Introduction
Malignant tumors consist of cancer cells that
usually grow rapidly and spread to other parts of the body via the
bloodstream or lymph system by degrading the surrounding tissues
and subsequently adapting to form secondary foci known as
metastases (1). During the
malignant progression, the acquisition of genetic and/or epigenetic
alterations causes uncontrolled cell proliferation and resistance
to chemotherapy and radiotherapy. In addition, metastatic
conversion is one of the main reasons for high mortality rates and
a leading cause of the poor clinical outcome of cancer patients,
because of its systemic nature and the resistance of disseminated
cells to existing therapeutic agents (2). The process of metastasis involves
several interdependent processes, including a loss of cell adhesion
to the extracellular matrix (ECM), the proteolytic breakdown of
tissue barriers, intravasation into the lumina of blood vessels,
survival in the circulation, extravasation and the subsequent
invasion into new tissues, and metastatic colonization. Therefore,
controlling the metastasis and proliferation of already-established
tumors is an important therapeutic strategy for the management of
malignant tumors (3,4).
Chemotherapy is commonly used to treat cancer.
However, current chemotherapy modalities using cytotoxic
anti-cancer drugs also destroy normal cells and provide little
survival benefit. These modalities also have significant side
effects, dose-limiting toxicities and low bioavailability. In many
studies, natural herbal medicines control various human diseases,
including cancer (5–7). Chinese/Oriental herbal medicines in
the form of herbal cocktails have been used to treat a wide range
of human diseases and improve the physical strength of patients.
Herbal cocktails containing a number of phytochemicals in a single
formula have the merit of functioning synergistically by affecting
multiple biological and pathological processes, leading to maximal
therapeutic effects with minimal side effects (8,9).
Recently, we formulated a novel herbal cocktail
termed MA, which is composed of herbal plants that have been used
traditionally to control allergic disease and inflammation,
including Glycyrrhizae radix, Polygoni cuspidati
radix, Sophorae radix, Cnidii rhizoma and
Arctii fructus (10,11). To improve the therapeutic efficacy
by increasing the absorption and bioavailability of the active
components, we fermented MA using Lactobacillus rhamnosus
(L. rhamnosus) and designated the product of MA128. MA and
MA128 are effective for the treatment of asthma by modulating
ovalbumin (OVA)-specific cytokines and atopic dermatitis (AD) by
reducing Ige production (10,11).
In addition, MA128 had significantly increased efficacy against
asthma and AD compared with MA. Recently, MA128 was demonstrated to
have anti-melanogenic effects by inhibiting tyrosinase activity
(12).
In the present study, we examined the anti-cancer
effects of MA128 in terms of the induction of cell death and
suppression of metastatic potential in highly malignant tumor
cells. We also assessed whether the administration of MA128 was
useful for the management of malignant tumor cells in mice without
exerting adverse effects.
Materials and methods
Cells and animals
Human fibrosarcoma HT1080 (KCLB no. 10121) and
murine melanoma B16F10 (KCLB no. 80008) cells were purchased from
the Korean Cell Line Bank (Seoul, Korea) and maintained in
RPMI-1640 or Dulbecco’s modified eagle’s medium (DMEM; Cellgro,
Manassas, VA, USA) supplemented with 10% (v/v) fetal bovine serum
(FBS; Cellgro), 100 U/ml penicillin and 100 µg/ml
streptomycin (Cellgro) at 37°C in a humidified incubator with 5%
CO2. Human umbilical vein endothelial cells (HUVECs)
were purchased from InnoPharmaScreen (Asan, Korea) at passage 2,
maintained in endothelial cell growth medium-2 (EGM-2; PromoCell,
Heidelberg, germany), and used for experiments at passages 3–8. For
the animal experiments, female athymic nude mice were purchased
from nara Biotech (Seoul, Korea). Female C57BL/6 mice and male
Sprague Dawley rats were obtained from Taconic Farms Inc. (Samtako
Bio Korea, Osan, Korea). All animals were housed under controlled
conditions using a 12/12-h light/dark cycle at 22±1°C and 55±5%
humidity under specific pathogen-free conditions. All animal
experiments were approved by the Animal Care and Use Committee of
the Korea Institute of Oriental Medicine (KIOM, Daejeon, Korea),
with the reference numbers #13–42, #13–48 and #14–27, and were
performed in accordance with the guidelines of the Animal Care and
Use Committee at KIOM.
Preparation of the fermented herbal
cocktail MA128
Herbs used for the preparation of MA128, including
Glycyrrhizae radix, Polygoni cuspidati radix,
Sophorae radix, Cnidii rhizoma, and Arctii
fructus, were purchased from yeongcheon Oriental Herbal Market
(yeongcheon, Korea), and all voucher specimens were deposited in
the herbal bank in KIOM. The authenticity of the plant species was
validated by Professor Ki Hwan Bae (Chungnam national university,
Daejeon, Korea). Pure L. rhamnosus KFRI128 cultures were
obtained from the Korea Food Research Institute (KFRI) and
incubated in MRS medium for 24 h at 37°C as described previously
(12). A total of 1,840 g MA was
soaked in 18.4 l distilled water (DW) and then heat-extracted for 3
h at 115°C using an extractor (Cosmos-600 Extractor; Gyeonseo Co.,
Incheon, Korea). After filtration through standard testing sieves
(150 µm; Retsch, Haan, Germany), the pH of decoction MA was
adjusted to 8.0 using 1 M NaOH. It was then sterilized at 5 min at
121°C using an autoclave. For the fermentation of decoction MA, 10
ml L. rhamnosus (1×108 CFU/ml) was added to 10
liter decoction MA and incubated at 37°C for 48 h. The fermented
MA, designated as MA128, was filtered through a 60-µm nylon
net filter (Millipore, Bedford, MA, USA), freeze-dried and stored
in a desiccator at 4°C. A total of 376 g MA128 powder was produced
and the yield was 20.44%. The freeze-dried MA128 powder was
dissolved in 10% (v/v) DMSO in DW and centrifuged at 14,000 rpm for
10 min. The supernatant was filtered (0.2-µm, pore size) and
kept at 4°C prior to use.
Reagents and antibodies
Type A gelatin from porcine skin, fibronectin from
human plasma, 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyltetrazolium
bromide (MTT), phorbol 12-myristate 13-acetate (PMA) and mitomycin
C were obtained from Sigma Chemical Co. (St. Louis, MO, USA). Type
I collagen solution (0.3%, Cellmatrix type I-A) and the growth
factor-reduced Matrigel basement membrane matrix were purchased
from Nitta Zerachin, Inc. (Osaka, Japan) and BD Biosciences
(Bedford, MA, USA), respectively. Antibodies against p21, p27,
cyclin B, cyclin D, p38, p-p38 (Thr180/Tyr182), ERK1/2, p-ERK1/2
(Thr202/Tyr204), JNK, p-JNK (Thr183/Tyr185), Akt, p-Akt (Ser473),
mTOR, p-mTOR (Ser2481), IκBα, p-IκBα (Ser32/36), p-4e-BP1
(Thr37/46), MMP-9 and tubulin were purchased from Cell Signaling
Technology (Danvers, MA, USA). Anti-HIF-1α antibodies were obtained
from BD Biosciences.
Chemicals
For high-performance liquid chromatography (HPLC),
HPLC-grade acetonitrile and trifluoroacetic acid (TFA) were
purchased from J.T. Baker (Philipsburg, NJ, USA) and Sigma,
respectively. Icariin and glycyrrhizin were obtained from the Tokyo
Chemical Industry Co. (Tokyo, japan). Matrine, chlorogenic acid,
nodakenin and arctiin were purchased from Faces Biochemical Co.
(Wuhan, China). Arctigenin and decursin were obtained from Tocris
Biosciences (Bristol, UK) and KFDA (Osong, Korea),
respectively.
In vivo assays used to assess the effect
of MA128 on tumor growth and pulmonary metastasis
Five-week-old female athymic nude mice were
inoculated subcutaneously with 2×106 HT1080 cells/mouse
in the femoral region. On day 5, the mice were divided randomly
into 3 groups (n=5 per group) and treated daily with saline
(control) or MA128 (75 or 150 mg/kg) in a volume of 100 µl
for 12 days. Tumor volume and weight were measured as previously
described (13). To induce
pulmonary metastasis, C57BL/6 mice were injected intravenously with
B16F10 cells (3×105 cells/mouse) via the tail vein.
After being divided randomly into 3 groups (n=4 per group), the
mice were administered saline (control) or MA128 at 75 or 150
mg/kg. After daily treatment for 17 days, the mice were sacrificed,
their lungs were fixed in Bouin’s solution (Sigma), and
metastasized black colonies were counted macroscopically. To assess
the safety of MA128, 5-week-old athymic nude mice and C57BL/6 mice
(n=3 for each group) were treated daily with vehicle (saline) or
MA128 (75 or 150 mg/kg) for 14 and 15 days, respectively. Body
weight was measured daily, and the weights of the major organs,
including the liver, heart, lung, spleen and kidneys, were measured
at the time of sacrifice. Serological and hematological parameters
were analyzed using XL 200 (erba Diagnostics, Mannheim, germany)
and an ADVIA 2120i hematology system (Siemens Healthcare
Diagnostics, Tarrytown, NY, USA), respectively.
Cell viability assay
To examine the effects of MA128 on cell
proliferation, 5×103 cells/well in a 96-well plate were
treated with various concentrations of MA128 (50–1,000
µg/ml) for 48 h, and MTT assays were then performed as
previously described (13).
Colony formation assay
To assess anchorage-independent cell growth,
5×104 cells were suspended in 2 ml medium containing the
indicated concentrations of MA128, 10% FBS, and 0.3% agar, and then
applied over the bottom agar that had been pre-solidified with 3 ml
medium containing 10% FBS and 0.6% agar. During incubation, colony
formation was observed daily under a phase-contrast microscope, and
images were captured.
Adhesion assays on immobilized FN and
type I collagen
Cell-to-ECM adhesion was measured in 96-well culture
plates as reported previously with slight modifications (14). After coating the plates with 50
µl Fn (5 µg/ml) or 0.3% type I collagen solution
overnight at room temperature, the wells were washed with cold PBS,
blocked with medium containing 0.3% BSA (200 µl/well) and
then washed again. MA128 pre-treated cells were suspended in
serum-free medium at a density of 5×105/ml, plated on
Fn- or type I collagen-coated wells (200 µl/well), and then
allowed to adhere for 1 h at 37°C. Unattached cells were removed by
washing and the attached cells were stained using 0.2% crystal
violet/20% methanol (w/v) solution. After staining, the cells were
dissolved in 1% SDS solution, and the spectrophotometric absorbance
was measured at 560 nm.
Wound-healing cell migration assay
The cells were pre-incubated with mitomycin C (25
µg/ml) for 30 min to inhibit proliferation. Subsequently,
they were injured by scraping across the cell monolayer. After
washing to remove floating cell debris, the cells were allowed to
migrate in the presence of MA128 and monitored under a
phase-contrast microscope.
Transwell migration/invasion assays
Migration/invasion assays were performed using
Transwell chambers with polyethylene tetraphthalate membranes
(6.5-mm diameter and 8-µm pore size) as described previously
with slight modifications (14,15).
For the invasion assays, the membranes were coated with 50
µl Matrigel (diluted in RPMI-1640, 1:8) to form an
intervening invasive barrier.
Gelatin zymography
Cells plated in 12-well culture plates at 80%
confluence were pre-incubated with MA128 in serum-free media for 12
h and then stimulated with 5 nM PMA for an additional 24 h.
Conditioned media (CM) were collected and electrophoresed using 8%
SDS-PAGE containing 0.1% gelatin. After washing, the gels were
incubated for 24 h and then stained and de-stained as previously
described (14,15). MMP-9 was detected as a clear 92-kDa
band against the blue background.
Tube formation assay
Capillary-like tube formation was assayed using
HUVECs, an ECMatrix assay kit (Millipore, Temecula, CA, USA), and
µ-Slide Angiogenesis ibiTreat chambers (ibidi gmbH,
germany), as preivously described (16). HUVECs (5×103 cells) were
suspended in 50 µl MA128-treated HT1080 CM, seeded onto the
surface of polymerized ECMatrix, and incubated for 4–8 h at 37°C in
an incubator containing 5% CO2. The cellular tube
networks were observed using a phase-contrast microscope and
photographed, and the relative tube area was quantified in five
fields of view per well.
Rat aortic ring assay
Dorsal aortas isolated from Sprague Dawley rats were
cut transversely into 1-mm thick rings. Each aortic ring was then
placed in 48-well culture plates pre-coated with 100 µl
Matrigel. After sealing the rings with 40 µl Matrigel, 400
µl EGM-2 was added to the wells and the plates were
incubated for 3 days to allow microvessel sprouting from the aortic
rings. The EGM-2 was then replaced with control or MA128-treated
CM, and microvessel outgrowths were observed and photographed for
an additional day.
Western blotting
Whole cell lysates were extracted using M-PER
Mammalian Protein extraction reagent (Thermo Scientific, Rockford,
IL, USA). Equal amounts of protein were resolved using 8–15%
SDS-PAGE, incubated with specific antibodies, and detected as
previously described (13). The
band intensities were analyzed using ImageJ software.
Determining VEGF, PDGF and bFGF levels
using ELISA
The cells were incubated with the indicated
concentrations of MA128 for 48 h. The levels of VEGF, PDGF, and
bFGF in control and MA128-treated CM were determined using a Human
ELISA Development kit (Peprotech, Rocky Hill, NJ, USA) according to
the manufacturer’s instructions.
Chromatographic analysis of MA128
The phytochemical profile of MA128 was measured
using HPLC analysis on HPLC-DAD (LaChrom elite, Hitachi
High-Technologies Co., Tokyo, Japan), and chromatographic
separation was achieved on a Phenomenex Luna C18 column (4.6×250
mm, 5 µm). A gradient elution of 10% acetonitrile in
deionized water containing 0.05% TFA (A) and 60% acetonitrile in
deionized water containing 0.05% TFA (B) was performed as follows:
0–25 min with 0% B; 25–100 min with 0–45% B; 100–110 min with
15–100% B; 110–120 min with 100% B; 120–122 min with 100–0% B; and
122–140 min with 0% B. The flow rate and injection volume were 1
ml/min and 10 µl, respectively. HPLC chromatograms were
obtained using UV at 190–400 nm. Standard (ST), ST-spiked sample
(SP), and sample solutions were dissolved and diluted in
methanol.
Statistical analysis
Data are presented as mean ± standard deviation
(SDs). The statistical significance of the differences between
groups was analyzed using a two-way analysis of variance (ANOVA)
and the Student’s t-test with SigmaPlot 8.0 software. P<0.05 was
considered to indicate significant differences.
Results
MA128 inhibits the in vivo tumorigenic
growth of HT1080 cells with no adverse effects
Prior to determining the anticancer effects of MA128
in athymic nude mice, we assessed whether the repeated
administration of 75 or 150 mg/kg MA128 elicited systemic toxicity.
The MA128 doses used were according to the amounts administered to
human adults (36.8 g/day/60 kg of body weight) and the yield of the
extraction. Fourteen days of MA128 administration did not cause
death or abnormal behavior. Body and organ weights, the ratios of
GOT/GPT and BUN/CRE, and the hematological parameters were
comparable between MA128-treated and control mice (Table I), suggesting that MA128 caused no
systemic toxicity, including hepatic and renal dysfunction. To
assess the inhibitory effects of MA128 on in vivo tumor
growth, athymic nude mice (n=5 per group) inoculated with highly
malignant HT1080 cells were treated with saline or 75 or 150 mg/kg
MA128 for 12 days beginning 5 days after tumor inoculation. As
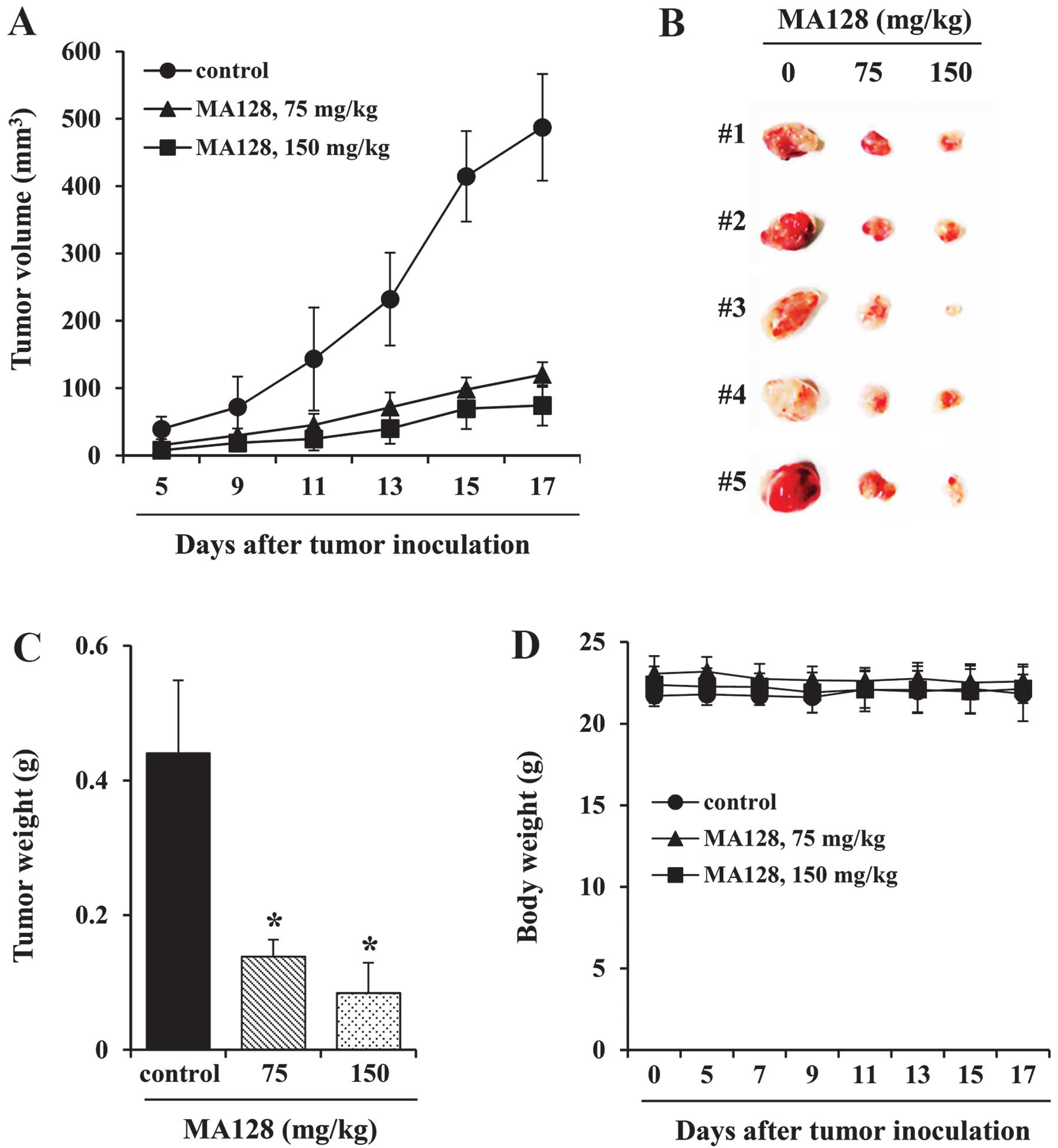
shown in Fig. 1A and B, the
administration of 75 or 150 mg/kg MA128 retarded tumor growth
successfully and suppressed tumor volume by ~75 and 85%,
respectively, compared with the saline-treated control mice on day
17. The control mice exhibited a mean tumor weight of 0.441±0.108
g, whereas the mice treated with 75 and 150 mg/kg MA128 had tumor
weights of 0.139±0.025 g and 0.085±0.045 g, reflecting a decrease
of 68.5 and 80.8%, respectively (Fig.
1C). In addition, there were no significant differences in body
weight between the control and MA128-treated mice during the
experimental period, suggesting that MA128 did not exhibit any
severe toxic effects (Fig. 1D).
 | Table ISafety measurement in athymic nude
mice after oral administration of 75 or 150 mg/kg MA128. |
Table I
Safety measurement in athymic nude
mice after oral administration of 75 or 150 mg/kg MA128.
| Variables | Control | 75 mg/kg | 150 mg/kg |
|---|
| Body weight
(g) |
| Day 0 | 24.66±0.46 | 23.98±1.59 | 24.52±0.48 |
| Day 14 | 25.05±0.48 | 24.54±1.31 | 25.23±0.54 |
| Organ weight
(g) |
| Liver | 1.15±0.04 | 1.11±0.11 | 1.14±0.04 |
| Heart | 0.12±0.01 | 0.11±0.01 | 0.12±0.01 |
| Lung | 0.17±0.01 | 0.18±0.01 | 0.18±0.02 |
| Spleen | 0.11±0.02 | 0.12±0.02 | 0.12±0.01 |
| Kidney (L) | 0.15±0.01 | 0.12±0.01 | 0.14±0.01 |
| Kidney (R) | 0.14±0.01 | 0.13±0.01 | 0.14±0.02 |
| Chemical
analysis |
| GOT (IU/l) | 58.21±5.46 | 54.67±8.33 | 53.68±3.61 |
| GPT (IU/l) | 28.67±8.08 | 31.33±8.08 | 26.04±8.72 |
| Bun (mg/dl) | 22.07±1.36 | 23.47±1.29 | 19.34±1.10 |
| CRE (mg/dl) | 0.47±0.01 | 0.53±0.23 | 0.48±0.04 |
| Hematological
analysis |
| RBC
(×106 cells/µl) | 8.85±0.41 | 8.51±0.49 | 8.83±0.18 |
| Hb (g/dl) | 13.77±0.51 | 13.69±0.62 | 13.95±0.35 |
| PLT
(×105/µl) | 11.67±0.24 | 10.71±2.37 | 11.27±1.11 |
| NEUT (%) | 38.71±2.77 | 31.87±5.21 | 33.05±3.89 |
| LYM (%) | 53.51±4.00 | 60.57±8.11 | 60.55±4.03 |
| MONO (%) | 1.93±0.75 | 1.54±0.21 | 1.43±0.14 |
MA128 decreases cell viability, increases
cell cycle- and cell death-associated protein expression, and
activates p38 and ERK in HT1080 cells
MTT assays were used to assess the effect of MA128
on the growth of cells treated with specified concentrations of
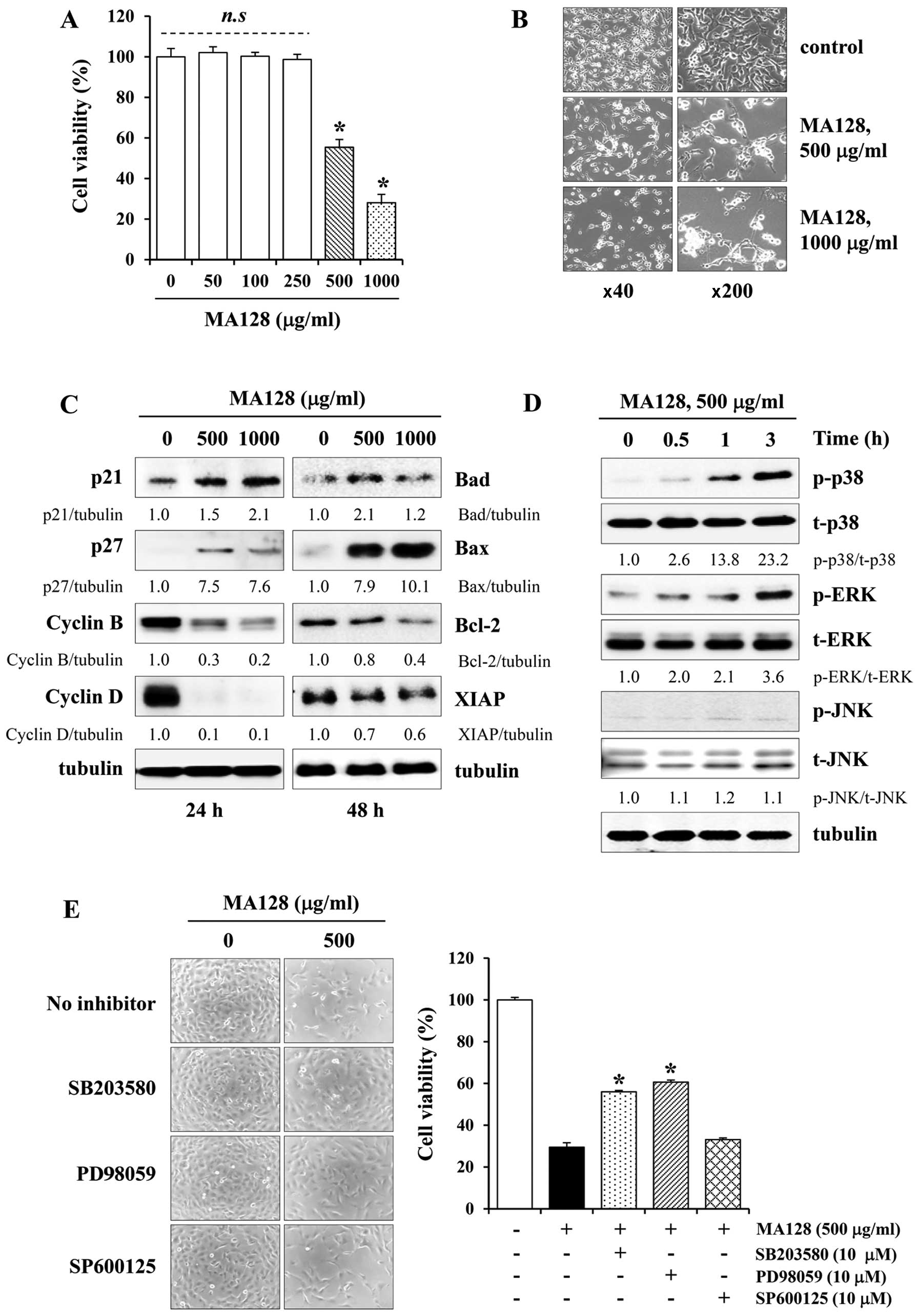
MA128 for 48 h. As shown in Fig.
2A, cell viability was not altered by 50–250 µg/ml MA128
compared with the untreated control cells, whereas treatment with
500 and 1,000 µg/ml MA128 caused a marked decrease in cell
viability and induced most cells to shrink and float (Fig. 2B). Western blotting revealed that
MA128 increased the levels of the cyclin-dependent kinase
inhibitors p21 and p27 in a dose-dependent manner, whereas it
decreased the levels of cyclin B and D compared with the control
cells (Fig. 2C). In addition, MA128
increased the levels of the pro-apoptotic proteins Bad and Bax and
decreased the levels of anti-apoptotic Bcl-2 and XIAP (Fig. 2C). It was reported previously that
the MAPK signaling pathways can induce cell proliferation or death
depending on the stimulus and cell type (17–19).
As shown in Fig. 2D, cytotoxic
doses of MA128 elevated the levels of phosphorylated p38 and ERK
significantly but had little effect on JNK activation. To confirm
the roles of p38 and ERK activation in MA128-mediated cell death,
the cells were pre-incubated with or without pharmacological
inhibitors of p38 (SB203580), ERK (PD98059), or JNK (SP600125) for
1 h and then treated with MA128 for 48 h. As shown in Fig. 2E, SB203580 and PD98059 moderately
protected MA128-treated cells from death by ~50%, whereas SP600125
had little effect. This result suggested that p38 and ERK
activation play roles in MA128-mediated cell death.
MA128 inhibits the in vivo pulmonary
metastasis of B16F10 cells with no adverse effects
Prior to assessing the anti-metastatic effects of
MA128 in C57BL/6 mice, the safety of MA128 treatment was assessed
over a 15-day period. As shown in Table II, MA128 did not induce any adverse
effects, such as loss of body weight or differences in organ
weights. In addition, there were no critical differences in the
serological or hematological parameters between MA128-treated and
control mice. After confirming safety, we evaluated whether MA128
reduced the pulmonary colonization of B16F10 cells injected
intravenously into the tail vein of C57BL/6 mice. As shown in
Fig. 3A and B, B16F10 cells
metastasized to the lungs of control mice and formed a considerable
number of black colonies (939.81±83.98). However, the incidence of
metastatic colonies in mice treated with 75 and 150 mg/kg MA128 was
decreased significantly by ~57.9 (395.33±75.72) and 61.1%
(365.67±105.19), respectively, compared with the control mice.
These results suggested that MA128 intake suppressed the pulmonary
metastasis of B16F10 cells efficiently without causing systemic
toxicity. To examine the anti-metastatic effects of MA128 in
vitro, we monitored the cell-to-ECM adhesion, migration, and
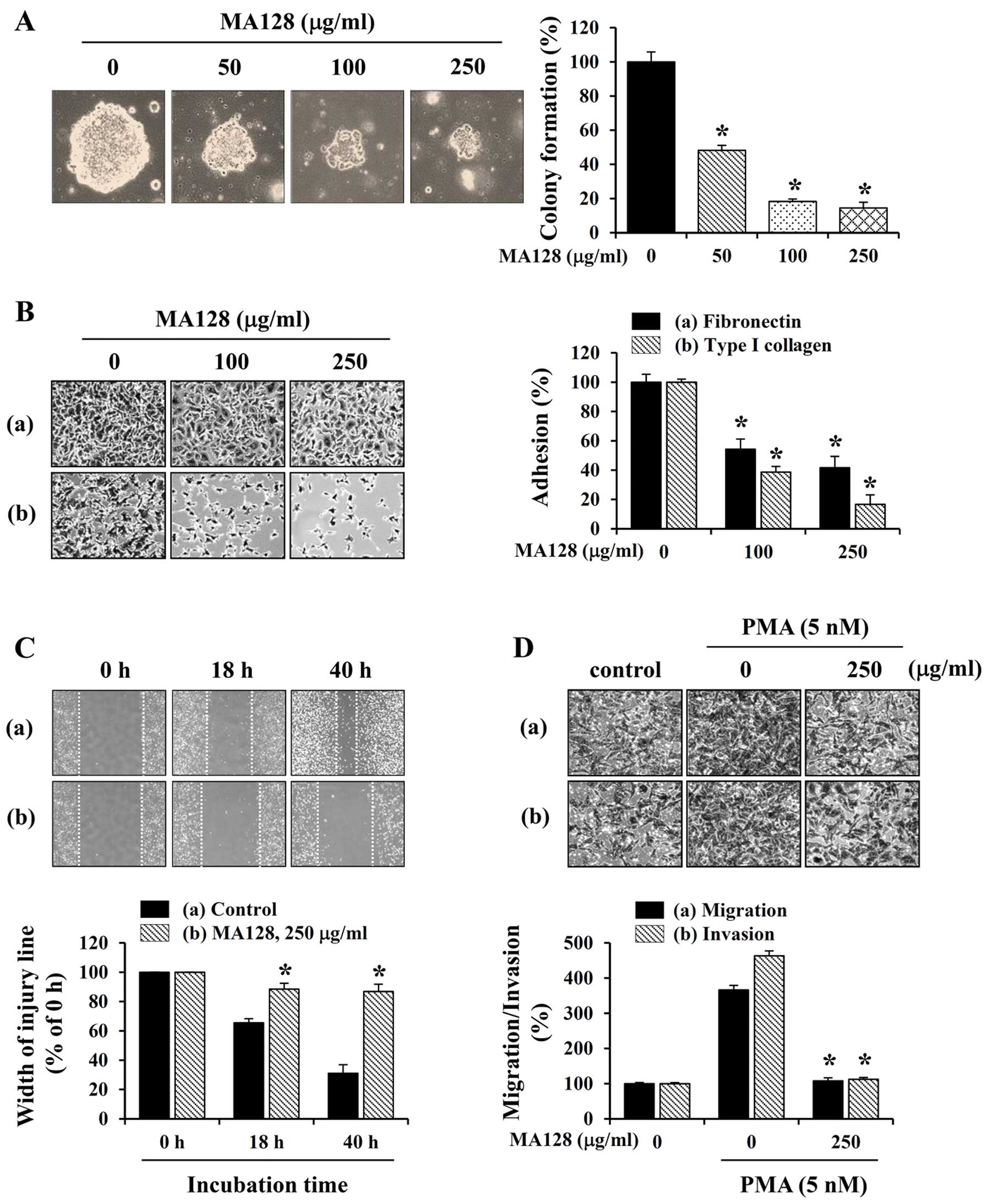
invasion of B16F10 cells in the presence or absence of MA128.
Pre-treatment with MA128 inhibited the attachment of cells to FN
and collagen in a dose-dependent manner (Fig. 3C). In the Transwell culture system,
serum-induced migration and invasion were markedly reduced by MA128
to ~10.6–25.8 and 15.7–41.2% in the control cells, respectively
(Fig. 3D).
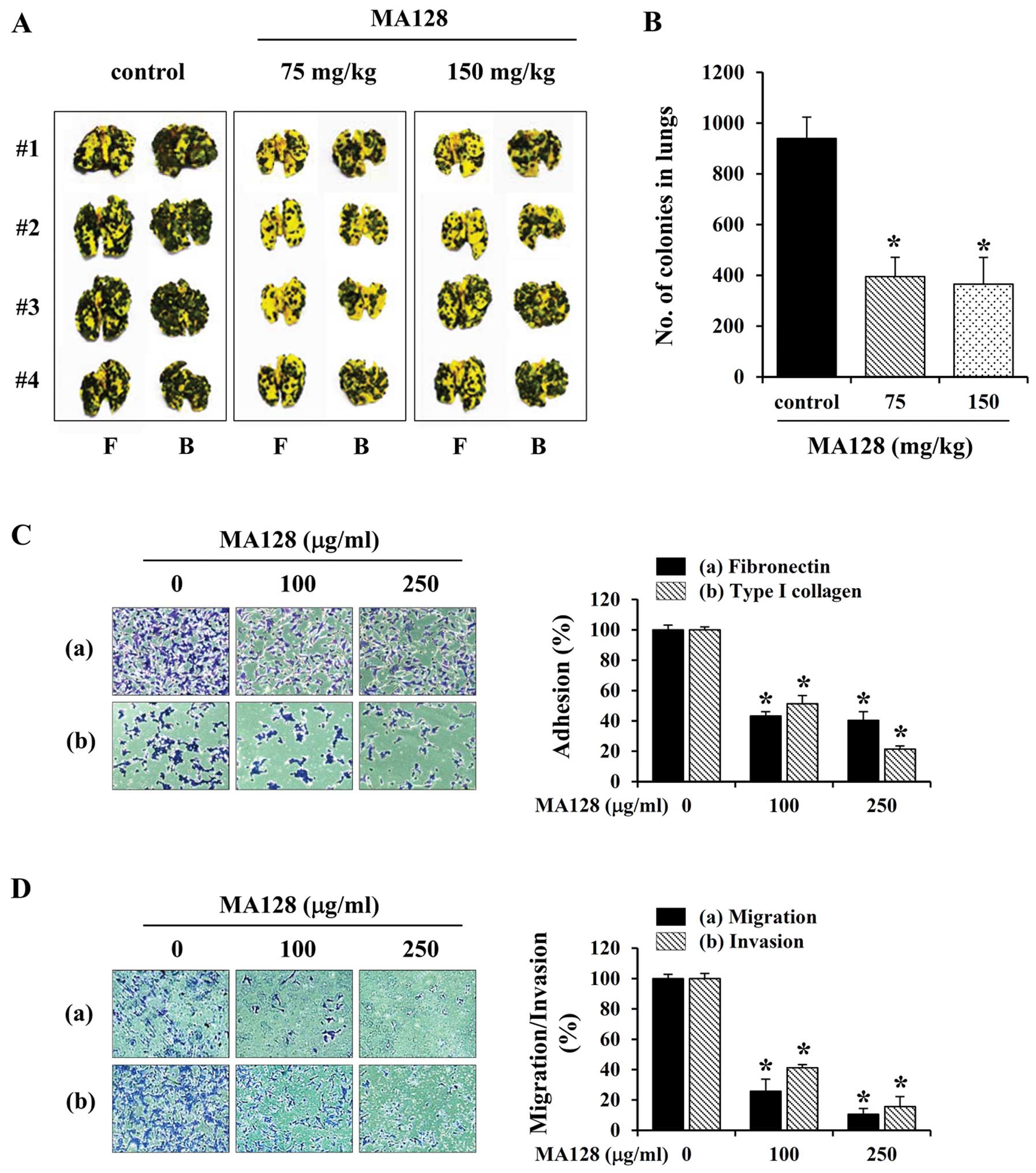 | Figure 3MA128 markedly inhibits the in
vivo pulmonary metastasis and in vitro adhesion,
migration, and invasion of B16F10 cells at non-toxic
concentrations. (A) Cells (3×105/mouse in 200 µl
PBS) were injected into the tail vein of C57BL/6 mice, which were
then treated daily with 75 and 150 mg/kg MA128 (n=4 per group). On
day 17, the mice were sacrificed, and the F, front and B, back of
the lungs were photographed. (B) Black colonies on the lung surface
were counted macroscopically, and data are presented as mean ± SD.
*P<0.05 vs. control (saline). (C) B16F10 cells
pre-incubated with or without MA128 were seeded in Fn- or
collagen-coated wells, incubated for 1 h, and then washed to remove
unattached cells. (D) B16F10 cells pre-incubated with or without
MA128 were added into the upper well of Transwell chambers and
incubated for 24 h (migration) or 48 h (invasion). (C and D) Cells
were stained with 0.2% crystal violet/20% methanol (w/v) solution,
and the relative degrees of adhesion, migration, and invasion were
calculated. Data are presented as mean ± SD. *P<0.05
vs. untreated control. |
| Table IISafety measurement in C57BL/6J mice
after oral administration of 75 or 150 mg/kg MA128. |
Table II
Safety measurement in C57BL/6J mice
after oral administration of 75 or 150 mg/kg MA128.
| Variables | Control | 75 mg/kg | 150 mg/kg |
|---|
| Body weight
(g) |
| Day 0 | 17.43±0.36 | 17.26±0.24 | 17.85±0.35 |
| Day 15 | 19.20±0.41 | 19.93±0.72 | 20.58±0.87 |
| Organ weight
(g) |
| Liver | 1.08±0.04 | 1.13±0.03 | 1.05±0.07 |
| Heart | 0.10±0.02 | 0.11±0.01 | 0.11±0.01 |
| Lung | 0.15±0.01 | 0.16±0.02 | 0.18±0.01 |
| Spleen | 0.08±0.01 | 0.09±0.01 | 0.09±0.01 |
| Kidney (L) | 0.12±0.01 | 0.13±0.01 | 0.12±0.01 |
| Kidney (R) | 0.12±0.01 | 0.13±0.01 | 0.12±0.01 |
| Chemical
analysis |
| GOT (IU/l) | 52.74±3.11 | 53.37±2.32 | 54.08±2.04 |
| GPT (IU/l) | 29.31±4.62 | 27.38±2.47 | 27.14±1.42 |
| BUN (mg/dl) | 36.52±1.54 | 34.78±1.84 | 33.49±2.66 |
| CRE (mg/dl) | 0.71±0.16 | 0.59±0.15 | 0.57±0.21 |
| Hematological
analysis |
| RBC
(×106 cells/µl) | 9.52±0.30 | 9.47±0.56 | 9.51±0.21 |
| Hb (g/dl) | 13.62±0.30 | 13.58±0.08 | 13.57±0.08 |
| PLT
(×105/µl) | 9.78±1.96 | 9.52±0.18 | 10.43±1.37 |
| NEUT (%) | 9.78±1.96 | 9.52±0.18 | 10.43±1.37 |
| LYM (%) | 86.55±1.85 | 84.86±0.29 | 84.38±2.22 |
| MONO (%) | 0.61±0.11 | 0.58±0.08 | 0.71±0.18 |
MA128 suppresses PMA-induced migration
and invasion by suppressing MMP-9 activity and blocking NF-κB
activation in HT1080 cells
To clarify the anti-metastatic mechanism of action
of MA128, HT1080 cells and PMA were used under the conditions as
previously described (20–22). At the non-toxic concentrations of
50–250 µg/ml, MA128 treatment prevented HT1080
anchorage-independent colony formation (Fig. 4A) and the adhesion to FN and
collagen (Fig. 4B) in a
dose-dependent manner. Results of the wound-healing assay showed
that the control HT1080 cells repaired the wounded area efficiently
to ~35 and 70% levels after 18 and 40 h, respectively. By contrast,
MA128 treatment inhibited wound repair significantly, allowing only
~10 and 15% healing at 18 and 40 h, respectively (Fig. 4C). In addition, the ability of
HT1080 cells to undergo Transwell migration and invasion was
markedly increased by PMA stimulation, whereas MA128 almost
completely inhibited migration and invasion under PMA-stimulated
conditions (Fig. 4D), suggesting
that MA128 exerted potent anti-metastatic effects. To elucidate the
mechanism by which MA128 inhibited the metastatic potential, we
examined whether MA128 suppressed the expression and activity of
MMP-9, which is essential for facilitating tumor metastasis by
degrading the surrounding ECM.
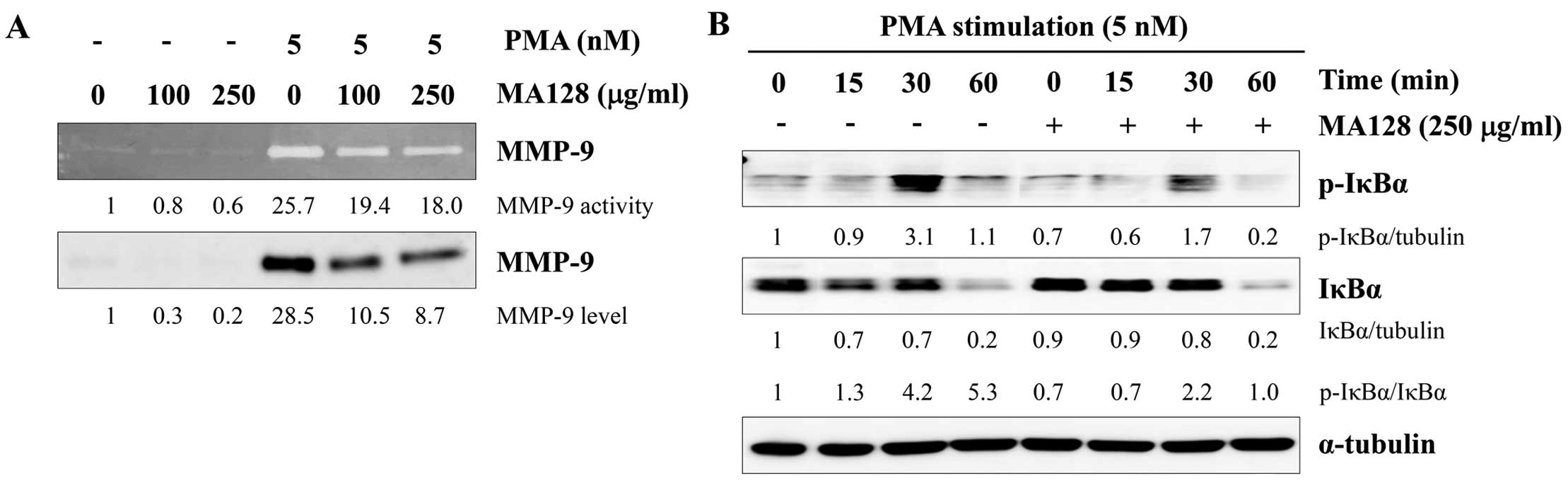
As shown in Fig. 5A,
PMA strongly induced the gelatinolytic activity and secretion of
MMP-9, as determined using gelatin zymography and western blotting.
By contrast, treatment with MA128 markedly inhibited the
PMA-induced increase in MMP-9 activity and expression in a
dose-dependent manner. Since the transcription factor nF-κB is
critical for MMP expression and invasion in various cancer cells
(23,24), we examined the effects of MA128 on
PMA-induced NF-κB activation. As shown in Fig. 5B, PMA treatment markedly increased
the phosphorylation and degradation of IκBα in HT1080 control
cells. However, MA128 significantly inhibited PMA-induced IκBα
phosphorylation and degradation. Consequently, the p-IκBα/IκBα
ratio was much lower in the MA128-treated cells compared with that
in the control cells. Taken together, these results suggested that
non-toxic doses of MA128 suppress the metastatic potential of
HT1080 cells by reducing MMP-9 activity and inhibiting nF-κB
activation.
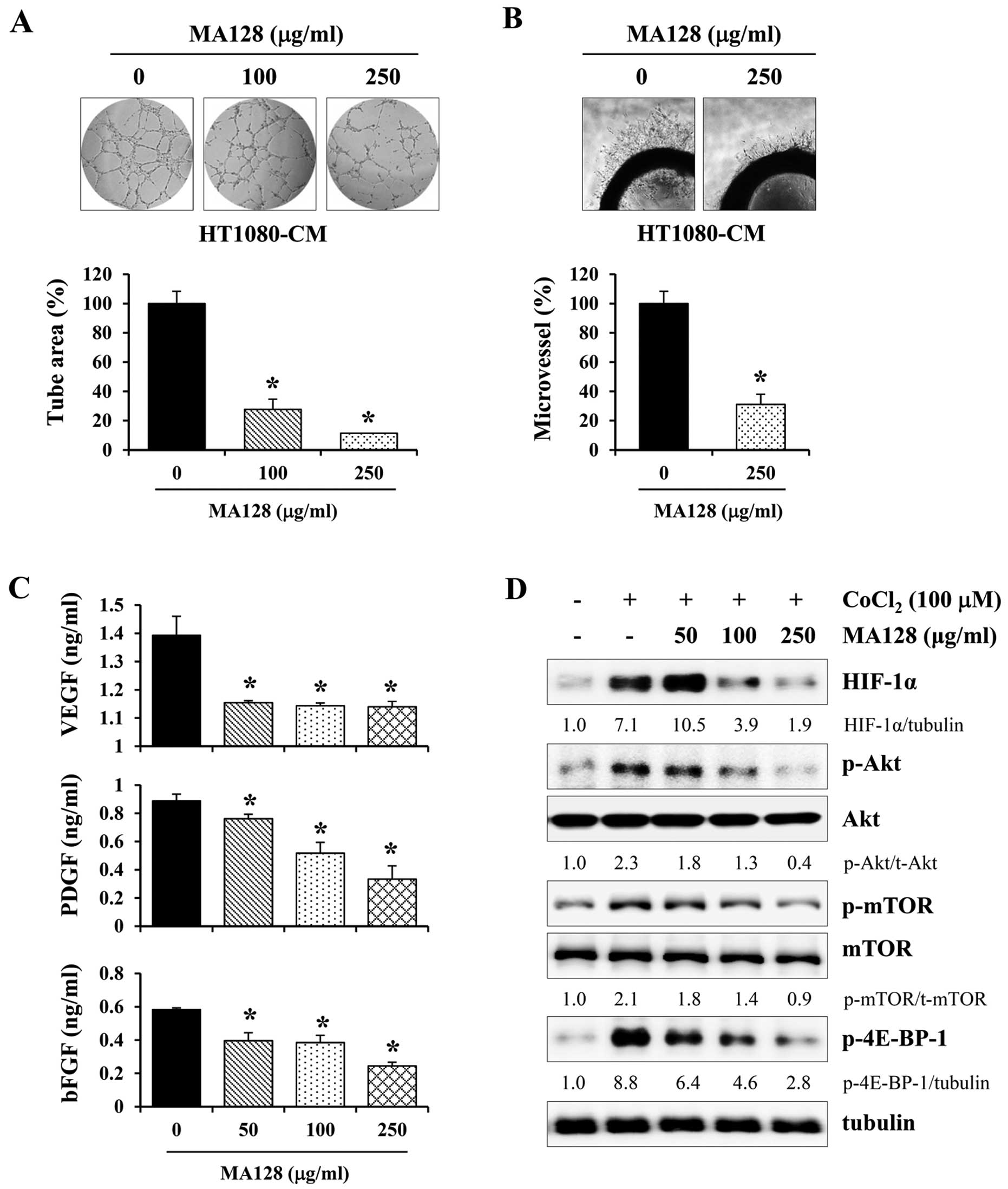
MA128 attenuates tumor-induced
angiogenesis by reducing the production of angiogenic factors
Tumor-induced angiogenesis, the formation of new
blood vessels from existing vasculature, is induced by tumor cells
and is critical for sustained tumor growth and metastasis (25–27).
It has been reported that supernatants harvested from various tumor
cells triggered an angiogenic response. Therefore, we examined the
ability of HUVECs to form network structures on Matrigel after
incubation with control or MA128-treated CM derived from HT1080
cell cultures. As shown in Fig. 6A,
control CM stimulated HUVECs to assemble into tube-like structures
on Matrigel, whereas MA128-treated CM significantly reduced tube
formation in terms of the number, length, and area of
capillary-like structures in a dose-dependent manner. Using ex
vivo angiogenesis assay, we assessed microvessel sprout
formation in rat aortic rings, after incubation with control or
MA128-treated CM from HT1080 cells. As reported previously, control
CM from HT1080 cell cultures triggered microvessel sprouting around
the aortic rings (16), whereas the
length of the sprouting microvessels around the aortic rings
incubated with MA128-treated CM was markedly shorter than that with
control CM (Fig. 6B). As
tumor-induced angiogenesis is affected by tumor cell-produced
angiogenic factors, the levels of pro-angiogenic factors, including
VEGF-α, PDGF and bFGF, were measured in the control and
MA128-treated CM from HT1080 cells. As shown in Fig. 6C, ELISA revealed that MA128
treatment significantly reduced the production of angiogenic
factors in a dose-dependent manner (Fig. 6C). Angiogenic factors are immediate
downstream targets of HIF-1α (28).
Therefore, we assessed the effect of MA128 on HIF-1α production.
MA128 reduced CoCl2-stimulated HIF-1α protein
accumulation, which was accompanied via inhibition of
Akt/mTOR/4E-BP-1 signaling (Fig.
6D). This result suggested that MA128 decreased tumor-induced
angiogenesis by inhibiting the production of angiogenic factors via
the suppression of HIF-1α accumulation.
Identifying the main components in MA128
using HPLC
HPLC identified eight phytochemicals in MA128:
matrine, chlorogenic acid, nodakenin, arctiin, icariin, arctigenin,
glycyrrhizin and decursin. To achieve the desired separation, a
gradient elution of water and acetonitrile was applied. TFA was
added to advance the peak shape and inhibit peak tailing, and the
UV wavelengths of the eight components were adjusted based on the
maximum UV spectra absorption of each component. As shown in
Fig. 7, matrine, nodakenin,
arctiin, icariin, arctigenin, and decursin were detected at 203 nm,
and chlorogenic acid and glycyrrhizin were detected at 254 and 320
nm, respectively. Each component in MA128 was identified by
comparing the retention time (tR) and uV spectra
with known data. Profiles corresponding to matrine (1,
tR 5.3 min), chlorogenic acid (2,
tR 11.3 min), nodakenin (3, tR
52.7 min), arctiin (4, tR 76.6 min), icariin (5,
tR 89.1 min), arctigenin (6, tR
105.2 min), glycyrrhizin (7, tR 109.6 min), and
decursin (8, tR 119.7 min) were identified in
MA128.
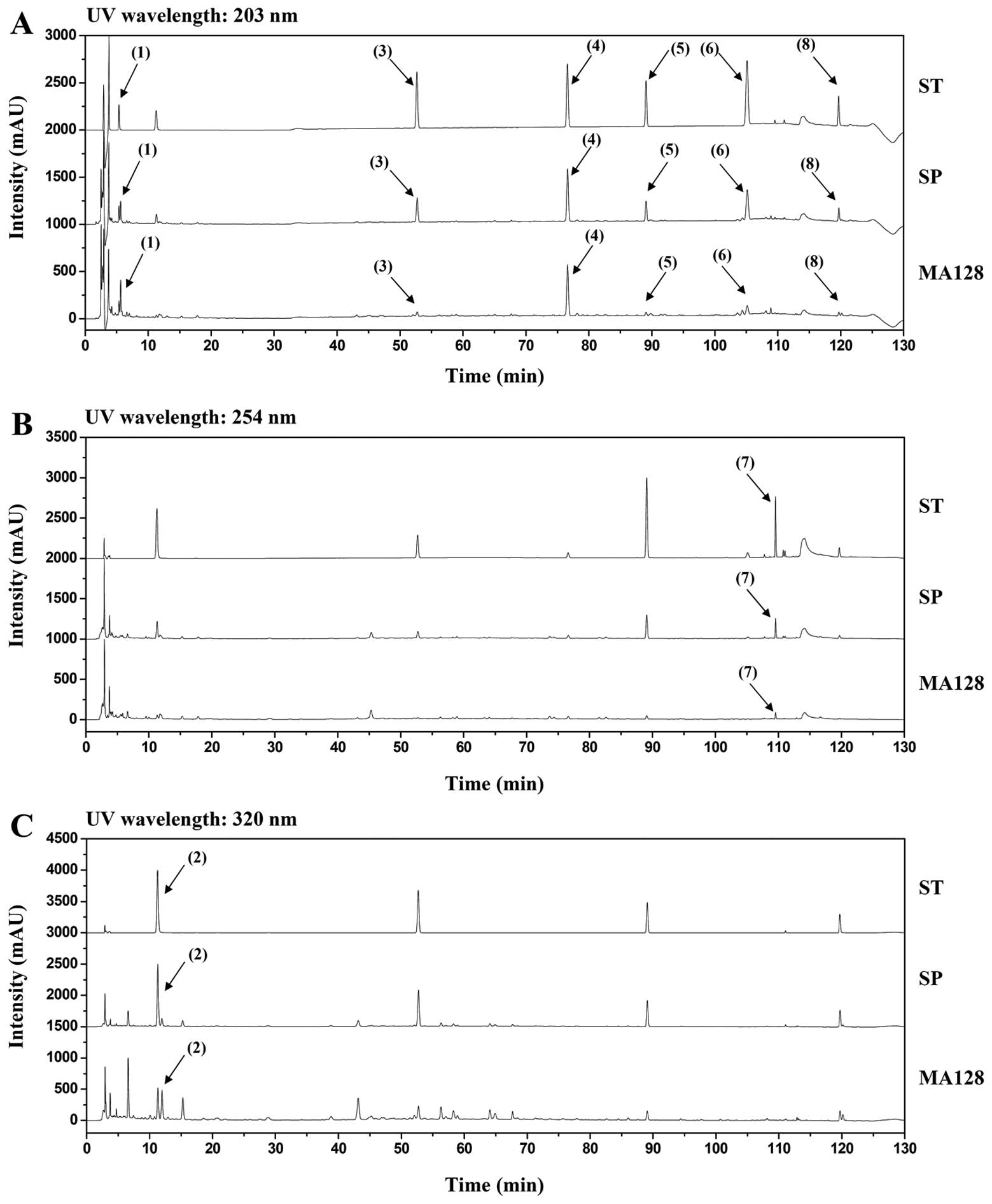 | Figure 7Chromatograms of eight major standard
compounds (ST), spiked samples (SP), and MA128 identified at UV
wavelengths of (A) 203 nm, (B) 254 nm, and (C) 320 nm. Matrine
(1), chlorogenic acid (2), nodakenin (3), arctiin (4), icariin (5), arctigenin (6), glycyrrhizin (7) and decursin (8) were identified. |
Discussion
It has been reported that interfering with critical
steps of cancer growth and progression, including metastasis and
angiogenesis, using naturally occurring herbal agents and
phytochemicals is important for reducing the morbidity and
mortality of cancer patients (7,29,30).
Properly formulated herbal cocktails have the advantage of
synergism among individual herbs and improved therapeutic efficacy
by targeting various cell pathways while minimizing side effects.
Although Chinese/Oriental herbal medicine has long been used to
control malignancies, quality control of the active ingredients,
safety assessment and elucidation of the mechanism of action must
be accompanied by medicinal efficacy for their therapeutic value in
modern medicine (31). Rigorous and
systematic cell-based in vitro and in vivo
pre-clinical animal studies that support their efficacy, safety,
and underlying mechanisms are essential for transforming
traditional herbal practice into ‘evidence-based medicine’.
We developed a novel mixture of medicinal herbs
known as MA128, comprising several herbs that have been used
traditionally to treat inflammatory and allergic diseases and have
been fermented with L. rhamnosus to improve their
therapeutic potential. In previous studies, we demonstrated that
the oral administration of MA and MA128 ameliorated the major
clinical signs of AD including erythema/darkening, edema,
excoriations and dryness in AD-induced BALB/c mice. It also
exhibited anti-asthmatic effects by suppressing the infiltration of
eosinophils into the airway and blood and reducing allergic airway
inflammation in asthma-induced BALB/c mice. In addition, the
therapeutic efficacy of MA128 in these two experimental models was
increased significantly by fermentation (11,21).
In the present study, we have demonstrated that
MA128 inhibits the proliferation, metastasis, and tumor-induced
angiogenesis of malignant tumor cells in vitro and in
vivo, with no apparent side effects. This finding suggests that
MA128 may be used as a safe and effective remedy for controlling
malignant tumor cells. In previous studies, individual herbs in
MA128, including Sophorae radix, Polugoni cuspidati
radix, Arctii fructus, and Angelica gigantis
radix, were shown to elicit anti-cancer effects by inhibiting
cell proliferation and inducing apoptosis (32–35).
We used HPLC to identify eight phytochemicals in MA128. Of these,
previous studies have shown that decursin inhibits the lung
metastasis of CT-26 murine colon carcinoma by targeting MMP-9
activity via the suppression of ERK and JNK phosphorylation,
inducing cell cycle arrest and apoptosis in human carcinomas,
including prostate, colon, and bladder, and by inhibiting
VEGF-induced angiogenesis, suggesting that it plays a crucial role
in mediating anti-cancer activity (36–38).
In addition, matrine and glycyrrhizin inhibited cell proliferation
and induced apoptosis in many human carcinomas (39–41),
whereas matrine reduced the invasion of A375 human malignant
melanoma cells (42). Icariin
exerted negative effects on the invasion and migration of BGC-823
human gastric cancer cells by reducing Rac1 and VASP expression
(43). It also induced apoptosis in
SMMC-7721 human hepatoma cells by generating ROS via the activation
of JNK (44). These results suggest
that MA128 has the potential to suppress cell proliferation,
migration, invasion, and angiogenesis via these active
components.
Tumor-induced angiogenesis is dependent on the
release of pro-angiogenic factors, such as VEGF, PDGF, bFGF,
angiopoietin and MMPs by tumor cells. These factors in turn
stimulate existing blood vessels to initiate angiogenesis. Without
angiogenesis, tumor masses are restricted to a few millimeters due
to a lack of oxygen and nutrients, and they cannot escape into the
circulation to metastasize (25).
It has been reported that MMP-2, MMP-9, and VEGF expression levels
were positively correlated with tumor size, lymphatic and venous
invasion, and lymph node metastasis, as well as malignant
progression (45). In the present
study, treatment with non-toxic concentrations of MA128 decreased
the production of MMP-9 and pro-angiogenic factors in CM
significantly by inhibiting NF-κB activation and HIF-1α
accumulation. Therefore, the proteolytic activity against gelatin,
migration/invasion through Matrigel, and tumor-induced angiogenesis
were decreased in MA128-treated cells compared with the control
cells. In addition, higher concentrations of MA128 induced cell
death efficiently by inducing p38 and ERK phosphorylation. In
animal models, the oral administration of 75 and 150 mg/kg MA128
markedly suppressed the growth of HT1080 cells and pulmonary
metastasis of B16F10 cells. Safety evaluation of MA128 in athymic
nude mice and C57BL/6 mice revealed that the oral administration of
75 and 150 mg/kg MA128 during the experimental period did not cause
any systemic toxicity with respect to body weight loss, organ
abnormalities, or changes in serological/hematological parameters.
This finding suggests that MA128 is a safe and effective cancer
treatment. Of note, none of the individual herbs exhibited
anti-cancer activities in HT1080 cells at the concentrations
present in MA128, suggesting that the herbs present in MA128
exerted synergistic and reciprocal effects.
In summary, the current data clearly demonstrate
that higher concentrations of MA128 induced cell death, primarily
via p38 and ERK activation, and non-toxic concentrations of MA128
suppressed metastatic activity by inhibiting the production of
MMP-9 and pro-angiogenic factors in malignant tumor cells.
Furthermore, MA128 exhibited potent anti-cancer efficacy against
tumor growth and pulmonary metastasis in mice with no adverse
effects. These results collectively suggest that MA128 is a safe
and efficacious herbal remedy against malignant tumors.
Acknowledgments
This study has been supported by the grant K14050
awarded to the Korea Institute of Oriental Medicine (KIOM) from the
Ministry of Science, ICT and Future Planning (MSIP), Republic of
Korea.
References
|
1
|
Liotta LA, Steeg PS and Stetler-Stevenson
WG: Cancer metastasis and angiogenesis: An imbalance of positive
and negative regulation. Cell. 64:327–336. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Maddika S, Ande SR, Panigrahi S,
Paranjothy T, Weglarczyk K, Zuse A, Eshraghi M, Manda KD, Wiechec E
and Los M: Cell survival, cell death and cell cycle pathways are
interconnected: Implications for cancer therapy. Drug Resist Updat.
10:13–29. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Brooks SA, Lomax-Browne HJ, Carter TM,
Kinch CE and Hall DM: Molecular interactions in cancer cell
metastasis. Acta Histochem. 112:3–25. 2010. View Article : Google Scholar
|
|
4
|
Valastyan S and Weinberg RA: Tumor
metastasis: Molecular insights and evolving paradigms. Cell.
147:275–292. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Petrylak DP: The current role of
chemotherapy in metastatic hormone-refractory prostate cancer.
Urology. 65(Suppl): 3–7; discussion 7–8. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Evans T: Chemotherapy in advanced
non-small cell lung cancer. Semin Respir Crit Care Med. 26:304–313.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mondal S, Bandyopadhyay S, Ghosh MK,
Mukhopadhyay S, Roy S and Mandal C: Natural products: Promising
resources for cancer drug discovery. Anticancer Agents Med Chem.
12:49–75. 2012. View Article : Google Scholar
|
|
8
|
Corson TW and Crews CM: Molecular
understanding and modern application of traditional medicines:
Triumphs and trials. Cell. 130:769–774. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kiyohara H, Matsumoto T and Yamada H:
Combination effects of herbs in a multi-herbal formula: expression
of Juzen-taiho-to’s immuno-modulatory activity on the intestinal
immune system. Evid Based Complement Alternat Med. 1:83–91. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kim DS, Kim SH, Kim BK, Yang MC and Ma JY:
Antiasthmatic effects of herbal complex MA and its fermented
product MA128. Evid Based Complement Alternat Med. 2012:7695082012.
View Article : Google Scholar
|
|
11
|
Chung TH, Kang TJ, Cho WK, Im GY, Lee GS,
Yang MC, Cho CW and Ma JY: Effectiveness of the novel herbal
medicine, KIOM-MA, and its bioconversionproduct, KIOM-MA128, on the
treatment of atopic dermatitis. Evid Based Complement Alternat Med.
2012:7629182012. View Article : Google Scholar
|
|
12
|
Kim A and Ma JY: Anti-melanogenic activity
of the novel herbal medicine, MA128, through inhibition of
tyrosinase activity mediated by the p38 mitogen-activated protein
kinases and protein kinase signaling pathway in B16F10 cells.
Pharmacogn Mag. 10(Suppl 3): S463–S471. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim A, Im M, Yim NH, Kim T and Ma JY: A
novel herbal medicine, KIOM-C, induces autophagic and apoptotic
cell death mediated by activation of JNK and reactive oxygen
species in HT1080 human fibrosarcoma cells. PLoS One. 9:e987032014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim A, Im M, Yim NH, Jung YP and Ma JY:
Aqueous extract of Bambusae Caulis in Taeniam inhibits PMA-induced
tumor cell invasion and pulmonary metastasis: Suppression of NF-κB
activation through ROS signaling. PLoS One. 8:e780612013.
View Article : Google Scholar
|
|
15
|
Kim A, Yim NH, Im M, Jung YP, Kim T and Ma
JY: Suppression of the invasive potential of highly malignant tumor
cells by KIOM-C, a novel herbal medicine, via inhibition of NF-κB
activation and MMP-9 expression. Oncol Rep. 31:287–297. 2014.
|
|
16
|
Kim A, Im M, Yim NH and Ma JY: Reduction
of metastatic and angiogenic potency of malignant cancer by
Eupatorium fortunei via suppression of MMP-9 activity and VEGF
production. Sci Rep. 4:69942014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jin Z and El-Deiry WS: Overview of cell
death signaling pathways. Cancer Biol Ther. 4:139–163. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ouyang L, Shi Z, Zhao S, Wang FT, Zhou TT,
Liu B and Bao JK: Programmed cell death pathways in cancer: A
review of apoptosis, autophagy and programmed necrosis. Cell
Prolif. 45:487–498. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Reddy KB, Nabha SM and Atanaskova N: Role
of MAP kinase in tumor progression and invasion. Cancer Metastasis
Rev. 22:395–403. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Roomi MW, Monterrey JC, Kalinovsky T, Rath
M and Niedzwiecki A: Patterns of MMP-2 and MMP-9 expression in
human cancer cell lines. Oncol Rep. 21:1323–1333. 2009.PubMed/NCBI
|
|
21
|
Choi JH, Han EH, Hwang YP, Choi JM, Choi
CY, Chung YC, Seo JK and Jeong HG: Suppression of PMA-induced tumor
cell invasion and metastasis by aqueous extract isolated from
Prunella vulgaris via the inhibition of NF-κB-dependent MMP-9
expression. Food Chem Toxicol. 48:564–571. 2010. View Article : Google Scholar
|
|
22
|
Hwang YP, Yun HJ, Kim HG, Han EH, Lee GW
and Jeong HG: Suppression of PMA-induced tumor cell invasion by
dihydroartemisinin via inhibition of PKCα/Raf/MAPKs and
NF-κB/AP-1-dependent mechanisms. Biochem Pharmacol. 79:1714–1726.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kim A, Kim MJ, Yang Y, Kim JW, Yeom YI and
Lim JS: Suppression of NF-κB activity by NDRG2 expression
attenuates the invasive potential of highly malignant tumor cells.
Carcinogenesis. 30:927–936. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yeh CB, Hsieh MJ, Hsieh YH, Chien MH,
Chiou HL and Yang SF: Antimetastatic effects of norcantharidin on
hepatocellular carcinoma by transcriptional inhibition of MMP-9
through modulation of NF-kB activity. PLoS One. 7:e310552012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Risau W: Mechanisms of angiogenesis.
Nature. 386:671–674. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jang YJ and Kim DS, Jeon OH and Kim DS:
Saxatilin suppresses tumor-induced angiogenesis by regulating VEGF
expression in NCI-H460 human lung cancer cells. J Biochem Mol Biol.
40:439–443. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Grant DS, Yenisey C, Rose RW, Tootell M,
Santra M and Iozzo RV: Decorin suppresses tumor cell-mediated
angiogenesis. Oncogene. 21:4765–4777. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hirota K and Semenza GL: Regulation of
angiogenesis by hypoxia-inducible factor 1. Crit Rev Oncol Hematol.
59:15–26. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Woo SM, Choi YK, Cho SG, Park S and Ko SG:
A new herbal formula, KSG-002, suppresses breast cancer growth and
metastasis by targeting NF-κB-dependent TNF α production in
macrophages. Evid Based Complement Alternat Med. 2013:7282582013.
View Article : Google Scholar
|
|
30
|
Lee HJ, Lee EO, Rhee YH, Ahn KS, Li GX,
Jiang C, Lü J and Kim SH: An oriental herbal cocktail,
ka-mi-kae-kyuk-tang, exerts anti-cancer activities by targeting
angiogenesis, apoptosis and metastasis. Carcinogenesis.
27:2455–2463. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Buchanan DR, White JD, O’Mara AM, Kelaghan
JW, Smith WB and Minasian LM: Research-design issues in
cancer-symptom-management trials using complementary and
alternative medicine: Lessons from the national Cancer Institute
Community Clinical Oncology Program experience. J Clin Oncol.
23:6682–6689. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kim SC, Byun SH, Yang CH, Kim CY, Kim JW
and Kim SG: Cytoprotective effects of Glycyrrhizae radix extract
and its active component liquiritigenin against cadmium-induced
toxicity (effects on bad translocation and cytochrome c-mediated
PARP cleavage). Toxicology. 197:239–251. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xiao ZM, Wang AM, Wang XY and Shen SR:
Effects of ethanol extract of Radix Sophorae Flavescentis on
activity of colon cancer HT29 cells. Afr J Tradit Complement Altern
Med. 10:352–355. 2013.PubMed/NCBI
|
|
34
|
Yang S, Ma J, Xiao J, Lv X, Li X, Yang H,
Liu Y, Feng S and Zhang Y: Arctigenin anti-tumor activity in
bladder cancer T24 cell line through induction of cell-cycle arrest
and apoptosis. Anat Rec (Hoboken). 295:1260–1266. 2012. View Article : Google Scholar
|
|
35
|
Shin JA, Shim JH, Jeon JG, Choi KH, Choi
ES, Cho NP and Cho SD: Apoptotic effect of Polygonum cuspidatum in
oral cancer cells through the regulation of specificity protein 1.
Oral Dis. 17:162–170. 2011. View Article : Google Scholar
|
|
36
|
Jung MH, Lee SH, Ahn EM and Lee YM:
Decursin and decursinol angelate inhibit VegF-induced angiogenesis
via suppression of the VEGFR-2-signaling pathway. Carcinogenesis.
30:655–661. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kim WJ, Lee SJ, Choi YD and Moon SK:
Decursin inhibits growth of human bladder and colon cancer cells
via apoptosis, G1-phase cell cycle arrest and extracellular
signal-regulated kinase activation. Int J Mol Med. 25:635–641.
2010.PubMed/NCBI
|
|
38
|
Son SH, Park KK, Park SK, Kim YC, Kim YS,
Lee SK and Chung WY: Decursin and decursinol from Angelica gigas
inhibit the lung metastasis of murine colon carcinoma. Phytother
Res. 25:959–964. 2011. View Article : Google Scholar
|
|
39
|
Zhang Y, Zhang H, Yu P, Liu Q, Liu K, Duan
H, Luan G, Yagasaki K and Zhang G: Effects of matrine against the
growth of human lung cancer and hepatoma cells as well as lung
cancer cell migration. Cytotechnology. 59:191–200. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ma L, Wen S, Zhan Y, He Y, Liu X and Jiang
J: Anticancer effects of the Chinese medicine matrine on murine
hepatocellular carcinoma cells. Planta Med. 74:245–251. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Thirugnanam S, Xu L, Ramaswamy K and
Gnanasekar M: Glycyrrhizin induces apoptosis in prostate cancer
cell lines DU-145 and LNCaP. Oncol Rep. 20:1387–1392.
2008.PubMed/NCBI
|
|
42
|
Liu XY, Fang H, Yang ZG, Wang XY, Ruan LM,
Fang DR, Ding YG, Wang YN, Zhang Y, Jiang XL, et al: Matrine
inhibits invasiveness and metastasis of human malignant melanoma
cell line A375 in vitro. Int J Dermatol. 47:448–456. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang Y, Dong H, Zhu M, Ou Y, Zhang J, Luo
H, Luo R, Wu J, Mao M, Liu X, et al: Icariin exterts negative
effects on human gastric cancer cell invasion and migration by
vasodilator-stimulated phosphoprotein via Rac1 pathway. Eur J
Pharmacol. 635:40–48. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li S, Dong P, Wang J, Zhang J, Gu J, Wu X,
Wu W, Fei X, Zhang Z, Wang Y, et al: Icariin, a natural flavonol
glycoside, induces apoptosis in human hepatoma SMMC-7721 cells via
a ROS/JNK-dependent mitochondrial pathway. Cancer Lett.
298:222–230. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gondi CS, Lakka SS, Dinh DH, Olivero WC,
Gujrati M and Rao JS: Downregulation of uPA, uPAR and MMP-9 using
small, interfering, hairpin RnA (siRnA) inhibits glioma cell
invasion, angiogenesis and tumor growth. Neuron Glia Biol.
1:165–176. 2004. View Article : Google Scholar
|