Introduction
Human glioma remains a refractory and
life-threatening cerebral disease with poor prognosis, despite the
improvement in current available therapies, including surgery,
radiotherapy and chemotherapy. It is a histologically and
molecularly heterogeneous central nervous system (CNS) malignancy
(1). Malignant glioma accounts for
32–45% of all primary brain tumors and 70–80% of malignant cerebral
tumors (2–5); it is the second major cause of
cancer-related deaths in both children and young adults. The
overall survival time of most patients with glioma is less than two
years after diagnosis (6), with a
the median life expectancy of only 12–14 months (7) and a 5-year survival rate of less than
10% (8). This poor prognosis
illustrates the urgent need to unveil the novel molecular
mechanisms involved in glioma, with the hope of finding novel
molecular targets for the treatment of this disease.
Close associations have been found between several
cancers and eukaryotic initiation factors (eIFs) (9), which are the key factors of
translation initiation, particularly in the first steps of
translation; indeed, eIFs regulate protein synthesis, and control
cell growth, size and proliferation. Specifically, the mRNA is
activated for pre-initiation complex (PIC) binding by eIFs that
recognize the mRNA m7G cap structure at the 5′-end or the poly(A)
tail at the 3′-end (10). Among
eIFs, eIF2 brings the Met-tRNA to the 40S ribosomal subunit; the
elF4 complex stabilizes the mRNA by binding to the cap
[7-methylguanosine (m7GpppN)] (11,12);
eIF4G establishes a bridge between elF3 and elF4E (13,14);
the eIF3 complex which comprises 13 subunits (eIF3a to m) (9) serves as a scaffold to mediate
translation initiation and recognizes the first AUG initiation
codon closest to the 5′-end of the mRNA (9). It has been suggested that interactions
between eIF3 and other translation initiation factors control the
binding of the ternary complex (eIF2-GTP-methionine) to the small
(40S) ribosomal subunit, position the mRNA on the 40S subunit and
modulate the stringency of the start codon selection (15,16).
Importantly, misregulation of eIF3 expression has been implicated
in oncogenesis and the maintenance of the cancerous state (17–19),
indicating the importance of the eIF3 complex.
eIF3c (913 amino acids, located at 16p11.2), a
housekeeping gene localized in the cytoplasm, is essential for the
assembly of the eIF3 complex, as well as the general initiation
complex (20–22). In the past few years, the eIF3c gene
has been demonstrated to be essential for cell proliferation in
numerous human tumors, including testicular seminomas (23), meningiomas (24) and colon cancer cells (25). However, little is known concerning
the relationship between eIF3c and human glioma.
In the present study, we first assessed the eIF3c
expression in human glioma tissues and determined its correlation
with pathologic grades. Then, the eIF3c gene was knocked down in
the human glioma U251 cells using the RNA interference (RNAi)
technology in order to explore its functions in cell tumorigenesis,
proliferation, apoptosis and cycle. The findings presented here
provide new insights into the biological role of the eIF3c gene in
human gliomas and identify eIF3c as a potential diagnostic or
therapeutic target for human gliomas.
Materials and methods
Patients and glioma specimens
The present study included 95 Chinese patients with
cerebral glioma treated surgically at the Department of
Neurosurgery, the Second Affiliated Hospital of Hebei Medical
University between January 2008 and December 2013. There were 50
males and 45 females, aged 49.65±15.15 (ranging from 16 to 73
years). Inclusion criteria; all the specimens were obtained at the
initial surgery and the patients had not received preoperative
radiotherapy, chemotherapy or immunotherapy. The pathologic grades
of the samples were confirmed independently by two pathologists
according to the revised World Health Organization criteria of
tumors for the central nervous system (1): respectively 11, 23, 26 and 35 patients
were found with grade I–IV tumors. Normal cerebral tissues used as
controls were obtained from 31 patients suffering from severe
cranio-cerebral injury who underwent internal decompression
operation. For each resected specimen, a portion was immediately
snap-frozen in liquid nitrogen, and the remaining part was fixed
with formalin and embedded in paraffin for histological studies.
Written informed consent forms were obtained from all the patients
involved in the present study. All the experiments using human
samples were approved by the Ethics Committee of the Second
Hospital of Hebei Medical University and complied with the current
laws of China.
Immunohistochemistry staining
Immunohistochemistry (IHC) staining of pathological
sections was performed following the standard procedures. Paraffin
blocks were cut in 4-μm sections and mounted on glass
slides. After being dewaxed in xylene, the sections were rehydrated
in ethanol gradients and immerged in water. Antigen retrieval was
performed by microwave exposure in 0.01 mmol/l sodium citrate
buffer solution (pH 6.0). Afterwards, the sections were blocked
with 10% bovine serum albumin for 30 min and incubated overnight
with primary rabbit polyclonal anti-eIF3c antibodies (1:50),
followed by biotinylated secondary anti-goat antibodies (1:200) for
60 min. Then, the sections were treated with diaminobenzidine (DAB)
activated with H2O2, used according to the
manufacturer’s instructions (Vector Laboratories). The slides were
finally counterstained with hematoxylin and examined using a Leica
DM1000 microscope (Leica, Germany).
Assessment of eIF3c gene expression in 95
glioma tissue samples and 31 normal cerebral specimens by real-time
quantitative PCR
Total RNA was extracted from the clinical specimens
(glioma and normal cerebral tissues) using TRIzol reagent
(Invitrogen, USA) according to the manufacturer’s instructions.
Total RNA (2 μg) was reverse-transcribed with M-MLV reverse
transcriptase (Promega, USA) and oligo(dT) primers (Sangon,
Shanghai). cDNA (1 μl) was used for real-time PCR, which was
performed to detect eIF3c using SYBR Master Mixture (Takara, Japan)
according to the manufacturer’s protocol. Sequences of eIF3c and
GAPDH primers were as follows: GAPDH (internal control forward,
5′-TGACTTCAACAGCGACACCCA-3′ and reverse,
5′-CACCCTGTTGCTGTAGCCAAA-3′; eIF3c forward,
5′-AGATGAGGATGAGGATGAGGAC-3′ and reverse,
5′-GGAATCGGAAGATGTGGAACC-3′.
Real-time PCR was carried out with initial
denaturation at 95°C for 15 sec, followed by 45 cycles of 95°C for
5 sec and 60°C for 30 sec. Data were analyzed using GraphPad Prism
4.0 software. The 2−ΔΔCt method was used to quantitate
the relative gene expression.
Cell culture
Human glioma cells (U87, U251, A172 and U373) were
purchased from the National Platform of Experimental Cell Resources
for Sci-Tech (Shanghai, China). The cells were maintained in
Dulbecco’s modified Eagle’s medium (D-MEM) supplemented with 10%
fetal bovine serum (FBS), 2 mM L-glutamine (all from Gibco, USA),
penicillin (100 U/ml) and streptomycin (100 μg/ml)
(Gen-View, USA). All the cells were cultured at 37°C in a
humidified atmosphere containing 5% CO2.
Assessment of eIF3c gene expression in
four human glioma cell lines (U87, U251, A172 and U373) by
semi-quantitative RT-PCR
Total RNA from the human glioma cell lines U87,
U251, A172 and U373 was extracted under RNase-free conditions using
TRIzol reagent according to the manufacturer’s instructions. For
semi-quantitative RT-PCR analysis, glyceraldehyde-3-phosphate
dehydrogenase (GAPDH) was used as an internal reference. Total RNA
(2 μ) from each sample was reverse transcribed to
single-stranded cDNA. cDNA (1 μg) was used as template for
PCR with the following primers: eIF3c forward,
5′-AGATGAGGATGAGGATGAGGAC-3′ and reverse,
5′-GGAATCGGAAGATGTGGAACC-3′, product size of 175 bp; GAPDH forward,
5′-TGACTTCAACAGCGACACCCA-3′ and reverse,
5′-CACCCTGTTGCTGTAGCCAAA-3′, product size of 121 bp. The
semi-quantitative RT-PCR was carried out with initial denaturation
at 95°C for 15 sec followed by 22 cycles of 95°C for 5 sec, and
60°C for 30 sec. PCR products were separated on a 2% agarose gel
and analyzed by imaging.
Construction and transfection of the
eIF3c-siRNA lentivirus
The sequences used for the eIF3c-siRNA and scrambled
siRNA were: 5′-GAC CAT CCG TAA TGC CAT GAA-3′ and 5′-TTC TCC GAA
CGT GTC ACG T-3′, respectively. These nucleotide sequences were
inserted into the plasmid using the siRNA expressing vector
pGCSIL-GFP and lentivirus packing eIF3c-siRNA Lentivector
Expression Systems (both from GeneChem, Shanghai, China). The
identities of the generated lentiviral based siRNA expressing
vectors were confirmed by DNA sequencing. Human renal epithelial
293T cells were infected with eIF3c-siRNA lentivirus and scramble
siRNA lentivirus (negative control) and the interference efficiency
was determined by western blot analysis.
Human glioma U251 cells were infected with
eIF3c-siRNA lentivirus and scrambled siRNA lentivirus (negative
control) at a multiplicity of infection (MOI) of 100.
Non-transfected cells were also included as controls. After three
days of infection, GFP expression was observed by fluorescence
microscopy. After five days of infection, the cells were harvested
to determine knockdown efficiency by real-time quantitative
PCR.
Western blot analysis
Western blot analysis was performed to evaluate the
eIF3c expression levels in the eIF3c-siRNA lentivirus-infected 293T
cells compared to the scrambled siRNA lentivirus (negative
control)-infected cells. For protein isolation, the cells were
washed with cold phosphate-buffered saline (PBS) and lysed with
radio-immunoprecipitation assay (RIPA) buffer [100 mmol/l Tris-HCl
(pH 6.8), 2% nonidet P-40, 4% sodium dodecyl sulfate (SDS)].
Proteins were separated by SDS-PAGE, transferred onto PVDF
membranes (Amersham, USA), probed with anti-eIF3c antibody (1:200;
Abcam, USA), and detected using the electrochemiluminescence (ECL)
kit (Amersham). Bands were obtained after exposure to X-ray film.
GAPDH was used as a control and was detected by anti-GAPDH antibody
(Santa Cruz Biotechnology, USA). The bands on the X-ray film were
quantified with an ImageQuant densitometric scanner (Molecular
Dynamics, USA).
BrdU cell proliferation assay
The U251 cells infected with lentivirus expressing
eIF3c-siRNA or scrambled siRNA (negative control) were cultured for
48 h. Cell proliferation was assessed with the BrdU Cell
Proliferation ELISA kit (Roche Applied Science, Switzerland)
according to the manufacturer’s instructions. The cells were seeded
at appropriate density into 96-well plates and cultured for 1–4
days. During the final 2–24 h, BrdU reagents were diluted at 1:100
and added to the cells (10 μl/well). Then, the cells were
fixed with FixDenat (200 μl/well) for 30 min and blocked
with 5–10% BSA for 30 min at room temperature. Anti-BrdU-POD
antibody was added (100 μl/well) to the cells for 90 min at
room temperature. After three washes with washing buffer (200–300
μl/well), the substrate solution (100 μl/well) was
added and incubated for 5–30 min in the dark. Color was developed
with 10% H2SO4 (50 μl/well) for 30 min and
BrdU amounts were determined at 450 nm on an ELx800 Absorbance
Mircoplate Reader (BioTek, USA).
Colony formation assay
The U251 cells that were transfected with
eIF3c-siRNA lentivirus or scrambled siRNA lentivirus (negative
control) for five days were harvested and seeded in 6-well plates
at a density of 500 cells/well. The medium was changed every three
days. After two weeks of culture, the cells were fixed with 4%
paraformaldehyde and stained by adding freshly prepared diluted
Giemsa stain for 20 min. Finally, the cells were rinsed with
distilled water and colonies with >50 cells were counted by
fluorescence microscopy (Olympus IX71, Japan).
Cell cycle analysis
The evaluation of cell cycle distribution was
carried out using flow cytometry. The U251 glioma cells infected
with lentivirus expressing eIF3c-siRNA or scrambled siRNA (negative
control) were incubated for 96 h with conditioned medium. Then, the
cells were resuspended, seeded in 6 cm dishes, and grown until ~80%
confluency. Afterwards, the cells were harvested and fixed with 70%
ice-cold alcohol for at least 1 h. After being washed with PBS, the
cells were treated with the staining solution containing propidium
iodide (PI), RNase and PBS. Finally, the cells were filtered
through a 50-μm nylon mesh and cell cycle profiles were
analyzed by flow cytometry (FACSCalibur; Becton-Dickinson, USA). At
least 1×106 cells/sample were prepared for cell cycle
analysis and triplicate experiments were performed.
Evaluation of cell apoptosis
Apoptosis in the cells was assessed using the
Annexin V-APC apoptosis detection kit (BioVision, USA). The U251
cells infected with lentivirus expressing eIF3c-siRNA or scrambled
siRNA (negative control) were incubated for five days, harvested
and washed with PBS. Then, the cells at a final density of
1×106–1×107/ml were resuspended in staining
buffer. For the apoptosis assessment, 100 μl of cell
suspension were mixed with 5 μl of Annexin V solution at
room temperature in the dark for 10–15 min. Finally, cell apoptosis
was analyzed by flow cytometry (FACSCalibur). All the assays were
performed in triplicates.
Statistical analysis
Data were analyzed using the statistical package SAS
version 8 software (SAS, USA). All the data were expressed as mean
± standard deviation (SD). Significant differences between the
groups were analyzed by a Chi-square test and the one-way ANOVA
followed by the Student-Newman-Keuls (SNK) test. P<0.05 was
considered to indicate a statistically significant result.
Results
eIF3c is overexpressed in glioma and
associated with pathologic grade
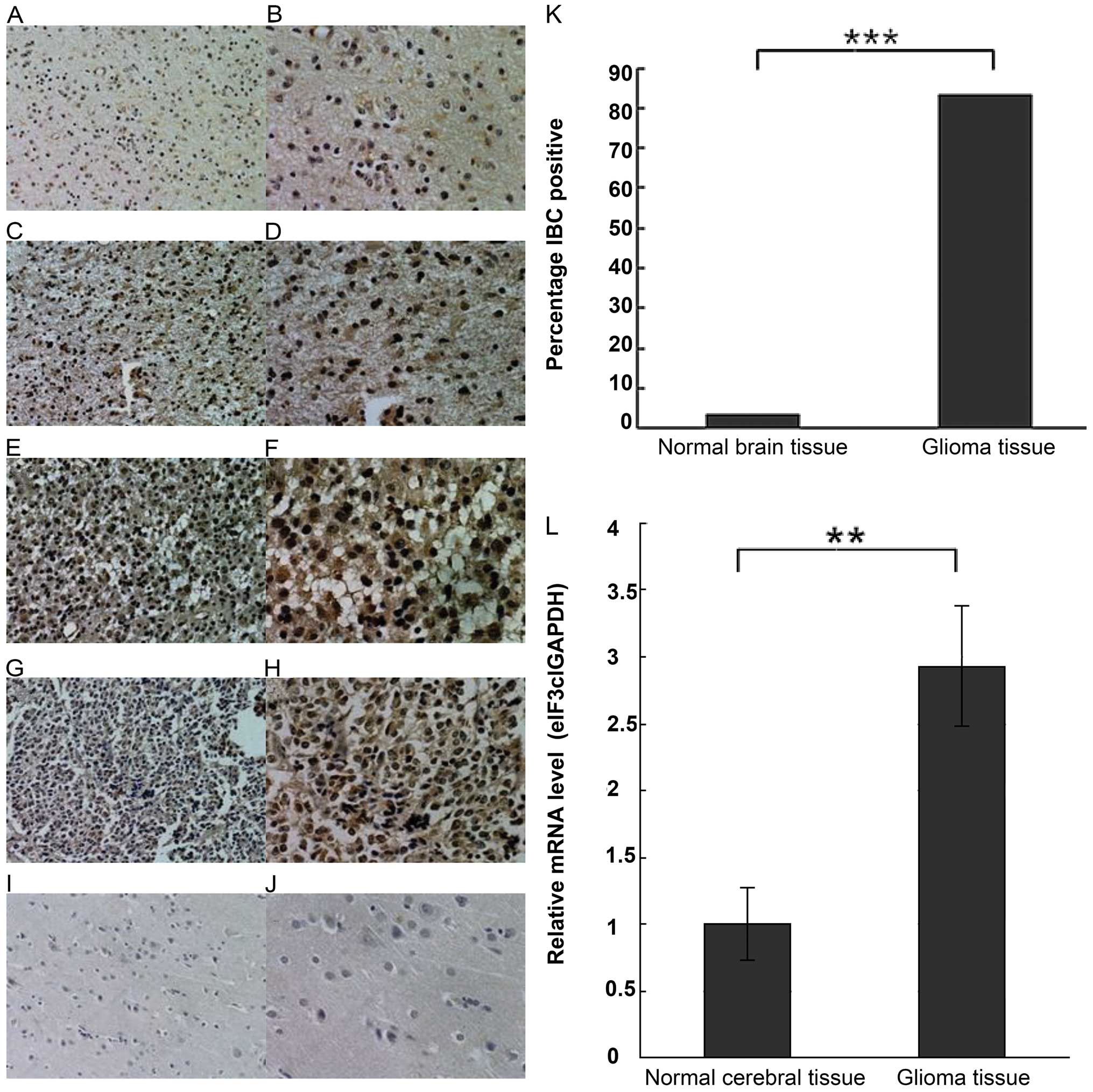
As shown in Fig. 1,
the eIF3c protein was overexpressed in the human glioma samples,
yet hardly detected in the normal brain specimens. In
immunopositive glioma cells, the labeling was primarily cytoplasmic
as observed by light microscopy. The positive expression rate of
the eIF3c protein was significantly different
(χ2=55.0385, P<0.0001) between the human glioma
(83.16%, 79/95) and the normal brain (3.23%, 1/31) tissues. In
addition, a significant difference (χ2=9.0958, P=0.0026)
was found between the low grade (grade I and II, 23/34, 67.65%) and
the high grade (grade III and IV, 56/61, 91.80%) gliomas. Notably,
no significant correlation between eIF3c protein level and gender,
age, tumor size and site was found (all P>0.05). Representative
images of the eIF3c immunostaining are shown in Fig. 1A–K, and the related results are
summarized in Tables I and II.
 | Table IExpression of eIF3c in human gliomas
and normal brain samples by IHC staining. |
Table I
Expression of eIF3c in human gliomas
and normal brain samples by IHC staining.
| Group | Case no. | Expression status of
eIF3c
| Percentage (%) | χ2 | P-value |
|---|
| Negative | Positive |
|---|
| Glioma tissues | 95 | 16 | 79 | 83.16 | 55.0385 | <0.0001 |
| Normal brain
tissues | 31 | 30 | 1 | 3.23 | | |
| Table IIRelationship between eIF3c expression
and clinicopathological parameters (cases,%) by IHC staining. |
Table II
Relationship between eIF3c expression
and clinicopathological parameters (cases,%) by IHC staining.
| Clinicopathological
parameters | Case no. | eIF3c expression
pattern
| Positive percentage
(%) | χ2 | P-value |
|---|
| Negative | Positive |
|---|
| Age (years) |
| <50 | 39 | 7 | 32 | 82.05 | 0.0578 | 0.8099 |
| ≥50 | 56 | 9 | 47 | 83.93 | | |
| Gender |
| Male | 50 | 8 | 42 | 84.00 | 0.0534 | 0.8172 |
| Female | 45 | 8 | 37 | 82.22 | | |
| Tumor diameter
(cm) |
| <3 | 43 | 6 | 37 | 86.05 | 0.4680 | 0.4939 |
| ≥3 | 52 | 10 | 42 | 80.77 | | |
| Glioma grades |
| I+II | 34 | 11 | 23 | 67.65 | 9.0958 | 0.0026 |
| III+IV | 61 | 5 | 56 | 91.80 | | |
Glioma tissues and cells display higher
eIF3c mRNA expression
Real-time quantitative PCR data showed that eIF3c
mRNA levels were significantly higher in human gliomas compared
with the normal cerebral tissues (P<0.01, Fig. 1L).
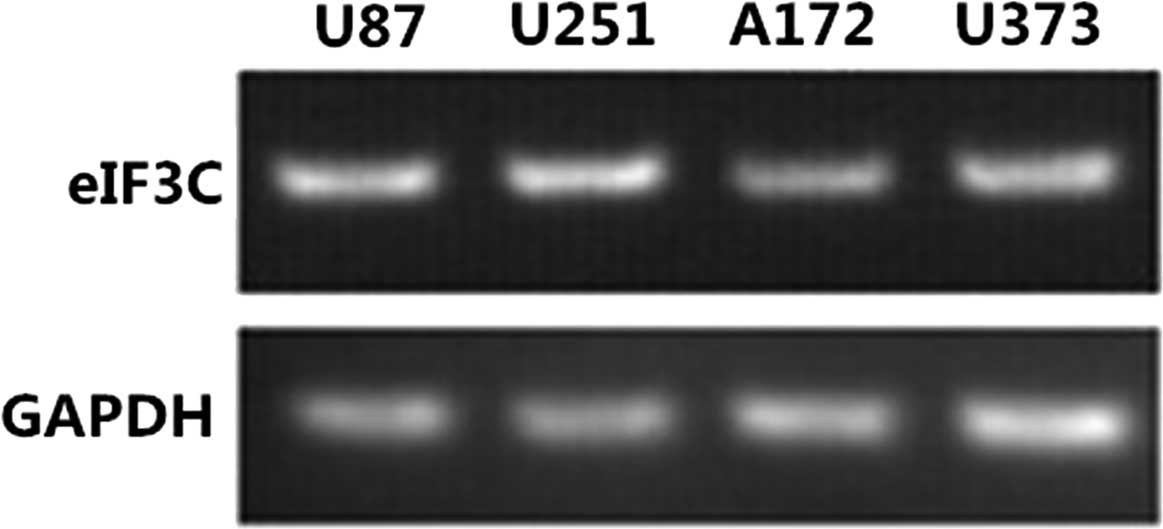
In addition, semi-quantitative RT-PCR revealed the
presence of the eIF3c gene in all the human glioma cell lines
tested, including U87, U251, A172 and U373 (Fig. 2).
eIF3c-siRNA delivery results in decreased
eIF3c expression in 293T and U251 cells
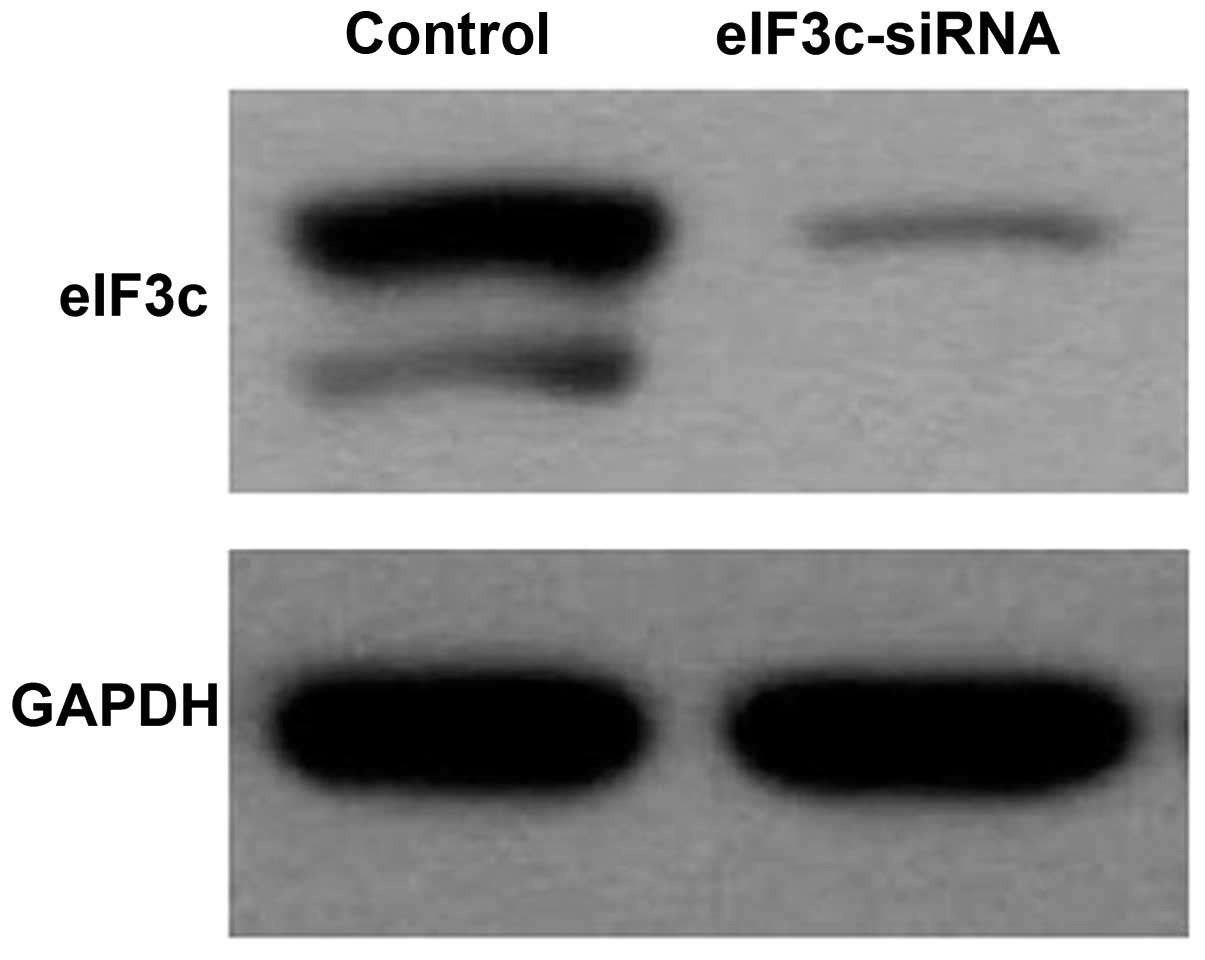
Human renal epithelial 293T cells were infected with
eIF3c-siRNA lentivirus or scrambled siRNA lentivirus (negative
control). As shown in Fig. 3, the
eIF3c protein levels detected by western blot analysis were
markedly decreased in eIF3c-siRNA infected cells compared with
those infected with scrambled siRNA. These data indicated that the
eIF3c gene was effectively silenced.
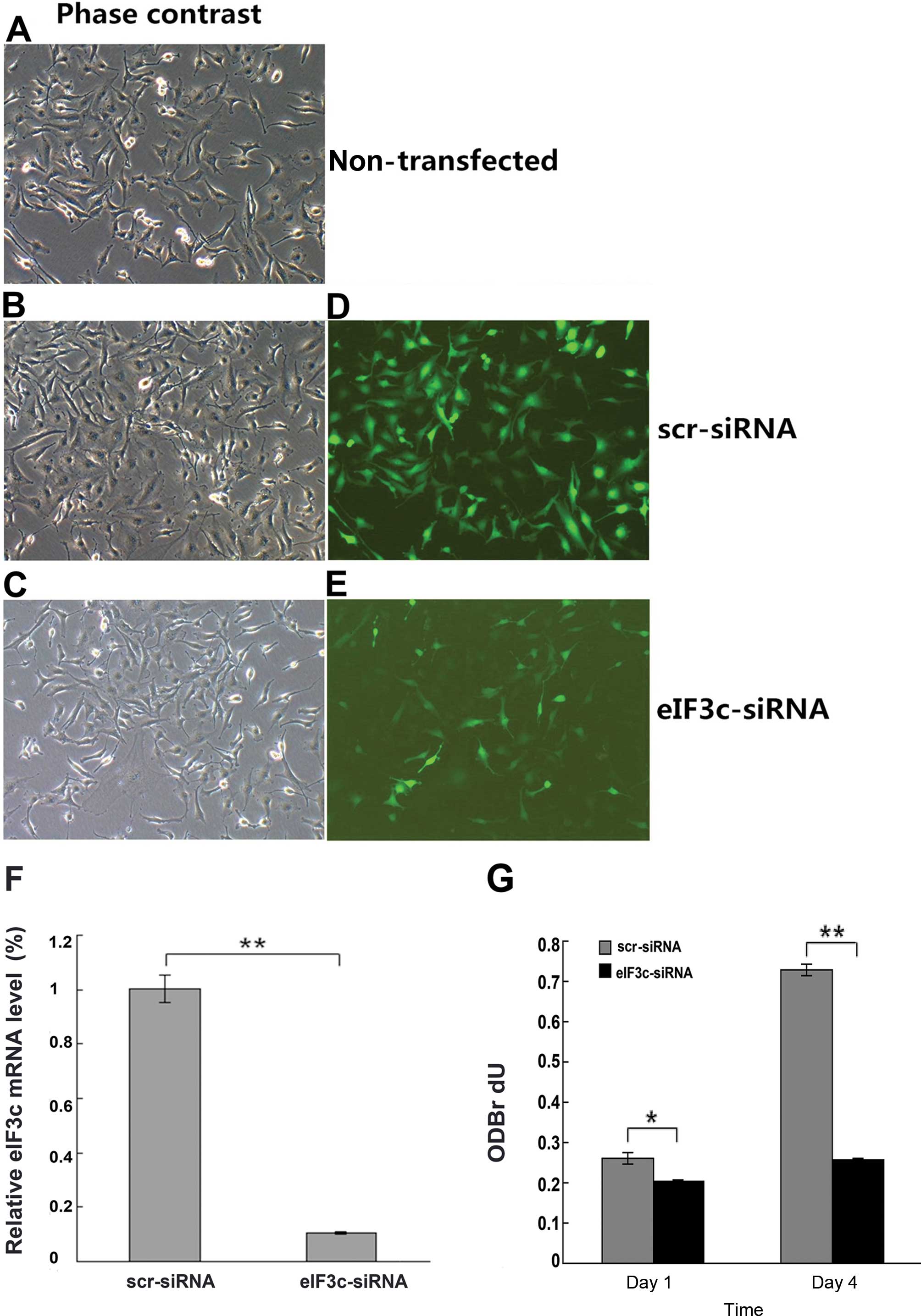
The successful transfection of eIF3c-siRNA or
scrambled siRNA into the U251 cells was confirmed by microscopic
green fluorescence detection and real-time PCR. As shown in
Fig. 4A–E, transfection efficiency
was >80% by 3 days after infection for both eIF3c-siRNA and
scrambled siRNA sequences. As shown in Fig. 4F, eIF3c-siRNA treatment resulted in
markedly downregulated gene expression of eIF3c, i.e. by 80%,
compared with the scrambled siRNA group. Therefore, the high
expression of eIF3c in U251 cells was suppressed by siRNA
infection.
eIF3c silencing results in decreased
proliferation of the U251 cells (BrdU assay)
In order to determine the eIF3c function on the cell
growth, the U251 cells expressing either eIF3c-siRNA or scrambled
siRNA sequences were analyzed by BrdU incorporation. The amounts of
DNA synthesized decreased significantly on days 1 and 4 after
infection with eIF3c-siRNA lentivirus (P<0.01, Fig. 4G) compared with the scrambled siRNA
lentivirus (negative control) group. These results indicated that
cell proliferation and DNA synthesis were significantly suppressed
by silencing eIF3c in the human glioma U251 cells.
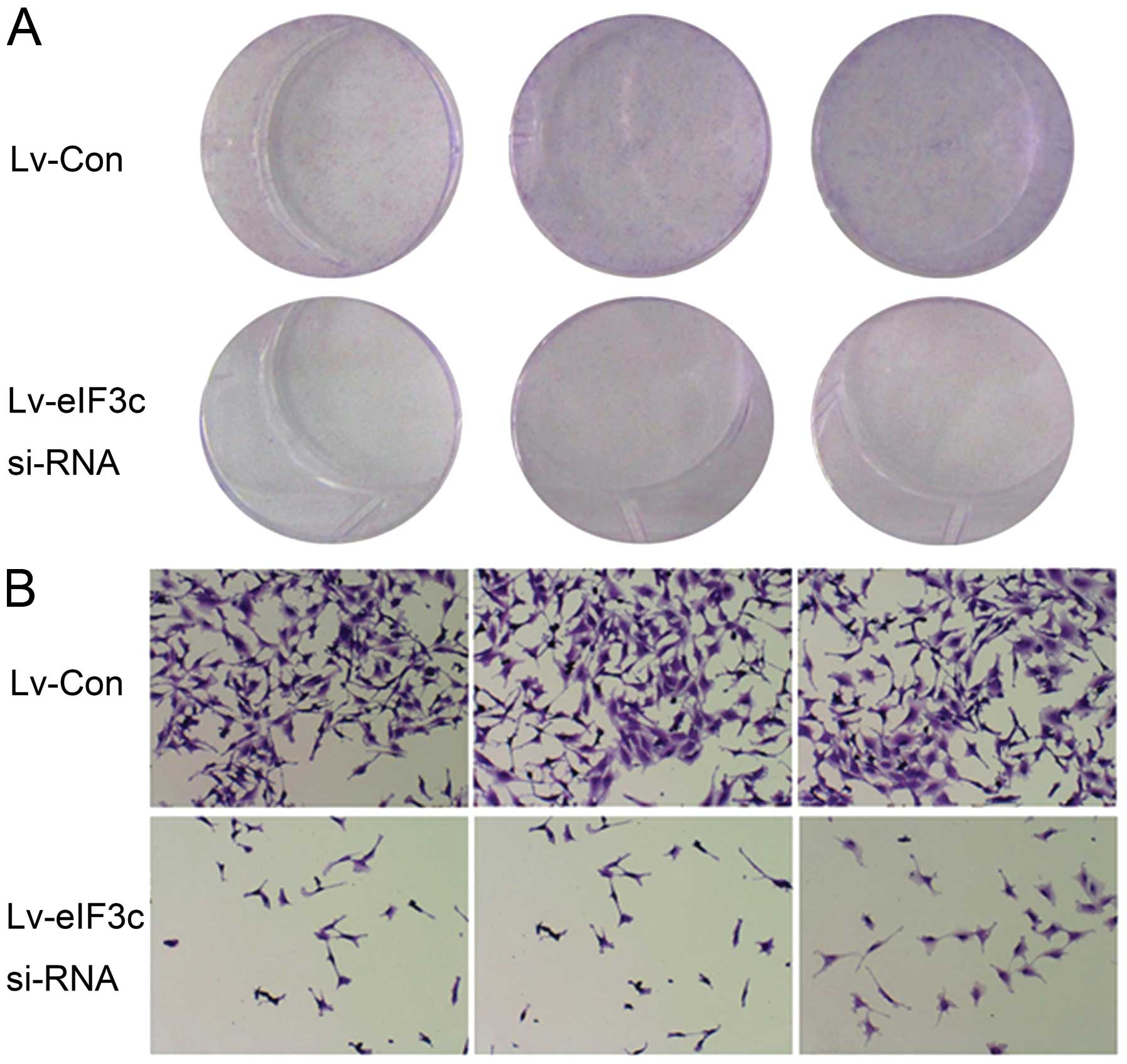
eIF3c silencing decreases U251 cell
proliferation as assessed by colony formation assay
As shown in Fig. 5,
no typical clone formation was achieved, indicating that the U251
cells have poor ability to form colonies. However, U251 cell
numbers decreased significantly in eIF3c-siRNA infected-cells when
compared with the scrambled siRNA (negative control) group. These
results further indicated that eIF3c gene knockdown resulted in
decreased U251 cell proliferation.
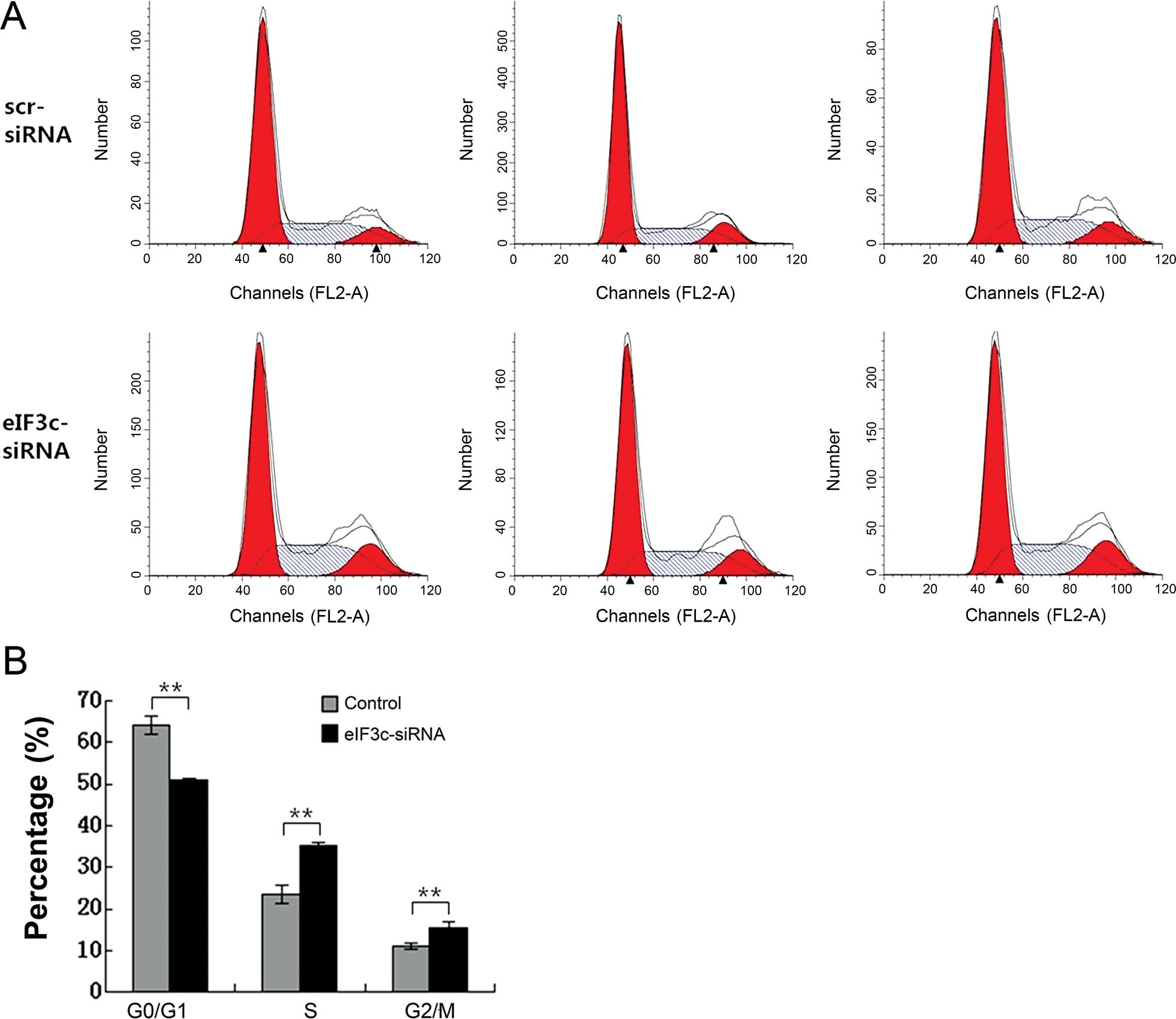
eIF3c gene silencing results in cell
cycle arrest in the U251 cells
We assessed the role of eIF3c in cell cycle
progression of the human glioma by PI-FACS in U251 cells (Fig. 6A). In the scrambled siRNA (negative
control) group, 64.05±2.20, 23.54±2.31 and 11.08±0.69% cells were
found in the G0/G1, S and G2/M phases, respectively; in the
eIF3c-siRNA group, 50.83±0.55, 35.26±0.82 and 15.64±1.38% cells
were in the G0/G1, S and G2/M, respectively. As shown in Fig. 6B, eIF3c-siRNA lentivirus cultures
displayed a significant increase in the percentage of S (P<0.01)
and G2 (P<0.01 phase cells, concomitantly with a significant
decrease in G1 phase cells (P<0.01). Therefore, the eIF3c gene
is closely related to U251 cell cycle distribution.
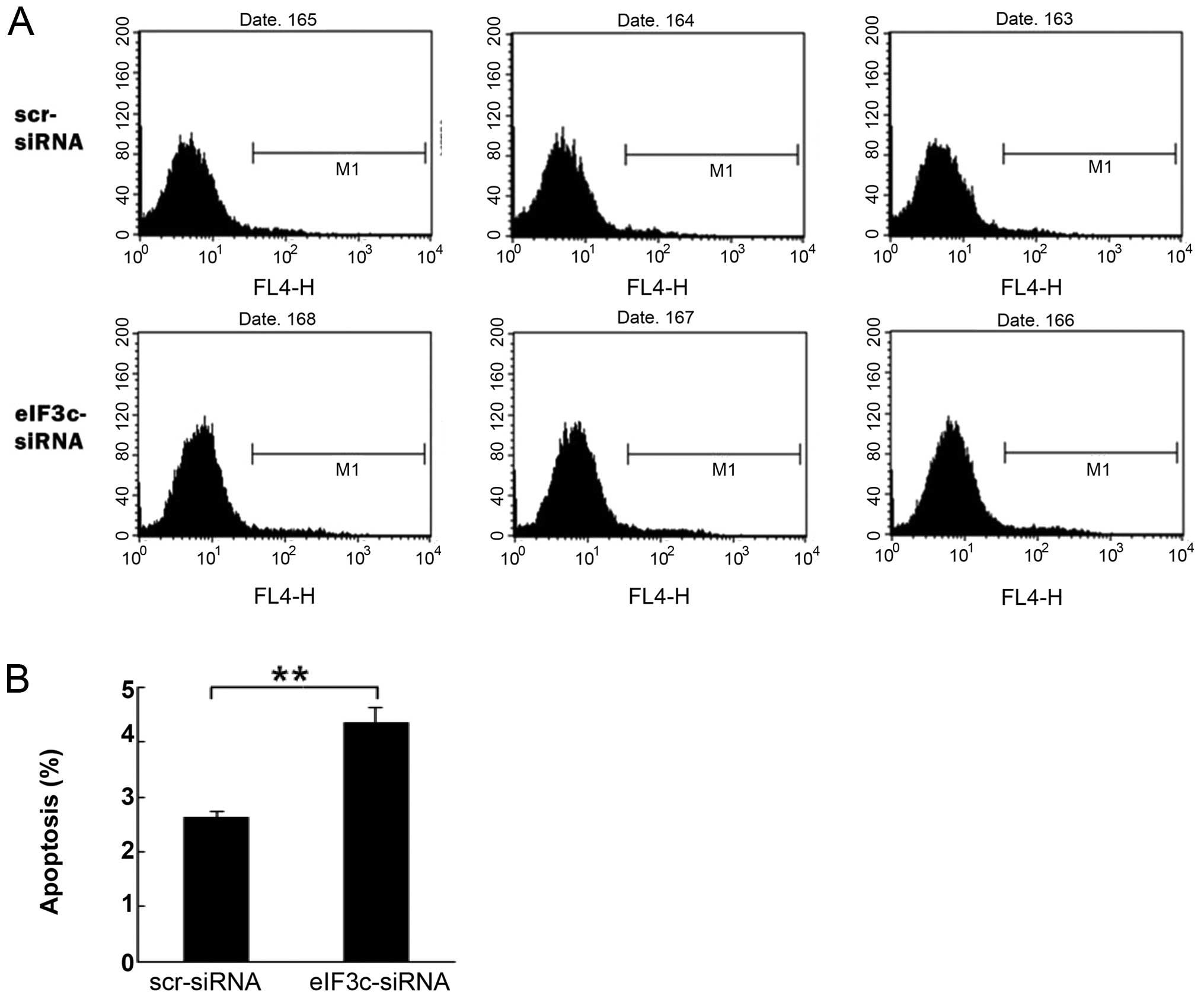
The eIF3c gene knockdown induces
apoptosis in human glioma U251 cells
Cell apoptosis was assessed by Annexin V staining
and flow cytometry (Fig. 7A). As
shown in Fig. 7B, cell apoptosis
was significantly increased in the eIF3c-siRNA group when compared
with the cells that were transfected with scrambled siRNA
(4.34±0.27 vs. 2.62±0.11, P<0.01). Therefore, knockdown of the
eIF3c gene induced apoptosis in human glioma U251 cells. These data
suggested a major role for the eIF3c gene in U251 cell survival; in
its absence, decreased survival rates resulted from apoptosis
induction.
Discussion
Malignant glioma, one of the most common and fatal
types of primary brain tumors in humans (26), arises in a multistep process which
relates to sequential and cumulative genetic alterations resulting
from intrinsic and environmental carcinogenic factors. Gene
expression is regulated at multiple levels, including the
translation of mRNAs into the proteins. Protein translation is
known to be the critical step of gene expression and translational
regulation for normal cell homeostasis and physiology (10). Remarkable progress has been made in
understanding the role of mRNA translation and protein synthesis in
human types of cancers. Studies have described that the abnormal
mRNA transcription and protein synthesis play a key role in the
tumorigenesis pathways (10). In
the present study, we showed that eIF3c was expressed in initial
surgery specimens from 95 patients with glioma and four glioma cell
lines. Knockdown of eIF3c by siRNA subsequently resulted in
decreased proliferation, increased cycle arrest and induced
apoptosis in the U251 cells. These results picture eIF3c as a
tumor-related gene in glioma cells and suggest this gene to be a
potential target for anti-glioma therapy.
To date, only few studies have investigated the
relationship between eIF3c and tumorigenesis, showing that eIF3c
gene expression is upregulated in several tumor types including
colon cancer (25), testicular
seminomas (23), meningiomas
bearing mutation in the tumor suppressor protein neurofibromatosis
2 (NF2) (25) and immortal
fibroblast cells (27). After
exploring the therapeutic potential of the eIF3c gene in five
different cancer cell lines [NCI-ADR/RES (NAR), HeLa, MCF7, HCT116
and B16F10] and analyzing the polysome profile following
downregulation of eIF3c gene expression in NAR cells, Emmanuel
et al found that eIF3c regulated cell cycle progression;
indeed, eIE3c knockdown caused polysome run-off and resulted in
cell death, justifying the conclusion that the translation
machinery was inhibited at the initiation stage and presenting
eIF3c as an anticancer target in different malignancies (20). However, in human glioma, the
expression, function and molecular mechanisms of the eIF3c gene
have not been reported and remain largely unknown.
In the present study, we found significantly higher
eIF3c expression in human glioma tissues. In addition, the positive
expression rate of eIF3c in high glioma grades (WHO III and IV) was
significantly higher than in low grades (WHO I and II)
(P<0.001). It is therefore possible that eIF3c plays a critical
role in the survival of the glioma cells as described above with
other types of cancer, likely by promoting translation
initiation.
In agreement, eIF3c mRNA were detected in all four
human glioma cell lines studied, including U87, U251, U373 and
A172, again suggesting a critical role for this gene in glioma. To
further characterize the eIF3c gene function in the glioma, we used
RNAi to knockdown this gene in human glioma U251 cells. Compared to
the control group, eIF3c-siRNA cells displayed reduced
proliferation, decreased S phase cell numbers, increased G1 phase
rates and enhanced apoptosis. These findings suggest that eIF3c may
be related to cell cycle checkpoints in the U251 cells. Indeed, the
S phase represents a critical period for cells to commit to
proliferation or undergo growth arrest (18). Therefore, these findings demonstrate
that silencing the eIF3c subunit causes a dramatic reduction of
translation initiation in glioma cells. Thus, the eIF3c gene plays
an important role in promoting U251 cell growth and is
significantly associated with U251 cell cycle distribution.
Clearly, the important oncogene eIF3c overexpressed in human
gliomas plays a critical role in proliferation and apoptosis of the
glioma cells through translational control in the protein
translation initiation phase.
Notably, a few studies have assessed miRNA profiling
in gliomas and found that miRNA regulates various cancer-associated
genes and oncogenic functions in gliomas (28). It is reasonably deduced that eIF3c
by dysregulating translational initiation with other
cancer-associated genes correlates significantly with
tumorigenesis, proliferation and apoptosis in human gliomas.
Therefore, eIF3c may be a key regulator of human gliomas and is a
novel and attractive therapeutic target to be used for designing
anticancer therapies. Inhibition of eIF3c may help substantially to
improve the clinical outcome and prognosis of patients with
gliomas.
References
|
1
|
Louis DN, Ohgaki H, Wiestler OD, Cavenee
WK, Burger PC, Jouvet A, Scheithauer BW and Kleihues P: The 2007
WHO classification of tumours of the central nervous system. Acta
Neuropathol. 114:97–109. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wakabayashi T: Clinical trial updates for
malignant brain tumors. Rinsho Shinkeigaku. 51:853–856. 2011.In
Japanese. View Article : Google Scholar
|
|
3
|
Ostrom QT, Gittleman H, Farah P, Ondracek
A, Chen Y, Wolinsky Y, Stroup NE, Kruchko C and Barnholtz-Sloan JS:
CBTRUS statistical report: Primary brain and central nervous system
tumors diagnosed in the United States in 2006–2010. Neuro Oncol1.
5(Suppl 2): ii1–ii56. 2013.
|
|
4
|
Omuro A and DeAngelis LM: Glioblastoma and
other malignant gliomas: A clinical review. JAMA. 310:1842–1850.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ohgaki H: Epidemiology of brain tumors.
Cancer Epidemiology. Springer; pp. 323–342. 2009, View Article : Google Scholar
|
|
6
|
Grossman SA, Ye X, Piantadosi S, Desideri
S, Nabors LB and Rosenfeld M: Survival of patients with newly
diagnosed glioblastoma treated with radiation and temozolomide in
research studies in the United States. Clin Cancer Res.
16:2443–2449. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ballman KV, Buckner JC, Brown PD, Giannini
C, Flynn PJ, LaPlant BR and Jaeckle KA: The relationship between
six-month progression-free survival and 12-month overall survival
end points for phase II trials in patients with glioblastoma
multiforme. Neuro Oncol. 9:29–38. 2007. View Article : Google Scholar
|
|
8
|
Grossman S, Ye X, Piantadosi S, Desideri
S, Nabors L and Rosenfeld M: Current survival statistics for
patients with newly diagnosed glioblastoma treated with radiation
and temozolomide on research studies in the United States. J Clin
Oncol. 27:20032009. View Article : Google Scholar
|
|
9
|
Zhou M, Sandercock AM, Fraser CS, Ridlova
G, Stephens E, Schenauer MR, Yokoi-Fong T, Barsky D, Leary JA,
Hershey JW, et al: Mass spectrometry reveals modularity and a
complete subunit interaction map of the eukaryotic translation
factor eIF3. Proc Natl Acad Sci USA. 105:18139–18144. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sonenberg N and Hinnebusch AG: Regulation
of translation initiation in eukaryotes: Mechanisms and biological
targets. Cell. 136:731–745. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li BD, Liu L, Dawson M and De Benedetti A:
Overexpression of eukaryotic initiation factor 4E (eIF4E) in breast
carcinoma. Cancer. 79:2385–2390. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lamphear BJ, Kirchweger R, Skern T and
Rhoads RE: Mapping of functional domains in eukaryotic protein
synthesis initiation factor 4G (eIF4G) with picornaviral proteases.
Implications for cap-dependent and cap-independent translational
initiation. J Biol Chem. 270:21975–21983. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Morley SJ, McKendrick L and Bushell M:
Cleavage of translation initiation factor 4G (eIF4G) during
anti-Fas IgM-induced apoptosis does not require signalling through
the p38 mitogen-activated protein (MAP) kinase. FEBS Lett.
438:41–48. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Damoc E, Fraser CS, Zhou M, Videler H,
Mayeur GL, Hershey JW, Doudna JA, Robinson CV and Leary JA:
Structural characterization of the human eukaryotic initiation
factor 3 protein complex by mass spectrometry. Mol Cell Proteomics.
6:1135–1146. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hinnebusch AG and Lorsch JR: The mechanism
of eukaryotic translation initiation: New insights and challenges.
Cold Spring Harb Perspect Biol. 4:42012. View Article : Google Scholar
|
|
16
|
Seal SN, Schmidt A and Marcus A: Ribosome
binding to inosine-substituted mRNAs in the absence of ATP and mRNA
factors. J Biol Chem. 264:7363–7368. 1989.PubMed/NCBI
|
|
17
|
Silvera D, Formenti SC and Schneider RJ:
Translational control in cancer. Nat Rev Cancer. 10:254–266. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hershey JW: Regulation of protein
synthesis and the role of eIF3 in cancer. Braz J Med Biol Res.
43:920–930. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sesen J, Cammas A, Scotland SJ, Elefterion
B, Lemarié A, Millevoi S, Mathew LK, Seva C, Toulas C, Moyal EC, et
al: Int6/eIF3e is essential for proliferation and survival of human
glioblastoma cells. Int J Mol Sci. 15:2172–2190. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Emmanuel R, Weinstein S, Landesman-Milo D
and Peer D: eIF3c: A potential therapeutic target for cancer.
Cancer Lett. 336:158–166. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Asano K, Kinzy TG, Merrick WC and Hershey
JW: Conservation and diversity of eukaryotic translation initiation
factor eIF3. J Biol Chem. 272:1101–1109. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Moldave K: Eukaryotic protein synthesis.
Annu Rev Biochem. 54:1109–1149. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rothe M, Ko Y, Albers P and Wernert N:
Eukaryotic initiation factor 3 p110 mRNA is overexpressed in
testicular seminomas. Am J Pathol. 157:1597–1604. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Scoles DR, Yong WH, Qin Y, Wawrowsky K and
Pulst SM: Schwannomin inhibits tumorigenesis through direct
interaction with the eukaryotic initiation factor subunit c
(eIF3c). Hum Mol Genet. 15:1059–1070. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Song N, Wang Y, Gu XD, Chen ZY and Shi LB:
Effect of siRNA-mediated knockdown of eIF3c gene on survival of
colon cancer cells. J Zhejiang Univ Sci B. 14:451–459. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Surawicz TS, McCarthy BJ, Kupelian V,
Jukich PJ, Bruner JM and Davis FG: Descriptive epidemiology of
primary brain and CNS tumors: Results from the Central Brain Tumor
Registry of the United States, 1990–1994. Neuro Oncol. 1:14–25.
1999.
|
|
27
|
Zhang L, Pan X and Hershey JW: Individual
overexpression of five subunits of human translation initiation
factor eIF3 promotes malignant transformation of immortal
fibroblast cells. J Biol Chem. 282:5790–5800. 2007. View Article : Google Scholar
|
|
28
|
Sherr CJ: Growth factor-regulated G1
cyclins. Stem Cells. 12(Suppl 1): 47–57. 1994.PubMed/NCBI
|