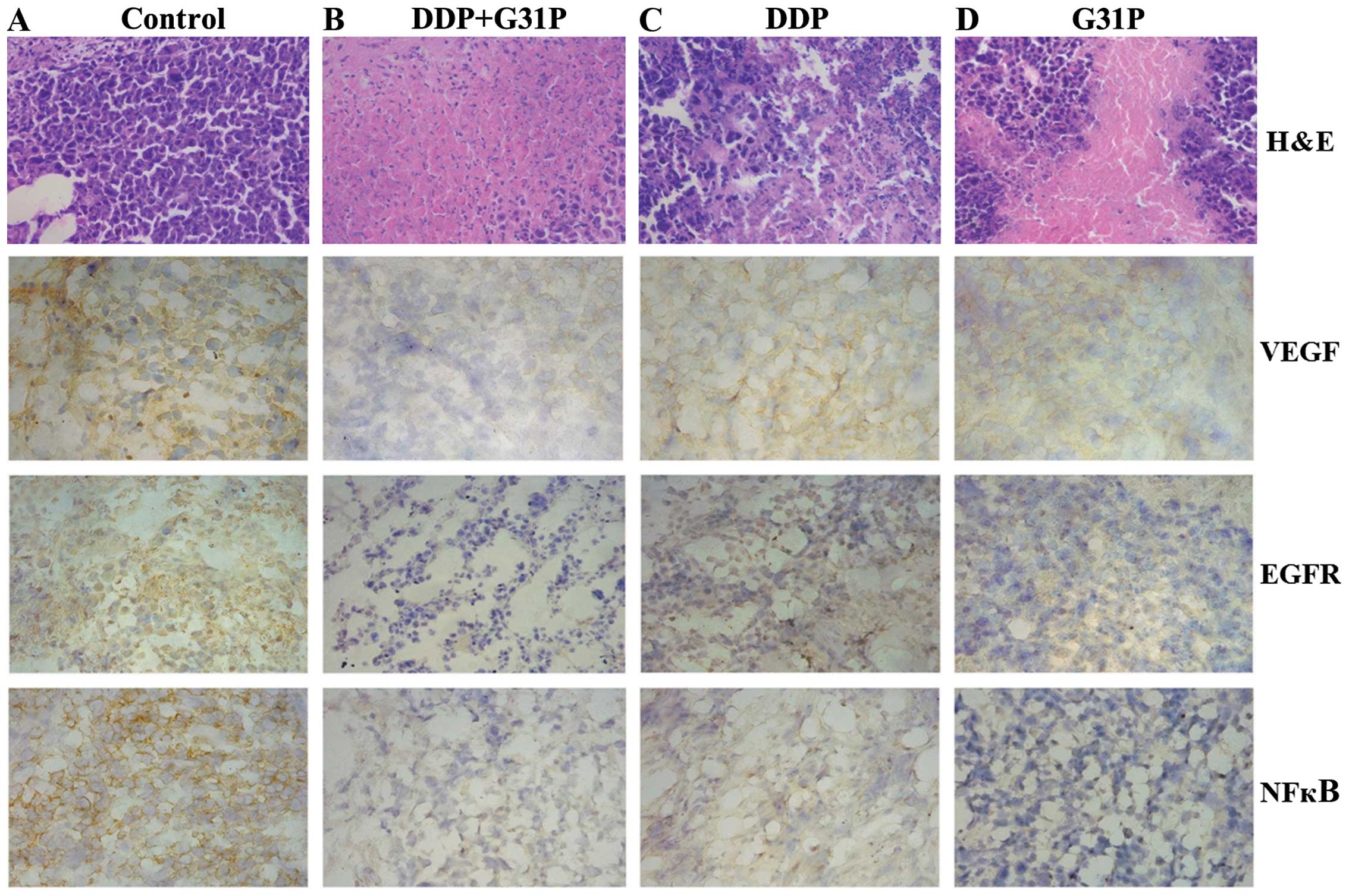
Primary tumors were sectioned and stained with
H&E. H22 cells of different shape and size were patchy or
diffused in tumor tissue with large, hyperchromatic nucleus. A
large proportion of the nucleus but less cytoplasm was evident in
cells. Significant necrosis, cytoplasmic shrinkage and cell
fragmentation were not identified in the control group as compare
to the remaining three groups. The DDP+G31P group showed a
significant tumor necrotic area and cell debris with large amounts
of nuclear fragmentation as compared to the DDP and G31P groups
(Fig. 2).
G31P reduces the expression of EGFR,
VEGF, MMP-2 and NF-κB in tumor tissue
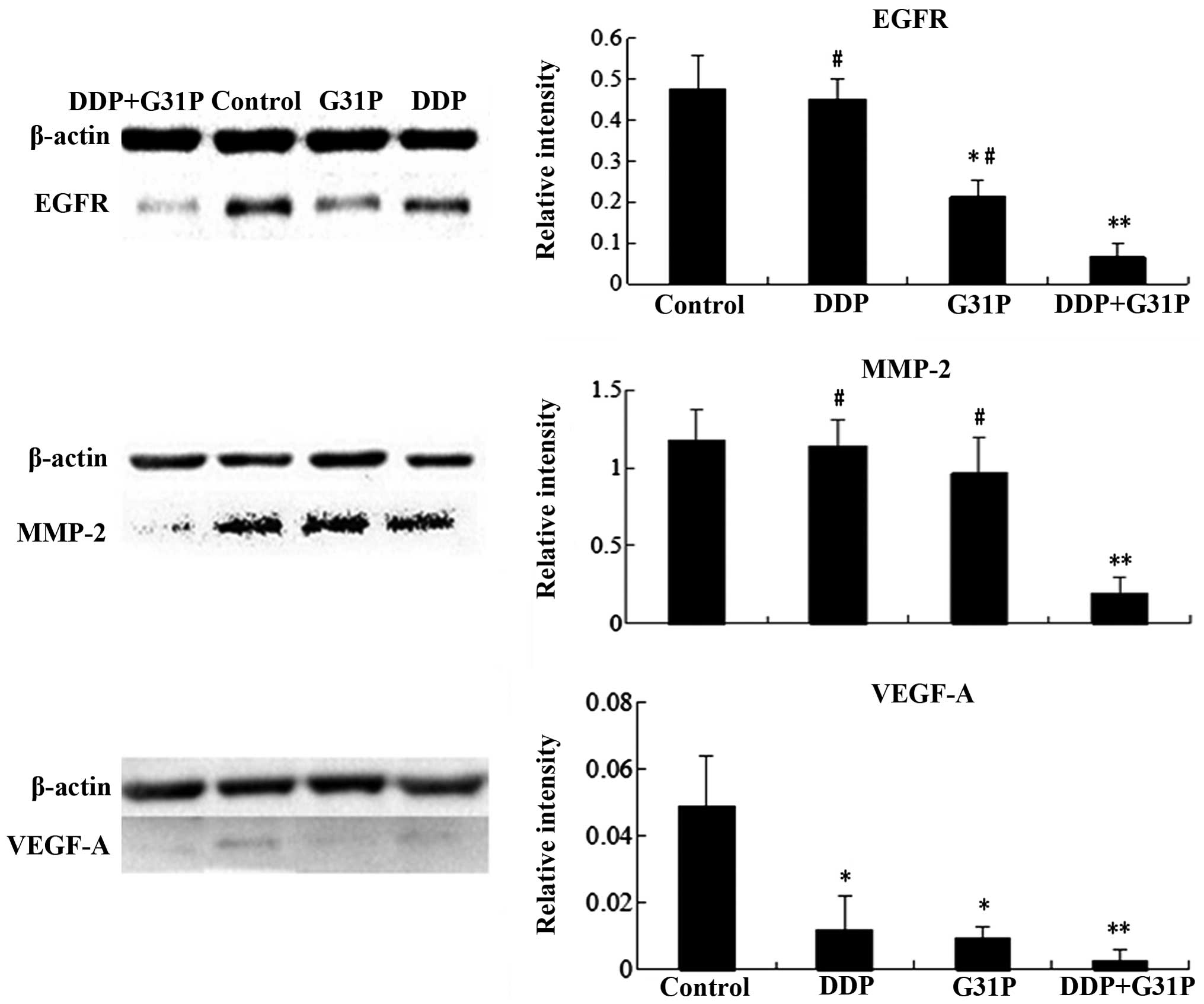
Western blot analysis was performed to evaluate
EGFR, MMP-2 and VEGF-A expression in tumor tissue of the control,
DDP, G31P and DDP+G31P groups (Fig.
3). We found that EGFR, MMP-2 and VEGF-A levels in the DDP+G31P
group were significantly lower than those of the control group
(P<0.01 vs. control). The difference of EGFR and MMP-2 between
the DDP+G31P and alone drug treatment groups (DDP and G31P) was
significant (P<0.05 vs. DDP+G31P). The G31P treatment group
markedly inhibited the expression of EGFR and VEGF-A (P<0.05 vs.
control). Significant VEGF-A inhibition was also identified in the
DDP treatment group (P<0.05 vs. control).
Immunohistochemical analysis was performed to
evaluate EGFR, VEGF-A and NF-κB expression in the control, DDP,
G31P and DDP+G31P groups (Fig. 2).
The evident difference of EGFR, VEGF-A and NF-κB was detected
between the DDP+G31P and control groups. A major difference in
NF-κB expression between the G31P or DDP and control group was
found.
G31P improves renal function and reduces
kidney tissue damage in cisplatin-induced ARF in mice
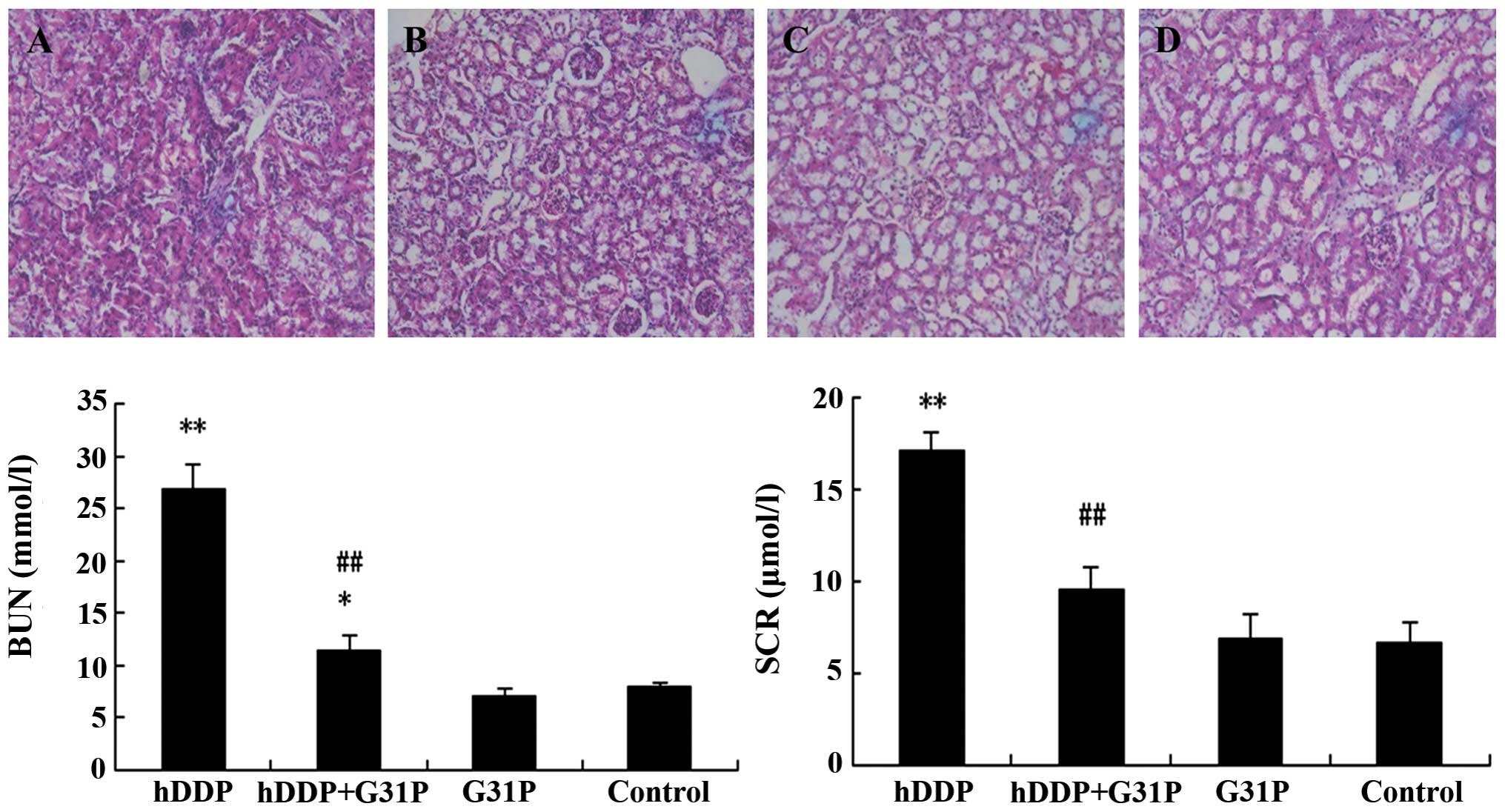
Renal function was assessed on the 9th day after
drug treatment. As shown in Fig. 4,
high-dose cisplatin injection significantly increased BUN and SCr
levels (P<0.01 vs. control). By contrast, combined therapy of
G31P+DDP significantly reduced BUN and SCr (P<0.01 vs. DDP)
while the G31P group did not show any significant change of BUN and
SCr compared with the control group. Tissues were resected from the
kidney for histopathological examination using H&E staining
from mice of the DDP, DDP+G31P, G31P and control groups (Fig. 4). In the DDP group, cisplatin caused
renal damage with loss of the brush border, and necrosis of tubular
cells. Tubular structure was disorganized, and fibrous connective
tissue hyperplasia was evident. The tubular injury score of the DDP
group was 4.0±0.36. In the G31P+DDP group, we found that G31P
treatment significantly reduced these changes with improvement of
the tubular injury score (1.0±0.20) while the G31P and control
groups did not show significant tubular structure destruction, with
scores of 0.3±0.47.
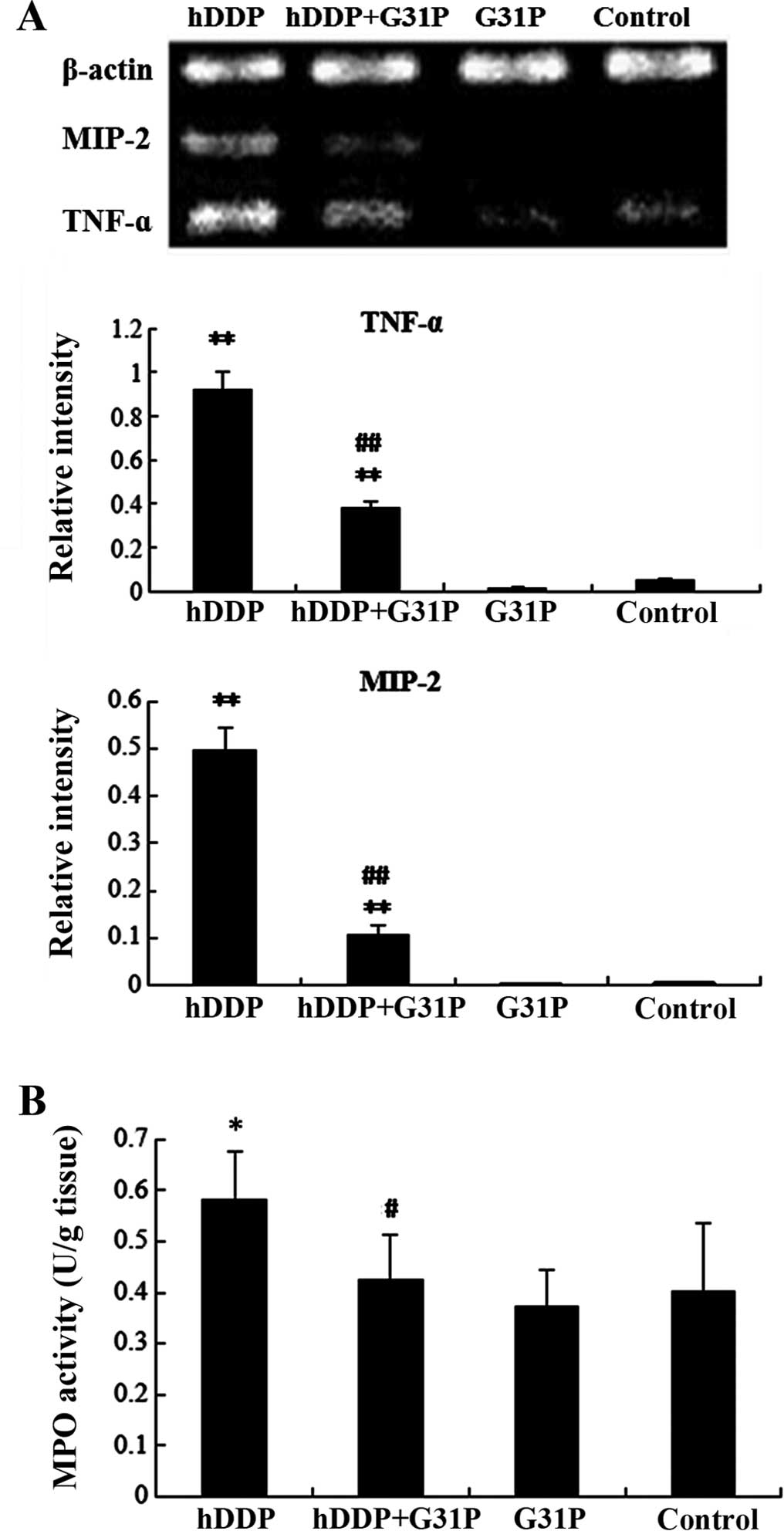
G31P reduces inflammatory cytokines and
MPO activity in kidney tissue
TNF-α and MIP-2 levels in kidney of the DDP group
were significantly higher than those of the control group
(P<0.01). However, G31P treatment significantly reduced the
levels of TNF-α and MIP-2 in kidney. There was an obvious
difference in the mRNA expression of TNF-α and MIP-2 between the
DDP and DDP+G31P groups (P<0.01, Fig. 5).
To determine the neutrophil infiltration in kidney
tissue when treated with a high dose of DDP, MPO activity was
measured (Fig. 5). MPO activity of
the DDP group was 0.58±0.10 U/gm of tissue, which was significantly
higher than that of the control group at 0.40±0.13 U/gm of tissue
(P<0.05). MPO activity of the DDP+G31P group was 0.42±0.09 U/gm
of tissue, which was significantly lower than that of the DDP group
(P<0.05). However, MPO showed no significant difference between
the G31P group (0.37±0.07 U/gm of tissue) and the control group
(0.40±0.13 U/gm of tissue).
Discussion
Results from the present study have shown that
combined therapy of G31P and DDP increased tumor deterioration and
reduced high-dose DDP-induced nephrotoxicity in H22 HCC-bearing
mice by inhibiting renal tubular endothelial cell necrosis and
blocking inflammatory cell infiltration. Similar results were
considered as G31P highly effective in blocking neutrophil
recruitment into the microbial and non-microbial inflammatory
response (17).
DDP is a chemotherapeutic drug used clinically for
the treatment of many types of tumors including HCCs. DDP induces
apoptosis in tumor cells by affecting the metabolism of proteins
and damages the structure of the cell membrane (18). The antitumor effect of DDP is
dose-dependent and as it is mainly excreted by kidneys, causes
substantial dose-dependent toxic renal side effects. Nephrotoxicity
is frequent and is the major limitation in cisplatin-based
chemotherapy (5,6). There are several mechanisms that
contribute to renal dysfunction following exposure to DDP including
direct tubular toxicity in the form of necrosis of endothelial
cells cause the excretion of cytokines, with neutrophil
infiltration leading to inflammation (19,20).
Due to limited effectiveness when any anti-carcinogen is used alone
and an obviously increased toxicity with dose increase, there is no
exception of combined therapy. Furthermore, combined chemotherapy
contributes to improving therapeutic effectiveness, dispersing
toxicity and overcoming drug resistance (21).
IL-8 is considered one of the major mediators of the
inflammatory response and this cancer-derived chemokine contributes
to tumor progression through the chemoattractive function and
regulation of angiogenesis, cancer cell growth and survival, as
well as tumor cell migration (22).
Expression of IL-8 was detected in human malignant liver tumor
tissue (23,24) where endothelial cells are present
and are responsible for lymphocyte recruitment particularly in
neutrophils through the activation of CXCR1/2 (25). Consequently, by antagonizing IL-8 by
blocking its interaction with CXCR1/2, G31P inhibits the biological
activity of IL-8, thereby inhibiting the proliferation of H22
carcinoma, as mentioned above, with decreased MTT activity.
Furthermore, G31P deteriorates the development of cancer in
vivo by inhibiting the implanted hepatocarcinoma in mice and
prolongs the survival phase of mice with no obvious toxicity. These
result are in concordance with previously reported findings
demonstrating G31P tumor growth inhibition and apoptosis in
prostate and lung cancers (15).
HCCs are vascular tumors and increased levels of
VEGF, MMP-2 and NFκB expression has been associated with increased
microvessel density (MVD) and tumor growth (26,27).
G31P suppression of the angiogenic effect of ELR-CXC chemokines was
also proven in prostate cancer (15). IL-8 increased tumor proliferation
through the transactivation of EGFR (28). Our results showed that G31P
treatment reduced EGFR expression, which downstream leads to tumor
growth inhibition and cause apoptosis.
TNF-α is crucial in the pathogenesis of
cisplatin-induced ARF (29). Our
results confirm that high-dose DDP therapy increased proinflamatory
cytokines including TNF-α and MIP-2. MPO is an enzyme mainly
released by activated neutrophils and characterized by efficient
pro-oxidative and proinflammatory properties in mouse kidney
(30). Treatment with G31P reduced
TNF-α and MIP-2 expression and MPO activity was evidently
decreased, confirming that G31P significantly reduced neutrophil
chemotaxis to the site of the infection and effectively alleviated
the inflammatory response and reduced unnecessary tissue damage to
improve the survival rate.
In conclusion, combined G31P and DDP significantly
inhibited the growth of mouse hepatoma cells and the expression of
EGFR, VEGF, MMP-2 and NF-κB of tumor tissues. G31P enhances the
efficacy of DDP in cancer treatment, mitigates cisplatin-induced
nephrotoxicity and improves the survival rate of mice by reducing
renal pathological damage, infiltration of neutrophils in kidney,
and the expression of TNF-α and MIP-2 in kidney tissues, caused by
DDP. These findings may provide a new approach for the prevention
and treatment of cancer.
Acknowledgements
This study was supported by a grant from the
National Science Foundation of China (grant no. NSFC30772023).
References
|
1
|
Forner A, Llovet JM and Bruix J:
Hepatocellular carcinoma. Lancet. 379:1245–1255. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shen FZ, Wang J, Liang J, Mu K, Hou JY and
Wang YT: Low-dose metronomic chemotherapy with cisplatin: can it
suppress angiogenesis in H22 hepatocarcinoma cells? Int J Exp
Pathol. 91:10–16. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ozdogan O, Ertay T, Arslan G, et al: Does
cisplatin chemotherapy decrease the MDP uptake of normal bone? An
experimental study. Ann Nucl Med. 22:357–362. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Arany I and Safirstein RL: Cisplatin
nephrotoxicity. Semin Nephrol. 23:460–464. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bonventre JV: Pathophysiology of ischemic
acute renal failure. Inflammation, lung-kidney cross-talk, and
biomarkers. Contrib Nephrol. 144:19–30. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ramesh G and Reeves WB: TNFR2-mediated
apoptosis and necrosis in cisplatin-induced acute renal failure. Am
J Physiol Renal Physiol. 285:610–618. 2003.
|
|
7
|
Faubel S, Lewis EC, Reznikov L, Ljubanovic
D, Hoke TS, Somerset H, Oh DJ, Lu L, Klein CL, Dinarello CA and
Edelstein CL: Cisplatin-induced acute renal failure is associated
with an increase in the cytokines interleukin (IL)-1β, IL-18, IL-6,
and neutrophil infiltration in the kidney. J Pharmacol and Exp
Ther. 322:8–15. 2007. View Article : Google Scholar
|
|
8
|
Mukaida N: Pathophysiological roles of
interleukin-8/CXCL8 in pulmonary diseases. Am J Physiol Lung Cell
Mol Physiol. 284:L566–L577. 2003.PubMed/NCBI
|
|
9
|
Wang Y, Wang W, Wang L, Wang X and Xia J:
Regulatory mechanisms of interleukin-8 production induced by tumor
necrosis factor-α in human hepatocellular carcinoma cells. J Cell
Mol Med. 16:496–506. 2012. View Article : Google Scholar
|
|
10
|
Zhao X, Li F, Town JR, Zhang X, Wang W and
Gordon JR: Humanized forms of the CXCR1/CXCR2 antagonist, bovine
CXCL8(3-74)K11R/G31P, effectively block ELR-CXC
chemokine activity and airway endotoxemia pathology. Int
Immunopharmacol. 7:1723–1731. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhao X, Town JR, Li F, Zhang X, Cockcroft
DW and Gordon JR: ELR-CXC chemokine receptor antagonism targets
inflammatory responses at multiple levels. J Immunol.
182:3213–3222. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gordon JR, Li F, Zhang X, Wang W, Zhao X
and Nayyar A: The combined CXCR1/CXCR2 antagonist
CXCL8(3-74)K11R/G31P blocks neutrophil infiltration,
pyrexia, and pulmonary vascular pathology in endotoxemic animals. J
Leukoc Biol. 78:1265–1272. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao X, Town JR, Li F, Li W, Zhang X and
Gordon JR: Blockade of neutrophil responses in aspiration pneumonia
via ELR-CXC chemokine antagonism does not predispose to airway
bacterial outgrowth. Pulm Pharmacol Ther. 23:22–28. 2010.
View Article : Google Scholar
|
|
14
|
Zhao X, Town JR, Yang A, Zhang X, Paur N,
Sawicki G and Gordon JR: A novel ELR-CXC chemokine antagonist
reduces intestinal ischemia reperfusion-induced mortality, and
local and remote organ injury. J Surg Res. 162:264–273. 2010.
View Article : Google Scholar
|
|
15
|
Liu X, Peng J, Sun W, Yang S, Deng G, Li
F, Cheng JW and Gordon JR: G31P, an antagonist against CXC
chemokine receptors 1 and 2, inhibits growth of human prostate
cancer cells in nude mice. Tohoku J Exp Med. 228:147–156. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Qin Y, Fan F, Zhao Y, Cui Y, Wei X, Kohama
K, Gordon JR, Li F and Gao Y: Recombinant human CXCL8 (3-72)
K11R/G31P regulates smooth muscle cell proliferation and migration
through blockage of interleukin-8 receptor. IUBMB Life. 65:67–75.
2013. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wei J, Peng J, Wang B, Qu H, Wang S,
Faisal A, Cheng JW, Gordon JR and Li F: CXCR1/CXCR2 antagonism is
effective in pulmonary defense against Klebsiella pneumoniae
infection. Biomed Res Int. 2013:7209752013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shen DW, Pouliot LM, Hall MD and Gottesman
MM: Cisplatin resistance: a cellular self-defense mechanism
resulting from multiple epigenetic and genetic changes. Pharmacol
Rev. 64:706–721. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Friedewald JJ and Rabb H: Inflammatory
cells in ischemic acute renal failure. Kidney Int. 66:486–491.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Okusa MD: The inflammatory cascade in
acute ischemic renal failure. Nephron. 90:133–138. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang W, Qin SK, Chen BA and Chen HY:
Experimental study on antitumor effect of arsenic trioxide in
combination with cisplatin or doxorubicin on hepatocellular
carcinoma. World J Gastroenterol. 7:702–705. 2001.
|
|
22
|
Yuan A, Chen JJ, Yao PL and Yang PC: The
role of interleukin-8 in cancer cells and microenvironment
interaction. Front Biosci. 10:853–865. 2005. View Article : Google Scholar
|
|
23
|
Xie K: Interleukin-8 and human cancer
biology. Cytokine Growth Factor Rev. 12:375–391. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Harimoto N, Shirabe K, Abe T, et al:
Interleukin-8 producing hepatocellular carcinoma with pyrexia. HPB
Surg. 1:461–492. 2009.
|
|
25
|
Yoong KF, Afford SC, Jones R, Aujla P, Qin
S, Price K, Hubscher SG and Adams DH: Expression and function of
CXC and CC chemokines in human malignant liver tumors: a role for
human monokine induced by gamma-interferon in lymphocyte
recruitment to hepatocellular carcinoma. Hepatology. 30:100–111.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang ZF and Poon RT: Vascular changes in
hepatocellular carcinoma. Anat Rec. 291:721–734. 2008. View Article : Google Scholar
|
|
27
|
Tanaka H, Yamamoto M, Hashimoto N,
Miyakoshi M, Tamakawa S, Yoshie M, Okusashi Y, Yokoyama K, Yaginuma
Y and Ogawa K: Hypoxia-independent overexpression of
hypoxia-inducible factor 1α as an early change in mouse
hepatocarcinogenesis. Cancer Res. 66:11263–11270. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Luppi F, Longo AM, de Boer WI, Rabe KF and
Hiemstra PS: Interleukin-8 stimulates cell proliferation in
non-small cell lung cancer through epidermal growth factor receptor
transactivation. Lung Cancer. 56:25–33. 2007. View Article : Google Scholar
|
|
29
|
Wu Y and Zhou BP: TNF-α/NF-κB/Snail
pathway in cancer cell migration and invasion. Br J Cancer.
102:639–644. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Guilpain P, Servettaz A, Batteux F,
Guillevin L and Mouthon L: Natural and disease associated
anti-myeloperoxidase (MPO) autoantibodies. Autoimmun Rev.
7:421–425. 2008. View Article : Google Scholar : PubMed/NCBI
|