Introduction
TXNIP (also known as vitamin D3-upregulated
protein or thioredoxin-interacting protein), was originally
isolated in HL60 leukemia cells treated with 1,25-dihydroxyvitamin
D3 (1). TXNIP has
subsequently been identified as a key modulator of the redox
system. It binds to the active cysteine residue of thioredoxin
(TRX) and inhibits its antioxidative function (2). In addition, TXNIP can act
independently of its binding to TRX, contributing to inhibition of
cell growth by arresting the domain-mediated suppression of glucose
uptake and metabolic reprogramming (3,4).
TXNIP is important in cell growth and cancer,
acting as a tumor suppressor gene. Downregulation or loss of
TXNIP expression has been shown in various tumors including
renal, breast, lung, gastric, colon and hepatocellular carcinoma
(5). Genetic alterations of
TXNIP are not common, and its aberrant expression in cancer
has been mainly regulated via post-transcriptional and
translational mechanisms. It seems that cancer cells develop
various ways, such as hypermethylation, histone deacetylation,
histone methylation and post-translational inhibition by miRNA, to
inactivate TXNIP expression, indicating the importance of
its repression in tumorigenesis (6).
Altered TXNIP expression may be associated
with disease progression and poor prognosis (7–9). The
role of TXNIP in breast cancer has been previously
investigated. A lower expression of TXNIP mRNA was identified in
breast cancer tumors as compared to matched normal controls
(10). Cadenas et al
demonstrated that a low TXNIP expression was associated with
worse prognosis in breast cancer, while a higher expression of
TXNIP was associated with longer metastasis-free interval
(7). In terms of breast cancer cell
biology, Butler et al demonstrated that ectopic expression
of TXNIP in MCF7 cells induced cell senescence (10).
Studies on stem cells led to the identification of
paused genes (11). These
genes are characterized by the contemporary presence of activator
and repressor epigenetic markers (bivalent marking). These markers
are based on histone post-translational modifications; for example,
lysine 4 trimethylation of H3 histone (H3K4me3) was associated with
transcriptional activation, while lysine 27 trimethylation of H3
histone (H3K27me3) was associated with transcriptional silencing
(12,13). Thus, a bivalent marking is typical
of paused genes, i.e., genes that are expressed at low
levels but that can be expressed at high levels following
differentiation or by external stimulation. By contrast, genes in
which only markers of transcriptional silencing are detected are
definitively silenced.
The epigenetic concept of paused genes has
been generated by studying epigenetic marking and gene expression
during differentiation of stem cells (11). However, if the bivalent marking
hypothesis of paused genes is true for the gene expression
of cancer cells, it may be useful for delineation of new
therapeutic strategies. For example, the expression of
paused oncosuppressor genes may be reactivated by external
stimulation more easily than the expression of oncosuppressor genes
that are definitively silenced.
Treatment of breast cancer is challenging as its
heterogeneity may be due to the differential expression of estrogen
receptor (ER), progesterone receptor (PR) and amplification of
HER2/neu. Therapeutic options are significantly affected by the
expression of the three markers (14). Triple-negative breast cancer (TNBC),
characterized by the absence of ER, PR and
HER-2 gene expression, is an important clinical challenge
because it does not respond to endocrine or monoclonal antibody
anti HER-2 therapies.
In the present study, the hypothesis that epigenetic
bivalent marking defines the possibility for re-activation of
TXNIP gene expression was evaluated in TNBC-derived cell
lines.
Materials and methods
Cell lines and immunohistochemistry
MDA157 and MDA468 human cell lines derived from
triple negative breast cancer, were grown in DMEM medium
supplemented with 10% fetal bovine serum (Gibco Invitrogen, Milan,
Italy), 2 mM L-glutamine (EuroClone, Milan, Italy) and 50 mg/ml
gentamicin (Gibco Invitrogen) in a humidified incubator (5%
CO2 in air at 37°C).
Core biopsies of triple negative breast tumors were
examined. The tissues were formalin-fixed for 16–24 h.
Formalin-fixed paraffin tissue sections (5 μm) mounted on
Superfrost slides (Surgipath, Richmond, IL, USA) were
immunohistochemically stained, by using the peroxidase/DAB Plus
Dako Real EnVision™ detection system (Dako A/S, Glostrup, Denmark).
Antigen retrieval was performed in a water bath at 98°C with 0.01 M
citrate buffer, at pH 6.0 for 40 min. Endogenous peroxidase
activity was blocked by incubation in the Peroxidase Block solution
(Dako A/S) for 10 min. Primary rabbit polyclonal antiserum to TXNIP
(Abcam, Cambridge, UK) diluted at 1:100 was applied and incubated
for 60 min at room temperature. After being washed, the slides were
incubated with the peroxidase/DAB Plus Dako Real EnVision™
detection system (Dako) according to the manufacturer’s
instructions. For reaction visualization, 3,3′-diaminobenzidine
tetrahydrochloride was used as chromogen. The sections were
counterstained with Mayer hematoxylin. Normal breast tissue
sections were used as a positive control, whereas the negative
control was performed by replacing the primary antibody with PBS.
Nuclear and cytoplasmic immunostaining were evaluated by using
light microscopy, in which the entire section was scanned at
high-power magnification (×400). In the case of cell cultures,
MDA157 cells, treated or not with SAHA 3 μM and PJ34 5
μM alone or in combination, were immediately fixed with
ethanol containing 5% glacial acetic acid for 10 min at 4°C, and
rinsed with 0.1% saponin (Sigma Chemical Co., St. Louis, MO, USA)
in phosphate-buffered saline (PBS). This PBS-saponin solution was
also used for all subsequent washing steps. The cultures were
incubated overnight at 4°C with rabbit antiserum to TXNIP, diluted
at 1:100. After washing the Dako Real EnVision™ detection system
kit, peroxidase/DAB+, rabbit/mouse K5007 (Dako) was
used. Peroxidase activity was detected with 3,3′-diaminobenzidine
tetrahydrochloride followed by haematoxylin counterstaining.
Chromatin immunoprecipitation (ChIP)
In order to perform ChIP assay, MDA157 and MDA468
cells treated or not for 72 h with SAHA 3 μM and PJ34 5
μM, were subjected to cross-linking by using 1% formaldehyde
(10 min at 37°C). The cells were then scraped in PBS with
proteinase inhibitors and resuspended in cell lysis buffer (1% SDS,
10 mM EDTA, 50 mM Tris-HCl pH 8.1 and protease inhibitors). The
samples were subjected to sonication and diluted 10-fold with
dilution buffer (0.01% SDS, 1.1% Triton X-100, 1.2 mM EDTA, 16.7 mM
Tris-HCl pH 8.1, 167 mM NaCl); 8% of this was saved as total
input.
The samples were incubated for preclearing with
Dynabeads Protein A (Gibco Invitrogen). For immunoprecipitation, th
esamples were incubated overnight with 10 μg of rabbit
polyclonal anti-acetyl-histone H3 antibody (Upstate Temecula, CA,
USA) or anti-histone H3 trimethyl Lys4 antibody or anti-histone H3
trimethyl Lys27 antibody (both from Active Motif) or rabbit
polyclonal anti-histone H3 trimethyl K9 antibody (Abcam). The
following day, samples and negative controls (samples with purified
Rabbit IgG, Millipore) were added with Dynabeads Protein A.
After being washed, the immunocomplexes were eluted
from beads with elution buffer (1% SDS, 0.1 M, 50 mM
NaHCO3), crosslinks were reverted by heating and the
samples were treated with proteinase K. DNA was purified with
phenol/chloroform extraction followed by ethanol precipitation,
which was used as a template in quantitative absolute PCR, whose
reaction primers are presented in Table
I. After quantitative PCR reactions, the acetylated or
methylated H3 levels were determined as a ratio of signals recorded
after and prior to (input) immunoprecipitation.
 | Table IPrimers used for real-time PCR
analysis. |
Table I
Primers used for real-time PCR
analysis.
| Gene name | Forward | Reverse | Probe |
|---|
| TXNIP |
GTCAGTCACTCTCAGCCATAGCA |
CACACTTTCTGGCTGTAATAACTCTCA | |
| β-actin |
TTGTTACAGGAAGTCCCTTGCC |
ATGCTATCACCTCCCCTGTGTG | |
| TXNIP |
| promoter |
GAGCGCAACAACCATTTTCC |
GAGCCCGACCAATCAGTGA |
FAM-TGTCCACGCGCCACAGCGAT |
Quantitative RT-PCR
Total RNA from cell lines, treated for 72 h with
SAHA 3 μM, PJ34 5 μM alone or in combination, were
extracted with an RNeasy mini kit, according to the manufacturer’s
instructions (Qiagen, Hilden, Germany). Total RNA (500 ng) was
reverse transcribed to cDNA using random exaprimers and MMLV
reverse transcriptase (Invitrogen). Quantitative PCRs were
performed using Platinum SYBR-Green qPCR supermix (Life
Technologies) or TaqMan Universal PCR master mix (Applied
Biosystems, Foster City, CA, USA) with the ABI Prism 7300 sequence
detection systems (Applied Biosystems). The ΔΔ Ct method, by means
of the SDS software (Applied Biosystems), was used to calculate the
mRNA levels. Oligonucleotide primers for TXNIP and for the
endogenous control, β-actin, were purchased from Sigma (Table I).
Protein extraction and western
blotting
In order to detect the TXNIP protein, total protein
extraction was performed. To this purpose, MDA157 and MDA468 cells,
cultured for 72 h in growth medium, in presence or absence of SAHA
3 μM, PJ34 5 μM alone or in combination, were
harvested by scraping and lysed with Laemmli/SDS lysis buffer (1.0
M Tris, pH 6.8, 2% SDS, 0.001% bromophenol blue, 10% glycerol, 2%
β-ME). The samples were then subjected to sonication.
For the western blot analysis, the proteins were
electrophoresed on 10% SDS-PAGE and then transferred to
nitrocellulose membranes, which were saturated with 5% non-fat
dried milk in PBS/0.1% Tween-20. The membranes were then incubated
overnight with rabbit polyclonal anti-TXNIP antibody or
anti-β-actin antibody (both from Abcam). The following day, the
membranes were incubated for 2 h with anti-rabbit immunoglobulin
coupled with peroxidase (Sigma-Aldrich). The blots were developed
using Chemidoc XRS (Bio-Rad, Hercules, CA, USA) with the
chemiluminescence procedure (Amersham Biosciences, Buckinghamshire,
UK).
Cell viability and apoptosis assays
To test cell viability the MTT assay was used as
previously described (15). The
cells were seeded in 96-well plates in 200 μl medium. The
following day, the growth medium was replaced with fresh medium
(untreated cultures) or with medium containing SAHA 3 μM
(Cayman Chemical, Ann Harbor, MI, USA) and PJ34 5 μM (Merck
Chemicals Ltd., Nottingham, UK) alone or in combination, and the
plates were incubated for 72 h. Experimental points were run in
quadruplicate.
Caspase 3/7 activity was measured as a marker of
apoptosis using the ApoOne Homogeneous Caspase 3/7 assay kit
(Promega, Milan, Italy) according to the manufacturer’s
instructions. Experimental points were run in triplicate.
Statistical analysis
TXNIP mRNA and protein levels, cell viability
and apoptosis levels were expressed as the mean ± SD values, and
significances were analyzed with the t-test performed with GraphPad
software for science (San Diego, CA, USA).
Results
Immunohistochemical detection of TXNIP in
normal breast tissue and triple-negative breast cancer
Identification of specific therapeutic options for
TNBC remain a challenge. Therefore, we focused our investigation on
this type of cancer. TNBC tissues were immunostained and, despite a
certain degree of heterogeneity, a decrease of TXNIP expression was
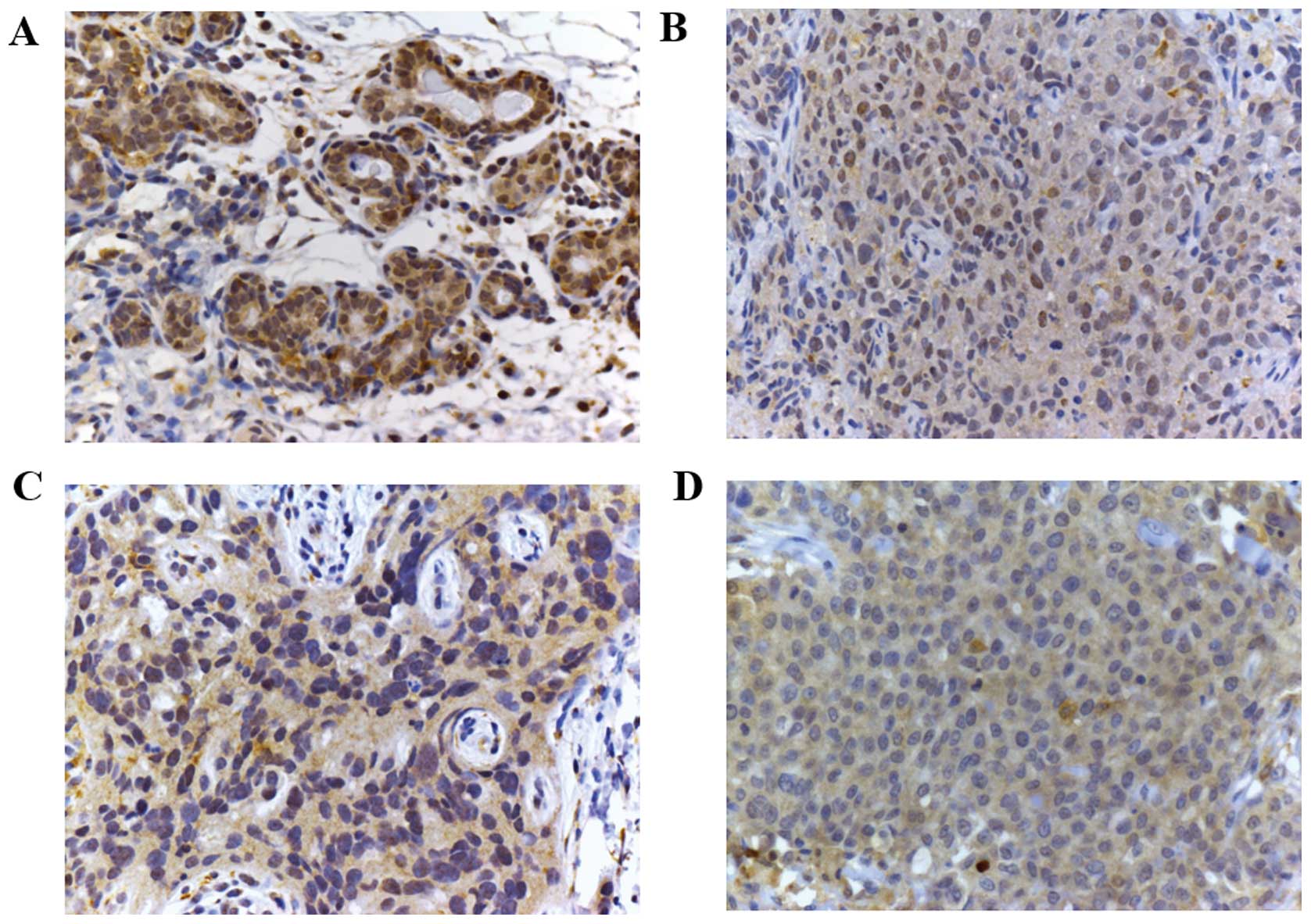
detected. Fig. 1 shows some
exemplifying cases: compared to normal breast tissue (A), TNBC
shows a weak cytoplasmic signal (B–D). However, in the majority of
cells no nuclear staining was identified (C and D).
Histone post-translational modifications
on TXNIP promoter
In order to evaluate the bivalent marking
hypothesis, we used the MDA157 and MDA468 TNBC-derived cell lines.
Histone modifications associated with transcriptional activation or
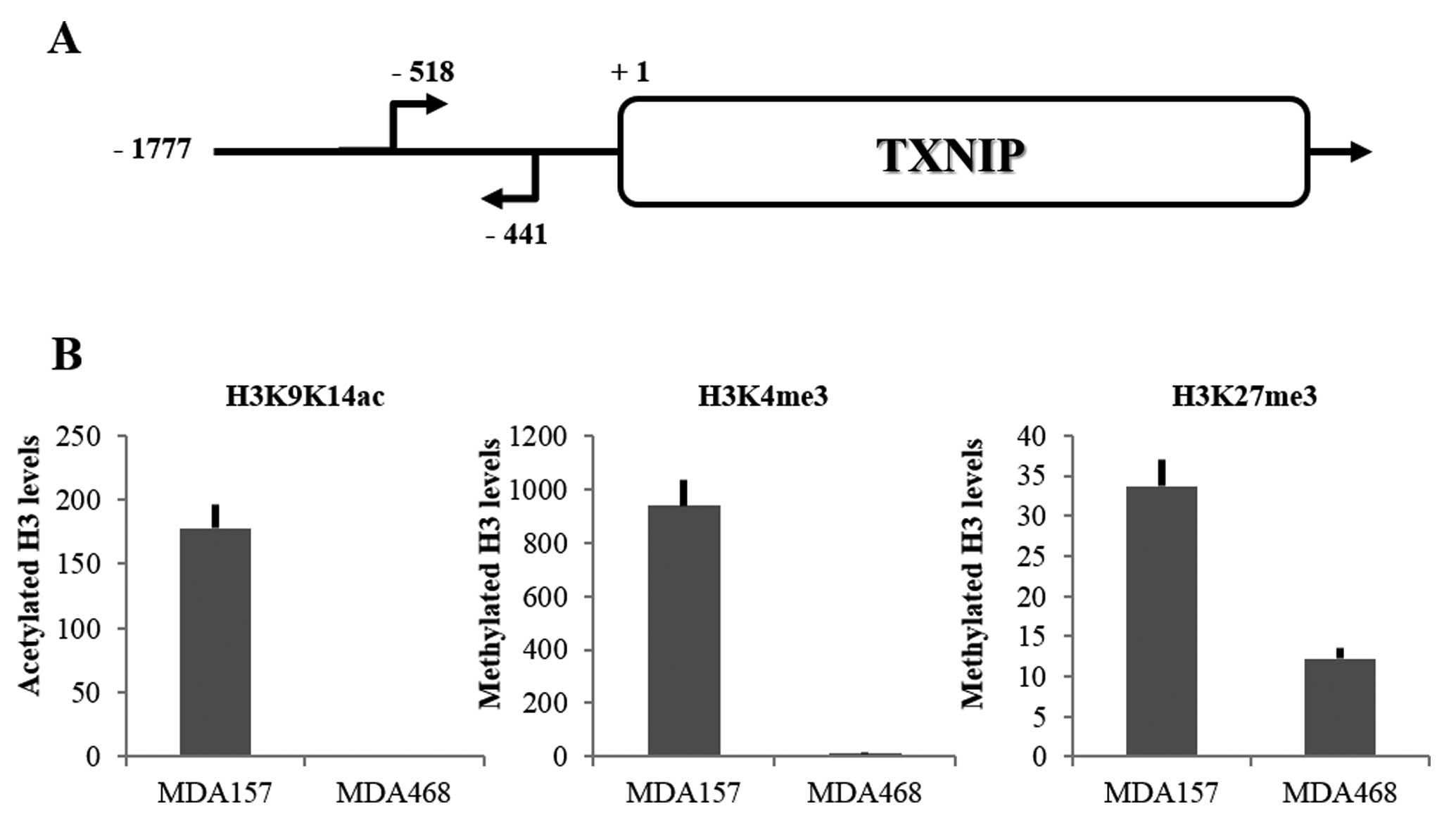
silencing were evaluated on TXNIP promoter (Fig. 2A). By using ChIP, three histone
modifications were investigated: two associated to transcriptional
activation, histone H3 lysines 9–14 acetylation (H3K9K14ac) and
histone H3 lysine 4 trimethylation (H3K4me3), and one associated
with transcriptional silencing, histone H3 lysine 27 trimethylation
(H3K27me3) (12,13). As shown in Fig. 2B, a difference was identified
between the two cell lines: in MDA157 all three markers were
detected, whereas in MDA468 only H3K27me3 was detectable. Thus, in
MDA157 cells the bivalent marking was observed in the promoter of
TXNIP gene, suggesting that its expression was prone to be
activated. Instead, in MDA468 cells only the marker associated with
gene silencing was detectable in the TXNIP promoter,
suggesting that transcription of this gene was totally blocked. The
basal levels of global histone modifications (evaluated by western
blotting) in MDA157 and MDA468 cells has been previously identified
(16). When results shown in
Fig. 2B were compared to those
data, differences of H3K9K14ac, H3K4me3 and H3K27me3 observed in
the TXNIP promoter were not due to differences of global
levels of these modifications. In particular, global levels of
H3K27me3 were much lower in MDA468 than in MDA157. Therefore, in
terms of relative amount, marker levels associated with gene
silencing (H3K27me3) in MDA468 were much higher than those in
MDA157.
Effects of SAHA and PJ34 on TXNIP gene
expression and histone post-translational modifications
To evaluate the possibility of activating
TXNIP gene expression in MDA157 we used two different
compounds. The first one was the histone deacetylase (HDAC)
inhibitor suberoylanilide hydroxamic acid (SAHA), already
FDA-approved for the treatment of several neoplastic diseases
(17,18), which is able to induce TXNIP
expression in transformed cells, including solid tumors and
leukemia, but not in normal cells (19,20).
The second compound was PJ34, a poly-ADP-ribose polymerases (PARP)
inhibitor. PARP is involved in numerous processes that are vital
for cells. In particular, PARP proteins are activated by the
presence of DNA damage, leading to poly-ADP-ribosylation of
proteins involved in DNA repair, genome integrity, regulation of
transcription, proliferation and apoptosis (21). In Fig.
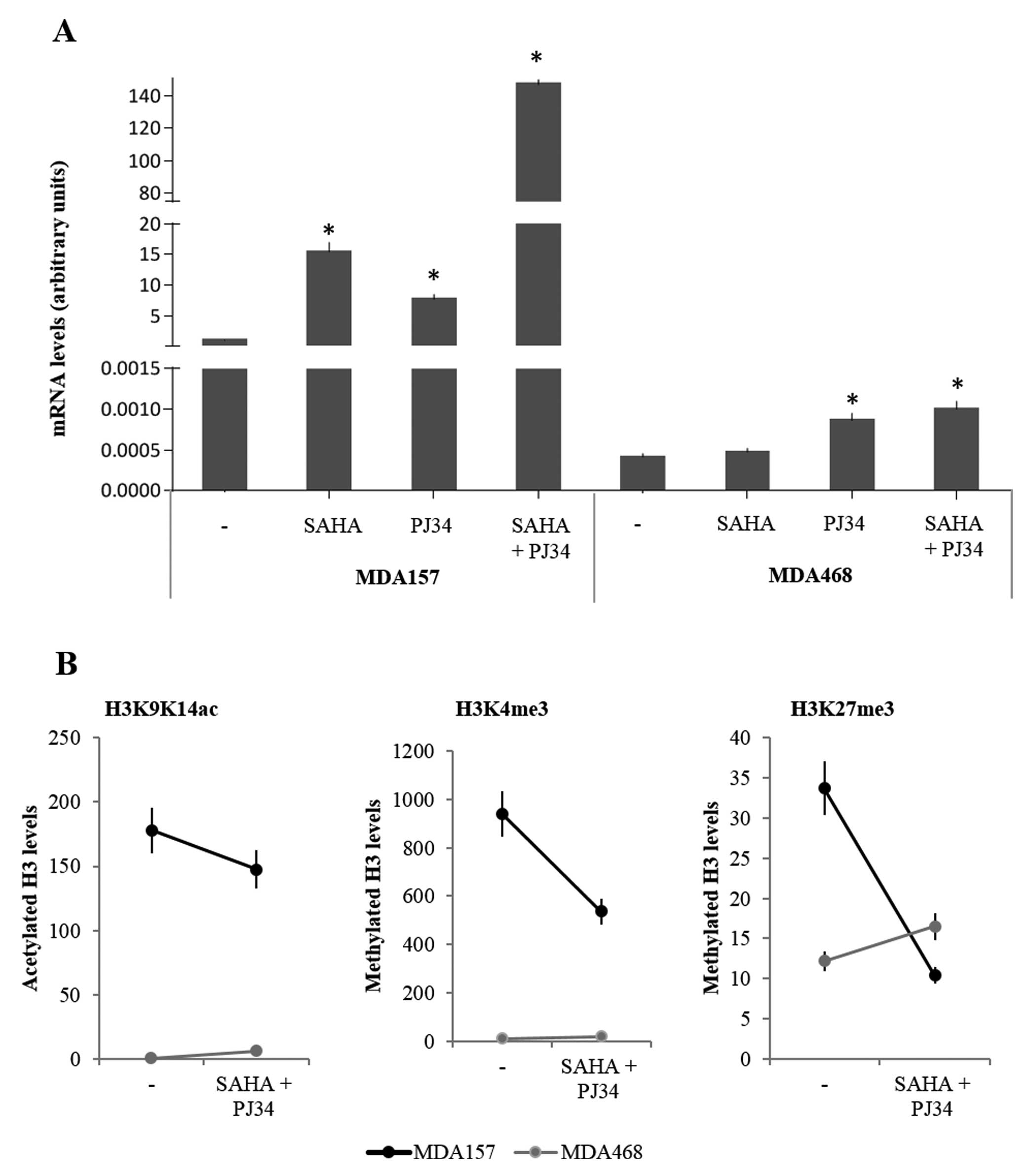
3A, the effects of either compound on TXNIP gene
expression are shown. Basal levels of TXNIP mRNA levels were
higher in MDA157 than in MDA468 cells. In MDA157 cells, treatment
with SAHA and PJ34 induced an increase of TXNIP mRNA levels
of 15- and 10-fold, respectively. When, in this cell line, the two
compounds were used together, a marked increase of TXNIP
mRNA levels was observed (140-fold over basal values). Minimal
effects were instead observed in MDA468 cells both with single
compounds and in combination. In this cell line, TXNIP mRNA values
remained markedly below the basal values observed in MDA157 cells.
Thus, only the paused cell line (MDA157) was responsive to
treatment with SAHA and PJ34 alone and synergy was observed when
the compounds were employed in combination. By contrast, MDA468
TXNIP gene expression appeared almost insensitive to
treatments, with only PJ34 having a very modest effect.
Subsequent to the synergy observed in MDA157 cells,
we tested whether the combined use of SAHA and PJ34 modified
histone post-translational modifications at the TXNIP
promoter level. As shown in Fig.
3B, a decrease for all three post-translational modifications
was observed in the MDA157 cells. However, this decrease was
minimal for H3 acetylation but marked for H3K27me3. In MDA468,
either marker associated with transcriptional activation remained
almost undetectable (H3K9K14ac or H3K4me3), while the marker
associated with silencing was increased. Thus, the combination of
SAHA and PJ34 did not erase the silencing epigenetic signature in
MDA468 cells.
Effects of SAHA and PJ34 on TXNIP protein
level
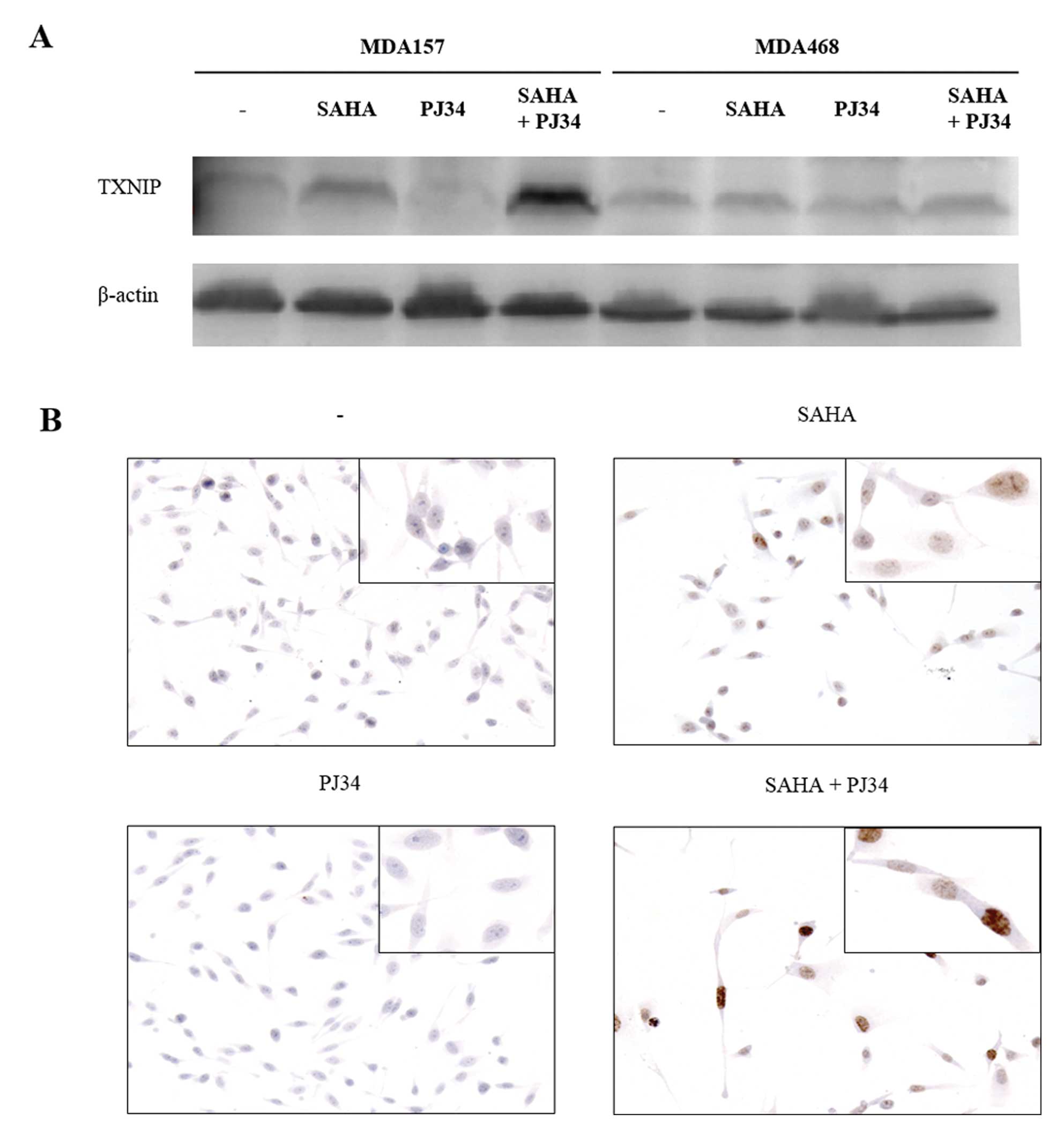
To verify whether the TXNIP mRNA increase was
associated with enhancement of TXNIP protein expression, a western
blot analysis was performed. As shown in Fig. 4A, the MDA468 cell line did not
express TXNIP protein in detectable amounts, either in basal
conditions or after stimulation with SAHA and PJ34 alone or in
combination. The MDA157 cell line showed an extremely low basal
TXNIP protein expression, which increased following SAHA treatment
and this increment was higher with the combination of SAHA and
PJ34. These findings were also supported by immunohistochemical
analysis of MDA157 cells. In Fig.
4B, the TXNIP protein shows nuclear localization, which was
weakly incremented following SAHA treatment, while no variation was
observed with PJ34. The combined addition of SAHA and PJ34 markedly
increased TXNIP protein levels.
Effects of HDAC and PARP inhibitors on
cell viability and apoptosis in MDA157 and MDA468 cells
In various tumor tissues, the increase of TXNIP
expression was correlated with growth inhibition and induction of
apoptosis (19,22). Therefore, we examined whether such a
correlation is present in our experimental system. We evaluated
whether SAHA and PJ34 cocktail may have a biological effect in
terms of cell proliferation and apoptosis. In particular, cell
viability was assessed in the two lines after treatment with SAHA
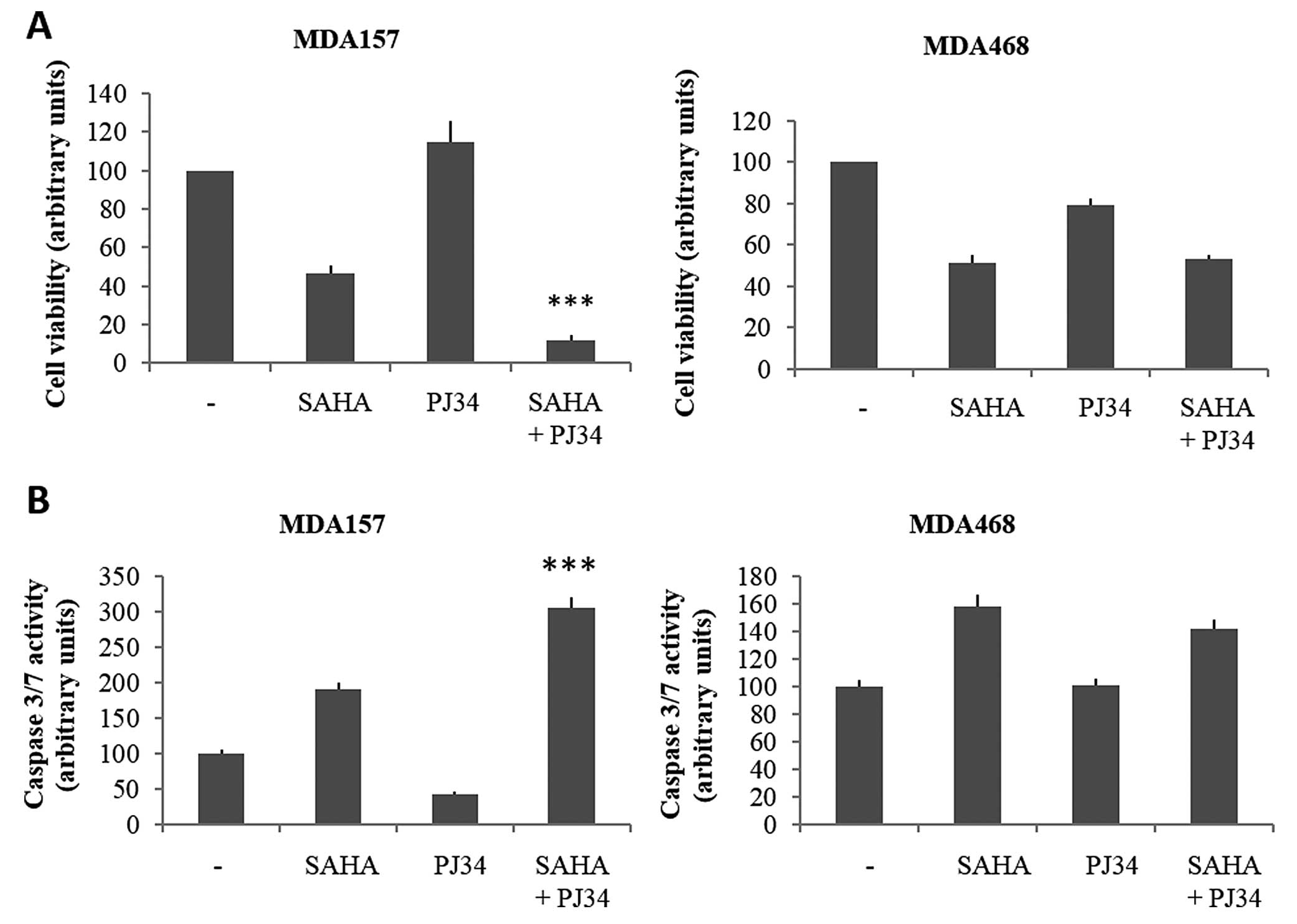
and PJ34 alone or in combination, for 72 h (Fig. 5A). In the two cell lines, the
treatment with SAHA alone determines a moderate decrease in
viability compared to untreated cells. The effect of PJ34, however,
appeared to be limited in the two lines, with a slight increase and
decrease in MDA157 and MDA468 cell viability, respectively. The
combination of the two compounds shows a decrease in growth
compared to untreated cells, which was significantly higher in
MDA157 than in MDA468. In the latter cell line, treatment with SAHA
alone or in combination with PJ34 affected cell proliferation in a
comparable manner.
To evaluate whether effects of the two drugs on cell
viability were associated with modification of apoptosis, the
caspase 3/7 activity was assessed (Fig.
5B). The MDA157 cell line exhibited an almost 2-fold increase
in caspase 3/7 activity after 72 h SAHA treatment, and a 2.5-fold
after SAHA and PJ34 treatment, while a slight reduction following
treatment with PJ34 alone was observed. In the MDA468 cell line, a
mild increase in apoptosis with SAHA alone or in combination with
PJ34 was observed, while treatment with PJ34 alone produced no
detectable effects. Therefore, treatment with SAHA and PJ34 in
combination determines an increase in apoptotic processes in MDA157
but not in MDA468 cells.
Taken together, these results suggested that cells
that are responsive to the SAHA and PJ34 combination, showing an
increase of TXNIP protein levels, were also responsive in terms of
cell growth parameters.
Discussion
For many years, cancer origin and progression have
been recognized to be secondary to the accumulation of genetic
mutations that may result in oncogene activation and tumor
suppressor gene inactivation. The recent finding that reversible
alterations in histone proteins and DNA can also lead to
tumorigenesis has introduced a novel term in the field of cancer
research known as ‘epigenetics’ (23). The importance of cancer epigenetics
has led many investigators to incorporate this novel and noteworthy
field in anticancer drug development. Several drugs that target
epigenetic alterations, including inhibitors of HDAC and DNA
methyltransferase (DNMT), are currently approved for the treatment
of hematological malignancies and are available for clinical
investigation in solid tumors (23). Epigenetic mechanisms may play a role
in TXNIP gene expression. It appears that in cancer cells
TXNIP expression is downregulated through epigenetic
mechanisms, such as deacetylation and hypermethylation at the
promoter level (6). Our data
indicate that epigenetic marking on TXNIP gene in MDA157 and
MDA468 cell lines is different. Histone post-translational
modifications are an important regulatory platform for gene
expression, replication, DNA repair, chromosome segregation and
cell death (24). Specific histone
modifications are associated with gene expression activation or
silencing. Therefore, we investigated histone H3 lysine 4
trimethylation (H3K4me3), and histone H3 lysines 9 and 14
acetylation (H3K9K14ac), which are markers of active transcription,
and histone H3 lysine 27 trimethylation (H3K27me3), which instead
characterizes silenced genes. In MDA157 cells basal levels of
H3K9K14ac, H3K4me3 and H3K27me3 are measurable. This signature
indicates that the TXNIP gene, in these cells, was paused,
and likely to be activated by exogenous stimulation. Treatment with
SAHA and PJ34 reduced the levels of H3K27me3, while maintaining
high levels of H3 acetylation, resulting in an increased
TXNIP expression. Conversely, in MDA468 cells, the
TXNIP gene was definitely silenced, with significant levels
of H3K27me3 being only observable in the basal state and increased
following the combined treatment of SAHA and PJ34. Thus, we showed
that the model of bivalent marking may accurately predict the cell
responsiveness of TXNIP gene following the combined
treatment of SAHA and PJ34. In MDA157 cells the pausing
signature was permissive for the synergy between SAHA and PJ34.
Epigenetic profiling of cancer cells may reveal oncosuppressor
genes that are paused and that, therefore, may be activated
by pharmacological treatment.
The synergy between SAHA and PJ34 observed in MDA157
cells was of great interest in terms of cancer treatment. A central
issue in the development of anticancer compounds was to increase
the therapeutic index, limiting at the same time, the development
of resistance. One solution was to combine multiple drugs that act
synergistically. A number of ongoing clinical trials are
investigating the effects of combined therapy against different
types of cancer (25–27). In our experimental system, the
effect of combined treatment with SAHA and PJ34 on TXNIP
expression was associated with variation in cell viability and
apoptosis. In the MDA157 cell line, the increase of TXNIP
expression was associated with a decrease in cell viability and an
increase in apoptosis. By contrast, MDA468 cells were not
responsive to the SAHA-PJ34 cocktail, both in terms of TXNIP
expression and cell proliferation parameters. These findings are
consistent with those of previous studies, in which a correlation
between TXNIP expression increment, growth inhibition and induction
of apoptosis has been shown (19,22).
Moreover, Jasek et al have recently shown that the combined
treatment of SAHA and PJ34 on leukemic cell lines has a synergistic
effect on proliferation inhibition and an increase in apoptosis
(28). Thus, our data support the
hypothesis that TXNIP is an effective target for the
treatment of breast cancer.
Acknowledgments
This study is funded by grants to GD from
Associazione Italiana per la Ricerca sul Cancro (AIRC) (project no.
IG 10296).
References
|
1
|
Chen KS and DeLuca HF: Isolation and
characterization of a novel cDNA from HL-60 cells treated with
1,25-dihydroxyvitamin D-3. Biochim Biophys Acta. 1219:1226–1232.
1994.
|
|
2
|
Kaimul AM, Nakamura H, Masutani H and
Yodoi J: Thioredoxin and thioredoxin-binding protein-2 in cancer
and metabolic syndrome. Free Radic Biol Med. 43:861–868. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Elgort MG, O’Shea JM, Jiang Y and Ayer DE:
Transcriptional and translational downregulation of thioredoxin
interacting protein is required for metabolic reprogramming during
G(1). Genes Cancer. 1:893–907. 2010. View Article : Google Scholar
|
|
4
|
Patwari P, Chutkow WA, Cummings K, et al:
Thioredoxin-independent regulation of metabolism by the
alpha-arrestin proteins. J Biol Chem. 284:24996–25003. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Spindel ON, World C and Berk BC:
Thioredoxin interacting protein: redox dependent and independent
regulatory mechanisms. Antioxid Redox Signal. 16:587–596. 2012.
View Article : Google Scholar :
|
|
6
|
Zhou J, Yu Q and Chng WJ: TXNIP (VDUP-1,
TBP-2): a major redox regulator commonly suppressed in cancer by
epigenetic mechanisms. Int J Biochem Cell Biol. 43:1668–1673. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cadenas C, Franckenstein D, Schmidt M, et
al: Role of thioredoxin reductase 1 and thioredoxin interacting
protein in prognosis of breast cancer. Breast Cancer Res.
12:R442010. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Frullanti E, Colombo F, Falvella FS, et
al: Association of lung adenocarcinoma clinical stage with gene
expression pattern in noninvolved lung tissue. Int J Cancer.
131:E643–E648. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nishizawa K, Nishiyama H, Matsui Y, et al:
Thioredoxin-interacting protein suppresses bladder carcinogenesis.
Carcinogenesis. 32:1459–1466. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Butler LM, Zhou X, Xu WS, et al: The
histone deacetylase inhibitor SAHA arrests cancer cell growth,
up-regulates thioredoxin-binding protein-2, and down-regulates
thioredoxin. Proc Natl Acad Sci USA. 99:11700–11705. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ku WL, Girvan M, Yuan GC, Sorrentino F and
Ott E: Modeling the dynamics of bivalent histone modifications.
PLoS One. 8:e779442013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Santos-Rosa H, Schneider R, Bannister AJ,
et al: Active genes are tri-methylated at K4 of histone H3. Nature.
419:407–411. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Barski A, Cuddapah S, Cui K, et al:
High-resolution profiling of histone methylations in the human
genome. Cell. 129:823–837. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chavez KJ, Garimella SV and Lipkowitz S:
Triple negative breast cancer cell lines: one tool in the search
for better treatment of triple negative breast cancer. Breast Dis.
32:35–48. 2010.
|
|
15
|
Lavarone E, Puppin C, Passon N, Filetti S,
Russo D and Damante G: The PARP inhibitor PJ34 modifies
proliferation, NIS expression and epigenetic marks in thyroid
cancer cell lines. Mol Cell Endocrinol. 365:1–10. 2013. View Article : Google Scholar
|
|
16
|
Baldan F, Lavarone E, Di Loreto C, Filetti
S, Russo D, Damante G and Puppin C: Histone post-translational
modifications induced by histone deacetylase inhibition in
transcriptional control units of NIS gene. Mol Biol Rep.
41:5257–5265. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Marks PA and Xu WS: Histone deacetylase
inhibitors: potential in cancer therapy. J Cell Biochem.
107:600–608. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Russo D, Damante G, Puxeddu E, Durante C
and Filetti S: Epigenetics of thyroid cancer and novel therapeutic
targets. J Mol Endocrinol. 46:R73–R81. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhou J and Chng WJ: Roles of thioredoxin
binding protein (TXNIP) in oxidative stress, apoptosis and cancer.
Mitochondrion. 13:163–169. 2013. View Article : Google Scholar
|
|
20
|
Ungerstedt JS, Sowa Y, Xu WS, et al: Role
of thioredoxin in the response of normal and transformed cells to
histone deacetylase inhibitors. Proc Natl Acad Sci USA.
102:673–678. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen A: PARP inhibitors: its role in
treatment of cancer. Chin J Cancer. 30:463–471. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yamaguchi F, Hirata Y, Akram H, et al:
FOXO/TXNIP pathway is involved in the suppression of hepatocellular
carcinoma growth by glutamate antagonist MK-801. BMC Cancer.
13:4682013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Connolly R and Stearns V: Epigenetics as a
therapeutic target in breast cancer. J Mammary Gland Biol
Neoplasia. 17:191–204. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Füllgrabe J, Kavanagh E and Joseph B:
Histone onco-modifications. Oncogene. 30:3391–3403. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lee JM, Hays JL, Annunziata CM, et al:
Phase I/Ib study of olaparib and carboplatin in BRCA1 or BRCA2
mutation-associated breast or ovarian cancer with biomarker
analyses. J Natl Cancer Inst. May 19–2014.Epub ahead of print.
View Article : Google Scholar
|
|
26
|
Chu BF, Karpenko MJ, Liu Z, et al: Phase I
study of 5-aza-2′-deoxycytidine in combination with valproic acid
in non-small-cell lung cancer. Cancer Chemother Pharmacol.
71:115–121. 2013. View Article : Google Scholar
|
|
27
|
Falchook GS, Fu S, Naing A, et al:
Methylation and histone deacetylase inhibition in combination with
platinum treatment in patients with advanced malignancies. Invest
New Drugs. 31:1192–1200. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jasek E, Gajda M, Lis GJ, Jasińska M and
Litwin JA: Combinatorial effects of PARP inhibitor PJ34 and histone
deacetylase inhibitor vorinostat on leukemia cell lines. Anticancer
Res. 34:1849–1856. 2014.PubMed/NCBI
|