Introduction
Colorectal cancer (CRC) is a well-known complication
of ulcerative colitis (UC) and is associated with significant
mortality and morbidity. In UC, CRC has an incidence of 0.4% and a
prevalence of 3.5%. CRC accounts for 15% of mortalities in UC
patients (1,2). The risk of developing CRC in UC
increases proportionally to the disease duration, early onset, the
extent of disease involvement, increased inflammatory burden and
presence of primary sclerosing cholangitis (3–6). The
occurrence of UC-associated CRC differs from that of sporadic CRC.
UC-associated CRC develops from the mucosa with focal or multifocal
dysplasia in the presence of inflammation rather than the adenomas
and follows the sequence of inflammation-dysplasia-carcinoma
(7,8). Several pathways of tumorigenesis and
progression including chromosomal instability and microsatellite
instability pathways, the β-catenin/Wnt intermediary pathway,
TGFβ/SMAD intermediary pathway, and RAF/RAS/MAPK intermediary
pathway have been described in CRC carcinogenesis. Each of these
pathways involves gradual cumulating of multiple mutations that
differentiates the pathogenic mechanism. Genetic alternations such
as Ki-RAS and B-RAF mutations, 18q deletion, APC and p53 mutations
as well as genetic and epigenetic alternations such as DNA CpG
islet hypermethylation, non-coding RNA (miRNA) expression and
specific histone modifications are observed in many genes involved
in these pathways (7,9,10).
Epigenetic changes, such as histone modifications, that result in
subsequent aberrant gene expression are known to have key roles as
ultimate predictors in inflammation-induced colorectal
carcinogenesis. Previous findings suggest that histone modification
genes including NEK6, AURKA, PAK1,
HDAC1, HDAC2, HDAC3, HDAC5 and
HDAC7 have key roles in colorectal carcinogenesis (11–19).
There is a need for more effective markers to
identify patients who are at a higher risk of developing cancer in
order to improve CRC screening efficacy and provide more selective
treatment strategies in UC patients. At present, there are no
studies comparing epigenetic chromatin histone modification
patterns of UC and CRC in the literature. We therefore aimed in the
present study to investigate whether genes, particularly involved
in chromatin histone modification, have value in patient monitoring
with regards to CRC development in UC.
Materials and methods
Study location
The study was performed at the Departments of
Gastroenterology and Medical Biology of Manisa Celal Bayar
University between July, 2012 and November, 2013. Patients
presenting to the Department of Gastroenterology, Manisa Celal
Bayar University, were included in the present study.
Ethics committee
The present study was performed in accordance with
the declaration of Helsinki, good clinical practice and applicable
regulatory requirements. The study was carried out with the
approval of the Institutional Ethics Review Board of the Celal
Bayar University Medical Center with the approval number 200 dated
26/06/2012. Informed written consent was obtained from each
patient.
Study design and samples
Fresh matched tissue samples were collected from a
total of 60 patients presenting at the Department of
Gastroenterology, Celal Bayar University, between July, 2012 and
November, 2013 [group 1 (20 patients) was classified as the
macroscopically and histopathologically confirmed CRC group],
[group 2 (20 patients) was classified as the macroscopically and
histopathologically confirmed UC group], and [group 3 (20 patients)
was the control group and included patients with normal colon
mucosa with no macroscopically and histopathologically confirmed UC
or CRC]. Patients who had received chemotherapy or radiotherapy at
any time for any types of cancer, patients diagnosed with any
malignancies other than CRC, and patients who received colectomy
for any reason, were excluded from the study. Age, gender and
history/family history were questioned and documented for all the
patients. The extent of colonic involvement of the disease,
duration of the disease, activation severity, evidence of
extraintestinal involvement, routine evaluations and examinations,
results of pathological assessments and drugs used in UC treatment
were documented for data purposes in the UC group. For all the
patients diagnosed with CRC, CT or PET/CT was performed for staging
purposes.
Colonoscopy
Colonoscopic examinations of all the patients were
performed at the Department of Gastroenterology, Celal Bayar
University by the five gastroenterologists (E.K., H.Y., E.E., E.G.
and A.B.) who conducted the study using an Olympus Luxera CFQ260AL
colonoscopy unit. Complete colonoscopic examination including the
terminal ileum was performed for controls and UC patients. In
patients with CRC, ileocolonoscopic examination was completed to
determine whether the tumor obliterated the passage access. In CRC
patients in whom access to the proximal of the tumor could not be
made, proximal of the tumor was assessed using abdominal CT.
For histopathologic and genetic (histone
modification gene expression analyses) assessments, mucosal biopsy
samples were collected from the tumor tissue in the CRC group, from
rectal mucosa with active inflammation in the UC group, and from
rectal, cecal and terminal ileal mucosa in the control group with
the Olympus biopsy forceps. Pathologic assessments of all the
biopsy samples were performed in the Department of Pathology, Celal
Bayar University, and the resulting data were documented.
The tissue samples were immediately frozen using dry
ice (a block of dry ice has a surface temperature of −78.5°C) and
stored at −80°C until RNA extraction.
RNA isolation from tissue
RNA was isolated using an RNeasy mini kit (Qiagen,
Germany) according to the manufacturer's instructions with small
modifications. Tissue samples (20–30 mg) from patients, 600
µl buffer RLT, and a metal ball with a 7 mm diameter were
homogenized in TissueLyser II homogenizer (Qiagen) at 25,000 Hz for
5 min. The lysate was centrifuged at maximum speed and the
supernatant was treated after this point following the RNeasy mini
kit protocol.
Quantity and purity of total RNA
RNA was quantified measuring the absorbance at 260
nm (A260) and RNA purity was determined by the ratio A260/A280
using a spectrophotometer. RNA quality was considered to be
acceptable with A260/A280 at ratio slightly >2.0 and A260/A230
ratio at slightly >1.8.
cDNA synthesis
The cDNA synthesis was performed using an RT First
Strand kit (C-03) (SABiosciences, Frederick, MD, USA). RNA sample
(8 µl) was incubated with 2 µl of GE (5X gDNA
elimination buffer) at 42°C for 5 min in a 0.2 ml PCR tube. In
another tube a PCR cocktail [4 µl BC3 (5X RT buffer 3), 1
µl P2 (primer and external control mix), 2 µl RE3 (RT
enzyme mix 3)], and 3 µl H2O were prepared and
added onto the RNA sample followed by a 15-min incubation at 42°C
and 5-min incubation at 95°C. The cDNA samples were later diluted
as required.
Human epigenetic chromatin modification
enzymes RT2 profiler™ PCR array
The Human Epigenetic Chromatin Modification Enzyme
RT2 Profiler™ PCR Array (PAHS-085A) (SABiosciences) was
used to detect the expression levels of 84 key genes listed in
Table I.
 | Table IHistone modification genes to be
investigated. |
Table I
Histone modification genes to be
investigated.
| Histone
modifications | Genes |
|---|
| DNA
methyltransferase | DNMT1,
DNMT3A, DNMT3B |
| Histone
methyltransferase | ATF2,
CDYL, CIITA, CSRP2BP, ESCO1,
ESCO2, HAT1, KAT2A (GCN5L2),
KAT2B (PCAF), KAT5 (HTATIP),
MYST1, MYST2, MYST3, MYST4,
NCOA1, NCOA3, NCOA6 |
| Histone
methyltransferase | CARM1
(PRMT4), DOT1L, EHMT2, MLL1,
MLL3, PRMT1, PRMT2, PRMT3,
PRMT5, PRMT6, PRMT7, PRMT8,
SETDB2, SMYD3, SUV39H1 |
| SET domain proteins
(activation of histone methyltransferase) | ASH1L,
MLL3, MLL5, NSD1, SETD1A,
SETD1B, SETD2, SETD3, SETD4,
SETD5, SETD6, SETD7, SETD8,
SETDB1, SUV39H1, SUV420H1, WHSC1 |
| Histone
phosphorylation | AURKA,
AURKB, AURKC, NEK6, PAK1,
RPS6KA3, RPS6KA5 |
| Histone
ubiquination | DZIP3,
MYSM1, RNF2, RNF20, UBE2A,
UBE2B, USP16, USP21, USP22 |
| DNA/histone
demethylase | AOF2,
JARID1B, JARID1C, JMJD2A, JMJD2C,
JMJD3, MBD2 |
| Histone
deacetylase | HDAC1,
HDAC2, HDAC3, HDAC4, HDAC5,
HDAC6, HDAC7, HDAC8, HDAC9,
HDAC10, HDAC11 |
A total of 2,300 µl RT-PCR array mix (102
µl diluted cDNA, 1,150 µl 2X RT2
SYBR-Green ROX FAST master mix and 1,048 µl H2O)
was prepared and loaded into 96-well Human Epigenetic Chromatin
Modification Enzymes RT2 Profiler™ PCR Arrays 20
µl/well. Arrays were placed in Rotor-Gene RG-3000 (Corbett
Research, Qiagen, Germany) and were initially activated at 95°C for
10 min followed by 40 cycles of 95°C for 15 sec and 60°C for 30 sec
incubation. Array data were analyzed and 8 genes were determined to
be the most overexpressed or underexpressed, and crossvalidates
using RT-qPCR.
RT2 qPCR primer assay
The cDNA samples of the 8 genes selected to be
overexpressed or underexpressed (AURKB, AURKA,
SETD8, PAK1, NEK6, KDM4C, HDAC1
and HDAC7) and the housekeeping gene (HPRT1) were
also examined by RT-qPCR. The primer assays and SYBR-Green master
mix (SABiosciences) specific for the 8 genes were purchased and
each PCR reaction mix was prepared by adding 12.5 µl of
SYBR-Green master mix and 1 µl of primer (10 pmol)/2.2
µl cDNA sample. The total volume was adjusted to 25
µl with dH2O and PCR mixes were loaded on the
plate of Rotor-Gene RG-3000 (Corbett Research) for amplification.
Amplification was performed with an initial denaturation step at
95°C for 5 min followed by 40 cycles of 94°C for 1 min, 61°C for 40
sec, and 72°C for 1 min with a final elongation step at 72°C for 2
min. Cycle threshold (Ct) values obtained from the RT-qPCR
reactions were normalized to the reference gene (HPRT1) and
evaluated using REST 2009 (Relative Expression Software Tool
V.2.0.13) in standard mode.
Data analysis
Data were analyzed using RT2 profiler PCR
array and RT2 qPCR primer assay data analysis software
(http://www.sabiosciences.com/pcrarraydataanalysis.php).
PCR array was quantified based on the Ct number. A gene was
considered to be not detectable when Ct was >32. Ct was defined
as 35 for the ΔCt calculation when the signal was under detectable
limits. Fold-change and fold-regulation values >2 were
indicative of upregulated genes, whereas the fold-change values
<0.5 and fold-regulation values <−2 were indicative of
downregulated genes.
Statistical analysis
Results were presented as the mean values ± standard
deviation and the p-values were calculated based on a Student's
t-test of the replicate 2-ΔCt values for each gene in the control,
CRC and UC groups. P<0.05 was considered to indicate a
statistically significant result.
Results
Patients and histone modification gene
expression analysis
Sixty patients were included in the study. They were
divided into three groups, 20 patients in the CRC group
(histopathologically confirmed), 20 patients in the UC group
(histopathologically confirmed), and 20 patients in the control
group (normal colon mucosa without any macroscopic and
histopathologic finding of UC or CRC). The patients in the study
group had a mean age of 57.80±12.3 years. Mean ages in individual
groups were 59.12±12.1, 56.85±12.6 and 57.45±11.5 years in the CRC,
UC and the control groups, respectively The three groups did not
differ significantly with regards to age (p=0.284). The proportion
of male gender in total was 51.7% (n=31). Gender distribution in
the CRC, UC and control groups did not statistically differ (CRC
M/F, 9/11; UC M/F, 11/9; control M/F, 9/11; and p=0.339).
Sites of CRCs were assessed and the majority of the
tumors were located in the distal colon. The proportion of the CRC
patients by site of involvement were as follows: rectum, 35% (n=7);
sigmoid colon, 35% (n=7); descending colon, 5% (n=1); transverse
colon, 10% (n=2); ascending colon, 10% (n=2); and cecum, 5% (n=1).
Pathological diagnosis was adenocarcinoma for all the CRC patients.
Of the patients, 25% (n=5) had poorly differentiated, 70% (n=14)
had moderately differentiated tumors and only 5% (n=1) had
well-differentiated tumors.
In CRC patients (n=20), overexpression of the
AURKA, AURKB, HDAC1, HDAC7, NEK6
and PAK1 genes was statistically significantly higher
compared to the control group (fold-change >2) (Figs. 1 and 2). CRC and control groups did not differ
statistically significantly with regards to the expression of
SETD8 or KDM4C genes (fold-change <2, p>0.05)
(Table II). In UC (n=20) patients,
overexpression of the AURKA, HDAC1, NEK6 and
PAK1 genes was greater as compared to the control group
(fold-change >2) (Figs. 1 and
3). The UC and control groups did
not differ statistically significantly with regard to the
expression of AURKB, HDAC7, SETD8 or
KDM4C genes (fold-change <2, p>0.05) (Table III). The overexpression of the
AURKB and HDAC7 genes was statistically significantly
higher in the CRC group compared to the UC group (p<0.05). The
UC and CRC groups did not statistically differ significantly with
regard to the overexpression of the AURKA, NEK6,
PAK1 and HDAC1 genes (Table IV).
| Table IIHistone modification gene expression
analysis in CRC. |
Table II
Histone modification gene expression
analysis in CRC.
| Histone gene | Fold-change | 95% CI | P-value |
|---|
| AURKB | 7.3871 | (1.54–13.23) | 0.00125 |
| AURKA | 6.9307 | (1.48–12.38) | 0.00271 |
| SETD8 | 1.3491 | (0.33–2.37) | 0.43853 |
| PAK1 | 6.4733 | (1.01–11.94) | 0.00152 |
| NEK6 | 7.7355 | (1.19–14.28) | 0.00090 |
| KDM4C | 0.6849 | (0.17–1.20) | 0.11222 |
| HDAC1 | 7.4876 | (1.33–13.65) | 0.00313 |
| HDAC7 | 8.0948 | (1.39–14.80) | 0.00293 |
| HPRT1 | 0.9211 | (0.69–1.16) | 0.58991 |
| Table IIIHistone modification gene expression
analysis in UC. |
Table III
Histone modification gene expression
analysis in UC.
| Histone gene | Fold-change | 95% CI | P-value |
|---|
| AURKB | 1.4162 | (0.36–2.47) | 0.16986 |
| AURKA | 7.4384 | (1.02–13.85) | 0.00486 |
| SETD8 | 0.8322 | (0.00–1.66) | 0.48752 |
| PAK1 | 7.953 | (1.02–14.89) | 0.00542 |
| NEK6 | 9.161 | (1.83–16.49) | 0.00107 |
| KDM4C | 0.7711 | (0.00–1.54) | 0.26249 |
| HDAC1 | 8.3195 | (1.19–15.45) | 0.00549 |
| HDAC7 | 1.5988 | (0.36–2.84) | 0.20862 |
| HPRT1 | 0.9727 | (0.72–1.23) | 0.96950 |
| Table IVComparison of the gene expression of
the CRC and UC groups. |
Table IV
Comparison of the gene expression of
the CRC and UC groups.
| Histone gene | Fold-change | 95% CI | P-value |
|---|
| AURKB | 5.2162 | (0.46–9.97) | 0.00796 |
| AURKA | 0.9317 | (0.00001–1.89) | 0.40211 |
| SETD8 | 1.6211 | (0.22–3.02) | 0.26484 |
| PAK1 | 0.8139 | (0.04–1.59) | 0.34676 |
| NEK6 | 0.8444 | (0.02–1.67) | 0.79039 |
| KDM4C | 0.8882 | (0.07–1.71) | 0.17068 |
| HDAC1 | 0.9 | (0.06–1.74) | 0.46234 |
| HDAC7 | 5.063 | (0.40–9.72) | 0.01954 |
| HPRT1 | 0.947 | (0.69–1.21) | 0.64247 |
UC subgroup gene expression analyses
Gene expression analyses by colon
involvement
The UC patients (n=20) were subdivided into two
subgroups by colonic involvement as follows: the patients with
involvement that is proximal to the splenic flexure (55%, n=11)
(extensive colitis or pancolitis), and left colon involvement
including pouchitis, (45%, n=9). In the UC subgroup with extensive
colitis or pancolitis, the overexpression of the NEK6 and
AURKA genes was statistically significantly higher than that
in the patients with left colon involvement (Fig. 4 and Table V).
| Table VGene expression analysis by colon
involvement in UC. |
Table V
Gene expression analysis by colon
involvement in UC.
| Histone gene | Fold-change | 95% CI | P-value |
|---|
| AURKB | 1.3494 | (0.00001–2.95) | 0.24876 |
| AURKA | 5.3678 |
(0.00001–12.55) | 0.02699 |
| SETD8 | 0.7484 | (0.00001–1.88) | 0.31268 |
| PAK1 | 2.2402 | (0.00001–5.16) | 0.06460 |
| NEK6 | 6.7131 |
(0.00001–13.96) | 0.01489 |
| KDM4C | 1.1199 | (0.00001–2.85) | 0.21034 |
| HDAC1 | 3.1064 | (0.00001–7.06) | 0.14044 |
| HDAC7 | 2.0928 | (0.00001–4.59) | 0.26969 |
| HPRT1 | 1 | (1.00–1.00) | – |
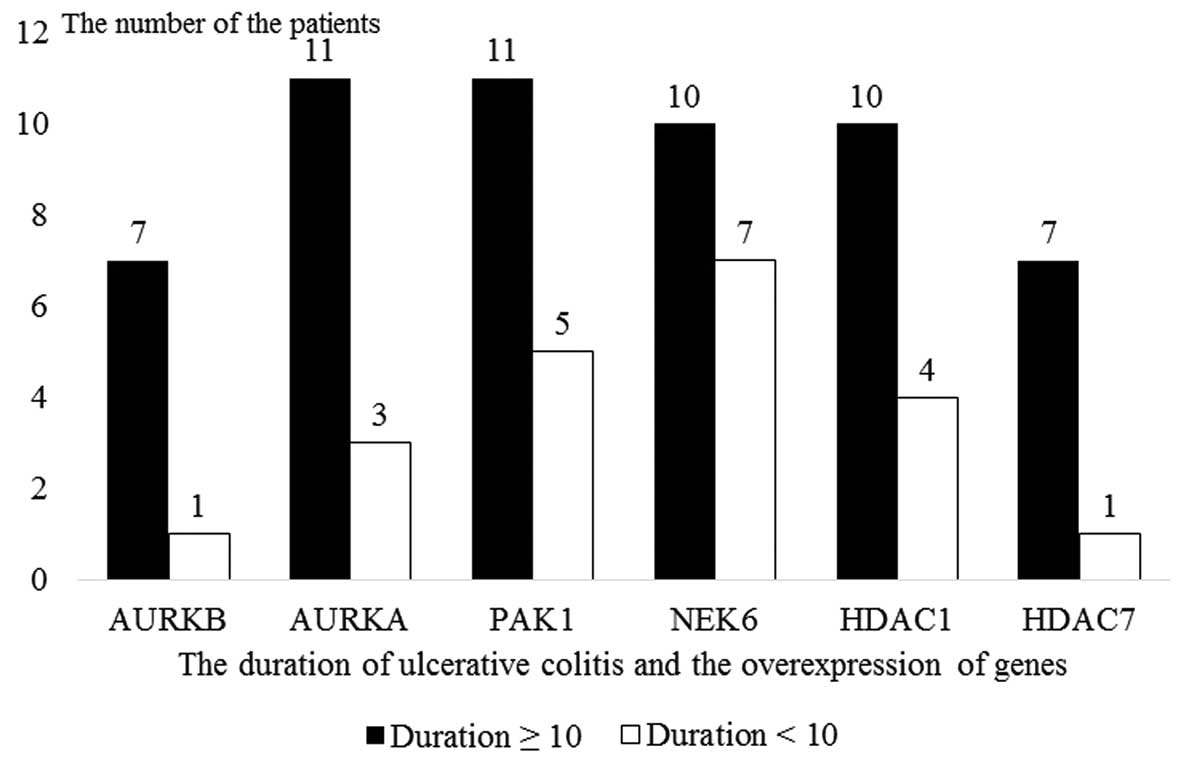
Gene expression analyses by disease
duration
The patients diagnosed with UC were subdivided into
two subgroups by disease duration as those with a disease duration
of ≥10 years (60%, n=12) and those a disease duration of <10
years (40%, n=8). In patients with a disease duration of ≥10 years,
the over-expression of the NEK6, AURKA, PAK1
and HDAC1 genes was statistically significantly higher
compared to those with a disease duration of <10 years (Fig. 5 and Table VI).
| Table VIGene expression analysis by disease
duration in UC. |
Table VI
Gene expression analysis by disease
duration in UC.
| Histone gene | Fold-change | 95% CI | P-value |
|---|
| AURKB | 5.8903 | (0.01–11.77) | 0.165124 |
| AURKA | 14.1641 | (0.43–27.90) | 0.012857 |
| SETD8 | 2.2211 | (0.00001–5.51) | 0.284641 |
| PAK1 | 10.1085 | (1.15–19.06) | 0.015182 |
| NEK6 | 3.388 | (0.00001–7.06) | 0.013988 |
| KDM4C | 3.1032 | (0.00001–7.46) | 0.223753 |
| HDAC1 | 8.959 | (1.39–16.53) | 0.013921 |
| HDAC7 | 4.2673 | (0.00001–8.67) | 0.231647 |
| HPRT1 | 1 | (1.00–1.00) | – |
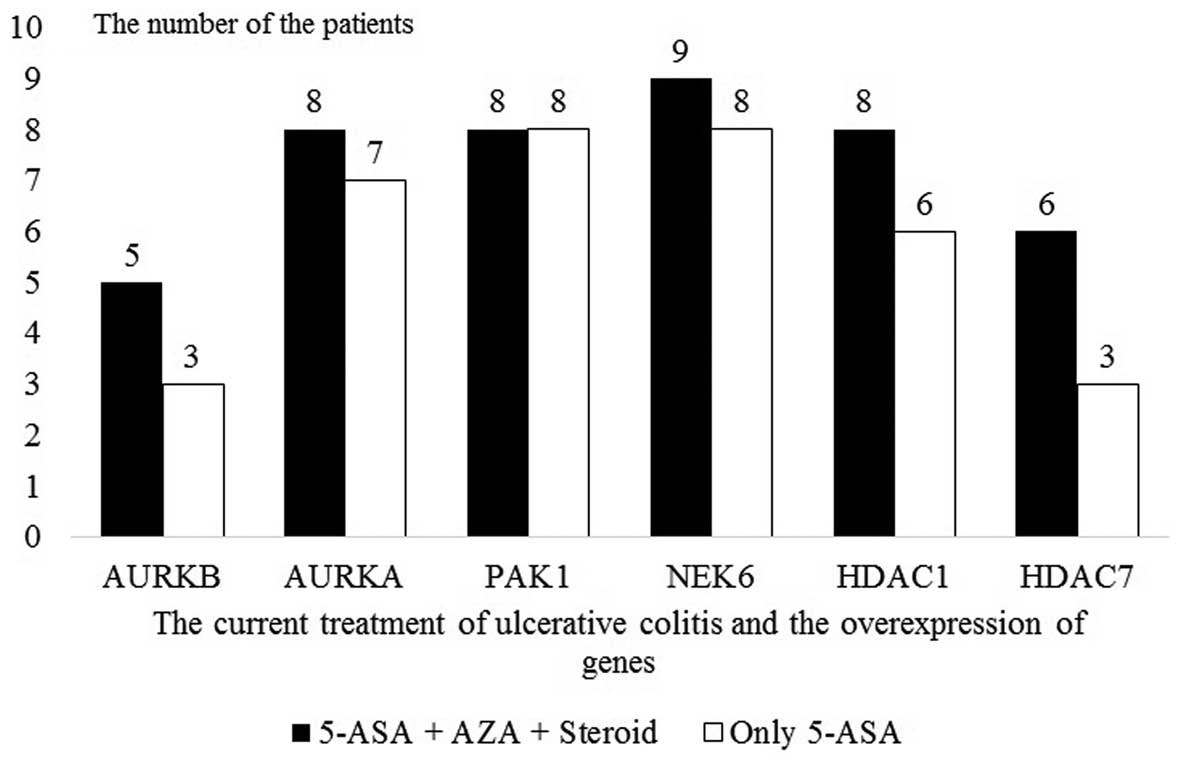
Gene expression analyses by disease
severity
None of the UC patients had a history of treatment
with an antitumor necrosis factor (anti-TNF) drug, a calcineurin
inhibitor such as cyclosporin and immunomodulator agents such as
azathiopurine and mycophenolate mofetil. The UC patients were on
oral and/or rectal 5-ASA treatment. The UC patients were divided
into two subgroups by the medications used during periods of
exacerbation and remission as an indicator of disease behavior as
follows: patients who received steroid treatment for remission
induction and azathioprine for remission maintenance (55%, n=11)
and those who received only 5-ASA (45%, n=9). The two subgroups did
not differ statistically significantly with regards to gene
expression (Fig. 6 and Table VII).
| Table VIIGene expression analysis by
treatments received by patients in UC. |
Table VII
Gene expression analysis by
treatments received by patients in UC.
| Histone gene | Fold-change | 95% CI | P-value |
|---|
| AURKB | 0.4597 | (0.00001–1.01) | 0.24561 |
| AURKA | 0.3842 | (0.00001–0.97) | 0.23059 |
| SETD8 | 0.2455 | (0.00001–0.58) | 0.14288 |
| PAK1 | 0.2564 | (0.00001–0.58) | 0.06694 |
| NEK6 | 0.698 | (0.00001–1.67) | 0.73757 |
| KDM4C | 0.3124 | (0.00001–0.77) | 0.17477 |
| HDAC1 | 0.4711 | (0.00001–1.12) | 0.42449 |
| HDAC7 | 0.5182 | (0.00001–1.12) | 0.23014 |
| HPRT1 | 1 | (1.00–1.00) | – |
Discussion
Chronic inflammation has a significant role in many
types of cancer including Helicobacter pylori-associated
gastric cancer, hepatitis C-associated hepatocellular cancer and
colitis-associated CRC (20).
Development of inflammation associated with cancer is characterized
by the gradual accrual of genetic and epigenetic alterations in
several proto-oncogene and tumor-suppressor genes (20). Understanding of the epigenetic
changes in cancer cells has led to gaining insight on the stages of
carcinogenesis. It has been shown that epigenetic changes has
similar effects as genetic changes on the stages of carcinogenesis
and that they trigger carcinogenesis (21). As our knowledge on cancer molecular
biology expands monitoring of precancerous and cancerous stages
improves (22,23). The fact that epigenetic changes,
unlike genetic ones, are reversible supports the hypothesis that
epigenetic treatments may be used in the future (24).
In the present study, NEK6 and AURKA
genes in particular were overexpressed in the patients with UC and
CRC, supporting our hypothesis that the two genes are of value as
diagnostic and prognostic biomarkers in UC and CRC patients.
HDAC1, HDAC7, PAK1 and AURKB genes were
significantly more overexpressed in CRC patients compared to the
controls. Additionally, HDAC1 and PAK1 genes were
more overexpressed in UC patients compared to the controls. The
NEK6 and AURKA genes are involved in the epigenetic
mechanisms of the formation of most of the gastrointestinal
cancers. Although dysplasia was not detected in any of the UC
patients there were no differences between NEK6 and
AURKA gene expression in UC and CRC patients. There was a
significant overexpression in the HDAC7 and AURKB
genes compared between the UC and CRC groups.
p-21 activated kinase (PAK1) coordinates
RAS/RAF/MAPK and Wnt/β-catenin signaling in the initiation and
progression of CRC carcinogenesis by acting on the flow of small
GTPases (25–27). Dammann et al demonstrated
that PAK1 has an important role in the carcinogenesis of
colitis-associated CRC and suggested that it may be a therapeutic
target with potential use in chemoprevention (28). Similarly, an experimental model by
He et al (18) demonstrated
that 5-ASA normalized the increased membranous expression of the
adhesion molecules E-cadherin and β-catenin in the carcinogenesis
of CRC. The authors also reported that 5-ASA suppressed PAK1
expression and neoplastic progression in the carcinogenesis of CRC
(17). In the stages of
colitis-associated carcinogenesis, intestinal PAK1 expression
increased in the presence of inflammation and malignant
transformation. PAK1 was involved in the carcinogenesis of
colitis-associated CRC by maintaining the colaboration of
mitogen-activated kinase, phosphoinositide 3-kinase (PI3K/AKT),
nuclear factor κB (NF-κB) and Wnt/β-catenin pathways and ensuring
continuance of these pathways (28).
Histone deacetylase 1 (HDAC1) controls cell
proliferation, differentiation, migration and apoptosis by
suppressing p21 and p27 cyclin-dependent kinase (CDK) (29). HDAC1 is involved in the
carcinogenesis stages of numerous GIS cancers including CRC, and
HDAC1 overexpression is associated with tumor differentiation,
increased proliferation, increased invasion, advanced disease and
poor prognosis (12,14,30–34).
Turgeon et al demonstrated that HDAC1 regulated the
proliferation and differentiation of the intestinal and colonic
epithelium in the inflammatory response (35). It has also been shown that
inflammation and tumor formation may be suppressed in experimental
colitis and inflammation-induced tumorigenesis models owing to the
antiproliferative properties of HDAC inhibitors (16).
The members of the aurora kinase family (AURKA,
AURKB and AURKC) are serine/threonine kinases involved as key
regulators in mitosis. AURKA regulates the G2/M phase of the cell
cycle by acting on the regulation of centrosome functions, spindle
montage (linkage), chromosome separation and cytokinesis. AURKA
mRNA is amplified in numerous types of cancers including CRC
(36–38). A study evaluating 386 CRC subjects
found that a high AURKA expression was associated with increased
recurrence/relapse frequency and suggested that AURKA may be used
as a prognostic marker (15). AURKA
is overexpressed in inflamed esophagitis tissue with precancerous
lesion in esophageal adenocarcinoma (39) and in Barrett's esophagus secondary
to chronic irritation/inflammation (40), suggesting that it is involved in the
carcinogenesis associated with inflammation. A study by Katsha
et al, investigated the role of AURKA in the tumorigenesis
of inflammation-associated gastric cancer, compared healthy stomach
tissue with premalignant and malignant lesions and found higher
levels of AURKA protein and nuclear NF-κB levels (41). The authors described that AURKA
overexpression initiated and maintained NF-κB and tumorigenesis
activation in a chronic inflammatory process. The authors also
suggested that AURKA inhibitors may be developed as therapeutic
agents for gastric cancer (41).
NEK6 is a serine/threonine kinase from the never in
mitosis gene A (NIMA)-related kinase 6 family involved in mitotic
division (42). Termination of
mitosis, chromatin spindle defects and abnormal chromosome
differentiation have been observed and apoptosis was triggered with
NEK6 dysfunction (43–45). Nassirpour et al demonstrated
that the kinase activity of NEK6 was increased in several malignant
human cancer cells including breast, uterus, colon, stomach, ovary,
lung, kidney, rectum, thyroid, cervix, prostate, pancreas and small
intestine cancers, and that suppression of NEK6 resulted in tumor
regression in a mice xenograft model (19). A study by Kasap et al
demonstrated that the NEK6 gene was overexpressed in tissues
with esophagitis to a similar extent as esophageal adenocarcinomas,
and that the extent of NEK6 gene overexpression increased
proportionally with the increasing severity of esophagitis
(39).
To the best of our knowledge, this is the first
study to show that the NEK6, AURKA, PAK1 and
HDAC1 genes were significantly elevated in human UC tissues.
In light of the results reported from the studies mentioned above
and those obtained in the present study, it was identified that the
NEK6 and AURKA genes, in particular, as well as the
PAK1 and HDAC1 genes play a significant role in the
stages of UC-associated CRC, and that they have a prognostic
relevance in CRC. Additionally, the four histone modification
enzyme genes can be used in anticancer treatments as therapeutic
targets. Of note, these results were identified despite the absence
of dysplasia in the subjects with UC. This finding suggests that
the four genes are involved in the early stages of
inflammation-associated carcinogenesis during which dysplasia did
not occur. The four genes, particularly NEK6 and
AURKA, can be used as early markers in the development of
CRC in UC patients.
The absence of a significant difference in
NEK6 and AURKA gene expression between the UC and CRC
patient groups, as determined in the present study, can be
considered to reflect the predisposition of UC patients to CRC.
The risk of developing CRC in UC increases
proportionally to the duration of disease, early age of disease
onset, the extent of colonic involvement, inflammation severity and
presence of primary sclerosing cholangitis (3–6). The
present results have demonstrated that the NEK6, AURKA,
PAK1 and HDAC1 genes were significantly more
overexpressed with an increasing UC disease duration, a risk factor
for CRC, in patients diagnosed with UC. In a meta-analysis of 116
individual studies by Eaden et al, the cumulative CRC risk
in UC patients was 2% after 10 years, 8% after 20 years and 18%
after 30 years, and the study thus recommended colonoscopic
screening for CRC risk after 8 years for patients with pancolitis
and after 12 years for those with left colitis (1). Based on these data, detection of these
genes can be of additional benefit to colonoscopic screening in UC.
The present study did not identify a link between the NEK6,
AURKA, PAK1 and HDAC1 genes and inflammation
severity. This indicates that the epigenetic changes involved in
CRC stages with long-standing chronic inflammation occur to a
greater extent with time regardless of the inflammation severity.
Consistent with the results of the study by Kasap et al
(39) comparing the histone
modification profiles in esophageal adenocarcinoma and esophagitis
tissues, the AURKA and NEK6 genes were more
overexpressed in UC patients with diffuse colon involvement,
whereas PAK1 and HDAC1 overexpression was independent of the extent
of disease involvement. While this may be due to the inadequate
number of subjects, it can also be explained by the chemoprevention
induced by 5-ASA, which was received by all UC patients in the
present study, or by the active presence of PAK1 and HDAC1 in the
stages of inflammation that precede the occurrence of
carcinogenesis (16,17,35).
In conclusion, the results show that the
overexpression particularly of the AURKA and NEK6
genes in patients with UC and CRC in the Turkish population.
Clarification of the molecular pathophysiology of CRC and UC
contributes to the treatment and prognosis of the diseases. Future
studies are needed to understand the roles of the four genes in the
carcinogenesis associated with colitis. Thus, NEK6 and AURKA can be
considered promising genetic markers.
Acknowledgments
This study was supported by the Celal Bayar
University Coordinator of the Scientific Research Projects
(2013-09) Manisa, Turkey.
Abbreviations:
|
AURKA
|
aurora kinase A
|
|
AURKB
|
aurora kinase B
|
|
CRC
|
colorectal adenocarcinoma
|
|
HDAC1
|
histone deacetylase 1
|
|
HDAC7
|
histone deacetylase 7
|
|
NEK6
|
never in mitosis gene A-related kinase
6
|
|
PAK1
|
p-21 activated kinase
|
|
UC
|
ulcerative colitis
|
References
|
1
|
Eaden JA, Abrams KR and Mayberry JF: The
risk of colorectal cancer in ulcerative colitis: A meta-analysis.
Gut. 48:526–535. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ekbom A, Helmick C, Zack M and Adami HO:
Ulcerative colitis and colorectal cancer. A population-based study.
N Engl J Med. 323:1228–1233. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Claessen MM, Lutgens MW, van Buuren HR,
Oldenburg B, Stokkers PC, van der Woude CJ, Hommes DW, de Jong DJ,
Dijkstra G, van Bodegraven AA, et al: More right-sided
IBD-associated colorectal cancer in patients with primary
sclerosing cholangitis. Inflamm Bowel Dis. 15:1331–1336. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gillen CD, Walmsley RS, Prior P, Andrews
HA and Allan RN: Ulcerative colitis and Crohn's disease: A
comparison of the colorectal cancer risk in extensive colitis. Gut.
35:1590–1592. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rutter M, Saunders B, Wilkinson K, Rumbles
S, Schofield G, Kamm M, Williams C, Price A, Talbot I and Forbes A:
Severity of inflammation is a risk factor for colorectal neoplasia
in ulcerative colitis. Gastroenterology. 126:451–459. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Winther KV, Jess T, Langholz E, Munkholm P
and Binder V: Long-term risk of cancer in ulcerative colitis: A
population-based cohort study from Copenhagen County. Clin
Gastroenterol Hepatol. 2:1088–1095. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kulaylat MN and Dayton MT: Ulcerative
colitis and cancer. J Surg Oncol. 101:706–712. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rhodes JM and Campbell BJ: Inflammation
and colorectal cancer: IBD-associated and sporadic cancer compared.
Trends Mol Med. 8:10–16. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jawad N, Direkze N and Leedham SJ:
Inflammatory bowel disease and colon cancer. Recent Results Cancer
Res. 185:99–115. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Migheli F and Migliore L: Epigenetics of
colorectal cancer. Clin Genet. 81:312–318. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Stypula-Cyrus Y, Damania D, Kunte DP, Cruz
MD, Subramanian H, Roy HK and Backman V: HDAC up-regulation in
early colon field carcinogenesis is involved in cell tumorigenicity
through regulation of chromatin structure. PLoS One. 8:e646002013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Higashijima J, Kurita N, Miyatani T,
Yoshikawa K, Morimoto S, Nishioka M, Iwata T and Shimada M:
Expression of histone deacetylase 1 and metastasis-associated
protein 1 as prognostic factors in colon cancer. Oncol Rep.
26:343–348. 2011.PubMed/NCBI
|
|
13
|
Wilson AJ, Byun DS, Popova N, Murray LB,
L'Italien K, Sowa Y, Arango D, Velcich A, Augenlicht LH and
Mariadason JM: Histone deacetylase 3 (HDAC3) and other class I
HDACs regulate colon cell maturation and p21 expression and are
deregulated in human colon cancer. J Biol Chem. 281:13548–13558.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Witt O, Deubzer HE, Milde T and Oehme I:
HDAC family: What are the cancer relevant targets? Cancer Lett.
277:8–21. 2009. View Article : Google Scholar
|
|
15
|
Belt EJ, Brosens RP, Delis-van Diemen PM,
Bril H, Tijssen M, van Essen DF, Heymans MW, Beliën JA, Stockmann
HB, Meijer S, et al: Cell cycle proteins predict recurrence in
stage II and III colon cancer. Ann Surg Oncol. 19(Suppl 3):
S682–S692. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Glauben R, Sonnenberg E, Zeitz M and
Siegmund B: HDAC inhibitors in models of inflammation-related
tumorigenesis. Cancer Lett. 280:154–159. 2009. View Article : Google Scholar
|
|
17
|
Khare V, Lyakhovich A, Dammann K, Lang M,
Borgmann M, Tichy B, Pospisilova S, Luciani G, Campregher C,
Evstatiev R, et al: Mesalamine modulates intercellular adhesion
through inhibition of p-21 activated kinase-1. Biochem Pharmacol.
85:234–244. 2013. View Article : Google Scholar :
|
|
18
|
He H, Huynh N, Liu KH, Malcontenti-Wilson
C, Zhu J, Christophi C, Shulkes A and Baldwin GS: P-21 activated
kinase 1 knockdown inhibits β-catenin signalling and blocks
colorectal cancer growth. Cancer Lett. 317:65–71. 2012. View Article : Google Scholar
|
|
19
|
Nassirpour R, Shao L, Flanagan P, Abrams
T, Jallal B, Smeal T and Yin MJ: Nek6 mediates human cancer cell
transformation and is a potential cancer therapeutic target. Mol
Cancer Res. 8:717–728. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chiba T, Marusawa H and Ushijima T:
Inflammation-associated cancer development in digestive organs:
Mechanisms and roles for genetic and epigenetic modulation.
Gastroenterology. 143:550–563. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Feinberg AP, Ohlsson R and Henikoff S: The
epigenetic progenitor origin of human cancer. Nat Rev Genet.
7:21–33. 2006. View Article : Google Scholar
|
|
22
|
Lao VV and Grady WM: Epigenetics and
colorectal cancer. Nat Rev Gastroenterol Hepatol. 8:686–700. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yi JM, Dhir M, Guzzetta AA,
Iacobuzio-Donahue CA, Heo K, Yang KM, Suzuki H, Toyota M, Kim HM
and Ahuja N: DNA methylation biomarker candidates for early
detection of colon cancer. Tumour Biol. 33:363–372. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yoo CB and Jones PA: Epigenetic therapy of
cancer: Past, present and future. Nat Rev Drug Discov. 5:37–50.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
He TC, Sparks AB, Rago C, Hermeking H,
Zawel L, da Costa LT, Morin PJ, Vogelstein B and Kinzler KW:
Identification of c-MYC as a target of the APC pathway. Science.
281:1509–1512. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Polakis P: The many ways of Wnt in cancer.
Curr Opin Genet Dev. 17:45–51. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Vogelstein B, Fearon ER, Hamilton SR, Kern
SE, Preisinger AC, Leppert M, Nakamura Y, White R, Smits AM and Bos
JL: Genetic alterations during colorectal-tumor development. N Engl
J Med. 319:525–532. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dammann K, Khare V and Gasche C: Tracing
PAKs from GI inflammation to cancer. Gut. 63:1173–1184. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lagger G, O'Carroll D, Rembold M, Khier H,
Tischler J, Weitzer G, Schuettengruber B, Hauser C, Brunmeir R,
Jenuwein T, et al: Essential function of histone deacetylase 1 in
proliferation control and CDK inhibitor repression. EMBO J.
21:2672–2681. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Choi JH, Kwon HJ, Yoon BI, Kim JH, Han SU,
Joo HJ and Kim DY: Expression profile of histone deacetylase 1 in
gastric cancer tissues. Jpn J Cancer Res. 92:1300–1304. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Miyake K, Yoshizumi T, Imura S, Sugimoto
K, Batmunkh E, Kanemura H, Morine Y and Shimada M: Expression of
hypoxia-inducible factor-1alpha, histone deacetylase 1, and
metastasis-associated protein 1 in pancreatic carcinoma:
Correlation with poor prognosis with possible regulation. Pancreas.
36:e1–e9. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Weichert W, Röske A, Gekeler V, Beckers T,
Ebert MP, Pross M, Dietel M, Denkert C and Röcken C: Association of
patterns of class I histone deacetylase expression with patient
prognosis in gastric cancer: A retrospective analysis. Lancet
Oncol. 9:139–148. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Weichert W, Röske A, Gekeler V, Beckers T,
Stephan C, Jung K, Fritzsche FR, Niesporek S, Denkert C, Dietel M,
et al: Histone deacetylases 1, 2 and 3 are highly expressed in
prostate cancer and HDAC2 expression is associated with shorter PSA
relapse time after radical prostatectomy. Br J Cancer. 98:604–610.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Weichert W, Röske A, Niesporek S, Noske A,
Buckendahl AC, Dietel M, Gekeler V, Boehm M, Beckers T and Denkert
C: Class I histone deacetylase expression has independent
prognostic impact in human colorectal cancer: Specific role of
class I histone deacetylases in vitro and in vivo. Clin Cancer Res.
14:1669–1677. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Turgeon N, Blais M, Gagné JM, Tardif V,
Boudreau F, Perreault N and Asselin C: HDAC1 and HDAC2 restrain the
intestinal inflammatory response by regulating intestinal
epithelial cell differentiation. PLoS One. 8:e737852013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Marumoto T, Zhang D and Saya H: Aurora-A -
a guardian of poles. Nat Rev Cancer. 5:42–50. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bischoff JR, Anderson L, Zhu Y, Mossie K,
Ng L, Souza B, Schryver B, Flanagan P, Clairvoyant F, Ginther C, et
al: A homologue of Drosophila aurora kinase is oncogenic and
amplified in human colorectal cancers. EMBO J. 17:3052–3065. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Carvalho B, Postma C, Mongera S, Hopmans
E, Diskin S, van de Wiel MA, van Criekinge W, Thas O, Matthäi A,
Cuesta MA, et al: Multiple putative oncogenes at the chromosome 20q
amplicon contribute to colorectal adenoma to carcinoma progression.
Gut. 58:79–89. 2009. View Article : Google Scholar
|
|
39
|
Kasap E, Boyacioglu SO, Korkmaz M, Yuksel
ES, Unsal B, Kahraman E, Ozütemiz O and Yuceyar H: Aurora kinase A
(AURKA) and never in mitosis gene A-related kinase 6 (NEK6) genes
are upregulated in erosive esophagitis and esophageal
adenocarcinoma. Exp Ther Med. 4:33–42. 2012.PubMed/NCBI
|
|
40
|
Rugge M, Fassan M, Zaninotto G, Pizzi M,
Giacomelli L, Battaglia G, Rizzetto C, Parente P and Ancona E:
Aurora kinase A in Barrett's carcinogenesis. Hum Pathol.
41:1380–1386. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Katsha A, Soutto M, Sehdev V, Peng D,
Washington MK, Piazuelo MB, Tantawy MN, Manning HC, Lu P, Shyr Y,
et al: Aurora kinase A promotes inflammation and tumorigenesis in
mice and human gastric neoplasia. Gastroenterology.
145:1312–1322.e1–8. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Jee HJ, Kim AJ, Song N, Kim HJ, Kim M, Koh
H and Yun J: Nek6 overexpression antagonizes p53-induced senescence
in human cancer cells. Cell Cycle. 9:4703–4710. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Belham C, Roig J, Caldwell JA, Aoyama Y,
Kemp BE, Comb M and Avruch J: A mitotic cascade of NIMA family
kinases. Nercc1/Nek9 activates the Nek6 and Nek7 kinases. J Biol
Chem. 278:34897–34909. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
O'Regan L and Fry AM: The Nek6 and Nek7
protein kinases are required for robust mitotic spindle formation
and cytokinesis. Mol Cell Biol. 29:3975–3990. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Cao X, Xia Y, Yang J, Jiang J, Chen L, Ni
R, Li L and Gu Z: Clinical and biological significance of never in
mitosis gene A-related kinase 6 (NEK6) expression in hepatic cell
cancer. Pathol Oncol Res. 18:201–207. 2012. View Article : Google Scholar
|