Introduction
Bone homeostasis is maintained by the coordinated
action of osteoclasts, osteoblasts and osteocytes in bone tissues
(1). Bone loss occurs when the rate
of osteoclastic bone resorption exceeds that of osteoblastic bone
formation (2,3). The consequence of bone loss is the
development of osteoporosis, a condition that significantly
increases the propensity for bone fractures, a major public health
threat. Numerous pathologic conditions promote bone loss including
estrogen deficiency, inflammation, bacterial and viral infection,
obesity and diabetes. Notably, several types of cancer, including T
cell lymphoma, multiple myeloma, prostate and breast cancer have a
propensity to metastasize to bone. Bone metastasis occurs in 70–80%
of patients with advanced breast cancer (4–7),
leading to severe pathological bone fractures, intractable pain,
hypercalcemia and spinal cord and nerve-compression syndromes
(6,8), that add significantly to patient
morbidity and increase mortality.
Tumor invasion into bone tissues often promotes
osteoclastic bone resorption, resulting in the liberation of growth
factors from the bone matrix, which feed back on cancer cells to
enhance tumor growth resulting in the so-called 'vicious cycle of
bone metastasis' (7,8). Breast cancer cells promote the
formation of osteoclasts through secretion of osteoporotic
cytokines, such as parathyroid hormone-related peptide (PTH-rP),
prostaglandin E2 (PGE2), tumor necrosis
factor-α (TNF-α), interleukins (IL-1, IL-6, IL-8, IL-11, IL-15 and
IL-17) and leukemia inhibitory factor (LIF) (7,9). These
factors ultimately converge on RANKL pathway to drive osteoclast
formation and activity leading to bone resorption and osteolytic
lesions.
Constitutively activated nuclear factor-κB (NF-κB)
in breast cancer cells has been shown to play a crucial role in the
osteolytic mechanism of bone metastasis of breast cancer by
stimulating production of granulocyte macrophage-colony stimulating
factor (GM-CSF) in breast cancer cells that enhance osteoclast
development from monocytes (10).
Progesterone receptor-positive mammary epithelial cancer cells
express receptor activator of NF-κB ligand (RANKL) that mediates
epithelial proliferation and carcinogenesis (11). Differentiation and activation of
osteoclasts are stimulated by production of RANKL, which is
stimulated by several osteoclastogenic cytokines including PTH-rP,
TNF-α and interleukins in osteoblasts (12), and other cells. Furthermore, breast
cancer causes an increase in osteoblast apoptosis and a decrease in
osteoblastic products required for new bone formation (9). Thus, breast cancer bone
metastasis-induced bone loss is due to both activated osteoclastic
bone resorption and suppressed osteoblastic bone formation
(13).
Therapeutic strategies to break the vicious cycle
have centered on bisphosphonates or anti-RANKL antibody (denosumab)
as the current standard of care for patients with bone metastasis
(13). Bisphosphonates inhibit
osteoclastic bone resorption by targeting active osteoclasts while
denosumab diminishes the differentiation, maturation and activity
of osteoclasts by inhibiting the association of RANKL with its
receptor RANK on preosteoclasts and mature osteoclasts. These
agents are only partly effective in alleviating breast
cancer-induced skeletal damage and may themselves be associated
with significant side-effects including further suppression of bone
formation, osteonecrosis of the jaw and atypical femoral
fractures.
Gentian violet (GV), a triaminophenylmethane dye,
has been used extensively in medicine for over a century owing to
its potent anti-microbial action (14). Recent studies further suggest
angiogenic and anticancer properties of GV and there is now renewed
interest in the medical applications of this dye (15,16).
In a recent study, we found that GV exhibits anti-NF-κB activity
and similar to other NF-κB antagonists can potently augment
osteoblast differentiation and mineralization, yet potently
suppresses osteoclast differentiation (17).
In the present study, we utilized the MDA-MB-231
breast cancer cell model that has previously been shown by us to
stimulate osteoclast formation and suppress osteoblast
differentiation in vitro (18) to investigate the hypothesis that GV
may break the vicious cycle between cancer cells and bone turnover,
by diminishing cancer-induced osteoclastogenesis and/or by
alleviating the suppressive action of cancer cells on
osteoblastogenesis. Our data revealed that GV does indeed prevent
the suppressed osteoblastogenesis and stimulated osteoclastogenesis
caused by coculture with MDA-MB-231 cells. Notably, GV was also
found to directly suppress MDA-MB-231 proliferation by a mechanism
involving apoptotic cell death. These data provide a proof of
concept for future in vivo studies to examine the potential
of GV to suppress breast cancer growth in vivo and to
alleviate skeletal impairments due to cancer cells.
Materials and methods
Materials
Dulbecco's modified Eagle's medium (DMEM) with 4.5
g/l glucose, L-glutamine and sodium pyruvate and antibiotics
(penicillin and streptomycin) were purchased from Corning Cellgro
(Mediatech, Inc. Manassas, VA, USA). α-minimum essential medium
(α-MEM) was purchased from Sigma-Aldrich (St. Louis, MO, USA).
Fetal bovine serum (FBS) was purchased from HyClone (Logan, UT,
USA). Alizarin red, lipopolysacharide (LPS), caspase-3 inhibitor
and all other reagents were purchased from Sigma-Aldrich unless
otherwise specified.
Cancer cells
The human breast cancer cell line MDA-MB-231 and
human pancreatic cancer cell line MIA PaCa-2 were obtained from the
American Type Culture Collection (Manassas, VA, USA). MDA-MB-231
cells were derived from a pleural effusion of mammary gland/breast
metastases and lacked estrogen, progesterone and human epithelial
growth factor type 2 (HER2) receptors and were therefore considered
as triple-negative (19).
Proliferation of the cancer cells
Breast cancer MDA-MB-231 or MIA PaCa-2 cells
(1×105/ml/well) were cultured in 24-well plates in DMEM
containing 10% FBS and 1% P/S in the presence or absence of GV (1,
10, 50, 100 or 200 nM) for 1, 3, 7 or 14 days in a water-saturated
atmosphere containing 5% CO2 and 95% air at 37°C. After
culture, the cells were detached from culture dishes and
quantitated as previously described (20,21).
In separate experiments, MDA-MB-231 cells
(1×105 cells/ml/well) were cultured using a 24-well
plate in DMEM containing 10% FBS and 1% P/S in the absence of GV
for 7 days until reaching confluency, and then the cells were
cultured in the presence of GV (1, 10, 50, 100 or 200 nM) with or
without caspase-3 inhibitor (5 µM) for 3 days (22). After culturing, the cells were
detached from each culture dish and counted.
Cell counting
After trypsinization of each culture dish using 0.2%
Trypsin plus 0.02% EDTA in Ca2+/Mg2+-free
phosphate-buffered saline (PBS) for 2 min at 37°C, detached cells
were collected after centrifugation. The cells were resuspended in
PBS and stained with eosin. Cell numbers were counted under a
microscope using a hemocytometer plate. For each dish, we took the
average of the two countings. The cell number was presented as cell
number/well.
Animals and bone marrow cells
Female mice (CD1-Elite, wild-type, 2-months old),
which were purchased from Charles River, were housed in a
pathogen-free facility, and all procedures and protocols were
approved by the Institutional Animal Care and Use Committee at
Emory University. The femur and tibia tissues were removed
immediately after sacrifice. Bone marrow cells were isolated with a
procedure of sterilization from the femoral and tibial tissues.
Osteoclastogenesis in bone marrow cell
culture
To determine the effects of breast cancer cells on
osteoclastogenesis in whole bone marrow, bone marrow cells
(2×105 cells/ml/well) were cultured in DMEM containing
10% FBS and 1% P/S in 24-well plates (1.0 ml/well) (18). The cells were cultured with or
without LPS (10 µg/ml of medium) for 3 days in the presence
or absence of GV (10, 50, 100 and 200 nM). The cells were fed by
replacing 50% (0.5 ml) of old medium with fresh medium containing
fresh LPS (10 µg/ml of medium) and GV (10, 50, 100 and 200
nM) as appropriate. Cultures were maintained for an additional 4
days (18). After 7 days of
culture, the cells were washed with PBS and fixed with 10%
neutralized formalin-phosphate (pH 7.2) for 10 min. The plates were
stained with tartrate-resistant acid phosphatase (TRACP), a
specific marker of osteoclasts (23) by incubating for 90 min at room
temperature in acetate buffer (pH 5.0) containing naphthol AS-MX
phosphate (Sigma) as a stain for the reaction product, in the
presence of 10 mM sodium tartrate. TRACP-positive multinucleated
cells (MNCs) containing 3 or more nuclei were counted as
osteoclast-like cells. MNCs scored were the mean ± SD of 6
cultures.
Osteoblast mineralization in cocultures
of bone marrow and breast cancer cells
To determine the effects of breast cancer cells on
bone marrow osteoblastogenesis and mineralization, we used
mineralization medium (MM) containing ascorbic acid (100 ng/ml) and
4 mM β-glycerophosphate in DMEM with 10% FBS and 1% P/S using
12-well plates. Bone marrow cells (1×106 cells/ml/well)
were cultured for 3 days at 37°C in a humidified 5% CO2
atmosphere, and then cocultured with breast cancer MDA-MB-231 cells
(1×104 cells/well) in the presence or absence of α-MEM
and MM with either vehicle or GV (1, 10 or 100 nM) for 18 days
(18). Medium was changed every 3
days. After culturing, the cells were washed with PBS and stained
with Alizarin red S stain. For quantification, 10% cetylpyridinium
chloride solution was added to each well to elute the dye (18). After complete elution, the
absorbance was measured at 570 nm on a microtiter plate reader.
Coculture of preosteoblastic MC3T3 cells
with breast cancer cells
Preosteoblastic MC3T3 cells (2×105
cells/1 ml/well) were cultured using a 12-well plare in α-MEM
containing 10% FBS and 1% P/S, and 3 days later the culture medium
was replaced with DMEM (containing 10% FBS and 1% P/S) in the
presence or absence of MM containing ascorbic acid (100 ng/ml) and
4 mM β-glycerophosphate. After 3 days, the osteoblastic cells were
cocultured with the addition of breast cancer MDA-MB-231 bone
metastatic cells (1×103 or 1×104
cells/ml/well) in α-MEM containing MM in the presence or absence of
GV (1, 10 and 100 nM) for 18 days (18). Medium was changed every 3 days.
After culturing, the cells were washed with PBS and stained with
Alizarin red S stain. For quantification of calcium deposition,
after washing of cells to remove unbound dye, Alizarin red S was
eluted with 10% cetylpyridinium chloride and the absorbance was
read at 570 nm on a microtiter plate reader.
Statistical analysis
Statistical significance was determined using
GraphPad InStat version 3 for Windows XP (GraphPad Software, Inc.,
La Jolla, CA, USA). Multiple comparisons were performed by one-way
analysis of variance (ANOVA) with Tukey-Kramer multiple comparisons
post-test for parametric data as indicated. p<0.05 was
considered to indicate a statistically significant result.
Results
GV suppresses proliferation of MDA-MB-231
and MIA PaCa-2 cells
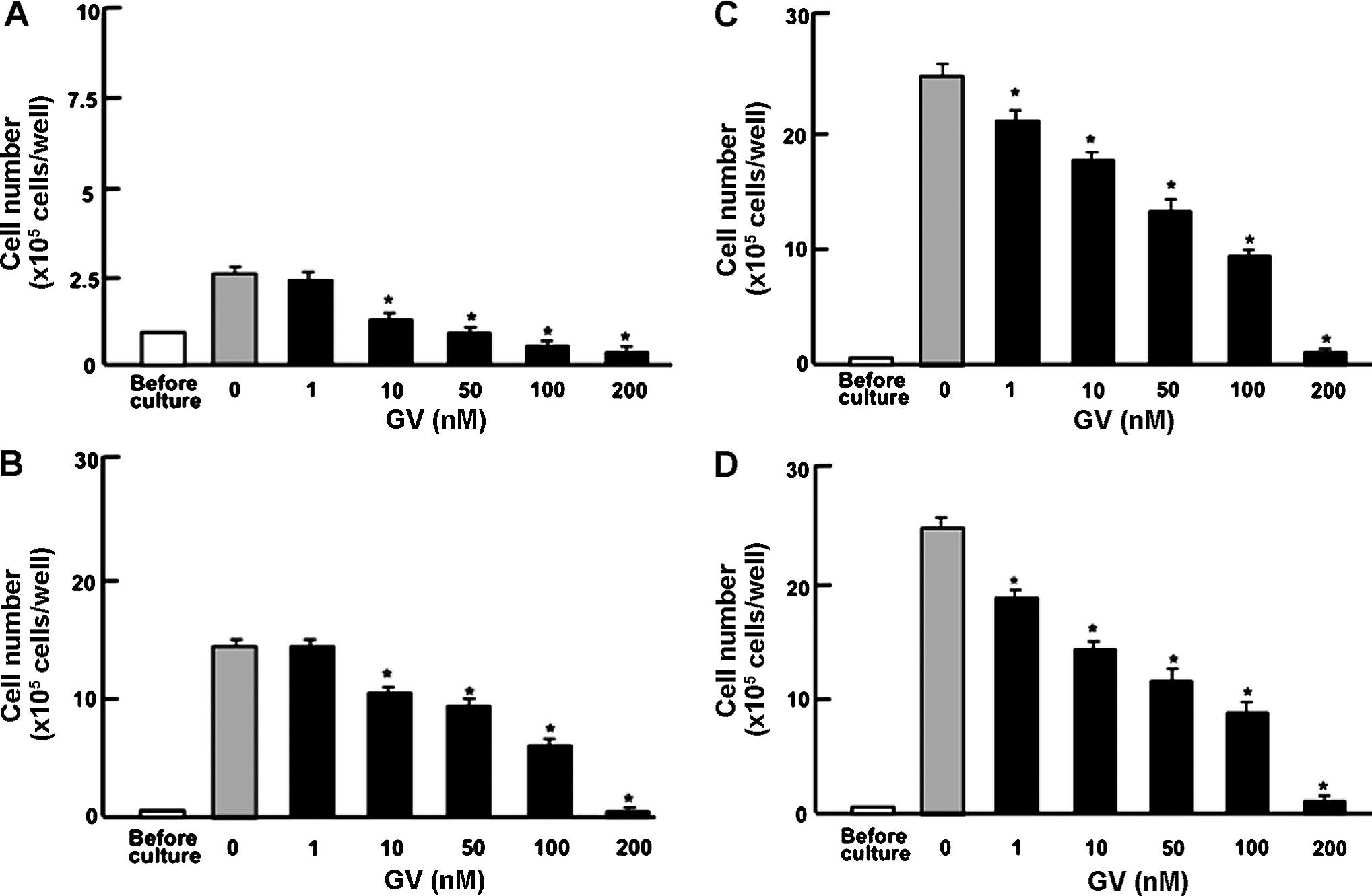
To determine the effects of GV on the proliferation
of human breast cancer cells, MDA-MB-231 cells were cultured in the
presence of GV for 1–14 days. Addition of GV (1–200 nM) led to a
significant dose dependent suppression of cell proliferation by day
1 (Fig. 1A), day 3 (Fig. 1B), day 7 (Fig. 1C) and day 14 (Fig. 1D). To investigate whether these
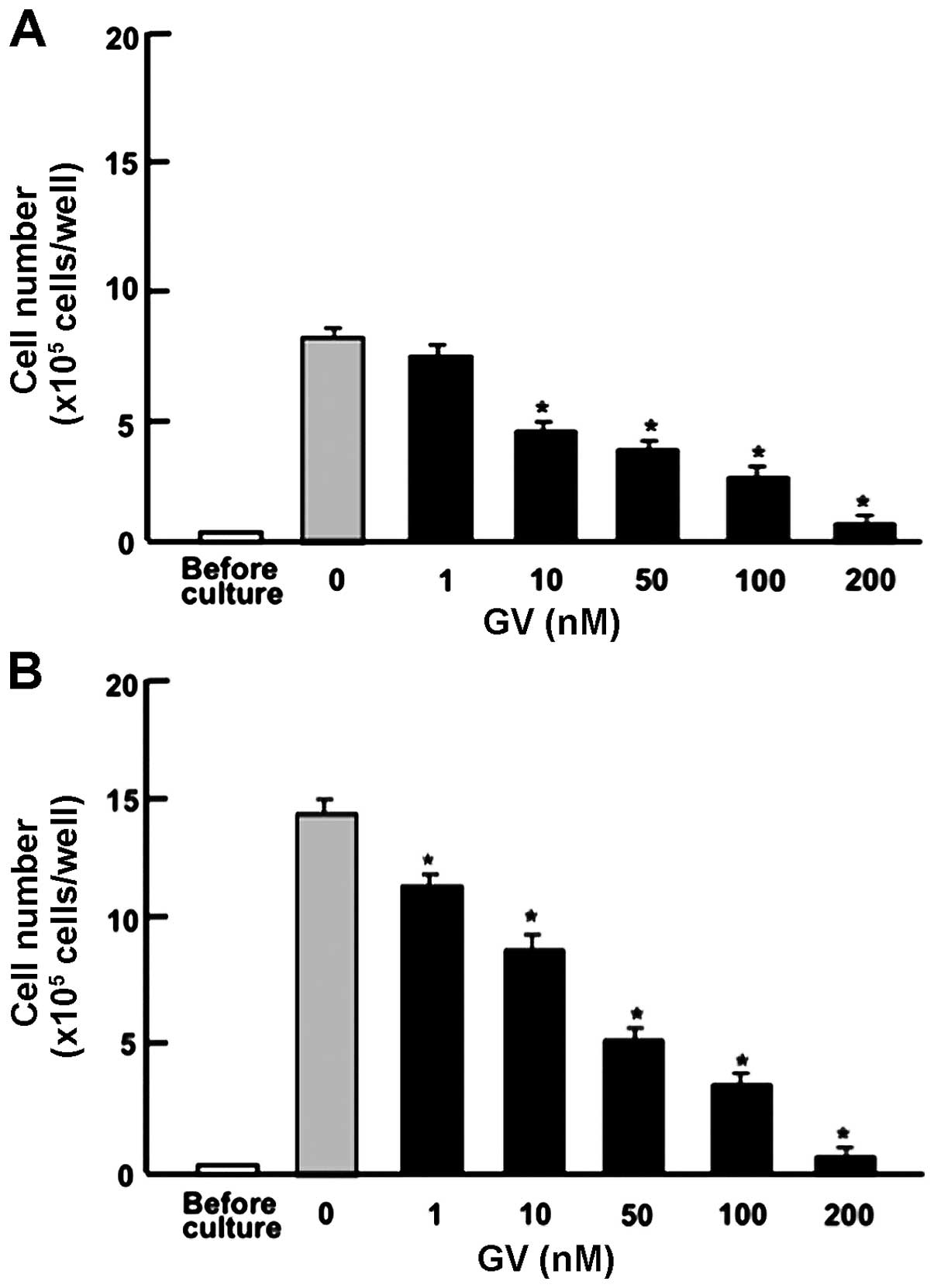
effects are unique to breast cancer cells, we further examined the
effects of GV on the cell proliferation of MIA PaCa-2 human
pancreatic cancer cells. As with MDA-MB-231 cells, proliferation
was suppressed by GV (1–200 nM) at day 3 (Fig. 2A) and day 7 (Fig. 2B). Taken together, these data reveal
for the first time the antiproliferative effects of GV on human
breast and pancreatic cancer cells in vitro.
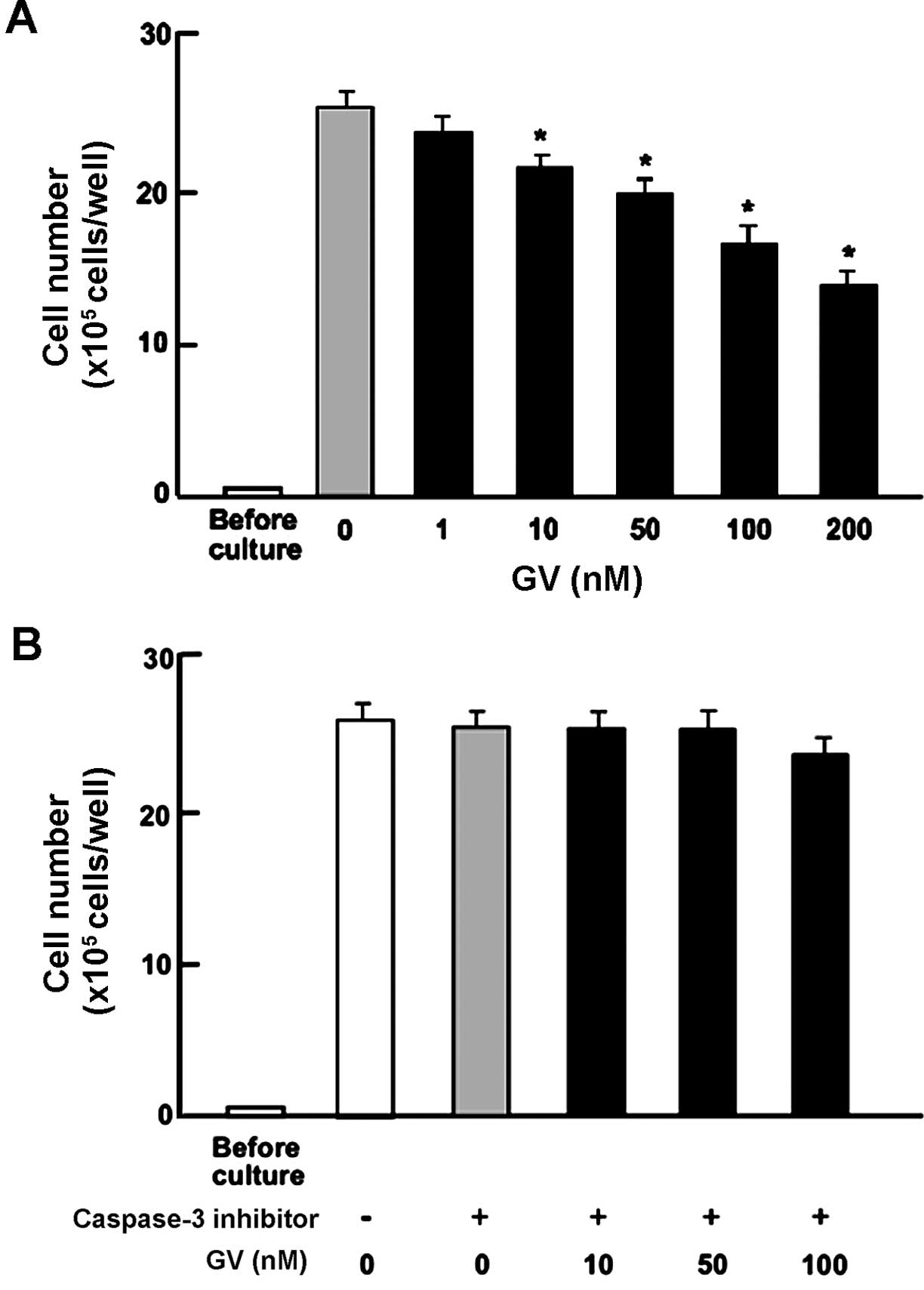
GV stimulates apoptotic cell death of
MDA-MB-231 breast cancer cells
We previously reported that GV promotes osteoblast
differentiation at doses of up to 100 nM in vitro. Even
though GV suppressed osteoclastic differentiation of the monocytic
(osteoclast precursor) cell line RAW264.7, GV did not promote
apoptosis at the same bioactive doses (10–100 nM) (17). To investigate whether the
suppression of proliferation by GV on MDA-MB-231 cells involves
apoptosis or another mechanism, confluent MDA-MB-231 cells were
treated with GV (10, 50 and 100 nM) with or without caspase-3
inhibitor (5 µM) for an additional 3 days (Fig. 3A and B). The data revealed that in
the presence of the caspase-3 inhibitor, MDA-MB-231 cells were
completely protected from the inhibitory effects of GV. These data
demonstrated that unlike the effect of GV on osteoblast and
osteoclast precursor cell lines, MDA-MB-231 breast cancer cells did
undergo apoptosis in the presence of low-dose GV.
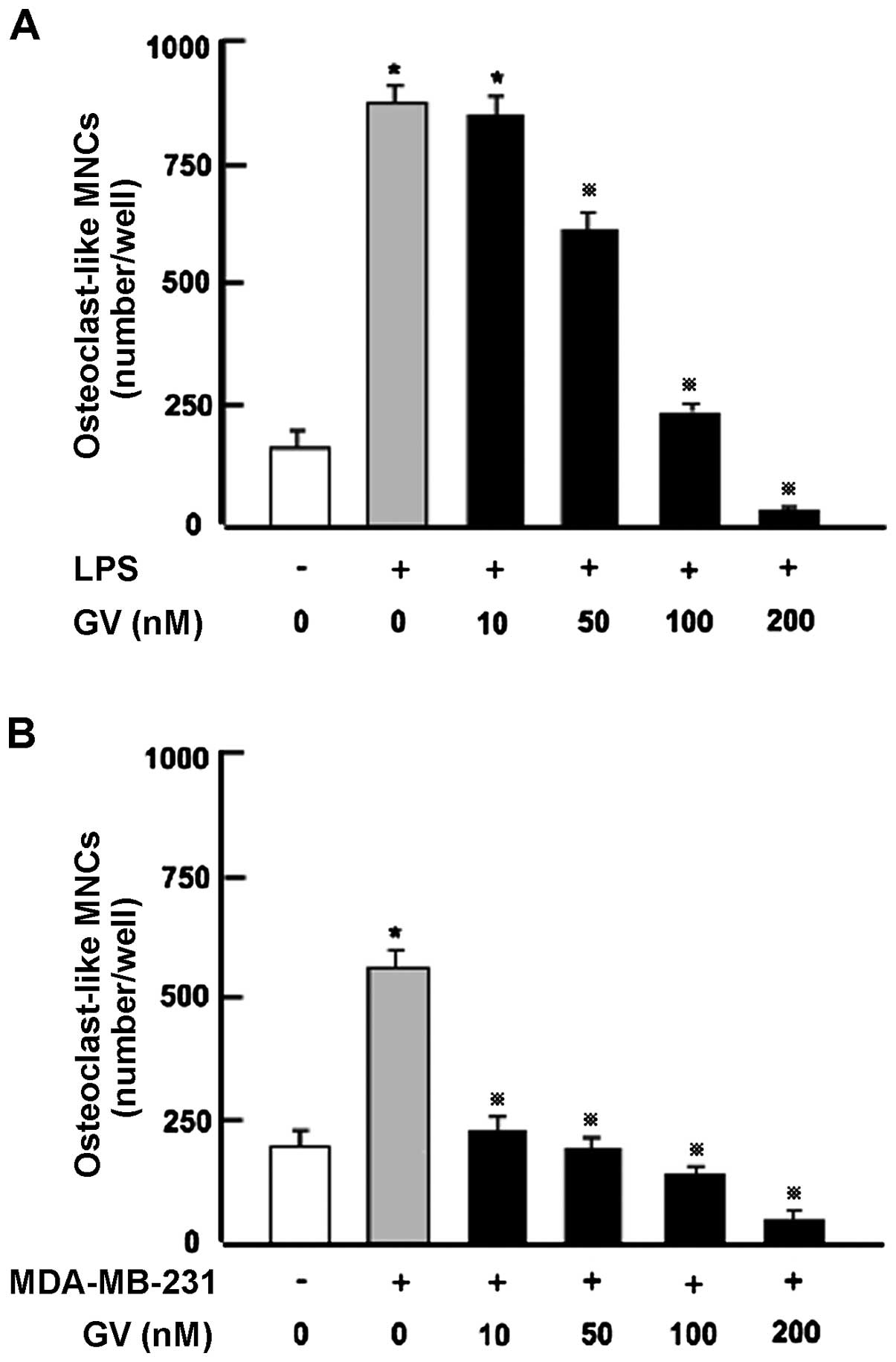
GV inhibits the osteoclast formation in
bone marrow cultures in vitro
We previously reported that GV blocks the
RANKL-induced differentiation of RAW264.7 cells into mature
osteoclasts (17). Although RANKL
is a key osteoclastic cytokine, activation of Toll-like receptors
on osteoblasts and monocytes by the bacterial-derived protein
lipopolysaccharide (LPS), leads to production of inflammatory
cytokines such as IL-1 and TNF-α from monocytes and by RANKL
production from osteoblasts causing significant osteoclastogenesis
(13). To test the ability of GV to
moderate osteoclastogenesis in a more physiological setting we used
a primary bone marrow culture system in which whole bone marrow was
treated with LPS in vitro with or without GV (50–200 nM) for
7 days (Fig. 4A). LPS caused a
marked increase in osteoclastogenesis in the bone marrow cells
in vitro. This increase was dose-dependently prevented by
addition of GV (10–200 nM). These data confirm the capacity of GV
to suppress the differentiation of osteoclasts in bone marrow and
further show for the first time that GV blocks osteoclastogenesis
in a primary bone marrow culture system.
GV alleviates the pro-osteoclastogenic
activity of MDA-MB-231 cells in vitro
Since GV blocks both osteoclastogenesis and
MDA-MB-231 breast cancer cell proliferation, we next investigated
whether GV alleviates the osteoclast formation promoted by human
breast cancer. To this end, MDA-MB-231 cells were cocultured with
mouse bone marrow cells in vitro as previously described
(18). Osteoclastogenesis was
markedly enhanced by coculture with MDA-MB-231 cells (Fig. 4B) and was significantly suppressed
by addition of GV (10–200 nM).
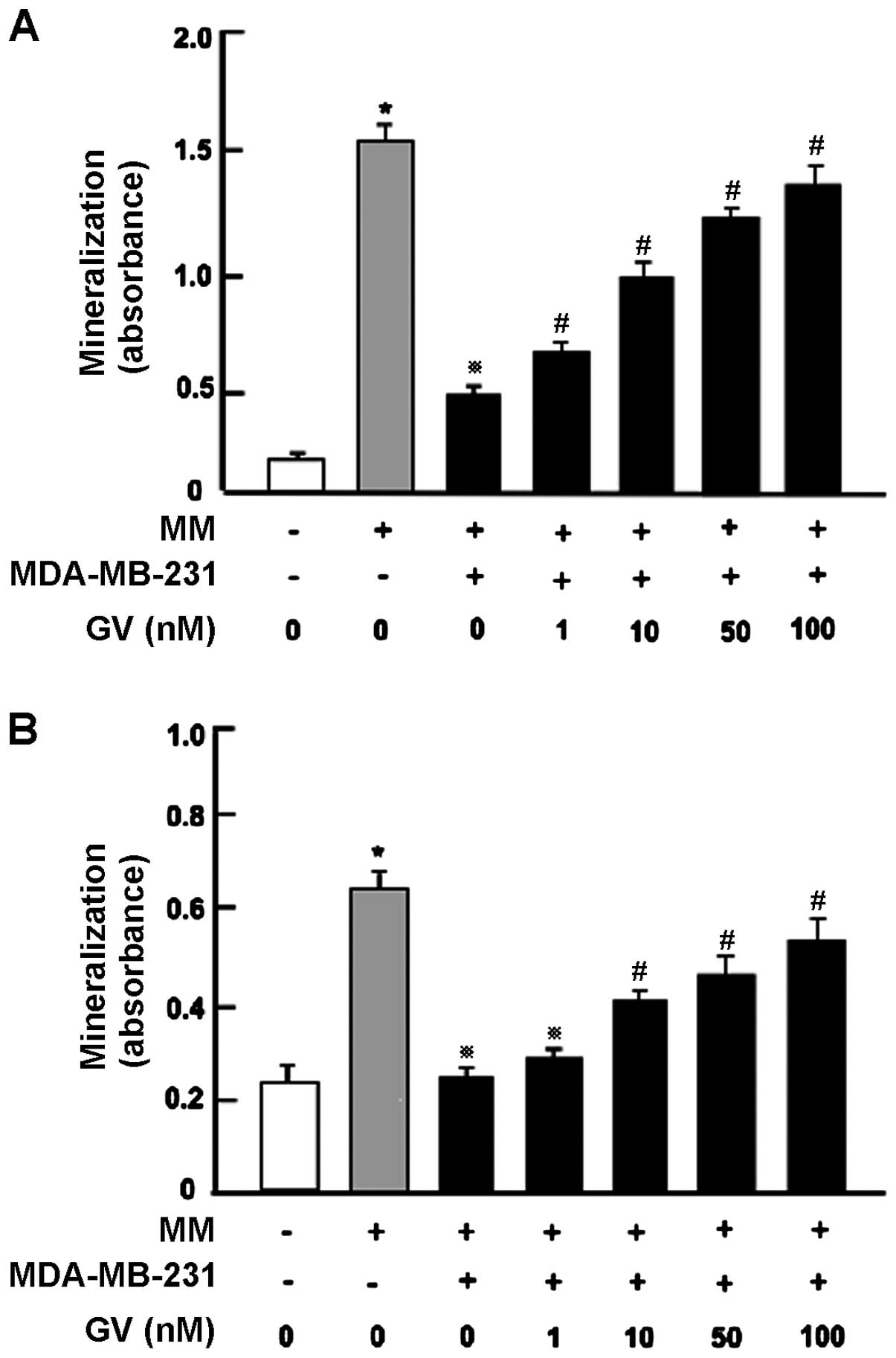
GV alleviates the anti-osteoblastogenic
activity of MDA-MB-231 cells in vitro
Next, we examined whether GV alleviates the
suppressive effect of MDA-MB-231 cells on osteoblast
differentiation and mineralization activity using the MC3T3
preosteoblastic cell line (Fig. 5A)
as well as primary bone marrow cultures (Fig. 5B). After 3 days of culture,
preosteoblastic MC3T3 cells were cocultured with MDA-MB-231 cells
in MM in the presence or absence of GV (1, 10 and 100 nM) for an
additional 18 days (Fig. 5A). We
previously reported (18), that
coculture with MDA-MB-231 cells potently suppressed mineralization
in osteoblastic cells. Importantly, this suppression was
significantly prevented by GV (1–100 nM) (Fig. 5A).
Finally, we cocultured primary bone marrow cells
containing osteoblast precursors (Fig.
5B) with MDA-MB-231 cells in vitro. Mouse bone marrow
cells were cultured in the presence or absence of MM. After 2 days,
the cells were cocultured with the addition of MDA-MB-231 cells in
the presence or absence of GV (1, 10 or 100 nM) for 18 days which
led to mineralization. Mineralization in the bone marrow cells was
potently suppressed by coculture with the MDA-MB-231 cells. This
suppression was prevented by GV (1–100 nM) (Fig. 5B).
Discussion
The central finding of the present study is that GV
mediates a suppressive effect on proliferation, and a stimulatory
effect on apoptotic cell death, in human breast cancer MDA-MB-231
cells in vitro. Moreover, both the suppressed
osteoblastogenesis and enhanced osteoclastogenesis induced by
coculture with MDA-MB-231 cells were prevented in the presence of
GV in vitro. Thus, GV has significant potential to alleviate
the skeletal abnormalities and metastatic engraftment of human
breast cancer cells in vivo.
We previously reported that GV stimulates
osteoblastic mineralization and suppresses RANKL-induced
osteoclastogenesis using preosteoblastic MC3T3 cells and RAW267.4
osteoclast precursors in vitro (17). Due to the limitations of the cell
lines we further demonstrated the capacity of GV to mediate these
responses in the context of whole bone marrow systems ex
vivo. These models represent a more physiological response as
would occur in vivo in the context of whole bone marrow.
In vivo, RANKL is supplied by bone marrow cells such as
osteoblasts. Consequently, rather than providing exogenous RANKL as
in our prior studies we stimulated RANKL production from bone
marrow cells using LPS. As previously reported for the cell lines,
GV potently suppressed osteoclastogenesis and enhanced osteoblast
mineralization in whole bone marrow. These studies provide a proof
of concept for future studies of GV action in intact animals in
vivo.
We previously reported that these differential
actions of GV on osteoclast and osteoblasts stem from the capacity
of GV to suppress activation of the NF-κB signal transduction
pathway that is required for osteoclastogenesis, yet inhibitory to
osteoblast differentiation and activity (17). We speculated that GV is an inhibitor
of NF-κB activation and holds promise for modulation of bone
turnover to promote a balance between bone formation and bone
resorption, favorable to the gain of bone mass. In the context of
breast cancer, one action of GV may thus be to restore bone
turnover, deregulated by cancer cells. Breast cancer cells are
known to produce RANKL, which plays a pivotal role in formation
from preosteoclastic cells to mature osteoclasts (7–13).
Indeed the capacity of MDA-MB-231 cells to promote
osteoclastogenesis in our bone marrow culture system may be due to
RANKL production.
As reported for in vivo studies of breast
cancer cells in mice (18,24) and in vitro using MC3T3 cell
lines (17), osteoblastic
mineralization in mouse bone marrow cells was markedly suppressed
after coculture with breast cancer MDA-MB-231 cells in
vitro. The mechanisms driving osteoblast suppression are not
presently clear, yet could be related to production of TNF-α, a
cytokine that is known to be produced by breast cancer cells
(10,11). TNF-α suppresses osteoblastic
mineralization by activation of NF-κB signaling (17,25).
Owing to the anti-NF-κB activity of GV previously reported by us
(17), we speculate that the
capacity of GV to relieve the inhibitory effects of MDA-MB-231
breast cancer cells on osteoblastic mineralization relates to
suppression of TNF-α-induced NF-κB signaling in osteoblasts, the
direct stimulation of osteoblastic mineralization, the induction of
apoptosis in MDA-MB-231 cells or a combination of these mechanisms.
Whether the actions of GV were mediated through actions on bone
cells or on cancer cells, or on both, remains to be determined.
Nonetheless, irrespective of the site of action, the data suggest
that GV is a useful agent to break the vicious cycle, diminish
growth and metastases of breast cancer cells, and repair
preexisting damage to the skeleton caused by bone metastases. GV
restores a favorable balance between bone formation and resorption
to negate the damaging effects of cancer metastases. Future studies
in vivo are needed to establish the therapeutic potential of
GV.
In conclusion, the present study demonstrated that
GV suppressed proliferation and stimulated apoptotic cell death in
human breast cancer MDA-MB-231 cells in vitro, and that it
alleviated the suppression of osteoblastogenesis and stimulated the
osteoclastogenesis induced by coculture with MDA-MB-231 cells in
vitro. Thus, GV may be a new and useful therapeutic tool for
breast cancer bone metastasis.
References
|
1
|
Raggatt LJ and Partridge NC: Cellular and
molecular mechanisms of bone remodeling. J Biol Chem.
285:25103–25108. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Weitzmann MN and Pacifici R: Estrogen
deficiency and bone loss: An inflammatory tale. J Clin Invest.
116:1186–1194. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Johnell O and Kanis JA: An estimate of the
worldwide prevalence and disability associated with osteoporotic
fractures. Osteoporos Int. 17:1726–1733. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Boyce BF, Yoneda T and Guise TA: Factors
regulating the growth of metastatic cancer in bone. Endocr Relat
Cancer. 6:333–347. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mundy GR: Metastasis to bone: Causes,
consequences and therapeutic opportunities. Nat Rev Cancer.
2:584–593. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Roodman GD: Mechanism of bone metastasis.
N Engl J Med. 350:1655–1664. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Akhtari M, Mansuri J, Newman KA, Guise TM
and Seth P: Biology of breast cancer bone metastasis. Cancer Biol
Ther. 7:3–9. 2008. View Article : Google Scholar
|
|
8
|
Coleman RE: Metastatic bone disease:
Clinical features, pathophysiology and treatment strategies. Cancer
Treat Rev. 27:165–176. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen YC, Sosnoski DM and Mastro AM: Breast
cancer metastasis to the bone: Mechanisms of bone loss. Breast
Cancer Res. 12:2152010. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Park BK, Zhang H, Zeng Q, Dai J, Keller
ET, Giordano T, Gu K, Shah V, Pei L, Zarbo RJ, McCauley L, Shi S,
Chen S and Wang CY: NF-kappaB in breast cancer cells promotes
osteolytic bone metastasis by inducing osteoclastogenesis via
GM-CSF. Nat Med. 13:62–69. 2007. View
Article : Google Scholar
|
|
11
|
Gonzalez-Suarez E, Jacob AP, Jones J,
Miller R, Roudier-Meyer MP, Erwert R, Pinkas J, Branstetter D and
Dougall WC: RANK ligand mediates progestin-induced mammary
epithelial proliferation and carcinogenesis. Nature. 468:103–107.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zaidi M, Blair HC, Moonga BS, Abe E and
Huang CL: Osteoclastogenesis, bone resorption, and osteoclast-based
therapeutics. J Bone Miner Res. 18:599–609. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Weilbaecher KN, Guise TA and McCauley LK:
Cancer to bone: a fatal attraction. Nat Rev Cancer. 11:411–425.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Berrios RL and Arbiser JL: Effectiveness
of gentian violet and similar products commonly used to treat
pyodermas. Dermatol Clin. 2011:69–73. 2011. View Article : Google Scholar
|
|
15
|
Perry BN, Govindarajan B, Bhandarkar SS,
Knaus UG, Valo M, Sturk C, Carrillo CO, Sohn A, Cerimele F, Dumont
D, et al: Pharmacologic blockade of angiopoietin-2 is efficacious
against model hemangiomas in mice. J Invest Dermatol.
126:2316–2322. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang X, Zheng Y, Fried LE, Du Y, Montano
SJ, Sohn A, Lefkove B, Holmgren L, Arbiser JL, Holmgren A, et al:
Disruption of the mitochondrial thioredoxin system as a cell death
mechanism of cationic triphenylmethanes. Free Radic Biol Med.
50:811–820. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yamaguchi M, Vikulina T, Arbiser JL and
Weitzmann MN: Suppression of NF-κB activation by gentian violet
promotes osteoblastogenesis and suppresses osteoclastogenesis. Curr
Mol Med. 14:783–792. 2014. View Article : Google Scholar
|
|
18
|
Yamaguchi M, Zhu S, Weitzmann MN, Snyder
JP and Shoji M: Curcumin analog UBS109 prevents bone marrow
osteoblastogenesis and osteoclastogenesis disordered by coculture
with breast cancer MDA-MB-231 bone metastatic cells in vitro. Mol
Cell Biochem. 401:1–10. 2015. View Article : Google Scholar
|
|
19
|
Yoneda T, Williams PJ, Hiraga T, Niewolna
M and Nishimura R: A bone-seeking clone exhibits different
biological properties from the MDA-MB-231 parental human breast
cancer cells and a brain-seeking clone in vivo and in vitro. J Bone
Miner Res. 16:1486–1495. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Misawa H, Inagaki S and Yamaguchi M:
Suppression of cell proliferation and deoxyribonucleic acid
synthesis in the cloned rat hepatoma H4-II-E cells overexpressing
regucalcin. J Cell Biochem. 84:143–149. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yamaguchi M and Daimon Y: Overexpression
of regucalcin suppresses cell proliferation in cloned rat hepatoma
H4-II-E cells: involvement of intracellular signaling factors and
cell cycle-related genes. J Cell Biochem. 95:1169–1177. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Izumi T and Yamaguchi M: Overexpression of
regucalcin suppresses cell death in cloned rat hepatoma H4-II-E
cells induced by tumor necrosis factor-alpha or thapsigargin. J
Cell Biochem. 92:296–306. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Minkin C: Bone acid phosphatase:
Tartrate-resistant acid phosphatase as a marker osteoclast
function. Calcif Tissue Int. 34:285–290. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yamaguchi M, Zhu S, Zhang S, Wu D, Moore
TM, Snyder JP and Shoji M: Curcumin analogue UBS109 prevents bone
loss in breast cancer bone metastasis mouse model: Involvement in
osteoblastogenesis and osteoclastogenesis. Cell Tissue Res.
357:245–252. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li Y, Li A, Strait K, Zhang H, Nanes MS
and Weitzmann MN: Endogenous TNFalpha lowers maximum peak bone mass
and inhibits osteoblastic Smad activation through NF-kappaB. J Bone
Miner Res. 22:646–655. 2007. View Article : Google Scholar : PubMed/NCBI
|