Introduction
Cancer of the uterine corpus is the second most
commonly diagnosed cancer among women in the USA, following breast
cancer. Although the majority of endometrial cancers (68%) are
detected at an early stage (stage I–II), 32% of the patients have
regional or distant metastasis at diagnosis (1). Endometrioid adenocarcinoma (EAC)
accounts for three-quarters of all uterine corpus cancers. Most
primary EACs are considered to be of low risk and are usually
treatable. Frequently, however, their potential to recur and
metastasize leads to a poor prognosis with significant mortality
(2). It is therefore necessary to
identify novel therapeutic targets and treatment strategies, both
of which may be obtained through an increased understanding of the
molecular mechanisms involved in EACs.
BAG3 is one of the six BAG family proteins, which
are involved in such cellular processes as proliferation,
apoptosis, adhesion and migration. BAG3 contains a BAG domain, a WW
domain near its N-terminus and a proline-rich region (multiple PXXP
motifs) (3,4). The BAG domain binds to the ATPase
domain of the molecular chaperone HSP70, where it acts as a
nucleotide exchange factor (5). The
WW domains act as protein interaction modules that bind a
proline-rich motif, XPPXY. Recognition of their peptide ligands by
some WW domains is phosphorylation-dependent, and WW domains have
been identified in several signal transduction proteins that
interact with plasma membrane receptor complexes or with components
of the submembranous cytoskeleton (3,4).
Proline-rich regions act by binding SH3 ligand motifs (6). Previous studies have demonstrated that
BAG3 is overexpressed in several epithelial cancers, mainly
adenocarcinomas (7). In addition, a
previous study from our group showed that BAG3 induced MMP2
expression and then bound to MMP2 to positively regulate the
process of cell invasion (8).
However, little is known about the role of BAG3 in EACs.
MicroRNAs (miRNAs) are small non-coding RNAs that
function as negative regulators of gene expression by targeting
mRNAs based on their complementarity to the mRNA 3′ untranslated
region (3′-UTR) (9). A growing body
of evidence indicates that a disordered expression of miRNA
contributes to the initiation and progression of human cancers. It
has been proven, for example, that miRNAs participate in human
carcinogenesis by acting as either tumor suppressors or oncogenes
(10). In addition, because miRNAs
exhibit tissue-specific expression patterns (11), they may be of value serving as
therapeutic targets for cancer treatment and/or as biomarkers.
Recently, changes in the patterns of miRNA
expression were observed in endometrial cancer specimens collected
from several study cohorts (11,12).
In both EAC and serous adenocarcinoma, numerous miRNAs are
overexpressed or underexpressed, as compared to healthy
endometrium. In serous endometrial carcinoma, for example, the
levels of miR-10b, -29b and -455-5p are all reduced, and this
effect appears to be involved in the cancer progression (13). Among these molecules, miR-29b
reportedly acts as a tumor suppressor in various cancers, including
acute myeloid leukemia, cholangiocarcinoma and hepatocellular
carcinoma, by regulating tumor apoptosis, invasion and metastasis
through its targeting of myeloid cell leukemia 1 (Mcl-1) (14–16)
and matrix metalloproteinase 2 (MMP-2) (17). In the present study, we explored the
relationship between BAG3 and miR29b in cultured EAC cells. Using
the Ishikawa (grade 1-derived) and HEC108 (grade 3-derived) EAC
cell lines, we assessed the expression of BAG3, MMP2 and miR29-b,
and evaluated their effects on cell motility and invasiveness. Our
findings have furthered our understanding of the function and
mechanism of BAG3 and miR-29b in EACs.
Materials and methods
Cells and cell culture
Two established uterine EAC cell lines were used in
this study. The Ishikawa line was established from a grade 1
cancer, while the HEC108 line was established from a grade 3
cancer. Ishikawa cells were cultured in RPMI-1640 (Gibco, Grand
Island, NY, USA) supplemented with 10% fetal bovine serum (Hyclone,
Logan, UT, USA) and 1% penicillin/streptomycin (Gibco). HEC108
cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM)
(Gibco) supplemented with 10% FBS and 1% penicillin/streptomycin.
Both cell lines were maintained in a CO2 incubator (5%
CO2) at 37°C.
Gene silencing using a short hairpin RNA
(shRNA) vector
A gene silencing vector (pLTRH1) containing a RNA
polymerase promoter producing shRNA specific for BAG3 was used for
transfection. Oligonucleotides
[5′-GATCCCGTACCTGATGATCGAAGAGTTTCAAGAGAACTCTTCGATCATCAGGTATTTTTGGAG-3′
(sense) and
5′-TCGACTTCCAAAAAATACCTGATGATCGAAGAGTTCTCTTGAAACTCTTCGATCATCAGGTACGG-3′
(antisense)] specific for mouse bag3 were synthesized and subcloned
into the Bg1II and Sa1I sites, downstream of the H1
promoter (18). G3T-hi amphotrophic
packaging cells (Takara Bio, Shiga, Japan) were transfected with
pLTRH1bag3 puro or empty pLTRH1 puro vector according to the
manufacturer’s instructions to obtain a retro-viral supernatant,
which was added at a 1:5 ratio to DMEM supplemented with 10% FBS
and then used to infect Ishikawa or HEC108 cells. Infected cells
were then selected by incubation with 1.0 μg/ml puromycin
(Gibco) for 48 h after infection. High-responder clones to BAG3
knockdown were selected for subsequent experiments.
Real-time quantitative reverse
transcription PCR (qRT-PCR) for mRNA
Total RNA was extracted from cells using TRIzol
reagent (Invitrogen, Carlsbad, CA, USA), after which cDNA was
synthesized from 1 μg of RNA using VILO master mix
(Invitrogen). qRT-PCR was carried out using Fast SYBR Green Master
Mix (Applied Biosystems, Foster, CA, USA) in a StepOnePlus™
Real-Time PCR system (Applied Biosystems). mRNA levels were
standardized to the level of glyceraldehyde 3-phosphate
dehydrogenase (GAPDH) mRNA. The PCR protocol entailed denaturation
at 95°C for 10 min followed by 40 cycles of 95°C for 15 sec and
60°C for 60 sec. The following primers were designed and used for
qRT-PCR: for BAG3, 5′-TGAGAAGTTTAACCCCGTTGCTTGT-3′ (forward) and
5′-CCCCATCTACCCCTCCAGTCCAG-3′ (reverse); for MMP2,
5′-ACCTGGATGCCGTCGTGGAC-3′ (forward) and 5′-TGTGGCAGCACCAGGGCAGC-3′
(reverse); for GAPDH, 5′-TGAACGGGAAGCTCACTGG-3′ (forward) and
5′-TCCACCACCCTGTTGCTGTA-3′ (reverse). Gene expression was
calculated using the 2−ΔCt method.
qRT-PCR for microRNA
Total RNA was extracted using TRIzol reagent
(Invitrogen), after which reverse transcription was performed with
10 ng of total RNA using a TaqMan® MicroRNA Reverse
Transcription kit (Applied Biosystems) and sequence-specific RT
primers from the TaqMan MicroRNA assays (Applied Biosystems)
according to the manufacturer’s instructions. Separate reverse
transcription reactions were run for each TaqMan MicroRNA assay on
each RNA sample. qRT-PCR was performed with cDNA using inventoried
TaqMan MicroRNA assays and TaqMan Universal Master Mix II (Applied
Biosystems). The assay was performed in triplicate, and the PCR
amplification was performed using a StepOnePlus™ Real-Time PCR
system. Gene expression was calculated using the 2−ΔCt
method.
Lysate production
Cell lysates were produced from subconfluent cell
cultures. After scraping the cells from the dishes, they were
placed in RIPA buffer [50 mM Tris-HCl (pH 8.0), 150 mM NaCl, 0.1%
SDS, 1% NP40 and 0.5% sodium deoxycholate] containing a protease
inhibitor cocktail (1:100 dilution; Thermo Scientific, Rockford,
IL, USA). The cells were then lysed by sonication, after which the
lysates were centrifuged at 15,000 rpm for 15 min at 4°C to pellet
the nuclei. The supernatant was then collected as the cell
lysate.
Western blotting
After measuring the protein content, lysates were
diluted in 2X sample buffer [0.5 M Tris-HCl (pH 6.8), 10% SDS,
β-mercaptoethanol and 1% BPB] and boiled for 5 min at 100°C.
Samples containing 20 μg of protein were then
electrophoresed (200 V for 35 min) on 12% SDS polyacrylamide gel,
after which the separated proteins were transferred onto PVDF
membranes. After being blocked with 5% non-fat dry milk in TBS [10
mM sodium phosphate (pH 7.8), 150 mM NaCl and 0.05% Tween-20], the
membrane was probed with the following primary antibodies: rabbit
monoclonal antibodies against BAG3 (1:1,000 dilution; gift of Dr S.
Takayama), mouse monoclonal antibodies against hMMP2 (1:150
dilution; F-68; Daiichi Fine Chemical Co., Ltd., Toyama, Japan) and
mouse monoclonal antibodies against β-actin (1:5,000 dilution;
A5441; Sigma-Aldritch, St. Louis, MO, USA). After being washed with
PBS-T, the membranes were incubated with secondary horseradish
peroxidase-conjugated antibodies. The protein was visualized using
ECL Prime Western Blotting Detection Reagent and ImageQuant LAS 500
(GE Healthcare, Buckinghamshire, UK).
Cell motility and invasion assays
BAG3 knockdown cells and miR29b mimic/inhibitor
transfectants were used for motility and invasion assays. To assess
motility, the cells were seeded into the upper chambers of the
24-well Transwell chambers (Corning, Inc., Corning, NY, USA) at the
density of 5×104 cells per well and cultured in medium
containing 0.5% BSA. Medium containing 10% FBS and 10 μg/ml
of fibronectin was added to the lower chamber. The plates were then
incubated for 7 h at 37°C, after which the cells that remained on
top of the membrane were removed with a cotton swab. The cells that
had migrated through the membrane were fixed in formalin, stained
with hematoxylin and eosin (H&E) and counted in five 20x
microscope (Olympus DP70) (Olympus, Tokyo, Japan) fields. To assess
invasion, cells were incubated for 60 h at 37°C in
Corning® BioCoat™ Matrigel® Invasion Chambers
with 8.0-μm pores (Discovery Labware, Inc., Bedford, MA,
USA). The data are from three independent experiments.
Mature miRNA transfection
Using Lipofectamine RNAiMAX (Invitrogen), cells
grown in 6-cm dishes were transfected with miRNA-29b mirVana miRNA
mimic (Ambion, Austin, TX, USA) to augment miR-29b activity, or
with an inactive negative control for miRNA-29b mirVana miRNA mimic
(Ambion). Alternatively, they were transfected with a miRNA-29b
mirVana miRNA inhibitor (Ambion) to diminish miR-29b activity or a
negative control for a miRNA-29b mirVana miRNA inhibitor
(Ambion).
Luciferase assay
HEC108 cells cultured in 96-well plates were
co-transfected using 17 nM miR-29b mimic or its control plus 25 ng
of the 3′-UTR of MMP2 mRNA, which contained the target sites for
miR-29b, or empty vector. The cells were collected 48 h after
transfection and analyzed using a LightSwitch Assay System (Switch
Gear Genomics, Menlo Park, CA, USA). Luciferase activity was
detected using an Infinite M 1000 PRO microplate reader (Tecan
Group Ltd., Männedorf, Switzerland). All the experiments were
performed in triplicate.
Statistical analysis
Student’s t-tests were used for statistical
evaluation of the data. Values of P<0.05 were considered
significant. SPSS 22.0 (IBM, Armonk, NY, USA) was used in the
analysis.
Results
Expression of BAG3 and MMP2 in EAC cell
lines
Ishikawa and HEC108 cells were used for western blot
and qRT-PCR analyses. There was little difference in MMP2 mRNA
expression between the two cell lines (Fig. 1B), but expression of MMP2 protein
was detected only in HEC108 cells (Fig.
1C). Similarly, BAG3 mRNA and protein were more strongly
expressed in HEC108, than Ishikawa cells (Fig. 1A and C).
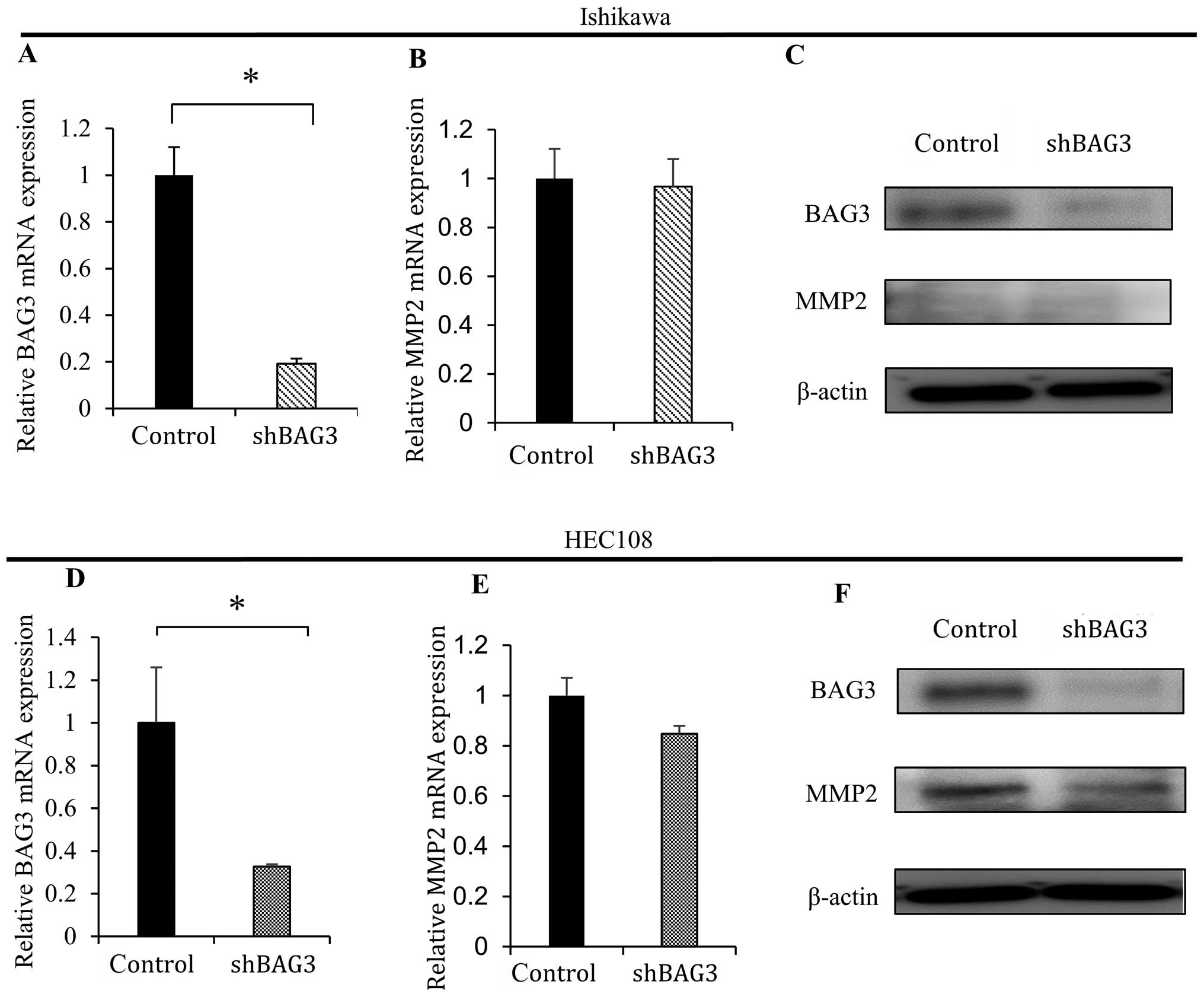
BAG3 knockdown reduces MMP2
expression
To determine the effect of reducing BAG3 expression
on MMP2 activity, Ishikawa and HEC108 cells were infected with a
retroviral vector encoding shRNA targeting BAG3 (shBAG3). In HEC108
cells, which exhibit substantial endogenous MMP2 expression, BAG3
knockdown (Fig. 2D and F, top blot)
led to reduction in the expression of MMP2 mRNA and protein
(Fig. 2E and F, middle blot). In
Ishikawa cells, which express little MMP2, knocking down BAG3
(Fig. 2A and C, top blot) had no
effect on MMP2 expression (Fig. 2B
and C, middle blot).
BAG3 knockdown reduces cell motility and
invasiveness
We next evaluated the effects of reducing endogenous
BAG3 levels on cell motility and invasiveness. To assess motility,
Transwell chambers treated with 10 μg/ml fibronectin were
loaded with Ishikawa or HEC108 cells expressing either shBAG3 or a
control sequence. As shown in Fig.
3A and B, BAG3 knockdown suppressed migration of both Ishikawa
and HEC108 cells through the Transwell filter.
To assess the role of BAG3 in determining the
invasiveness of tumor cells, Ishikawa or HEC108 cells expressing
shBAG3 were placed on a Matrigel ECM layer in a Transwell chamber,
and their rate of migration through the Matrigel was assayed. We
found that BAG3 knockdown significantly reduced the invasiveness of
the HEC108 cells, which strongly express MMP2 (Fig. 3D), but had little effect on the
invasiveness of Ishikawa cells, which express little if any MMP2
(Fig. 3C).
Reducing BAG3 expression led to increases
in miR29b levels in EAC cell lines
When we used real-time qRT-PCR to compare the levels
of miR29b expression between Ishikawa and HEC108 cells, we observed
that miR29b is more strongly expressed in Ishikawa than HEC108
cells (Fig. 4A). Moreover, knocking
down BAG3 expression using shBAG3 enhanced miR29b expression in
both Ishikawa and HEC108 cells (Fig.
4B and C).
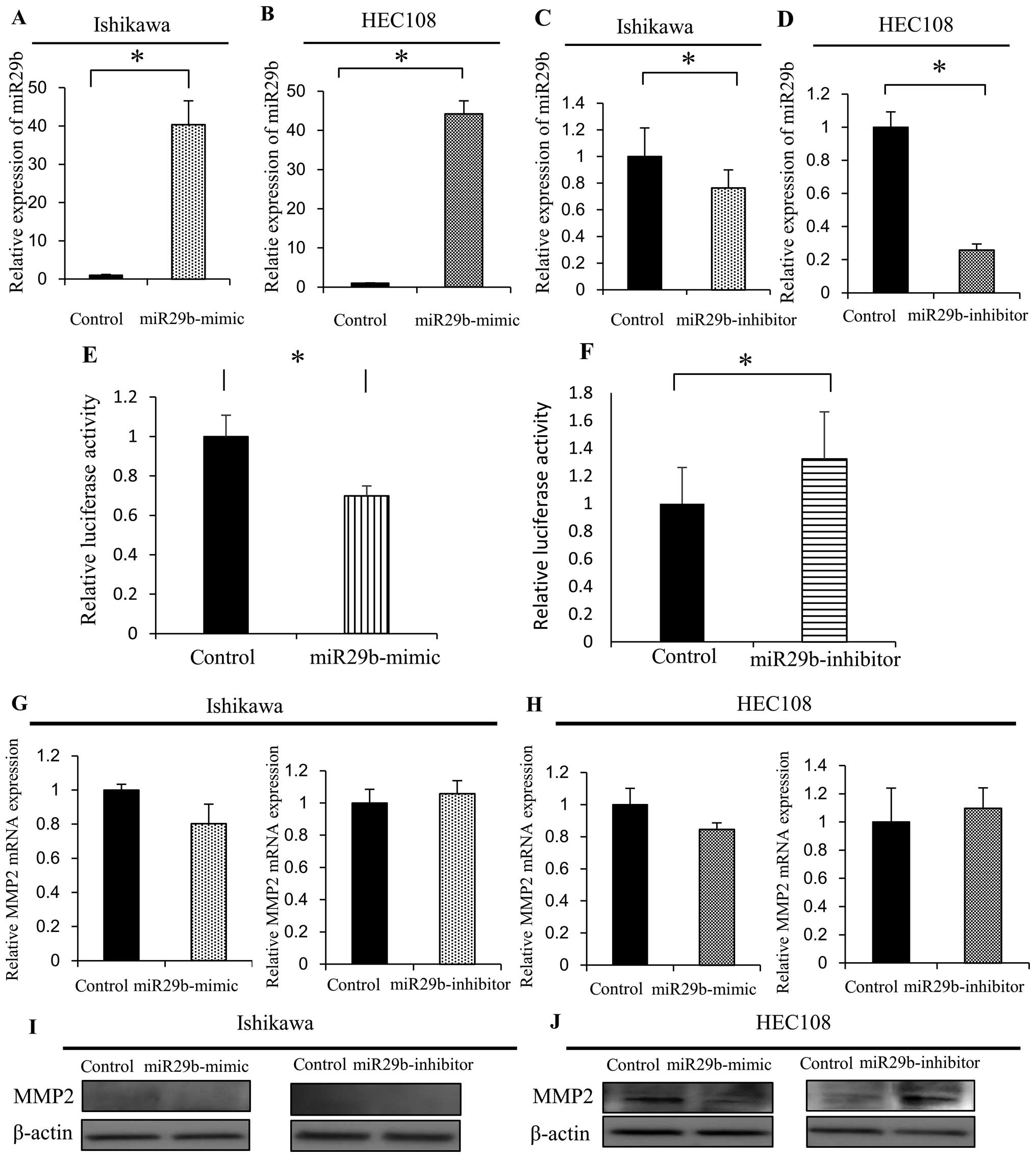
miR-29b directly targets the MMP2
3′-UTR
To further investigate the role of miR-29b in
determining the metastatic potential of EAC cells, we transfected
Ishikawa and HEC108 EAC cells with miRNA-29b mirVana miRNA mimic to
augment miR-29b activity or with miRNA-29b mirVana miRNA inhibitor
to diminish miR-29b activity. Using qRT-PCR, we observed that
miR-29b expression was markedly increased in both Ishikawa and
HEC108 cells transfected with miR-29b-mimic (Fig. 5A and B), and was diminished in both
cell types transfected with miR-29b-inhibitor, though the effect
appeared to be greater in HEC108 cells (Fig. 5C and D). Then using a luciferase
reporter system, we found that co-transfection of miR-29b-mimic
significantly suppressed the activity of a reporter gene containing
the MMP2 mRNA 3′-UTR (Fig. 5E).
Conversely, inhibiting of endogenous miR-29b using a
miR-29b-inhibitor increased the activity of the MMP2 reporter gene
(Fig. 5F). In addition, MMP2 mRNA
levels were lower than the control in Ishikawa and HEC108 cells
transfected with miR-29b-mimic and were higher than the control in
the cells transfected with miR-29b-inhibitor (Fig. 5G and H). Correspondingly, the levels
of MMP2 protein were lower than the control in the HEC108 cells
expressing miR-29b-mimic and higher than the control in the HEC108
cells expressing the miR-29b-inhibitor (Fig. 5J). In Ishikawa cells, by contrast,
levels of MMP2 protein were slightly affected by miR29b-mimic or
-inhibitor, as there was slight or no endogenous MMP2 expression in
the Ishikawa cells (Fig. 5I). All
things considered, these findings suggest miR-29b directly
suppresses MMP2 expression through its binding to the MMP2 mRNA
3′-UTR.
Cell migration and invasion is inhibited
by miR-29b in human EAC
We evaluated the effect of miR-29b in EAC cell
motility and invasiveness using migration and Matrigel invasion
assays after confirming the efficient transfection of miR-29b-mimic
or -inhibitor using qRT-PCR 48 h after transfection. The migration
assays revealed that motility of both Ishikawa and HEC108 cells was
significantly reduced by expression of miR-29b-mimic, as compared
with the miR-control (Fig. 6A and
C), and that motility was markedly increased by expression of the
miR-29b-inhibitor (Fig. 6B and
D).
Matrigel invasion assays demonstrated that the
invasiveness of cells endogenously expressing MMP2 (HEC108) was
significantly reduced by expression of miR-29b-mimic (Fig. 6G), but was markedly increased by
expression of miR-29b-inhibitor (Fig.
6H). On the other hand, expression of miR-29b-mimic or
miR-29b-inhibitor had a slight, if any, effect in Ishikawa cells,
which express a low level of MMP2 (Fig.
6E and F).
Discussion
Cell invasion is essential for cancer metastasis,
which is the major cause of cancer mortality. It is well known that
tumor metastasis is a complex process involving the loss of
cell-cell adhesion and the gain of cell-matrix adhesion as well as
the increase in the expression and activation of extracellular
proteases that degrade the ECM, enabling cell migration through the
space created. During this process, MMPs play a key role in
catalyzing proteolytic degradation of the ECM (19). Among these enzymes, MMP2 catalyzed
the degradation of type IV collagen and the overexpression of MMP2
has been observed in various types of cancers (20,21).
We found that the MMP2 expression was significantly weaker in the
Ishikawa EAC cells (grade 1) than in the HEC108 EAC cells (grade
3). In addition, we demonstrated that reducing BAG3 or varying
expression of miR-29b in HEC108 cells influenced both MMP2
expression and cell invasiveness, whereas Ishikawa cells,
expressing much less MMP2 than HEC108 cells, showed little
variation of MMP2 expression or cell invasiveness. These results
indicate that MMP2 is a key mediator of EAC cell invasion.
BAG3 is known to be an important regulator of cancer
cell migration, invasion and adhesion (7). A previous study from our group showed
that BAG3 increases the invasiveness of cultured ovarian cancer
cells by interacting with MMP2 (8).
We also found that BAG3 knockdown leads to a reduction in the MMP2
expression, and that BAG3 binds to MMP2 to positively regulate the
process of cell invasion. However, the molecular mechanism through
which BAG3 enhances MMP2 expression is not known.
The expression of oncogenes is often regulated by
miRNAs, small RNAs (21–25 nucleotides) that suppress gene
expression by interacting with their mRNAs (22). Consequently, dysfunction or
dysregulation of miRNAs can lead to the activation of oncogenes.
Because miR-29b is known to directly suppress MMP2 expression in
prostate cancer and hepatocellular carcinoma cells (17,23) we
examined the relationship between the expression of BAG3 and
miR-29b. Our results show that BAG3 suppresses miR-29b expression
in both Ishikawa and HEC108 cells. Moreover, luciferase reporter
assays revealed that miR-29b directly targets MMP2 in EAC cells. It
thus appears that inhibition of miR-29b correlates with enhanced
MMP2 expression, as well as with increases in cell motility and
invasiveness. Conversely, restoration of miR-29b reduced MMP2
expression and inhibited EAC cell motility and invasiveness. All
things considered, these results suggest that inhibition of miR-29b
by BAG3 leads to MMP2 overexpression and, in turn, increased EAC
cell invasion.
The miR-29 family contains three members (miR-29a,
-29b and -29c), which are highly similar and share the same seed
sequence. Reduced expression of the miR-29 family miRNAs has been
observed in several cancers, including acute myeloid leukemia,
non-small cell lung cancer, cholangiocarcinoma and hepatocellular
carcinoma. Moreover, reduced expression of miR-29b is significantly
correlated with poorer overall survival and poorer progression-free
survival in endometrial serous adenocarcinoma (13). However, miR-29 expression is not
always diminished in cancer, as enhanced expression of miR-29 has
been reported in both breast and colon cancer (24,25).
It will therefore be important to fully elucidate the mechanism by
which miR-29 expression is regulated, which may enable miR-29 to
serve as a therapeutic target. It has been reported that c-Myc
negatively regulates miR-29a/b1 expression at the transcription
level (26). miR-29 expression also
appears to be regulated via the NF-κB-YY1 pathway. Within that
pathway, YY1 inhibits miR-29 transcription, whereas miR-29 blocks
YY1 translation, and YY1 is also regulated by NF-κB (27). In the present study, we found that
BAG3 knockdown restores miR-29b expression in EAC cells, suggesting
that BAG3 also inhibits miR-29b expression. Though further
elucidation of molecular mechanism controlling the expression of
miR-29b via BAG3 is necessary, understanding the involvement of
BAG3 in regulating miR-29b expression may provide new insight into
the process by which miR-29b activity is suppressed in EAC.
In conclusion, our results indicate that BAG3
suppresses expression of miR-29b. The reduction in miR-29b leads to
enhanced expression of MMP-2, which in turn promotes cell migration
and invasion. This scenario suggests that the BAG3-miR-29b-MMP2
pathway is a novel means by which BAG3 promotes cell invasion and
contributes to EAC met astasis. It thus warrants further
investigation.
Acknowledgments
We would like to thank Dr S. Takayama for providing
a gene silencing vector (pLTRH1) specific for BAG3 and rabbit
anti-BAG3 antibody.
References
|
1
|
DeSantis CE, Lin CC, Mariotto AB, Siegel
RL, Stein KD, Kramer JL, Alteri R, Robbins AS and Jemal A: Cancer
treatment and survivorship statistics, 2014. CA Cancer J Clin.
64:252–271. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Barrena Medel NI, Bansal S, Miller DS,
Wright JD and Herzog TJ: Pharmacotherapy of endometrial cancer.
Expert Opin Pharmacother. 10:1939–1951. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Einbond A and Sudol M: Towards prediction
of cognate complexes between the WW domain and proline-rich
ligands. FEBS Lett. 384:1–8. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sudol M, Chen HI, Bougeret C, Einbond A
and Bork P: Characterization of a novel protein-binding module -
the WW domain. FEBS Lett. 369:67–71. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Doong H, Vrailas A and Kohn EC: What’s in
the ‘BAG’? - A functional domain analysis of the BAG-family
proteins. Cancer Lett. 188:25–32. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Doong H, Price J, Kim YS, Gasbarre C,
Probst J, Liotta LA, Blanchette J, Rizzo K and Kohn E: CAIR-1/BAG-3
forms an EGF-regulated ternary complex with phospholipase C-gamma
and Hsp70/Hsc70. Oncogene. 19:4385–4395. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Iwasaki M, Homma S, Hishiya A, Dolezal SJ,
Reed JC and Takayama S: BAG3 regulates motility and adhesion of
epithelial cancer cells. Cancer Res. 67:10252–10259. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Suzuki M, Iwasaki M, Sugio A, Hishiya A,
Tanaka R, Endo T, Takayama S and Saito T: BAG3 (BCL2-associated
athanogene 3) interacts with MMP-2 to positively regulate invasion
by ovarian carcinoma cells. Cancer Lett. 303:65–71. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nelson KM and Weiss GJ: MicroRNAs and
cancer: Past, present, and potential future. Mol Cancer Ther.
7:3655–3660. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Banno K, Yanokura M, Kisu I, Yamagami W,
Susumu N and Aoki D: MicroRNAs in endometrial cancer. Int J Clin
Oncol. 18:186–192. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Devor EJ, Hovey AM, Goodheart MJ,
Ramachandran S and Leslie KK: microRNA expression profiling of
endometrial endometrioid adenocarcinomas and serous adenocarcinomas
reveals profiles containing shared, unique and differentiating
groups of microRNAs. Oncol Rep. 26:995–1002. 2011.PubMed/NCBI
|
|
13
|
Hiroki E, Akahira J, Suzuki F, Nagase S,
Ito K, Suzuki T, Sasano H and Yaegashi N: Changes in microRNA
expression levels correlate with clinicopathological features and
prognoses in endometrial serous adenocarcinomas. Cancer Sci.
101:241–249. 2010. View Article : Google Scholar
|
|
14
|
Garzon R, Heaphy CE, Havelange V, et al:
MicroRNA 29b functions in acute myeloid leukemia. Blood.
114:5331–5341. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sugio A, Iwasaki M, Habata S, Mariya T,
Suzuki M, Osogami H, Tamate M, Tanaka R and Saito T: BAG3
upregulates Mcl-1 through downregulation of miR-29b to induce
anticancer drug resistance in ovarian cancer. Gynecol Oncol.
134:615–623. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mott JL, Kobayashi S, Bronk SF and Gores
GJ: miR-29 regulates Mcl-1 protein expression and apoptosis.
Oncogene. 26:6133–6140. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fang JH, Zhou HC, Zeng C, Yang J, Liu Y,
Huang X, Zhang JP, Guan XY and Zhuang SM: MicroRNA-29b suppresses
tumor angiogenesis, invasion, and metastasis by regulating matrix
metalloproteinase 2 expression. Hepatology. 54:1729–1740. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Homma S, Iwasaki M, Shelton GD, Engvall E,
Reed JC and Takayama S: BAG3 deficiency results in fulminant
myopathy and early lethality. Am J Pathol. 169:761–773. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Stetler-Stevenson WG, Aznavoorian S and
Liotta LA: Tumor cell interactions with the extracellular matrix
during invasion and metastasis. Annu Rev Cell Biol. 9:541–573.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ogasawara S, Yano H, Momosaki S, Nishida
N, Takemoto Y, Kojiro S and Kojiro M: Expression of matrix
metalloproteinases (MMPs) in cultured hepatocellular carcinoma
(HCC) cells and surgically resected HCC tissues. Oncol Rep.
13:1043–1048. 2005.PubMed/NCBI
|
|
21
|
Giannelli G, Bergamini C, Marinosci F,
Fransvea E, Quaranta M, Lupo L, Schiraldi O and Antonaci S:
Clinical role of MMP-2/TIMP-2 imbalance in hepatocellular
carcinoma. Int J Cancer. 97:425–431. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Negrini M, Ferracin M, Sabbioni S and
Croce CM: MicroRNAs in human cancer: From research to therapy. J
Cell Sci. 120:1833–1840. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Steele R, Mott JL and Ray RB: MBP-1
upregulates miR-29b that represses Mcl-1, collagens, and
matrix-metalloproteinase-2 in prostate cancer cells. Genes Cancer.
1:381–387. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang Y, Zhang X, Li H, Yu J and Ren X: The
role of miRNA-29 family in cancer. Eur J Cell Biol. 92:123–128.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gebeshuber CA, Zatloukal K and Martinez J:
miR-29a suppresses tristetraprolin, which is a regulator of
epithelial polarity and metastasis. EMBO Rep. 10:400–405. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mott JL, Kurita S, Cazanave SC, Bronk SF,
Werneburg NW and Fernandez-Zapico ME: Transcriptional suppression
of miR-29b-1/miR-29a promoter by c-Myc, hedgehog, and NF-kappaB. J
Cell Biochem. 110:1155–1164. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang H, Garzon R, Sun H, et al:
NF-kappaB-YY1-miR-29 regulatory circuitry in skeletal myogenesis
and rhabdomyosarcoma. Cancer Cell. 14:369–381. 2008. View Article : Google Scholar : PubMed/NCBI
|