Introduction
Multiple myeloma (MM) is a malignant plasma-cell
disorder characterized by the abnormal proliferation of plasma
cells in the bone marrow microenvironment, monoclonal protein in
blood or urine, and relevant organ dysfunction (1). MM accounts for 1% of all types of
cancer and 13% of hematologic malignancies, and the incidence and
prevalence of MM increases with age, thus the median age at
diagnosis is 70 years (2).
Consequently, MM is prevalent in the elderly, and pre-existing
comorbidities are usually present at diagnosis, type 2 diabetes
being the most important one as its incidence is on the increase
worldwide, along with the aging of population (3). It has been established that the
morbidity of pre-existing diabetes in elderly patients with MM is
~6–11% (3). Besides pre-existing
diabetes, application of dexamethasone may cause steroid-induced
diabetes (4), contributing to the
increased prevalence of diabetes in elderly MM patients. Thus, more
attention should be focused on the treatment of MM patients with
diabetes.
The impact of pre-existing diabetes or
steroid-induced diabetes on patients with MM has been suggested in
a few clinical studies (3,5–7).
Diabetes is associated with increased risks of adverse events such
as peripheral neuropathy and venous thrombosis in MM patients who
received bortezomib, thalidomide or lenalidomide (3). In addition, another study published by
Jung et al (5) evaluated the
incidence of hyperglycemia during induction therapy in 155 MM
patients and its effect on serious infections during that period of
treatment. Those authors found that 12.9% of the patients developed
hyperglycemia (≥200 mg/dl) and serious infections occurred in 18.1%
of this population, while infection-associated mortality occurred
in one patient (5). These adverse
events may influence the integrity of the treatment. Regarding the
clinical outcomes, diabetes was proven to be an indicator of poor
prognosis. Chou et al enrolled 310 MM patients who were
identified with pre-existing diabetes (3). Compared with their non-diabetic
counterparts, MM patients with diabetes had a significantly higher
proportion of renal dysfunction, advanced clinical staging, and
higher all-cause mortality risk (HR=1.509, P=0.037). In accordance
with this result, Wu et al found that diabetes,
steroid-induced diabetes in particular, is associated with poor
clinical outcomes in MM (7). The
highest level of post-load glycemia was associated with an
evaluated risk of mortality in MM (HR=3.06, P<0.05) in another
study by Chiu et al (8).
These findings demonstrated the unfavorable impact of pre-existing
and steroid-induced diabetes on clinical features and OS in
patients with MM. However, little is known regarding how diabetes
or hyperglycemia affects the efficacy of treatment for patients
with MM.
Hyperglycemia influences the biological activities
in cells and tissues. Several biochemical pathways have been proven
to be closely associated with hyperglycemia, one of the best known
pathways is high-glucose-mediated upregulation of reactive oxygen
species (ROS) (9,10). ROS are the by-products of cellular
metabolism, and mitochondria are the main sources of ROS (11). Various antioxidant systems for
removing ROS exist in cells. In general, the generation and
clearance of ROS are maintained in the dynamic balance. When the
production of ROS is out of control, it usually leads to oxidative
stress, even apoptosis (12,13).
Normal cells in a high-glucose environment usually generate
excessive ROS, leading to cell damage (14,15).
It was hypothesized that hyperglycemia-induced ROS was useful in
improving the efficacy of anti-myeloma agents such as bortezomib or
dexamethasone. However, the existing clinical evidence mentioned
above did not support this hypothesis. Thus, the cause for the
impaired anticancer effect of chemotherapy on MM with hyperglycemia
remains to be determined.
Peroxisome proliferator-activated receptor γ
coactivator-1α (PGC-1α) is a transcriptional coactivator of nuclear
receptors and plays critical roles in various metabolic pathways,
including oxidative phosphorylation, fatty acid oxidation, glucose
metabolism, and cellular respiration (16). PGC-1α has been well studied in
diabetes and exercise, however, little is known concerning the role
of PGC-1α in MM. In a previously published study, we found that
PGC-1α was upregulated in MM cells and was associated with glucose
metabolism and angiogenesis (17).
It has been suggested that PGC-1α induces several key
ROS-detoxifying enzymes when cells experience hyperglycemia and
oxidative stress (18–21). However, it is not clear whether
PGC-1α is involved in regulating ROS in MM under hyperglycemic
conditions and thus affecting the efficacy of chemotherapy.
Based on the observations that diabetes affects MM
response to drugs and that hyperglycemia may interfere with the
cellular production of ROS in MM, we investigated whether
hyperglycemia influenced the axis PGC-1α-ROS-antioxidants in the
setting of the increased level of glucose. For this purpose, we
cultured MM cells in 10 mM glucose culture medium, and then treated
these cells with bortezomib. Our study provided in vitro
evidence to explain the reason for MM patients with high-glucose
condition suffering poor outcome when compared with
non-hyperglycemic MM patients, and suggested that PGC-1α is a
potential target for improving the efficacy of treatment in MM
patients with diabetes or steroid-induced diabetes.
Materials and methods
Cell culture
RPMI-8226, U266 and ARH77 MM cell lines were
purchased from the American Type Culture Collection (Manassas, VA,
USA). The cells were cultured in RPMI-1640 medium (HyClone, Thermo
Fisher Scientific, Rockford, IL, USA) containing 10% fetal bovine
serum and 100 U/ml penicillin/streptomycin, and were incubated at
37°C under 5% CO2 conditions. To create a hyperglycemic
condition in vitro, an additional 625 mg of glucose were
supplemented into 500 ml ordinary RPMI-1640 medium. Bortezomib
(kindly provided by Professor Yu Hu) and dexamethasone
(Sigma-Aldrich, St. Louis, MO, USA) were used as chemotherapeutic
agents to treat MM cells.
Small-interference RNA (siRNA)
To inhibit the expression of PGC-1α, we introduced
the siRNA-targeting PGC-1α which was designed and produced by
GenePharma (GenePharma, Suzhou, China). Briefly, MM cells were
treated with the Lipofectamine 2000™ reagent (Invitrogen, Carlsbad,
CA, USA) according to the manufacturer’s instructions. MM cells
were collected at 24 h after transfection. The sequence information
for siRNA is presented as: siRNA for GAPDH, sense and antisense
control DNA (provided with the kit); siRNA for PGC-1α,
5′-GCCAAACCAACAACUUAUU-3′ (forward) and 5′-AUAAAGUUGUUGGUUGGCUU-3′
(reverse).
MTT assay
MM cells cultured in normal or hyperglycemic
conditions were seeded in 96-well plates at a density of
3.5–5×104 cells/well. Subsequent to treatment with
different doses of chemotherapeutic agents including bortezomib and
dexamethasone for 24 h, 10 μl MTT
[3-(4,5-dimethylthiazolyl-2)-2,5-diphenyltetrazolium bromide]
solution was added to each well. The plates were then incubated for
4 h in the dark and detergent reagent was added until the purple
formazan was completely dissolved. The assay was performed by a
microplate reader (Bio-Rad, La Jolla, CA, USA) and the absorbance
at 490 nm was measured and recorded. The inhibition rate of
proliferation was defined as: Inhibition rate = (
ODcontrol - ODdrug - ODbackground
)/( ODcontrol - ODbackground ) ×100%.
cDNA synthesis and quantitative PCR
analysis
The procedures were performed according to
previously described methods (17).
Total RNAs in MM cells were extracted by using the TRIzol reagent
(Invitrogen). cDNA was then synthesized using a FSQ-101 cDNA
Reverse Transcription kit (Toyobo, Life Science Department, Osaka,
Japan). Quantitative RT-PCR (RT-qPCR) was performed using the
SYBR-Green PCR system on a Life ABI 7500 instrument (Applied
Biosystems, Foster City, CA, USA), and the results were normalized
to β-actin. Primer sequences used were: β-actin: 5′-TTC CAG CCT TCC
TTC CTG G-3′ (forward) and 5′-TTG CGCTCAGGAGCAAT-3′ (reverse);
PGC-1α: 5′-TGGTGCCAC CACCATCAAAGA-3′ (forward) and 5′-TCACCAAACAGC
CGCAGACTG-3′ (reverse); catalase (CAT): 5′-CCAGTCGGT GTATGCCTTCT-3′
(forward) and 5′-GGACGCCACATTCTC GATAAG-3′ (reverse); glutathione
peroxidase-1: 5′-CCA GTCGGTGTATGCCTTCT-3′ (forward) and 5′-GGACGC
CACATTCTCGATAAG-3′ (reverse); glutathione peroxidase-4:
5′-GCTGTGGAAGTGGATGAA-3′ (forward) and 5′-GATGAGGAACTGTGGAGAG-3′
(reverse); superoxide dismutase-1: 5′-ACTCATCTGTTATCCTGCTAG-3′
(forward) and 5′-GCCTCATAATAAGTGCCA TAC-3′ (reverse); superoxide
dismutase-2: 5′-TCACCGAGGAGAAGTACC-3′ (forward) and
5′-TTGATATGACCACCACCATT-3′ (reverse).
ROS
Briefly, MM cells cultured in normal or high glucose
were collected and incubated with 10 μM DCFH-DA (Sigma-Aldrich) for
30 min at 37°C. The suspended cells were rotated gently every 5 min
during the period of incubation. The cells were then washed and
resuspended in PBS. The mean fluorescence intensity of DCF was
detected by the FACSCalibur flow cytometer and analyzed using Cell
Quest software (both from BD Biosciences, San Jose, CA, USA).
Western blotting
Whole cell lysate as well as SDS-PAGE,
electrophoretic transfer and immunoblotting were prepared as
previously described (22). Protein
samples were analyzed using chemiluminescence detection (ECL;
Amersham Pharmacia, Buckinghamshire, UK, GB). Antibodies used in
this section were purchased from Cell Signaling Technology. Total
protein concentration was measured according to the manufacturer’s
instructions (Thermo Scientific Pierce, Rockford, IL, USA).
Statistical analysis
SPSS 15.0 software (SPSS Inc., Chicago, IL, USA) was
used to analyze all recorded data. Data are presented as means ± SD
values. Differences between groups were analyzed by ANOVA or
Student’s t-test and were considered to be statistically
significant if P<0.05. Each experiment was repeated at least
three times, independently.
Results
Hyperglycemia decreases the toxicity of
chemotherapeutic agents
Human RPMI-8226, U266 and ARH-77 MM cells were
cultured in hyperglycemic conditions continuously, and treated with
different chemotherapeutic agents including bortezomib and
dexamethasone independently. After 24 h of treatment, we examined
the toxicity of bortezomib and dexamethasone in RPMI-8226, U266 and
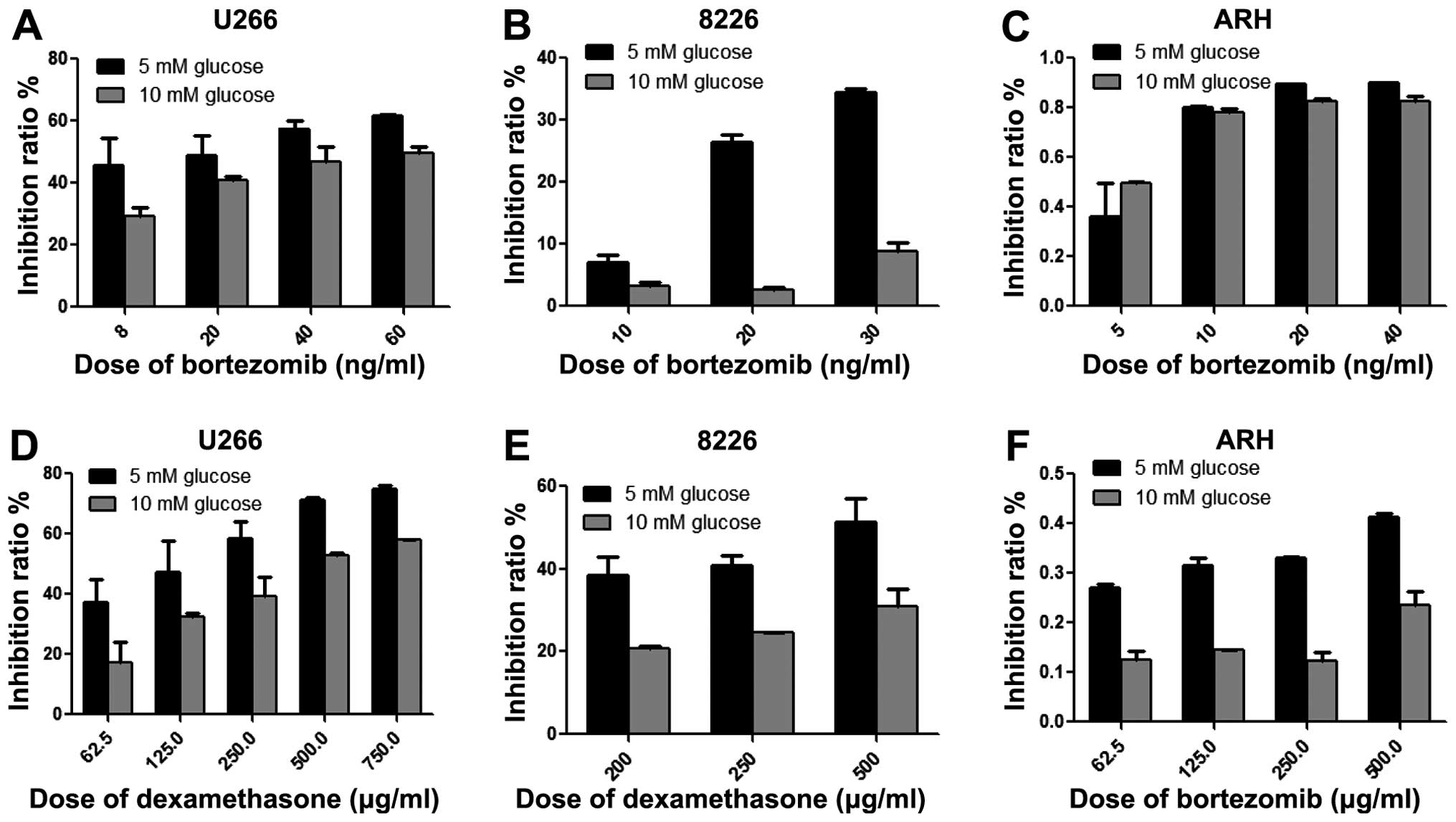
ARH77 by MTT. As shown in Fig. 1,
MM RPMI-8226, U266 and ARH77 cells cultured in high glucose showed
decreased toxicity of bortezomib. The inhibitory rates of
bortezomib at the same dose were much lower than those in cells
grown in normal glucose conditions. These data indicated that
hyperglycemia reduced the efficacy of chemotherapeutic agents. This
is in accordance with the results from studies showing that
elevated blood glucose levels are associated with a poor outcome in
cancer patients and hyperglycemia is a prognostic factor with
higher risk of death in lung and breast cancer patients (23,24).
In addition, 20 mM glucose treatment of U266, ARH77 and other cell
lines caused a decrease of toxicity of dexamethasome that may
contribute to the resistance of chemotherapy (25).
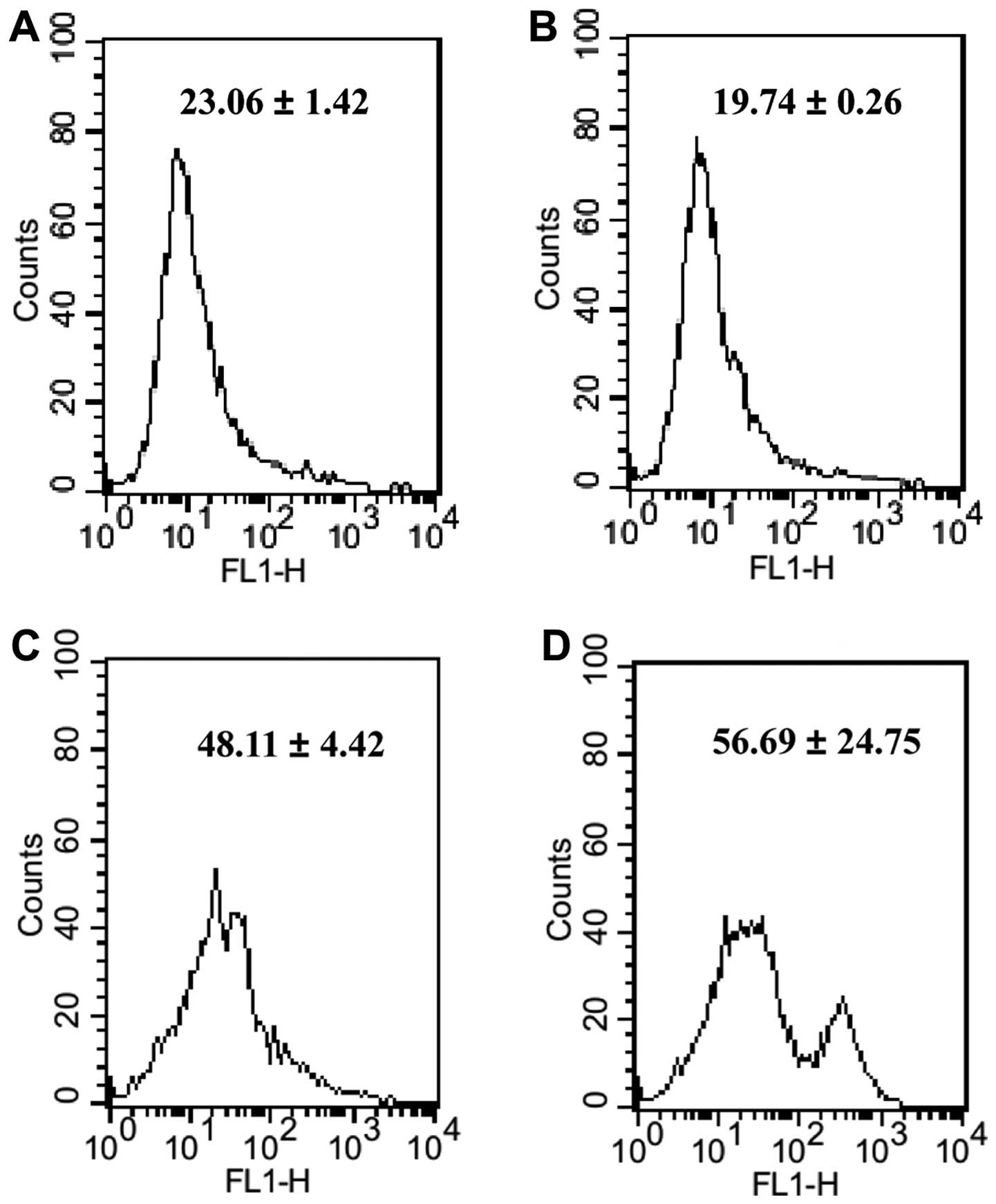
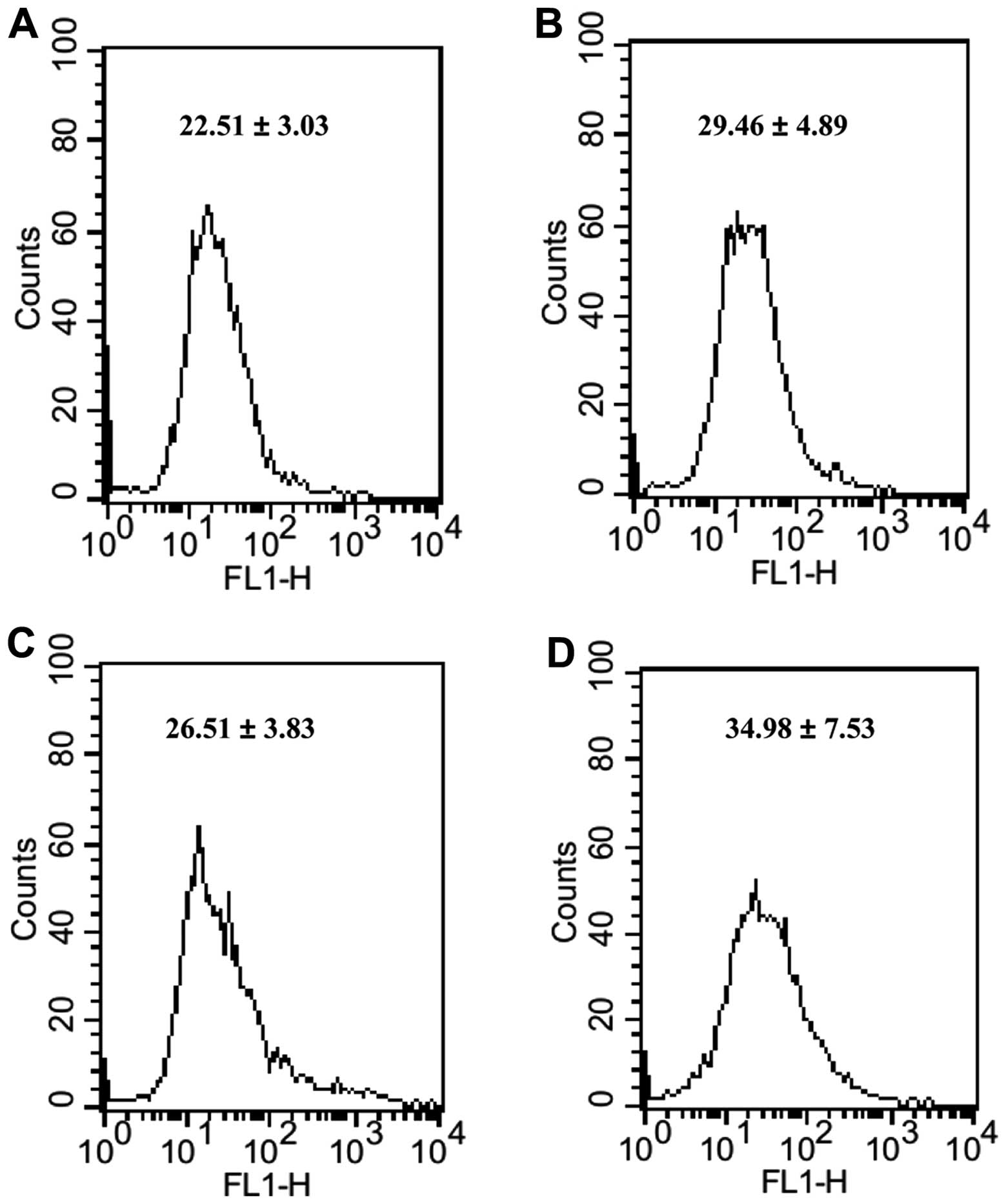
We examined the level of ROS in MM cells which were
incubated in high-glucose culture medium, before and after
treatment of bortezomib. The cellular ROS level was labeled by
CM-H2DCFDA and measured using a FACSCalibur flow cytometer. As
shown in Fig. 2, following
treatment of chemotherapeutic agents, we observed that the ROS
level in MM RPMI-8226 cells cultured in high-glucose culture medium
were a little higher than that of control. The fold-change of ROS
in the cells treated with bortezomib and 10 mM glucose was >2
when compared with 10 mM glucose alone.
PGC-1α and relevant antioxidant factors
are upregulated in MM cells treated with chemotherapeutic agents
under hyperglycemic conditions
Levels of PGC-1α are highly regulated by cellular
stress and therefore allow metabolic adaptation (26). Thus, we assessed the impact of high
glucose on the expression of PGC-1α in the RPMI-8226 MM cell line.
We measured the relative expression of PGC-1α mRNA using RT-PCR. As
shown in Fig. 3, levels of PGC-1α
mRNA in cells cultured in 10 mM glucose condition were higher than
those cultured in normal medium. We assessed PGC-1α expression in
cells treated with bortezomib, and found that levels of PGC-1α were
further increased when compared with cells without treatment of
bortezomib, especially in cells grown in 10 mM glucose medium.
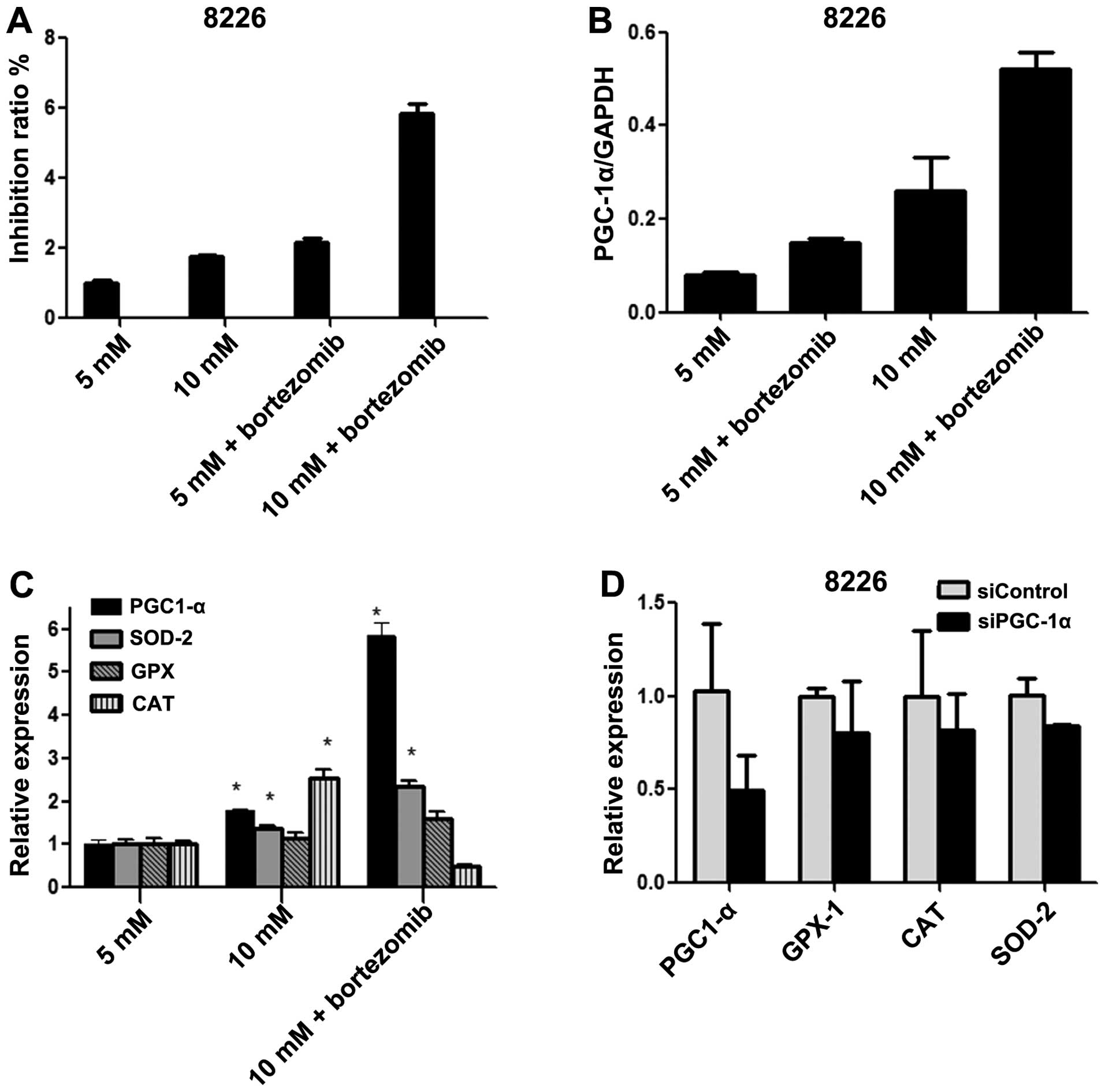 | Figure 3The expression of PGC-1α and related
antioxidant factors after chemotherapy or siRNA treatment. After
treating RPMI-8226 cells with bortezomib or siRNA, the relative
expression of PGC-1α, CAT, GPX-1 and SOD-2 are measured. (A, B and
C) Suppression of PGC-1α by siRNA resulted in a decreased
expression of PGC-1α, CAT, GPX-1 and SOD-2. (A and B) Expression of
PGC-1α in RPMI-8226 cells under normal glucose, high-glucose and
chemotherapy conditions. (C) Expression of PGC-1α, CAT, GPX-1 and
SOD-2 under normal glucose, high-glucose and chemotherapy
conditions, when compared to normal-glucose condition. (D) The
expression of PGC-1α, CAT, GPX-1 and SOD-2 in RPMI-8226 cells were
inhibited after treating cells with siPGC-1α, when compared to
siControl. MM, multiple, myeloma; SOD, superoxide dismutase; GPX,
glutathione peroxidase; CAT, catalase; Dex, dexamethasone; Bor,
bortezomib; Met, metformin; PGC-1α, peroxisome
proliferator-activated receptor γ coactivator-1α. |
The levels of PGC-1α protein in RPMI-8226 cells were
assessed by western blot analysis. We used metformin, an AMPK
activator, to induce the expression of PGC-1α in MM cells as a
positive control. We found an increased expression of PGC-1α
protein in cells grown in 10 mM glucose or treated with bortezomib
when compared with the control group. These findings indicate that
PGC-1α is also highly expressed in MM, as high glucose and
chemotherapeutic agents induce the expression of PGC-1α at the mRNA
and protein levels.
PGC-1α is known to be involved in the regulation of
the antioxidant system. However, it is unclear whether PGC-1α
retains the same function in MM. Thus, we assessed the impact of
PGC-1α on the expression of a set of genes relating to antioxidant
factors. Cells cultured in 5 or 10 mM glucose and treated with or
without 10 ng/ml bortezomib were collected, and mRNA expressions of
superoxide dismutase 2 (SOD-2), CAT and glutathione peroxidase 1
(GPX-1) in these cells were measured by RT-PCR. Increased
expression of SOD-2 and GPX-1 was observed in cells treated with 10
mM glucose and bortezomib (Fig. 3)
when compared to the control, and these cells also showed an
upregulated expression of PGC-1α.
Suppression of PGC-1α by siRNA reverses
the reduction of ROS and enhances the toxicity of chemotherapeutic
agents in MM cells
To elucidate the role of PGC-1α in the
transcriptional regulation of antioxidant factors, we suppressed
the expression of PGC-1α in cells with siRNA targeting PGC-1α. As
expected, the expression of PGC-1α was reduced by PGC-1α siRNA
(Fig. 3D). Addtionally, the
expression of SOD-2, CAT and GPX-1 were differentially reduced.
However, the levels of SOD-1, GPX-4 and PRDX-1 were not affected by
the suppression of PGC-1α (data not shown), indicating that other
pathways may exist that are independent of regulation by PGC-1α
during hyperglycemia.
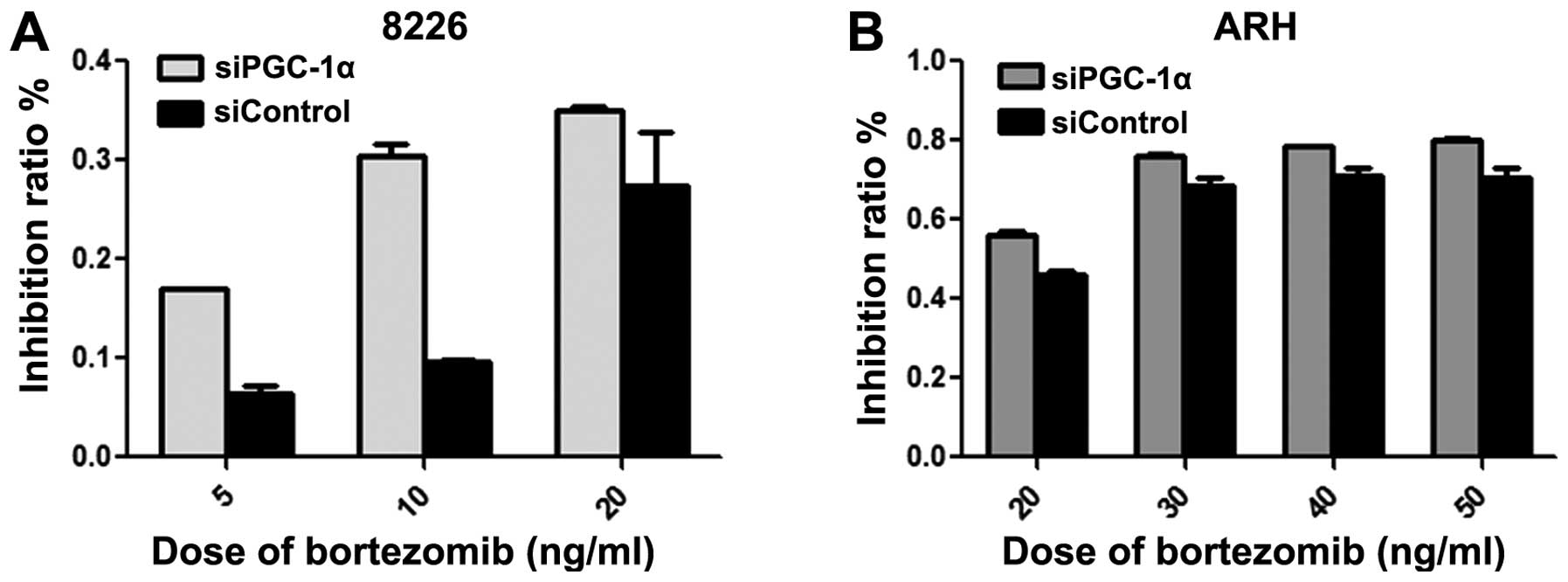
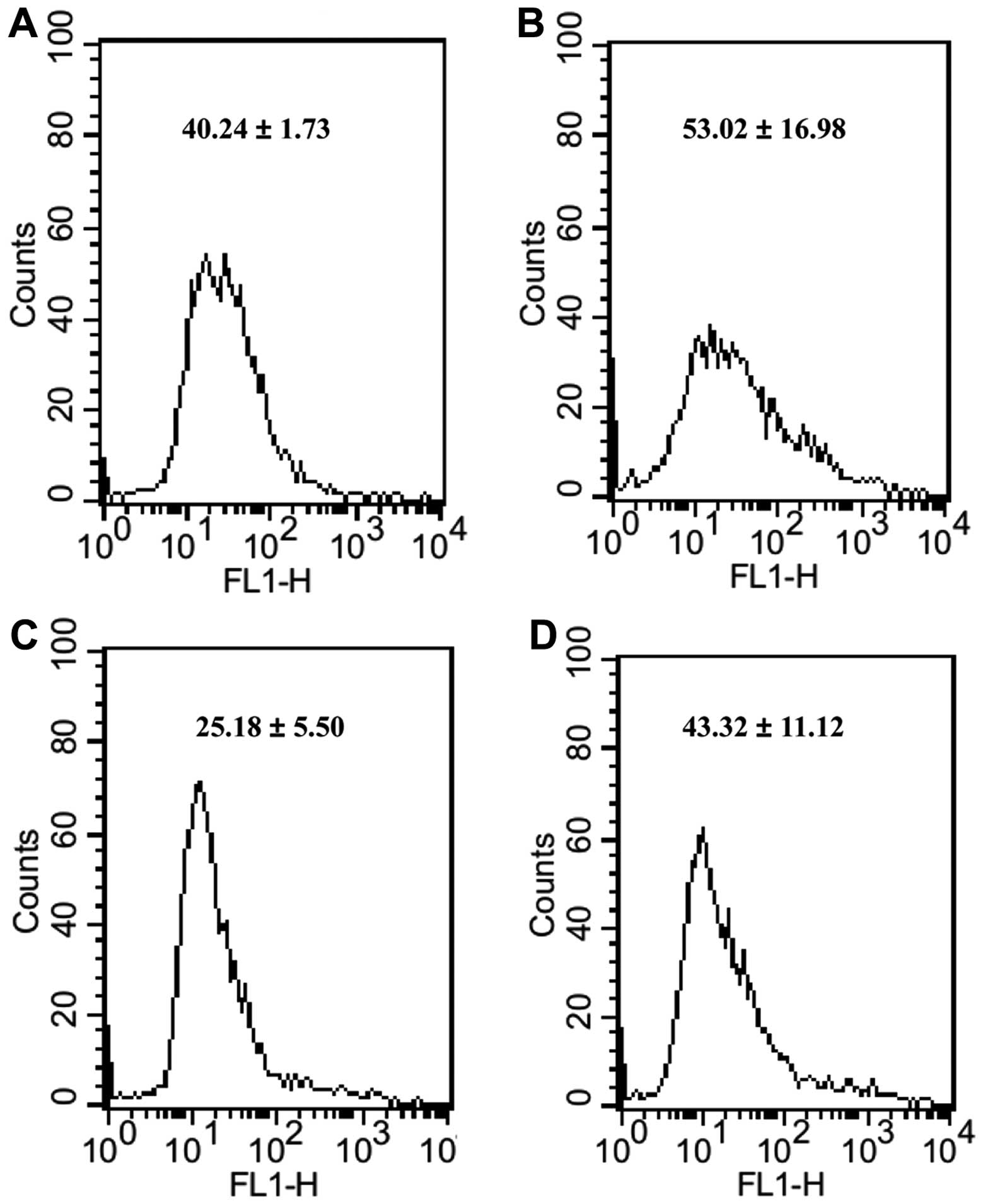
To determine the effect of inhibition of PGC-1α on
the amount of ROS and toxicity of bortezomib, we tested cellular
ROS level by FACSCalibur flow cytometer and toxicity of bortezomib
by MTT in cells transfected with siPGC-1α. The level of ROS was
increased (Figs. 4 and 5) and toxicity of bortezomib was enhanced
(Fig. 6) when compared with the
siControl group, suggesting that PGC-1α may induce the reduced ROS
level and toxicity of bortezomib regardless of hyperglycemia, thus
targeting PGC-1α may be beneficial for MM patients with or without
evaluated blood-glucose level.
Discussion
The impact of hyperglycemia on the therapeutic
effect of chemotherapeutic agents in MM, as well as how
hyperglycemia influences outcomes of MM during treatment, remains
poorly understood (6,25,27).
In the present study, we found that the cellular toxicity of
bortezomib was reduced when treating MM cells cultured under
high-glucose condition, accompanied with an increased expression of
PGC-1α and antioxidant factors such as SOD-2, CAT and GPX-1. The
levels of ROS in these MM cells were not significantly increased
following treatment with the agents, but a little decrease in ROS
levels. However, following treatment of cells with siPGC-1α, the
accumulation of ROS and the efficacy of chemotherapy agents were
enhanced in MM cells. Our results suggested that hyperglycemia did
not enhance or decrease the tumor-killing effect of
chemotherapeutic agents, and the upregulated PGC-1α and associated
antioxidant factors likely induced the indistinctive elevation of
ROS in MM cells cultured under high-glucose condition.
MM cells grown under high-glucose conditions readily
obtain more glucose as abnormal glucose uptake and metabolism exist
in MM, as identified in a PET-CT examination (28). Glucose is an important substance for
proliferation and survival of normal and malignant cells (29–31).
Glucose is mainly used to produce energy material such as ATP, and
can provide synthetic intermediates such as lipid and carbon
skeleton for cell proliferation. Studies have shown that glucose
participates in the regulation of a variety of key factors
associated with cell apoptosis, such as Mcl-1 and Bcl-2 (32–35).
In addition, glucose is capable of preventing the release of
cytochrome c and subsequent cell apoptosis by maintaining
the interaction of mitochondria and hexokinase II (36). In the present study, we found that
MM cells under hyperglycemia had a lower proportion of apoptosis
following treatment with chemotherapeutic agents, but a higher
level of PGC-1α, when compared with MM cells cultured at the normal
level of glucose. PGC-1α has been proved to be involved in the
regulation of glucose metabolism, and is a master regulator of
GLUT-4, which is an important glucose transporter in MM. We have
previously demonstrated that PGC-1α has the ability of upregulating
the expression of GLUT-4 in MM (17), and myeloma cells exhibit reliance on
constitutively cell surface-localized GLUT-4 for basal glucose
consumption, maintenance of Mcl-1 expression, growth, and survival
(28). Elevated glucose uptake and
metabolism enables MM cells to possess the advantages of
proliferation and anti-apoptosis mentioned above. Besides,
intracellular glucose can be used to generate intermediates such as
NADPH, and exert a fundamental role in maintaining the integrity of
the mitochondria (37). Molecules
such as NADPH are especially important for salvaging the oxidative
stress and reducing insult-associated-excessive ROS in cells. This
beneficial regulation allows the cells to enhance the ability to
preserve themselves from the damage of excessive ROS. Based on
these settings, hyperglycemia induced the expression of PGC-1α and
associated antioxidant factors (SOD-2, CAT and GPX-1), leading to
the relatively decreasing of ROS in MM cells, and eventually
resulting in reduced cellular toxicity of the chemotherapeutic
agents. This is in accordance with the results of Friday et
al who have reported that hyperglycemia increased the
IC50 value of dexamethasone in MM cells (25).
Apart from hyperglycemia induced by drugs, type 2
diabetes disease is present in ~10% of patients diagnosed as MM,
and these patients had a significantly higher mortality risk
compared with non-diabetic ones (3,4,6).
Therefore, hyperglycemia should be positively and effectively
controlled in order to improve survival outcomes. In a study
conducted by Vu et al it was shown that, the administration
of metformin and thiazolidinediones may be associated with improved
outcomes in acute lymphoblastic leukemia patients with
hyperglycemia (38). Our results
have demonstrated that high glucose attenuates the antitumor effect
of chemotherapeutic agents by inducing the expression of PGC-1α and
antioxidant factors. Although the treatment of patients with MM and
pre-existing diabetes or drug-induced hyperglycemia forms a complex
challenge for physicians, these findings suggest that it is
essential to treat diabetes and hyperglycemia to improve treatment
outcomes.
PGC-1α is a transcriptional coactivator initially
recognized as a functional activator of the peroxisome
proliferator-activated receptor (PPAR)-γ in brown adipose tissue
(39) and recently identified as an
important regulator of numerous aspects of metabolism by
interacting with nuclear receptors and transcription factors to
activate the transcription of their target genes (40–42).
In recent years, studies have determined that PGC-1α has a
regulatory mechanism for the expression of endogenous antioxidant
factors (40). In skeletal muscle
from PGC-1α knockout (KO) mice, reduced mRNA expression of SOD-1,
SOD-2, and/or GPX-1 was observed when compared to wild-type mice,
while PGC-1α overexpression showed an upregulation of SOD-2
(40). PGC-1α knockout fibroblasts
showed that a decrease in mRNA levels of SOD-2, CAT, and GPX-1
compared to wild-type fibroblasts and mice with loss of PGC-1α were
more vulnerable to oxidative stress (18). In addition, PGC-1α may increase the
uncoupling capacity and concomitantly reduce mitochondrial ROS
production. Furthermore, PGC-1α promotes the expression of SIRT-3
by interacting with an ER-α binding element mapped to the SIRT3
promoter region. SIRT3 can activate SOD-2 through a
post-translational mechanism (40).
Taken together, the results show that, PGC-1α has a role in
reducing the expression and activity of ROS damage by upregulating
antioxidant factors. As PGC-1α exerts its roles by modulating the
metabolism of non-malignant cells, the role of PGC-1α in tumors is
important (43–45). In our study, cells cultured under
high-glucose conditions showed a higher level of PGC-1α, and these
cells showed a higher level of SOD-2, CAT and GPX-1 during
chemotherapy than cells cultured in ordinary RPMI-1640 medium,
supporting the hypothesis that PGC-1α maintains the role of
regulating antioxidant factors in MM. Furthermore, we suppressed
the expression of PGC-1α by siRNA and these cells showed a
decreased expression of SOD-2, CAT and GPX-1, accompanied with an
increased ROS and enhanced toxicity of chemotherapeutic agents.
Thus, PGC-1α should be responsible for the decreased efficacy of
chemotherapeutic agents in MM, and it may be effective to improve
efficacy by targeting PGC-1α.
In conclusion, though this is an in vitro
study and there are several limitations, our study is sufficient to
show that hyperglycemia attenuates the anti-MM effect of
chemotherapeutic agents by upregulating PGC-1α and related
antioxidant factors, leading to the reduction of ROS. Treatment of
hyperglycemia holds may be benefiical for patients with MM and
diabetes or drugs induced hyperglycemia. Targeting PGC-1α may be
sufficient in improving the efficacy of chemotherapy in cancer
treatment.
References
|
1
|
Guglielmelli T and Palumbo A:
Incorporating novel agents in the management of elderly myeloma
patients. Curr Hematol Malig Rep. 8:261–269. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Palumbo A and Mina R: Management of older
adults with multiple myeloma. Blood Rev. 27:133–142. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chou YS, Yang CF, Chen HS, et al:
Pre-existing diabetes mellitus in patients with multiple myeloma.
Eur J Haematol. 89:320–327. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Issa ZA, Zantout MS and Azar ST: Multiple
myeloma and diabetes. ISRN Endocrinol. 2011:8150132011.
|
|
5
|
Jung SH, Jang HC, Lee SS, et al: The
impact of hyperglycemia on risk of severe infections during early
period of induction therapy in patients with newly diagnosed
multiple myeloma. Biomed Res Int. 2014:4131492014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ahmed YA and Eltayeb A: Clinical
challenges: myeloma and concomitant type 2 diabetes. Int J Hematol
Oncol Stem Cell Res. 7:34–41. 2013.
|
|
7
|
Wu W, Merriman K, Nabaah A, et al: The
association of diabetes and anti-diabetic medications with clinical
outcomes in multiple myeloma. Br J Cancer. 111:628–636. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chiu BC, Gapstur SM, Greenland P, Wang R
and Dyer A: Body mass index, abnormal glucose metabolism, and
mortality from hematopoietic cancer. Cancer Epidemiol Biomarkers
Prev. 15:2348–2354. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Baynes JW: Role of oxidative stress in
development of complications in diabetes. Diabetes. 40:405–412.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Busik JV, Mohr S and Grant MB:
Hyperglycemia-induced reactive oxygen species toxicity to
endothelial cells is dependent on paracrine mediators. Diabetes.
57:1952–1965. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zorov DB, Juhaszova M and Sollott SJ:
Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS
release. Physiol Rev. 94:909–950. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Edeas M: Strategies to target mitochondria
and oxidative stress by antioxidants: key points and perspectives.
Pharm Res. 28:2771–2779. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Vurusaner B, Poli G and Basaga H: Tumor
suppressor genes and ROS: complex networks of interactions. Free
Radic Biol Med. 52:7–18. 2012. View Article : Google Scholar
|
|
14
|
Valle I, Alvarez-Barrientos A, Arza E,
Lamas S and Monsalve M: PGC-1alpha regulates the mitochondrial
antioxidant defense system in vascular endothelial cells.
Cardiovasc Res. 66:562–573. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nishikawa T, Edelstein D, Du XL, et al:
Normalizing mitochondrial superoxide production blocks three
pathways of hyperglycaemic damage. Nature. 404:787–790. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Finck BN and Kelly DP: PGC-1 coactivators:
inducible regulators of energy metabolism in health and disease. J
Clin Invest. 116:615–622. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cao D, Zhou H, Zhao J, et al: PGC-1α
integrates glucose metabolism and angiogenesis in multiple myeloma
cells by regulating VEGF and GLUT-4. Oncol Rep. 31:1205–1210.
2014.PubMed/NCBI
|
|
18
|
St-Pierre J, Drori S, Uldry M, et al:
Suppression of reactive oxygen species and neurodegeneration by the
PGC-1 transcriptional coactivators. Cell. 127:397–408. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kong X, Wang R, Xue Y, et al: Sirtuin 3, a
new target of PGC-1 alpha, plays an important role in the
suppression of ROS and mitochondrial biogenesis. PLoS One.
5:e117072010. View Article : Google Scholar
|
|
20
|
Fujisawa K, Nishikawa T, Kukidome D, et
al: TZDs reduce mitochondrial ROS production and enhance
mitochondrial biogenesis. Biochem Biophys Res Commun. 379:43–48.
2009. View Article : Google Scholar
|
|
21
|
Zhang LN, Zhou HY, Fu YY, et al: Novel
small-molecule PGC-1α transcriptional regulator with beneficial
effects on diabetic db/db mice. Diabetes. 62:1297–1307. 2013.
View Article : Google Scholar :
|
|
22
|
Pearce EL, Walsh MC, Cejas PJ, et al:
Enhancing CD8 T-cell memory by modulating fatty acid metabolism.
Nature. 460:103–107. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Villarreal-Garza C, Shaw-Dulin R,
Lara-Medina F, et al: Impact of diabetes and hyperglycemia on
survival in advanced breast cancer patients. Exp Diabetes Res.
2012:7320272012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Luo J, Chen YJ and Chang LJ: Fasting blood
glucose level and prognosis in non-small cell lung cancer (NSCLC)
patients. Lung Cancer. 76:242–247. 2012. View Article : Google Scholar
|
|
25
|
Friday E, Ledet J and Turturro F: Response
to dexamethasone is glucose-sensitive in multiple myeloma cell
lines. J Exp Clin Cancer Res. 30:812011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Scarpulla RC, Vega RB and Kelly DP:
Transcriptional integration of mitochondrial biogenesis. Trends
Endocrinol Metab. 23:459–466. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sheean PM, Kilkus JM, Liu D, Maciejewski J
and Braunschweig CA: Incident hyperglycemia, parenteral nutrition
administration and adverse outcomes in patients with myeloma
admitted for initial auto-SCT. Bone Marrow Transplant.
48:1117–1122. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
McBrayer SK, Cheng JC, Singhal S, Krett
NL, Rosen ST and Shanmugam M: Multiple myeloma exhibits novel
dependence on GLUT4, GLUT8, and GLUT11: implications for glucose
transporter-directed therapy. Blood. 119:4686–4697. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhao Y, Butler EB and Tan M: Targeting
cellular metabolism to improve cancer therapeutics. Cell Death Dis.
4:e5322013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gitenay D, Wiel C, Lallet-Daher H, et al:
Glucose metabolism and hexosamine pathway regulate oncogene-induced
senescence. Cell Death Dis. 5:e10892014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhao Y, Liu H, Riker AI, et al: Emerging
metabolic targets in cancer therapy. Front Biosci (Landmark Ed).
16:1844–1860. 2011. View
Article : Google Scholar
|
|
32
|
DeBerardinis RJ, Mancuso A, Daikhin E, et
al: Beyond aerobic glycolysis: transformed cells can engage in
glutamine metabolism that exceeds the requirement for protein and
nucleotide synthesis. Proc Natl Acad Sci USA. 104:19345–19350.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhao Y, Altman BJ, Coloff JL, et al:
Glycogen synthase kinase 3alpha and 3beta mediate a
glucose-sensitive antiapoptotic signaling pathway to stabilize
Mcl-1. Mol Cell Biol. 27:4328–4339. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Danial NN, Gramm CF, Scorrano L, et al:
BAD and glucokinase reside in a mitochondrial complex that
integrates glycolysis and apoptosis. Nature. 424:952–956. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Rathmell JC, Fox CJ, Plas DR, Hammerman
PS, Cinalli RM and Thompson CB: Akt-directed glucose metabolism can
prevent Bax conformation change and promote growth
factor-independent survival. Mol Cell Biol. 23:7315–7328. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Mathupala SP, Ko YH and Pedersen PL:
Hexokinase II: cancer’s double-edged sword acting as both
facilitator and gatekeeper of malignancy when bound to
mitochondria. Oncogene. 25:4777–4786. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gendron MC, Schrantz N, Metivier D, et al:
Oxidation of pyridine nucleotides during Fas- and ceramide-induced
apoptosis in Jurkat cells: correlation with changes in
mitochondria, glutathione depletion, intracellular acidification
and caspase 3 activation. Biochem J. 353:357–367. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Vu K, Busaidy N, Cabanillas ME, et al: A
randomized controlled trial of an intensive insulin regimen in
patients with hyperglycemic acute lymphoblastic leukemia. Clin
Lymphoma Myeloma Leuk. 12:355–362. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Puigserver P, Wu Z, Park CW, Graves R,
Wright M and Spiegelman BM: A cold-inducible coactivator of nuclear
receptors linked to adaptive thermogenesis. Cell. 92:829–839. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kang C and Li Ji L: Role of PGC-1α
signaling in skeletal muscle health and disease. Ann NY Acad Sci.
1271:110–117. 2012. View Article : Google Scholar
|
|
41
|
Radak Z, Zhao Z, Koltai E, Ohno H and
Atalay M: Oxygen consumption and usage during physical exercise:
the balance between oxidative stress and ROS-dependent adaptive
signaling. Antioxid Redox Signal. 18:1208–1246. 2013. View Article : Google Scholar :
|
|
42
|
Wenz T: Regulation of mitochondrial
biogenesis and PGC-1α under cellular stress. Mitochondrion.
13:134–142. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Salem AF, Whitaker-Menezes D, Howell A,
Sotgia F and Lisanti MP: Mitochondrial biogenesis in epithelial
cancer cells promotes breast cancer tumor growth and confers
autophagy resistance. Cell Cycle. 11:4174–4180. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tennakoon JB, Shi Y, Han JJ, et al:
Androgens regulate prostate cancer cell growth via an
AMPK-PGC-1α-mediated metabolic switch. Oncogene. 33:5251–5261.
2014. View Article : Google Scholar
|
|
45
|
Girnun GD: The diverse role of the PPARγ
coactivator 1 family of transcriptional coactivators in cancer.
Semin Cell Dev Biol. 23:381–388. 2012. View Article : Google Scholar : PubMed/NCBI
|