Introduction
Bladder cancer (BC) is the fourth most common cancer
in men worldwide, with an estimated 70,530 new cases and a
projected number of 14,680 deaths in the United States in 2010
(1). In China, BC is the eighth
most common cancer among all malignant tumors and the most common
cancer of the genitourinary system in men (2). At initial diagnosis, ~70–75% of the
patients are afflicted with non-muscle invasive bladder cancer
(NMIBC), in which the papillary tumor invades the urothelium (Ta)
and lamina propria (T1), or carcinoma in situ (CIS), with a
relatively flat tumor within the urothelium, while the remaining
25–30% already have a muscle invasive bladder cancer (MIBC)
(3,4). More than 90% of BC cases belongs to
the pathological type, transitional cell (urothelial) carcinomas
(TCCs), and 3–7% are squamous cell carcinomas; adenocarcinoma is
rare and accounts for <2% of all BC cases (5). The majority of TCCs present as
papillary NMIBC; however, ~80% of patients with NMIBC suffer from
recurrence within 1 year to 2 years and ~8% will progress to MIBC
although standard treatment is performed, including transurethral
resection and adjuvant intravesical chemotherapy or immunotherapy
(3). Patients with MIBC are treated
by radical cystectomy (3). Despite
the radical cystectomy and systemic therapy, 50% of patients with
invasive tumors die from metastasis (6). Recent progress has been made in
diagnosis and treatment, but the molecular mechanisms underlying
the development and progression of BC remain poorly understood and
MIBC prognosis is still unsatisfactory (7). Further investigation of the molecular
mechanisms of development and progression of BC is necessary to
develop new methods for the treatment of BC.
Engrailed (EN) is a member of the homeobox gene
family and was first characterized in Drosophila. The
encoded homeodomain-containing transcription factor has important
functions in development and has been identified in annelids
(8), mollusks (9), insects (10), echinoderms, chordates (11) and vertebrates (12). In vertebrates, two engrailed (EN)
genes have been discovered, namely, EN1 and EN2, which differ
slightly in their specific functions (13,14).
High levels of both EN1 and EN2 are expressed in alar (dorsal)
cells of the midbrain/hindbrain border region during brain
development and affect the survival of mesencephalic dopaminergic
neurons (15,16). The EN2 gene has also been implicated
in early muscle development, with expression in the murine
mandibular arch, particularly myoblasts, which give rise to the
masseter, temporalis, and lateral and medial pterygoid muscles
involved in jaw closure (17).
Germline mutations in EN may have profound effects on embryological
development.
Evidence suggests that overexpression of EN2 protein
may be associated with tumor development in adult humans,
particularly in breast (18),
prostate (19–21) and ovarian cancers (22). A recent study indicated that EN2 is
expressed in and secreted by BC cell lines and patient tumor
specimens, justifying an evaluation of urinary EN2 as a diagnostic
biomarker in BC using archived samples from an established
biospecimen collection. Moreover, high expression of EN2 is
associated with the pathological grade and stage of BC (23). However, the mechanisms of how EN2
functions to promote BC progression remain elusive. Given the
significance of EN2 in cancer tumorigenesis, EN2 is hypothesized to
have critical functions in BC tumorigenesis and metastasis. In the
present study, the effects of EN2 knockdown on the proliferation,
apoptosis, and invasion of BC were investigated in vitro and
in vivo using small interference RNA (siRNA) for EN2. The
obtained results suggest that downregulation of EN2 may be
effective in the inhibition of tumor growth and progression of
BC.
Materials and methods
Cell lines and cell culture
T24, RT4 and EJ human BC cell lines as well as
normal uroepithelial SV-HUC-1 cells were obtained from the American
Type Culture Collection (ATCC; Manassas, VA, USA). Normal human
bladder epithelial cells (NHBECs) were purchased from PriCells Co.,
Ltd. (Wuhan, China). The cells were thawed and cultured at 37°C in
5% CO2 in Dulbecco’s modified Eagle’s medium (DMEM;
HyClone, Logan, UT, USA) containing 10% fetal bovine serum (FBS), 2
mmol/l glutamine, 1 mmol/l sodium pyruvate, 10 mmol/l
4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid, 100 U/ml
penicillin G, 100 mg/ml streptomycin and 50 ng/ml SDF-1 (Sigma, St.
Louis, MO, USA).
Construction of recombinant plasmids
A pair of primers with double restriction enzyme
cutting sites (EcoRI-forward primer,
5′-CCGGAATTCATGGAGGAGAATGACCCCAAGCCT GGCG-3′ and
BamHI-reverse primer, 5′-CCGCTCGAGCT
ACTCGCTGTCCGACTTGCCCTCCTTG-3′) was designed to amplify the
full-length cDNA of EN2 (GenBank Accession No. NM_001427) from the
cellular cDNA library of the human bladder epithelium. The
restriction enzymes EcoRI and BamHI were then used to
cut the PCR products and expression vector pcDNA3.1 (Invitrogen).
Finally, the EN2 Flag-tagged overexpression vector was obtained
through ligation reaction. In addition, the accurate sequence of
EN2 open reading frame was confirmed by DNA sequencing.
Construction of the plasmids for
knockdown of human EN2 expression
The following three siRNAs were designed based on
the EN2 sequence: siRNA 1, 5′-GGCTCAAGGCCGAGT TCCA-3′, siRNA 2,
5′-GGGTCTACTGTACGCGCTA-3′, and siRNA 3, 5′-CAAAGAAGAAGAACCCGAA-3′.
Oligonucleotides with a sequence predicted to induce efficient RNAi
of EN2 (containing sense and antisense sequences) were synthesized
by Sigma (Shanghai, China). The synthesized sequences are shown in
Table I. The oligonucleotides were
annealed in sodium chloride-Tris-EDTA buffer at 94°C for 5 min and
cooled gradually. The double-stranded products were cloned
downstream to the human U6 promoter of the pGCsilencer U6/Neo/GFP
(pGC) plasmid (Shanghai Genechem Co., Ltd., Shanghai, China). The
products were designated as EN2-siRNA1, EN2-siRNA2 and EN2-siRNA3.
A scramble sequence (5′-TTCTCCGAACGTGTCACGT-3′) without significant
homology to any mammalian gene sequence was inserted into the pGC
plasmid and used as a negative control siRNA.
 | Table IThe synthesized siRNA sequences for
construction of plasmids for knockdown of human EN2 expression. |
Table I
The synthesized siRNA sequences for
construction of plasmids for knockdown of human EN2 expression.
| siRNAs | Sequences
(5′-3′) |
|---|
| siRNA-1 | Sense:
GATCCCGGCTCAAGGCCGAGTTCCATTCAAGAGATGGAACTCGGCCTTGAGCCTTTTTT
Antisense:
CTAGAAAAAAGGCTCAAGGCCGAGTTCCATCTCTTGAATGGAACTCGGCCTTGAGCCCGG |
| siRNA-2 | Sense:
GATCCCGGGTCTACTGTACGCGCTATTCAAGAGATAGCGCGTACAGTAGACCCTTTTTT
Antisense:
CTAGAAAAAAGGGTCTACTGTACGCGCTATCTCTTGAATAGCGCGTACAGTAGACCCCGG |
| siRNA-3 | Sense:
GATCCCCAAAGAAGAAGAACCCGAATTCAAGAGATTCGGGTTCTTCTTCTTTGTTTTTT
Antisense:
CTAGAAAAAACAAAGAAGAAGAACCCGAATCTCTTGAATTCGGGTTCTTCTTCTTTGCGG |
Cell transfection
Cells were plated on 6-well plates at 80–90%
confluency (recombinant plasmid transfection) or 30–50% confluency
(siRNA transfection). All cell transfections were implemented using
Lipofectamine 2000 (Invitrogen) according to the manufacturer’s
protocol. An empty vector was used as a negative control. To
determine whether the recombinant plasmids were transduced into the
tumor cells, the green fluorescent protein (GFP) expression of EJ
cells in the NC-siRNA, EN2-siRNA1, EN2-siRNA2 and EN2-siRNA3 groups
was assessed using an inverted fluorescence microscope at 24, 48
and 72 h after transduction, respectively. Ten fields were randomly
selected, and the delivery efficiency was counted as follows:
number of positive cells/total number of cells × 100% under the
same field. The samples were considered positive regardless of the
degree of brightness.
Selection of stable transfected cell
clones
NHBECs transfected with pcDNA3.1 and pcDNA3.1+EN2 as
well as BC cells transfected with pGCsilencer plasmids were
cultured in the medium with added G418 (Gibco) for 2 weeks. Stable
transfected clones were selected using G418. Briefly, two EJ clones
stably expressing EN2 siRNA (siRNA1) (named as C1 and C2,
respectively) and one EJ clone stably expressing NC-siRNA (named as
NC) were identified using indirect immunofluorescent assay, which
were further verified with qRT-PCR and western blot analysis.
Quantitative RT-PCR
Total RNA was isolated from frozen tumor tissue or
1×106 cells using the TRIzol reagent (Invitrogen). Total
RNA was subsequently reverse transcribed to cDNA using M-MLV
reverse transcriptase (Promega) according to the manufacturer’s
instructions. DNase (Promega) was used to completely eliminate DNA
contamination in the total RNA before reverse transcription.
qRT-PCR was performed using the SYBR-Green qPCR SuperMix kit
(Invitrogen) in the ABI PRISM® 7500 Sequence Detection
system (Applied Biosystems, Foster City, CA, USA). Specific primers
were: EN2, F-5′-TCTTGGAGTGGCTGCTTCTG3′ and
R-5′-TCCTGGAGGATTCTGAGTTCTT3′; 18S rRNA, F-5′CCTGGATACCGCAGCTAGGA3′
and R-5′-GCGGCG CAATACGAATGCCCC-3′. Thermal cycle conditions were
as follows: 5 min at 95°C and 40 cycles of 1–5 sec at 95°C, 32 sec
at 60°C, and 30 sec at 72°C. The comparative threshold cycle method
was used to calculate the amplification fold. 18S rRNA was employed
as a reference control gene to normalize the expression value of
the target genes. Triple replicates were performed for each gene,
and the average expression value was calculated for subsequent
analysis. The relative expression levels of the genes were
calculated using the 2−ΔΔCt method (24).
Western blot analysis
In brief, the cells were rinsed thrice with ice-cold
PBS and then lysed in lysis buffer (PBS at pH 7.4, 1% Triton X-100,
10 mM sodium deoxycholate, 3 mM SDS, and 1 mM EDTA with protease
inhibitors) at 4°C for 30 min. Protein concentrations were
determined using a BCA protein assay (Thermo Fisher Scientific).
Equal quantities of protein were subjected to SDS-PAGE and
transferred to Immobilon-P transfer membranes (Millipore).
Successive incubations with primary antibodies (Table II) and horseradish
peroxidase-conjugated secondary antibody were conducted.
Immunoreactive proteins were then detected using the enhanced
chemiluminescence system (Bio-Rad Laboratories). For the protein in
xenograft tissues, the harvested xenograft samples were cut into
small pieces and dissolved in lysis solution. The complex solution
was homogenized and sonicated for 5 min on ice. Protein
concentrations were determined using a BCA protein assay. All
proteins to be validated were separated by SDS-PAGE with 20
μg of protein per lane and transferred to Immobilon-P
transfer membranes [in transfer buffer (pH 11.0), 25 mmol/l Tris,
0.2 mol/l glycine, 20% (v/v) methanol] for 45 min at 1.5
mA/cm2 on a semidry electroblotter (Bio-Rad
Laboratories, Richmond, VA, USA). The membranes were blocked with
1% skim milk for 1 h at 25°C and probed with the primary antibody
(Table II) and then horseradish
peroxidase-conjugated secondary antibody. GAPDH was used as an
internal control to ensure equal loading. Bands were scanned using
a densitometer (GS-700; Bio-Rad Laboratories, Hercules, CA, USA),
and relative quantification was performed using Image-Pro Plus 6.0
software (Media Cybernetics Inc., Silver Spring, MD, USA).
| Table IIPrimary antibodies for western blot
analysis. |
Table II
Primary antibodies for western blot
analysis.
| Primary
antibodies | Manufacturer | Product no | Dilutions |
|---|
| EN2 | Abcam | ab28731 | 1:20000 |
| Cyclin D1 | Abcam | ab16663 | 1:200 |
| E-cadherin | Abcam | ab40772 | 1:20000 |
| TP53 | Abcam | ab65021 | 1:500 |
| PTEN | Abcam | ab32199 | 1:500 |
| VEG | Abcam | ab46154 | 1:1000 |
| PDGF | Abcam | ab125268 | 1:800 |
| Bcl-2 | Abcam | ab59348 | 1:1000 |
| BAX | Abcam | ab69643 | 1:1000 |
| MMP-2 | Abcam | ab92536 | 1:1000 |
| MMP-9 | Abcam | ab38898 | 1:1000 |
| p-Akt (Ser
473) | Santa Cruz | sc-33437 | 1:500 |
| p-Akt (Thr
308) | Santa Cruz | sc-135650 | 1:500 |
| PI3K p110 | Santa Cruz | sc-7189 | 1:500 |
| PI3K p85 | Santa Cruz | sc-292114 | 1:500 |
| GAPDH | Abcam | ab9385 | 1:5000 |
Cell cycle assay
Cells (1×10 6) were obtained from
appropriate samples, and nuclei were stained with PI. Flow
cytometry was performed using FACSCalibur (BD Biosciences, San
Jose, CA, USA), and the cell fraction in each cell cycle phase was
determined using the cell cycle analysis platform in FlowJo
software (Tree Star Inc., Ashland, OR, USA).
Apoptosis assay
Annexin V-FITC/PI apoptosis kit was purchased from
Nanjing KeyGen Biotech Co., Ltd. (Nanjing, China). In brief, cells
(5×105) were collected, washed, resuspended in PBS and
Annexin V-FITC (1.25 μl) and PI (10 μl) were added.
The cells were then incubated for 20 min at 4°C. Tests were
performed in triplicate for each sample, and analyses were
performed by FACScan flow cytometry (Becton-Dickinson, San Jose,
CA, USA) with FlowJo software according to the manufacturer’s
guidelines. The TUNEL assay was performed using In Situ Cell Death
Detection kit, POD (Roche Applied Science, Basel, Switzerland)
according to the manufacturer’s protocol. The sections were
observed under a bright-field microscope, and the number of
apoptotic cells in the tumor tissue in each section was counted in
10 different microscopic fields.
In vitro invasion assay
In brief, a polycarbonate filter was coated with
Matrigel (BD Biosciences, Franklin Lakes, NJ, USA) and incubated at
37°C for half an hour. The cells were harvested by typsinization
and suspended in DMEM serum-free medium with 0.1% BSA at
5×105 cells/ml. Approximately 100 μl of the cell
suspensions was placed in the upper chambers, and 600 μl of
complete medium was filled into the lower chambers. The cells were
allowed to invade at 37°C in 5% CO2 for 48 h and fixed
with 4% formaldehyde for 15 min. The cells on the lower surface of
the membranes were stained with crystal violet for 10 min. The
invaded cells in five fields (i.e., upper, lower, left, right and
middle) were counted under a microscope at a ×200 magnification.
Experiments were repeated at least three times, and data are
presented as the number of cells passing through the filters.
Cell proliferation assay
Cell proliferation was assessed using CellTiter
96® Aqueous One Solution Cell Proliferation assay (MTS
assay; Promega) according to the manufacturer’s instructions. Cells
were plated on 96-well plates at 1×104 cells/well (i.e.,
100 μl/well) and incubated for 3 days. MTS (10 μl)
was added to each well after 0, 24, 48 and 72 h of incubation. The
cells were incubated at 37°C in 5% CO2 for another 4 h.
The value of the optical density was measured at a wavelength of
490 nm using a microplate reader (multiscan MK3; Thermo Fisher
Scientific).
In vivo assay
All animal studies were carried out in accordance
with the Guide for the Care and Use of Laboratory Animals published
by the US National Institutes of Health. The experimental
procedures and protocols were approved by the Ethics Committee for
Animal Experiments of Jinan University. In brief, male BALB/c nude
mice aged 4 weeks were obtained and maintained in a pathogen-free
facility. Cells from two EJ cell clones stably expressing EN2 siRNA
(C1 and C2) and one stable clone expressing a control scramble
siRNA (NC) were suspended in PBS at a concentration of
5×106 cells/ml and injected subcutaneously in the back
of the mice (n=6 for each group). The tumor size was measured every
7 days, and the tumor volume was determined using the simplified
formula of a rotational ellipsoid (L × W2 × 0.5). Six
weeks after inoculation, the mice were sacrificed and dissected for
collection of primary tumors and organs, including lungs and
livers. The tissues were then fixed in 4% paraformaldehyde and
embedded in paraffin for further study.
Immunohistochemistry
Five-micrometer sections of formalin-fixed,
paraffin-embedded tissue were cut onto silanized glass slides,
deparaffinized in xylene, rehydrated in graded ethanol
concentrations (100, 95, 70 and 50%), and finally submerged in PBS.
The sections were blocked for endogenous peroxidase with 3%
hydrogen peroxide solution for 15 min and placed in an autoclave
with 0.01 mol/l sodium citrate solution at 121°C for 3 min for
antigen retrieval. The sections were incubated with EN2 (1:500) and
Ki-67 (Abcam; ab66155, 1:200) primary antibodies overnight at 4°C
and then incubated with biotin-labeled secondary antibody at room
temperature for 1 h. Negative controls were performed by replacing
the primary antibody with PBS. Diaminobenzidine tetrahydrochloride
was used as a chromogen. All sections were then counterstained with
hematoxylin and observed under a bright-field microscope.
Statistical analysis
Quantitative data are expressed as mean ± SD
(standard deviation). The statistical significance of differences
between groups was determined by one-way ANOVA analysis.
Enumeration of the data was analyzed by Mann-Whitney U test.
P<0.05 was considered to indicate a statistically significant
result. All data were analyzed with SPSS 17.0 software.
Results
Significant expression of EN2 in BC
cells
The EN2 expression was examined using quantitative
real-time polymerase chain reaction (qRT-PCR) and western blot
analysis in human BC cell lines, including T24, RT4 and EJ cell
lines, as well as normal uroepithelium SV-HUC-1 cells. The EN2 mRNA
levels were significantly higher in all BC cell lines than the
level in the SV-HUC-1 cells. The EN2 mRNA level in the EJ cell line
was also significantly higher than levels in the T24 and RT4 cell
lines (Fig. 1A). The EN2 protein
levels showed the same trend with mRNA expression (Fig. 1B). Therefore, the EJ cell line was
selected for further study.
EN2 overexpression promotes cell
proliferation, invasion and apoptosis
To investigate the possible oncogenic function of
EN2 in normal bladder epithelial cells, we selected the
immortalized and non-transformed bladder epithelial cell line
SV-HUC-1 for further study. The stable cells transfected with
pcDNA3.1 and pcDNA3.1+EN2 as well as non-transfected cells were
examined for the effects of EN2 overexpression on cellular
proliferation, invasion and apoptosis. The expression of exogenous
EN2 mRNA and protein was verified using qRT-PCR and western blot
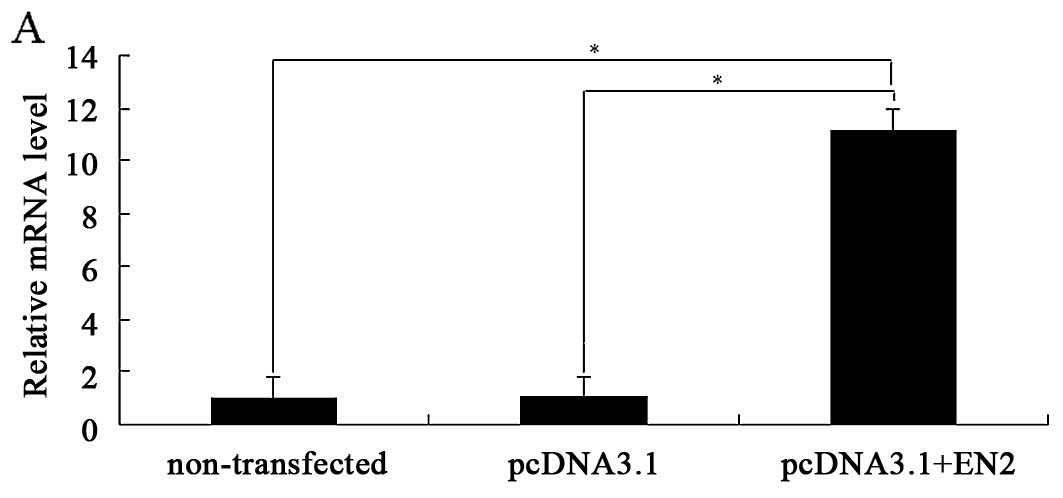
analysis, respectively (Fig. 2A and
B).
The cell growth curve drawn with absorbance at 490
nm showed that the stable SV-HUC-1 cells transfected with
pcDNA3.1+EN2 had larger values of cell viability than the
corresponding stable SV-HUC-1 cells transfected with pcDNA3.1 and
the non-transfected SV-HUC-1 cells (Fig. 2C). Moreover, SV-HUC-1 cells
ectopically expressing EN-2 were more invasive compared with the
non-transfected cells or pcDNA3.1-transfected cells (Fig. 2D). Apoptotic analysis with flow
cytometry showed a lower proportion of apoptotic SV-HUC-1 cells
ectopically expressing EN-2 compared with the non-transfected cells
or pcDNA3.1-transfected cells (Fig.
2E). In summary, the results indicated that EN2 overexpression
enhanced cellular proliferation and invasion while inhibiting
cellular apoptosis.
Effects of EN2 siRNA on the expression of
EN2 in EJ cells in vitro
The delivery efficiency of NC-siRNA, EN2-siRNA1,
EN2-siRNA2 and EN2-siRNA3 was >85% at 48 h by assessing the
green fluorescent protein (GFP) expression under an inverted
fluorescence microscope (data not shown). After EN2-siRNA1,
EN2-siRNA2 and EN2-siRNA3 transduction for 48 h in EJ cells, the
expression of EN2 mRNA was substantially reduced by 91.7, 89.7 and
81.0% while its protein expression was reduced by 85.4, 74.7 and
51.6%, respectively, compared with the blank group (data not
shown). Therefore, the EN2-siRNA1 plasmid was chosen for further
study.
Selection of stable clones of
siRNA-transfected EJ cells by G418
The results showed that the positive proportion of
GFP in the above three clones was >85% while the intensity of
red fluorescence in C1 and C2 cells was very weak, compared with
NC, which indicated that knockdown efficiency of C1 and C2 was very
high (data not shown).
EN2 knockdown promotes cell cycle arrest
and apoptosis of EJ cells but inhibits proliferation and invasion
in vitro
After identifying the siRNA with the best inhibition
for EN2 and two stable clones of EJ cells with EN2-siRNA
transfection (C1 and C2), the effects of EN2 knockdown on the
biological behavior of EJ cells were investigated. The results
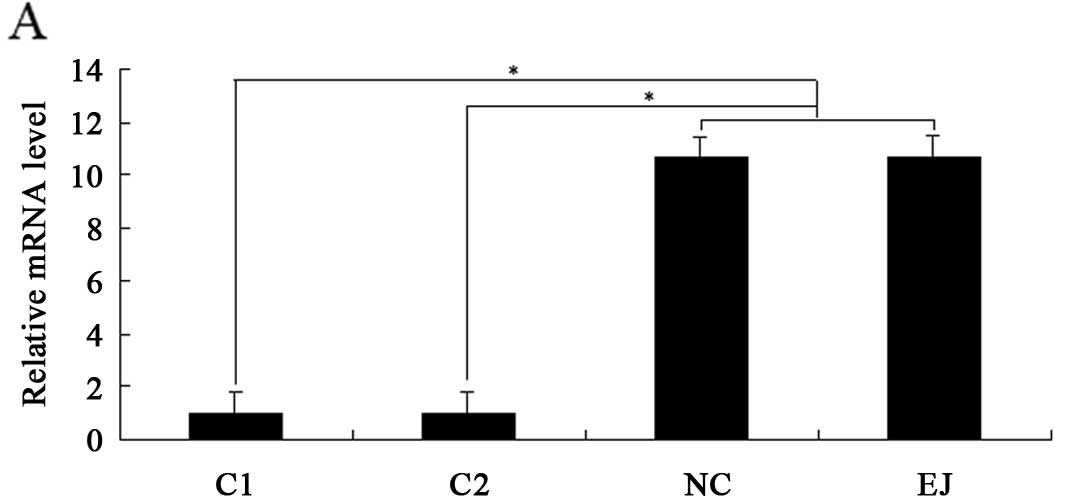
showed that the expression levels of EN2 mRNA and protein in C1 and
C2 were significantly downregulated than those in the NC and EJ
cells (Fig. 3A and B). The
percentages of cells in the S phase decreased from ~35.2 to 21.7
and 21.5% in the C1 and C2 cells, respectively, whereas the
percentages of cells in the G0/G1 phase increased from ~52.1 to
67.4 and 68.3% in C1 and C2, respectively. The percentages of cells
in the G2/M phase showed no significant differences among the
various samples (Fig. 3C). These
results suggest that EN2 downregulation leads to cell cycle arrest
at the G0/G1 phase.
 | Figure 3Effects of EN2 knockdown on the cell
cycle, apoptosis, proliferation and invasion of BC cells in
vitro. (A) Expression of EN2 mRNA was detected using qRT-PCR.
(B) Expression of EN2 protein was detected by western blot
analysis. (C) After cultivation for 48 h, analysis of cell cycle
distribution of various cells, including EJ, NC, C1 and C2, was
performed by flow cytometry. The fractions of cells in the G0/G1, S
and G2/M phases are shown in the representative histogram.
*P<0.05. (D) After cultivation for 48 h, the
apoptosis of various cells was detected, and quantitative analysis
of positive Annexin V cells in the different groups is shown in the
representative histogram. (E) After obtaining stable clones, cell
proliferation was assessed using the MTS assay. (F) Effects of EN2
knockdown on in vitro invasion. *P<0.05. |
After cultivation for 48 h, Annexin V/propidium
iodide (PI) staining of various cells showed that the proportions
of Annexin V-positive cells in C1 and C2 were ~39.9 and ~38.7%,
respectively, whereas the proportion of Annexin V-positive cells in
the EJ cells was only 9% (Fig. 3D).
These data suggest that EN2 downregulation promotes EJ cell
apoptosis.
MTS assay demonstrated that EN2 knockdown
significantly decreased the proliferation of EJ cells (Fig. 3E). Furthermore, EN2 knockdown also
significantly decreased the invasive ability of the EJ cells
(Fig. 3F).
Effects of EN2 knockdown on
proliferation, apoptosis, invasion and metastasis of BC in
vivo
EJ and NC groups showed a significantly more rapid
increase in tumor volume than that in the C1 and C2 groups
(P<0.05). However, no differences were observed in regards to
tumor size and growth pattern between the EJ and NC groups or
between the C1 and C2 groups (Fig.
4A and B).
 | Figure 4Inhibitory effects of EN2 knockdown
on bladder cancer in vivo. (A) Images of representative mice
and tumors are shown on day 42 after inoculation. (B) Tumor size
was measured, and tumor volume was determined as described in
Materials and methods every 7 days for 42 days, n=6. (C and D)
Adhesionrelated protein, tumor-suppressor protein,
apoptosis-related protein, matrix metalloproteinases, cell cycle
regulation protein and tumor angiogenesis factors were determined
using western blot analysis. (E) Immunohistochemical detection of
EN2, proliferation marker Ki-67 and apoptosis marker TUNEL.
EN2-positive staining was located in the tumor nuclei and
cytoplasm, and more positive cells were observed in the EJ and NC
groups than in the C1 and C2 groups; Ki-67-positive staining was
located in the cell nuclei, and more positive cells were observed
in the EJ and NC groups than in the C1 and C2 groups. In the TUNEL
assay, positive cells were located in the cell nuclei. Scale bars,
50 μm. (F) Representative images of hematoxylin and
eosin-stained sections of lung and liver micrometastases, and (G)
quantification of the liver and lung micrometastases. (H)
Representative images of lung and liver metastases in nude mice.
Scale bars, 100 μm. *P<0.05. |
To further investigate the potential mechanisms of
BC inhibition by EN2 knockdown, we detected the expression of
various representative proteins that have been confirmed to be
associated with the development and progression of tumors using
western blot analysis. The results demonstrated that
adhesion-related protein E-cadherin (CDH1), tumor-suppressor
proteins, including TP53 and PTEN, as well as anti-apoptosis
protein Bcl-2, were significantly downregulated in the EJ and NC
groups. Matrix metalloproteinases, including MMP-2 and MMP-9,
pro-apoptosis protein BAX, and cell cycle regulation protein cyclin
D1, as well as tumor angiogenesis factors, including VEGF and PDGF,
were also significantly downregulated in the C1 and C2 groups,
which were consistent with the expression of their own genes
(Fig. 4C and D).
Immunohistochemistry results showed that the protein
expression levels of EN2 and Ki-67 were significantly lower in the
C1 and C2 groups than those in the EJ and NC groups (Fig. 4E). The number of apoptotic cells was
significantly increased in tumors from the C1 and C2 groups than
the number in tumors from the EJ and NC groups as detected by TUNEL
assay (Fig. 4E).
In addition, the number of micrometastatic events
(observed under a microscope) in the lungs and livers was
significantly lower in mice from the C1 and C2 groups than the
events in lungs and livers from the EJ and NC groups (Fig. 4F and G). Similarly, the number of
metastatic nodules (macroscopic) in the lungs and livers was
apparently reduced in the C1 and C2 groups compared with that in
the EJ and NC groups (Fig. 4H; data
not shown).
EN2 appears to induce activation of the
phosphatidylinositol 3-kinase (PI3K) pathway by inhibiting
PTEN
The PI3K pathway possesses a critical function in
cell growth, proliferation and survival (25), and signaling via the P13K pathway is
upregulated in many types of cancer (26,27).
Given the expression levels of EN2 and PTEN (the main inhibitor of
PI3K-Akt pathway) in the aforementioned experiments, the levels of
Akt phosphorylated at serine 473 (pAkt-473) and threonine 308
(pAkt-308), PI3K and PTEN protein were determined by western blot
analysis in vitro. As shown in Fig. 5, the levels of pAkt-473, pAkt-308
and PI3K were significantly higher in the EJ and NC cell lines than
those in the C1 cell line (EN2-siRNA-transfected). This result
indicates that EN2 knockdown significantly decreased the levels of
pAkt-473, pAkt-308 and PI3K, but increased PTEN expression.
Discussion
EN2, a homeobox-containing transcription factor, was
identified as a candidate oncogene in breast and prostate cancer.
Previous studies indicate that EN2 is overexpressed in human breast
and prostate cancer cells, and downregulation of EN2 expression
causes a decrease in breast and prostate cancer cell proliferation
(18,19). In the present study, EN2 was highly
expressed in BC cell lines. The ectopic expression of EN2 in normal
urothelial cells significantly enhanced cellular proliferation and
invasion, but inhibited cellular apoptosis. Moreover, RNAi-mediated
EN2 silencing significantly downregulated the protein levels of EN2
in vitro and in vivo. More importantly, EN2 knockdown
significantly promoted cell cycle arrest and apoptosis of tumor
cells and inhibited tumor growth and distant metastasis. All of
these findings suggest that EN2 has potential functions in BC
development and progression and is a promising gene-targeting
therapy for BC.
The cell cycle has important functions in cell
proliferation, differentiation and tumor progression and is
regulated by intracellular and extracellular signal transduction
pathways (28). In the present
study, the number of cells in the G0/G1 phase was significantly
increased, whereas that in the S phase was significantly decreased
in the C1 and C2 cells. This result suggests that EN2 knockdown
induced G0/G1 phase arrest and apoptosis, as well as inhibited
tumor cell proliferation. Furthermore, the cell proliferation assay
indicated that EN2 knockdown led to a significant decrease in BC
cell proliferation. After constructing the xenograft tumor models
in nude mice, the tumor volumes in the C1 and C2 groups were
significantly lower than those in the EJ and NC groups on day 42.
Furthermore, cell apoptosis was detected using TUNEL assay, and the
Ki-67 protein expression was determined by immunohistochemistry
in vivo. The results showed that the number of apoptotic
cells was significantly increased and Ki-67 protein expression was
significantly decreased in tumors in the C1 and C2 groups compared
with those in the EJ and NC groups. These data indicate that
RNAi-mediated EN2 gene silencing inhibited tumor growth and induced
apoptosis of BC cells both in vitro and in vivo.
To further investigate the potential molecular
mechanisms of the effects of EN2 knockdown on tumor growth
inhibition, we detected the expression levels of various common
proteins that are related to the development and progression of
tumors in vivo. The results indicated that protumor proteins
were downregulated, whereas antitumor proteins were upregulated
after EN2 knockdown. Numerous genes and proteins are involved in
the development and progression of tumors, including BC. Loss of
TP53 and PTEN tumor suppressors is common in BC (29,30).
The Bcl-2 family functions as a ‘life/death switch’ that integrates
diverse intercellular and intracellular cues to determine whether
the stress apoptosis pathway should be activated (31). The p53 gene precisely regulates cell
apoptosis via activating apoptosis promoters of the Bcl-2 family,
such as BAX, or inhibiting antiapoptosis molecules, such as Bcl-2
(32). PTEN can negatively regulate
the PI3K/Akt pathway, which regulates several relevant processes
for tumor growth and progression, such as cell survival, motility,
invasion, cell cycle progression and angiogenesis (33,34).
Genetic aberrations in the regulatory circuits that govern transit
through the G1 phase of the cell cycle occur frequently in human
cancer, and overexpression of cyclin D1 is one of the most commonly
observed alterations, including BC (35,36).
Studies indicate that cyclin D1 downregulation contributes to
inhibition of BC proliferation by inducing cell cycle G0/G1 arrest
(37,38). VEGF and PDGF have important
functions in tumor angiogenesis for neoangiogenesis, which further
promotes tumor growth (39). CDH1
(E-cadherin) is a calcium-dependent cell adhesion molecule that
inhibits tumor invasion and metastasis through stabilization of
cell-cell adhesion (40). MMP-2 and
MMP-9 can degrade epithelial-mesenchymal transition (ECM) and
promote tumor metastasis (41).
The PI3K/AKT signaling pathway is a frequently
dysregulated pathway in cancer (42,43)
and possesses a major function in bladder carcinogenesis (44–48).
PTEN is the antagonist of PI3K, which removes the 3′ phosphate of
PIP3 and attenuates downstream signaling of activated PI3K
(49). Therefore, we further
investigated the effects of EN2 knockdown on the PI3K/Akt pathway
in vitro. EN2 knockdown significantly decreased the levels
of pAkt-473, pAkt-308 and PI3K protein, but increased PTEN protein.
This finding suggests that EN2 may activate the PI3K/Akt pathway by
inhibiting PTEN, thereby promoting the progression of BC.
In addition to transcription, EN2 protein may have a
regulatory function in translation as it can bind directly to the
eukaryotic translation initiation factor 4E (eIF4E) with high
affinity and specificity (50). EN2
should have the same characteristics as other homeoproteins since
it is a homeobox-containing transcription factor. Zhou et al
(51) indicated that Nkx3.1, a
homeoprotein transcription factor, functions within the cells that
produce them in a cell autonomous manner as well as can regulate
genes and inhibit cell proliferation in a non-cell autonomous
manner. Prin et al (52)
reported that Hox4 proteins regulate Eph/ephrins and other
cell-surface proteins during chick and mouse embryo development.
These proteins can also function in a non-cell-autonomous manner to
induce apical cell enlargement on both sides of their expression
border. Therefore, we hypothesized that EN2 may induce
pro-oncogenes, such as VEGF, or inhibit anti-oncogenes, such as
PTEN, to promote the development and progression of BC via the
aforementioned three mechanisms (i.e., regulating translation by
binding directly to eIF4E and functioning in a cell autonomous and
non-cell-autonomous manner). This hypothesis needs to be further
verified.
Notably, we found that most mice in the EJ and NC
groups presented with liver and lung metastases, whereas mice in
the C1 and C2 groups did not show liver and lung metastases. This
result suggests that EN2 knockdown significantly inhibited the
metastasis of BC cells to the liver and lung in vivo.
The effects of EN2 knockdown on gene expression
profiling of BC were not analyzed due to the limitation in
experimental conditions, which may further confirm the results in
the present study. However, the study indicated that EN2 may
promote cell proliferation, invasion, and metastasis in BC by
activating the PI3K/Akt pathway and inhibiting PTEN. Importantly,
EN2 knockdown significantly decreased tumor growth of BC through
regulating the cell cycle, apoptosis and ECM-related proteins.
Thus, EN2 may be a candidate oncogene in BC and may be used as a
therapeutic target for the treatment of BC.
Acknowledgments
The present study was supported by the Major
Research Fund for the People’s Livelihood of Guangzhou Science and
Technology Plan (2011Y-00003).
References
|
1
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Han S, Zhang S, Chen W and Li C: Analysis
of the status and trends of bladder cancer incidence in China.
Oncol Prog. 11:89–95. 2013.In Chinese.
|
|
3
|
Babjuk M, Burger M, Zigeuner R, Shariat
SF, van Rhijn BW, Compérat E, Sylvester RJ, Kaasinen E, Böhle A,
Palou Redorta J, et al: EAU guidelines on non-muscle-invasive
urothelial carcinoma of the bladder: update 2013. Eur Urol.
64:639–653. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gakis G, Witjes JA, Compérat E, Cowan NC,
De Santis M, Lebret T, Ribal MJ and Sherif AM; European Association
of Urology: EAU guidelines on primary urethral carcinoma. Eur Urol.
64:823–830. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Na Y and Guo Z: Practice of Urology.
People’s Medical Press; Beijing: pp. 280–296. 2009, In Chinese.
|
|
6
|
Wu X: Urothelial tumorigenesis: a tale of
divergent pathways. Nat Rev Cancer. 5:713–725. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Efstathiou JA, Spiegel DY, Shipley WU,
Heney NM, Kaufman DS, Niemierko A, Coen JJ, Skowronski RY, Paly JJ,
McGovern FJ, et al: Long-term outcomes of selective bladder
preservation by combined-modality therapy for invasive bladder
cancer: the MGH experience. Eur Urol. 61:705–711. 2012. View Article : Google Scholar
|
|
8
|
Wedeen CJ and Weisblat DA: Segmental
expression of an engrailed-class gene during early development and
neurogenesis in an annelid. Development. 113:805–814.
1991.PubMed/NCBI
|
|
9
|
Wanninger A and Haszprunar G: The
expression of an engrailed protein during embryonic shell formation
of the tusk-shell, Antalis entails (Mollusca, Scaphopoda). Evol
Dev. 3:312–231. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fjose A, McGinnis WJ and Gehring WJ:
Isolation of a homoeo box-containing gene from the engrailed region
of Drosophila and the spatial distribution of its transcripts.
Nature. 313:284–289. 1985. View
Article : Google Scholar : PubMed/NCBI
|
|
11
|
Holland LZ, Kene M, Williams NA and
Holland ND: Sequence and embryonic expression of the amphioxus
engrailed gene (AmphiEn): the metameric pattern of transcription
resembles that of its segment-polarity homolog in Drosophila.
Development. 124:1723–1732. 1997.PubMed/NCBI
|
|
12
|
Joyner AL, Kornberg T, Coleman KG, Cox DR
and Martin GR: Expression during embryogenesis of a mouse gene with
sequence homology to the Drosophila engrailed gene. Cell. 43:29–37.
1985. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hanks M, Wurst W, Anson-Cartwright L,
Auerbach AB and Joyner AL: Rescue of the En-1 mutant phenotype by
replacement of En-1 with En-2. Science. 269:679–682. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hanks MC, Loomis CA, Harris E, Tong CX,
Anson-Cartwright L, Auerbach A and Joyner A: Drosophila engrailed
can substitute for mouse Engrailed1 function in mid-hindbrain, but
not limb development. Development. 125:4521–4530. 1998.PubMed/NCBI
|
|
15
|
Simon HH, Saueressig H, Wurst W, Goulding
MD and O’Leary DD: Fate of midbrain dopaminergic neurons controlled
by the engrailed genes. J Neurosci. 21:3126–3134. 2001.PubMed/NCBI
|
|
16
|
Alberi L, Sgado P and Simon HH: Engrailed
genes are cell autonomously required to prevent apoptosis in
mesencephalic dopaminergic neurons. Development. 131:3229–3236.
2004. View Article : Google Scholar
|
|
17
|
Degenhardt K, Rentschler S, Fishman G and
Sassoon DA: Cellular and cis-regulation of En-2 expression in the
mandibular arch. Mech Dev. 111:125–136. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Martin NL, Saba-El-Leil MK, Sadekova S,
Meloche S and Sauvageau G: EN2 is a candidate oncogene in human
breast cancer. Oncogene. 24:6890–6901. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bose SK, Bullard RS and Donald CD:
Oncogenic role of engrailed-2 (en-2) in prostate cancer cell growth
and survival. Transl Oncogenomics. 3:37–43. 2008.PubMed/NCBI
|
|
20
|
Morgan R, Boxall A, Bhatt A, et al:
Engrailed-2 (EN2): a tumor specific urinary biomarker for the early
diagnosis of prostate cancer. Clin Cancer Res. 17:1090–1098. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pandha H, Sorensen KD, Orntoft TF, Langley
S, Hoyer S, Borre M and Morgan R: Urinary engrailed-2 (EN2) levels
predict tumour volume in men undergoing radical prostatectomy for
prostate cancer. BJU Int. 110:E287–E292. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
McGrath SE, Michael A, Pandha H and Morgan
R: Engrailed homeobox transcription factors as potential markers
and targets in cancer. FEBS Lett. 587:549–554. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Morgan R, Bryan RT, Javed S, Launchbury F,
Zeegers MP, Cheng KK, James ND, Wallace DM, Hurst CD, Ward DG, et
al: Expression of Engrailed-2 (EN2) protein in bladder cancer and
its potential utility as a urinary diagnostic biomarker. Eur J
Cancer. 49:2214–2222. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2 (-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
25
|
Cantley LC: The phosphoinositide 3-kinase
pathway. Science. 296:1655–1657. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Luo J, Manning BD and Cantley LC:
Targeting the PI3K-Akt pathway in human cancer: rationale and
premise. Cancer Cell. 4:257–262. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shaw RJ and Cantley LC: Ras, PI(3)K and
mTOR signalling controls tumour cell growth. Nature. 441:424–430.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Peter M and Herskowitz I: Joining the
complex: cyclin-dependent kinase inhibitory proteins and the cell
cycle. Cell. 79:181–184. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Al Hussain TO and Akhtar M: Molecular
basis of urinary bladder cancer. Adv Anat Pathol. 20:53–60. 2013.
View Article : Google Scholar
|
|
30
|
Cordes I, Kluth M, Zygis D, Rink M, Chun
F, Eichelberg C, Dahlem R, Fisch M, Höppner W, Wagner W, et al:
PTEN deletions are related to disease progression and unfavourable
prognosis in early bladder cancer. Histopathology. 63:670–677.
2013.PubMed/NCBI
|
|
31
|
Adams JM and Cory S: The Bcl-2 apoptotic
switch in cancer development and therapy. Oncogene. 26:1324–1337.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Haupt S, Berger M, Goldberg Z and Haupt Y:
Apoptosis - the p53 network. J Cell Sci. 116:4077–4085. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Maehama T and Dixon JE: The tumor
suppressor, PTEN/MMAC1, dephosphorylates the lipid second
messenger, phosphatidylinositol 3,4,5-trisphosphate. J Biol Chem.
273:13375–13378. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Nicholson KM and Anderson NG: The protein
kinase B/Akt signaling pathway in human malignancy. Cell Signal.
14:381–395. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Diehl JA: Cycling to cancer with cyclin
D1. Cancer Biol Ther. 1:226–231. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kopparapu PK, Boorjian SA, Robinson BD,
Downes M, Gudas LJ, Mongan NP and Persson JL: Expression of cyclin
d1 and its association with disease characteristics in bladder
cancer. Anticancer Res. 33:5235–5242. 2013.PubMed/NCBI
|
|
37
|
Fang Y, Cao Z, Hou Q, Ma C, Yao C, Li J,
Wu XR and Huang C: Cyclin d1 downregulation contributes to
anticancer effect of isorhapontigenin on human bladder cancer
cells. Mol Cancer Ther. 12:1492–1503. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gee JR, Burmeister CB, Havighurst TC and
Kim K: Cyclinmediated G1 arrest by celecoxib differs in low-versus
high-grade bladder cancer. Anticancer Res. 29:3769–3775.
2009.PubMed/NCBI
|
|
39
|
Appelmann L, Liersch R, Kessler T, Mesters
RM and Berdel WE: Angiogenesis inhibition in cancer therapy:
platelet-derived growth factor (PDGF) and vascular endothelial
growth factor (VEGF) and their receptors: biological functions and
role in malignancy. Recent Results Cancer Res. 180:51–81. 2010.
View Article : Google Scholar
|
|
40
|
Moh MC and Shen S: The roles of cell
adhesion molecules in tumor suppression and cell migration: a new
paradox. Cell Adh Migr. 3:334–336. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Rodriguez Faba O, Palou-Redorta J,
Fernández-Gómez JM, Algaba F, Eiró N, Villavicencio H and Vizoso
FJ: Matrix metalloproteinases and bladder cancer: what is new? ISRN
Urol. 2012:5815392012.PubMed/NCBI
|
|
42
|
Yuan TL and Cantley LC: PI3K pathway
alterations in cancer: variations on a theme. Oncogene.
27:5497–5510. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Courtney KD, Corcoran RB and Engelman JA:
The PI3K pathway as drug target in human cancer. J Clin Oncol.
28:1075–1083. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
López-Knowles E, Hernández S, Malats N,
Kogevinas M, Lloreta J, Carrato A, Tardón A, Serra C and Real FX:
PIK3CA mutations are an early genetic alteration associated with
FGFR3 mutations in superficial papillary bladder tumors. Cancer
Res. 66:7401–7404. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Platt FM, Hurst CD, Taylor CF, Gregory WM,
Harnden P and Knowles MA: Spectrum of phosphatidylinositol 3-kinase
pathway gene alterations in bladder cancer. Clin Cancer Res.
15:6008–6017. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Knowles MA, Platt FM, Ross RL and Hurst
CD: Phosphatidylinositol 3-kinase (PI3K) pathway activation in
bladder cancer. Cancer Metastasis Rev. 28:305–316. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Askham JM, Platt F, Chambers PA, Snowden
H, Taylor CF and Knowles MA: AKT1 mutations in bladder cancer:
identification of a novel oncogenic mutation that can co-operate
with E17K. Oncogene. 29:150–155. 2010. View Article : Google Scholar
|
|
48
|
Calderaro J, Rebouissou S, de Koning L,
Masmoudi A, Hérault A, Dubois T, Maille P, Soyeux P, Sibony M and
de la Taille A: PI3K/AKT pathway activation in bladder
carcinogenesis. Int J Cancer. 134:1776–1784. 2014. View Article : Google Scholar
|
|
49
|
Cully M, You H, Levine AJ and Mak TW:
Beyond PTEN mutations: the PI3K pathway as an integrator of
multiple inputs during tumorigenesis. Nat Rev Cancer. 6:184–192.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Nedelec S, Foucher I, Brunet I, Bouillot
C, Prochiantz A and Trembleau A: Emx2 homeodomain transcription
factor interacts with eukaryotic translation initiation factor 4E
(eIF4E) in the axons of olfactory sensory neurons. Proc Natl Acad
Sci USA. 101:10815–10820. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zhou J, Qin L, Tien JC, Gao L, Chen X,
Wang F, Hsieh JT and Xu J: Nkx3.1 functions as para-transcription
factor to regulate gene expression and cell proliferation in
non-cell autonomous manner. J Biol Chem. 287:17248–17256. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Prin F, Serpente P, Itasaki N and Gould
AP: Hox proteins drive cell segregation and non-autonomous apical
remodelling during hindbrain segmentation. Development.
141:1492–1502. 2014. View Article : Google Scholar : PubMed/NCBI
|