Introduction
Oral squamous cell carcinoma (OSCC) is one of the
life-threatening cancer types worldwide with a high frequency of
recurrence and a low survival rate (1,2).
Although many efforts have been made to improve the therapeutic
methods for the treatment of OSCC, the therapeutic efficacy and the
five-year survival rate remain unsatisfactory (3). Therefore, a better understanding of
the molecular pathologies underlying OSCC provides novel insight to
develop effective therapeutic strategies for OSCC.
In recent years, microRNAs (miRNAs), a small type of
non-coding RNAs, have been found to play an important role in many
diseases including cancer (4,5).
miRNAs are capable of targeting the 3′-untranslated region (3′-UTR)
of messenger RNA (mRNA) containing the corresponding complementary
sequences that results in mRNA destabilization and degradation,
thereby leading to protein translational inhibition (6). Given their ability to modulate gene
expression, miRNAs have been found to participate in many cell
processes, including cell growth and proliferation,
differentiation, apoptosis; thus, miRNAs are involved in the
pathologies of various diseases (7,8).
Numerous miRNAs have been identified in various studies on cancer
including OSCC (9). For instance,
Manikandan et al have reported that miR-21,
miR-125b-2*, miR-134, miR-155, miR-184 and miR-205 are
dysregulated in OSCC and are associated with clinical pathology
(10). A retrospective study of
miRNA expression in OSCC tumor tissues from early stage patients
has demonstrated that miR-375 and miR-214-3p have a predictive
value for prognosis (11).
Downregulation of miR-329 and miR-410 increased the proliferation
and invasion of OSCC cells by targeting and regulating the
expression of Wnt-7b (12).
Overexpression of mi-125a has been found to inhibit the
proliferation and invasion of OSCC cells by modulating
estrogen-related receptor α (13).
Therefore, targeting the pivotal miRNAs in the pathogenesis of OSCC
is expected to provide valid therapeutics for the treatment of
OSCC.
MicroRNA-138 (miR-138) has been suggested as a
tumor-suppressor gene that is frequently downregulated in various
types of cancer, including lung (14–16),
thyroid (17), liver (18), colorectal (19) and ovarian (20) cancer. Moreover, miR-138 is involved
in OSCC. Loss of heterozygosity in miR-138 precursor genes located
at the chromosome loci frequently occurs in OSCC (21,22). A
decreased expression of miR-138 was observed in the tumor samples
from OSCC (10). However, the
precise molecular mechanism of miR-138 in regulating OSCC remains
poorly understood. Yes-associated protein 1 (YAP1), a critical
transcription co-activator of Hippo pathway, has been suggested as
a candidate oncogene in human cancer (23–25).
Overexpression of YAP1 has been observed in a number of cancer
types and is associated with cell proliferation and tumor growth
(26,27). In OSCC, YAP1 has been found to be
amplified and overexpressed (28),
suggesting an important role of YAP1 in OSCC. Of note, we found
through bioinformatics analysis that YAP1 was the predicted target
gene of miR-138. Therefore, the aim of the present study was to
identify and validate whether miR-138 was involved in the
development and progression of OSCC by directly targeting and
regulating YAP1.
Materials and methods
Cell lines and animals
OSCC cell lines, including OC3, KB, OEC-M1, HSC3 and
SCC-4, were purchased from the Type Culture Collection of the
Chinese Academy of Sciences (Shanghai, China) and were cultured
according to the manufacturer's instructions. Normal human oral
keratinocytes (HOK) were obtained from ScienCell (Carlsbad, CA,
USA) and were grown in oral keratinocyte medium according to the
manufacturer's instructions. The cells were maintained in a
humidified atmosphere containing 5% CO2 at 37°C.
Five-week-old male BALB/c nude mice (weighing 25–30 g) were
purchased from the Laboratory Animal Centre of the Medical School
of Xi'an Jiaotong University (Xi'an, China) and were raised and
handled according to the guideline of the Institutional Animal Care
and Use Committee of Xi'an Jiaotong University.
Collection of clinical samples
Twenty paired adjacent non-tumorous and tumor
tissues from patients undergoing surgical resection were frozen in
liquid nitrogen. The present study was approved by and was in
accordance with the guidelines of Institutional Human Experiment
and the Ethics Committee of the First Affiliated Hospital, Medical
School of Xi'an Jiaotong University (Xi'an, China).
Reverse transcriptase-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted using TRIzol (Invitrogen,
Carlsbad, CA, USA) and was used to generate cDNA using M-MLV
reverse transcriptase (Clontech, Palo Alto, CA, USA) and One Step
PrimeScript miRNA cDNA Synthesis kit (Takara, Dalian, China) for
mRNA and miRNAs analysis, respectively, according to the
manufacturer's instructions. The gene expression was detected using
SYBR-Green qPCR Master Mix (Thermo Fisher, Shanghai, China). The
relative quantification of the gene expression level was compared
with the internal referee GAPDH (for mRNA) or U6 SnRNA (for miRNAs)
using the 2−ΔΔCt method.
Cell infection
Lentivirus (LV)-pre-miR-138, LV-anti-mR-138 and
their negative controls were purchased from GenePharma (Shanghai,
China). For cell infection, the cells were cultured in a normal
medium for 24 h and then cultured in a medium containing Polybrene
(Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA). The cells
were then infected with pre-miR-13, LV-anti-mR-138s or their
negative controls on a 0.5×105 plaque-forming unit
overnight. The old medium was discarded, and a new medium without
Polybrene was added and incubated overnight. Stable clones were
selected with puromycin dihydrochloride (Santa Cruz Biotechnology,
Inc.).
Cell proliferation assay
Cell proliferation was detected by 3-(4,5)-dimethylthiahiazol(-z-y1)-3,5-di-phenytetrazo-liumromide
(MTT) assay. Briefly, the cells infected with LV-miR-138 or
LV-anti-miR-138 were seeded in 96-well plates at 1×104
cells/well for 24, 48 and 72 h. The old medium was then replaced
with fresh medium containing MTT [20 μl/well; 5 mg/ml
diluted in phosphate-buffered saline (PBS)]. The cells were
continually cultured for another 4 h, the medium was discarded and
dimethylsulfoxide (150 μl/well; Sigma, St. Louis, MO, USA)
was added to dissolve the formed formazan crystals. The optical
density at 490 nm was then measured with a microtiter plate reader
(Thermo Electron Corporation, Vantaa, Finland).
Dual-luciferase reporter assay
The cDNA fragment of 3′-UTR of YAP1 containing the
putative binding site of miR-138 was subcloned into a pGL3
luciferase promoter vector (Promega, Madison, WI, USA). The cells
firmly expressing pre-miR-138 or anti-miR-138 were transfected with
0.1 μg of pGL3-YAP1-3′-UTR and incubated for 48 h. The cells
were collected and lysed. The luciferase activities were quantified
using the Dual-Luciferase reporter assay kit (Promega) according to
the manufacturer's instructions.
Western blot analysis
Proteins from cells or tumor tissues were extracted
using a protein extraction kit (Applygen Technologies, Beijing,
China). Protein concentrations in different samples were measured
using the Bradford method. For protein separation, a total of 50
μg of protein was loaded on 12.5% sodium dodecyl sulfate
polyacrylamide gel electrophoresis. The protein was then
transferred to a nitrocellulose membrane (Bio-Rad, Hercules, CA,
USA), which was blocked with 3.0% non-fat milk for 1 h at 37°C.
Primary antibodies were added and incubated at 4°C overnight.
Horseradish peroxidase-conjugated secondary antibodies (1:2,000;
Bioss, Beijing, China) were added and incubated for 1 h at room
temperature. After washing, the immunoreactive protein bands on the
membrane were visualized using an enhanced chemiluminescence
detection system (Amersham, Little Chalfont, UK). The primary
antibodies used in these experiments, including anti-YAP1 and
anti-GAPDH antibodies were purchased from Santa Cruz Biotechnology,
Inc.
Xenograft tumor assay
OEC-M1 cells (1×106 cells) infected with
LV-pre-miR-138 or LV-anti-miR-138 diluted in 200 μl of PBS
were inoculated subcutaneously into the left flank of nude mice.
The size of the tumor (length and width) was measured every 3 days
by a vernier caliper. The tumor volume was presented as length ×
width2 × π/6. At the end of the experiment, the tumor
was isolated, weighed and extracted for further analysis.
Statistical analysis
Data are presented as means ± standard deviation and
processed using SPSS version 11.5 (SPSS, Inc., Chicago, IL, USA).
Statistical differences were obtained by the Student's t-test or
one-way analysis of variance followed by Bonferroni post hoc test.
Differences were considered statistically significant at
P<0.05.
Results
Downregulated expression of miR-138 is in
OSCC tissues and cell lines
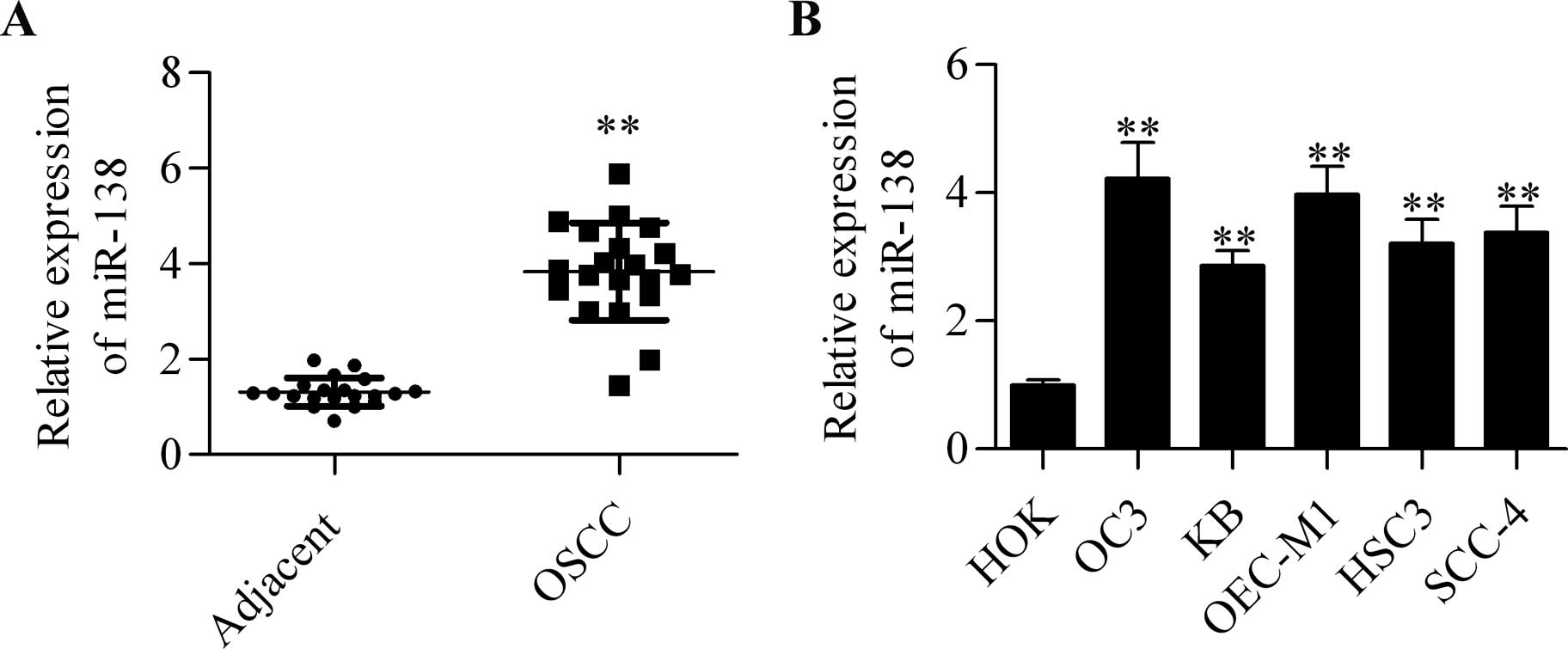
To examine the role of miR-138 in OSCC, the
expression profile of miR-138 in tumor and adjacent non-tumorous
epithelia was quantified by RT-qPCR analysis. Generally, miR-138
was significantly downregulated in OSCC tumor tissues as compared
with that in the adjacent non-tumorous tissues (Fig. 1A). In addition, the expression of
miR-138 in several different OSCC cell lines was detected.
Furthermore, the results showed that miR-138 expression was
markedly reduced to different degrees in OC3, KB, OEC-M1, HSC3 and
SCC-4 cell lines than that in the control HOK cells (Fig. 1B).
miR-138 suppresses the proliferation of
OSCC cells
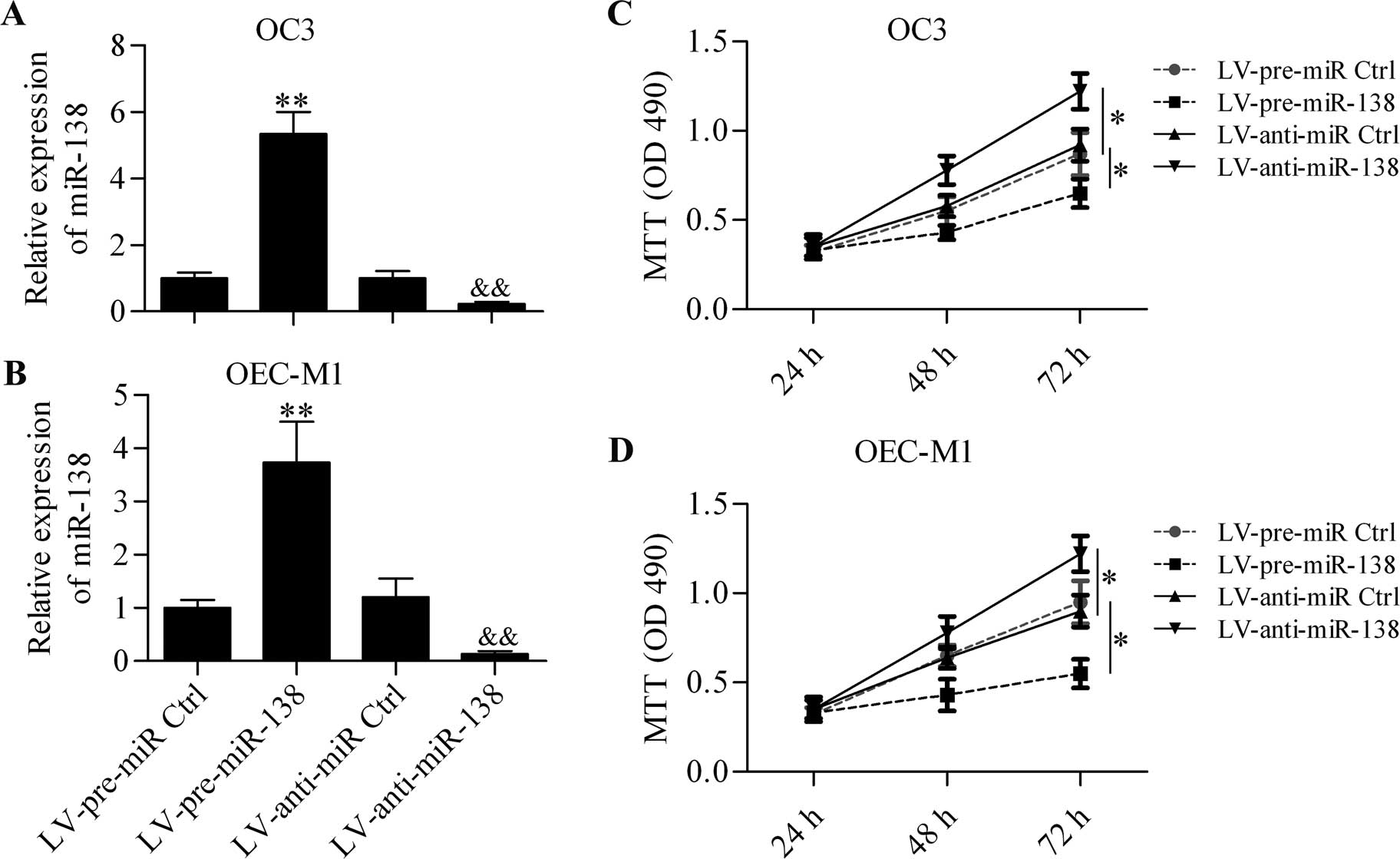
Considering the downregulated expression of miR-138
in OSCC, we investigated the function of miR-138 in OSCC cells.
OSCC cells with miR-138 overexpression or downregulation were
generated in OC3 (Fig. 2A) and
OEC-M1 (Fig. 2B) cells by infecting
with LV-pre-miR-138 or LV-anti-miR-138. MTT assay results showed
that miR-138 overexpression significantly decreased the
proliferation in OC3 cells, whereas miR-138 downregulation markedly
increased OC3 cell proliferation (Fig.
2C). Additionally, similar results were obtained using OEC-M1
cells (Fig. 2D).
YAP1 is the target gene of miR-138
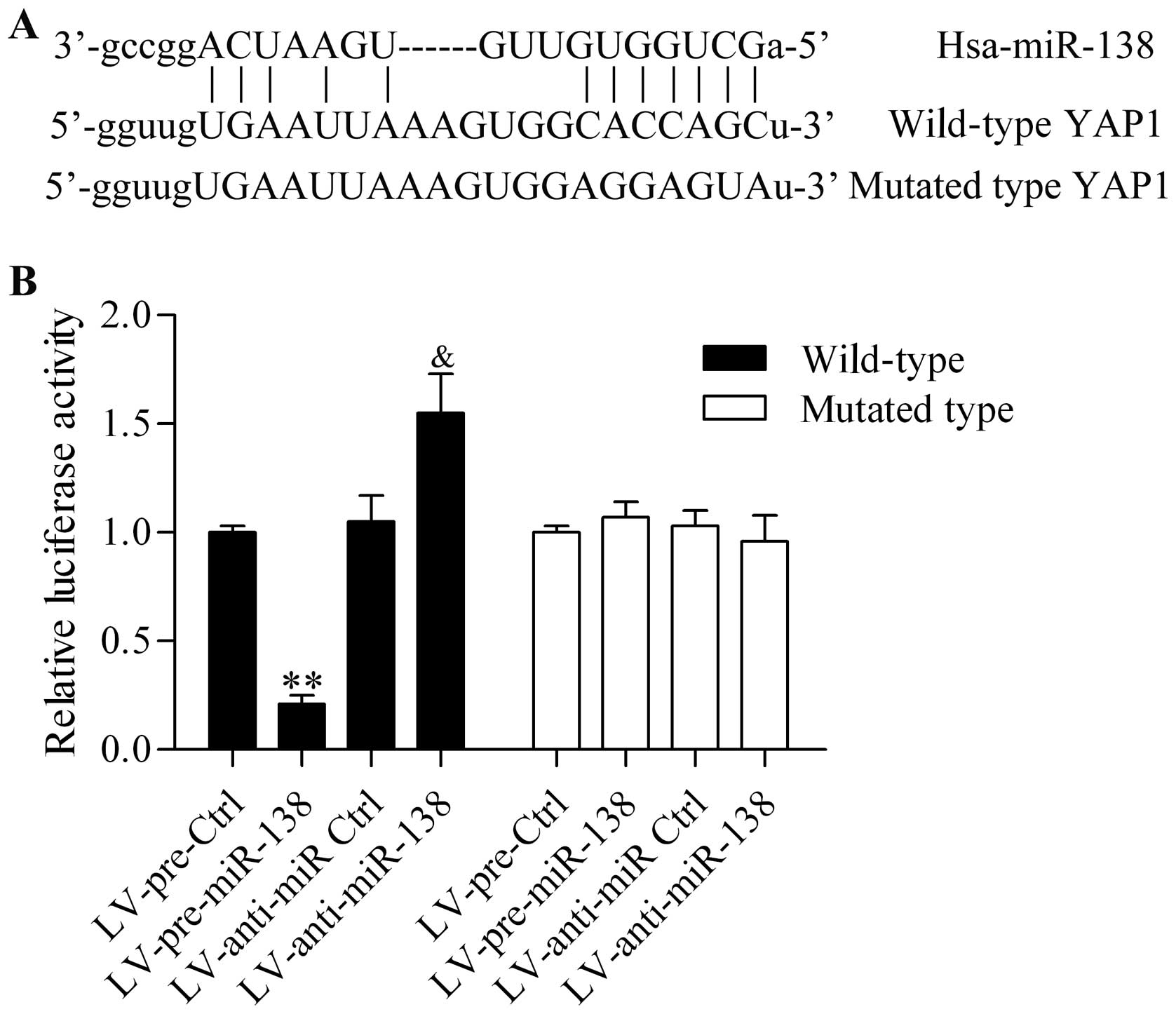
To investigate the potential mechanism of the
miR-138 regulative role in OSCC, we predicted the target gene of
miR-138 by bioinformatics analysis. Notably, we found that the
3′-UTR of YAP1, a candidate oncogene in various types of cancer
(29), processed the putative
binding sites for miR-138 (Fig.
3A). The wild-type and the mutants of 3′-UTR of YAP1 in the
predicted binding sequences were constructed into luciferase
reporters to confirm the direct binding between miR-138 and 3′-UTR
of YAP1. These reporters were then transfected into HEK293 cells
with miR-138 overexpression or downregulation by infection with
LV-pre-miR-138 and LV-anti-138. The reporter assay exhibited that
miR-138 overexpression significantly inhibited the luciferase
activity in wild-type transfected cells, whereas miR-138
downregulation markedly increased the luciferase activity in
wild-type transfected cells. Additionally, the aberrant expression
of miR-138 had no obvious effect on the luciferase activity in the
mutated transfected cells (Fig.
3B).
miR-138 reduces the expression of YAP1 in
OSCC cells
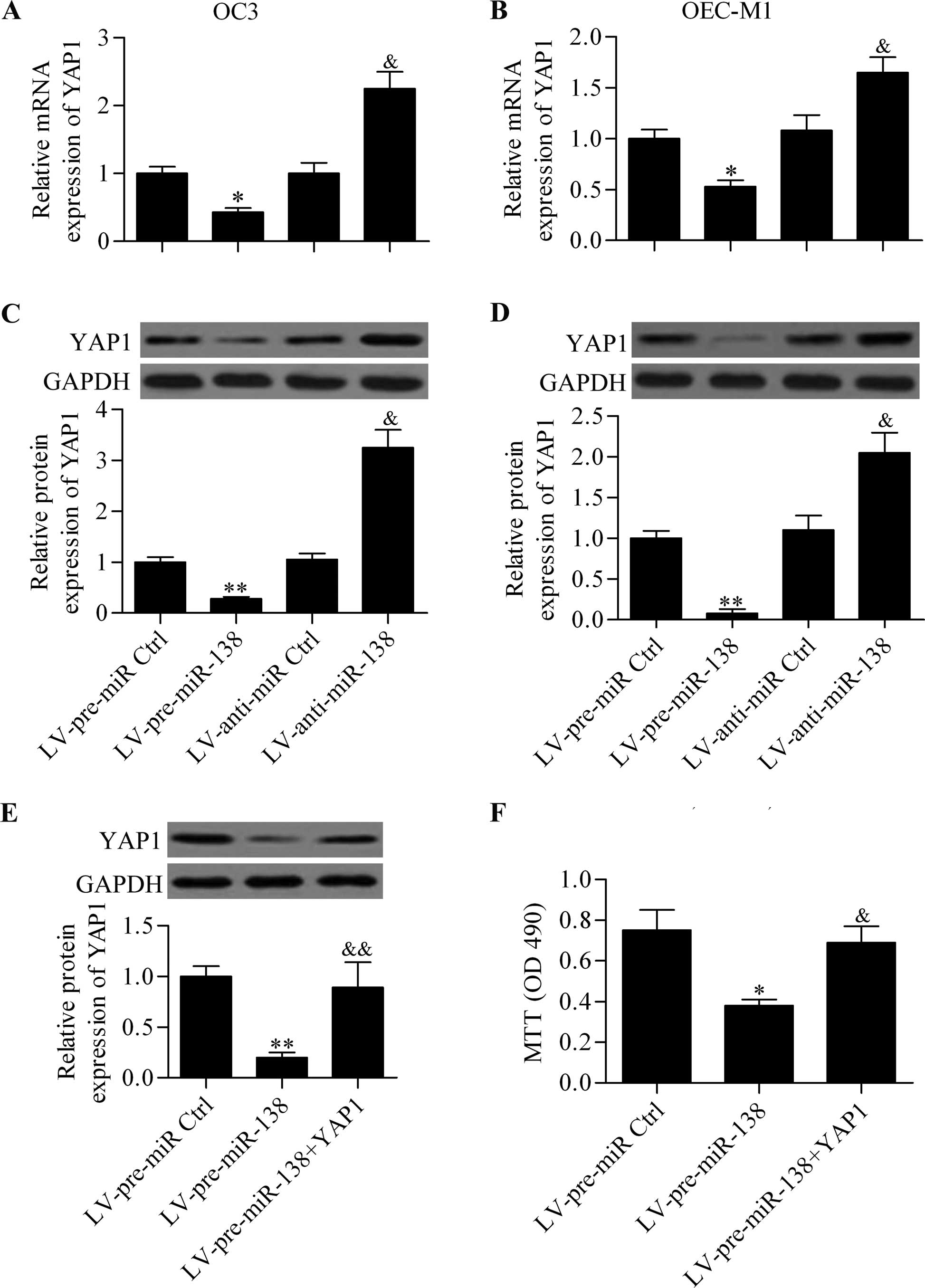
To verify the interaction between miR-138 and YAP1,
we observed the effect of the ectopic expression of miR-138 on YAP1
expression in OSCC cells. RT-qPCR analysis revealed that miR-138
overexpression significantly decreased the mRNA expression of YAP1
in OC3 (Fig. 4A) and OEC-M1
(Fig. 4B). By contrast, miR-138
inhibition markedly increased the YAP1 mRNA expression in OC3 and
OEC-M1 cells. Similar results were obtained in the western blot
analysis, which demonstrated that the protein expression of YAP1
was repressed by miR-138 overexpression or promoted by miR-138
downregulation (Fig. 4C and D). To
assess whether miR-138 regulated the cell proliferation of OSCC
cells by action on YAP1, we transfected YAP1 overexpression
vectors, which harbored no specific miR-138 binding specific
sequences in 3′-UTR in miR-138-overexpressing cells. The results
showed that the overexpression of YAP1 significantly abrogated the
inhibitory effect of miR-138 on the cell proliferation of OSCC
cells (Fig. 4E and F).
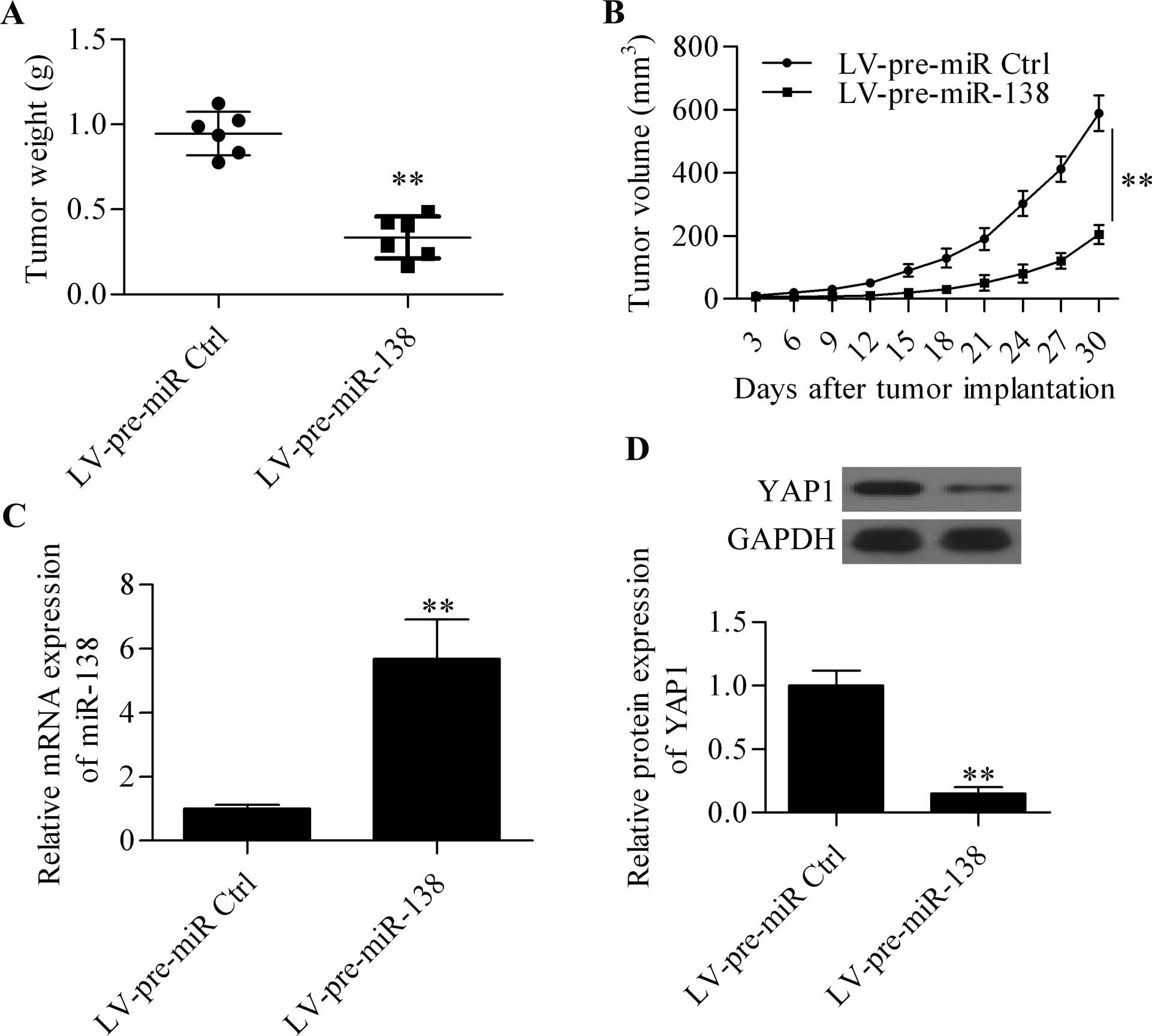
miR-138 represses the xenograft tumor
growth in vivo
To confirm the above findings of miR-138, an in
vivo xenograft assay was performed using overexpressed stable
OC3 cells to test the inhibitory effect of miR-138 on tumor growth
in vivo. Compared with the control group, the overexpression
of miR-138 significantly reduced the average tumor weight (Fig. 5A) and tumor volume (Fig. 5B) in nude mice subcutaneously
injected with OC3 cells. Moreover, the expression of miR-138 and
YAP1 in harvested tumor tissues were detected. The results showed
that miR-138 was significantly increased in the tumors derived from
miR-138-overexpressing OC3 cells (Fig.
5C), whereas the protein expression level of YAP1 was
significantly decreased (Fig.
5D).
Discussion
In the present study, we have reported that a
decreased expression of miR-138 in OSCC tumor tissues and OSCC
cells occurred frequently, leading to the overexpression of the
oncogene YAP1. Consistent with previous findings (30), our results confirm that miR-138 was
dysregulated in OSCC. Moreover, to the best of our knowledge, for
the first time, we have delineated a direct interaction of miR-138
and YAP1 that may be involved in the regulation of OSCC.
The important role of miR-138 in human types of
cancer has been reported. Studies have shown that miR-138 is
frequently decreased in non-small cell lung cancer tissues and
cells and that the overexpression of miR-138 inhibited cancer cell
growth in vitro and in vivo by targeting the zeste
homolog 2 enhancer (15). More
recently, miR-138 has been recognized as a potential prognostic
factor for overall survival in non-small cell lung cancer patients
by regulating 3-phosphoinositide-dependent protein kinase-1
(31). In addition, Ye et al
reported that miR-138 suppressed lung cancer cell proliferation by
inhibiting 3-phosphoinositide-dependent protein kinase-1 (16). In human anaplastic thyroid carcinoma
cells, the decreased miR-138 was associated with increased human
telomerase reverse transcriptase protein (17). Furthermore, miR-138 was demonstrated
to be involved in regulating hepatocellular and colorectal cancer
by targeting cyclin D3 and the Twist basic helix-loop-helix
transcription factor 2 gene (18,19).
In head and neck squamous cell carcinoma cell lines, the
overexpression of miR-138 inhibited cell invasion and increased
cell cycle arrest and apoptosis, whereas knockdown of miR-138
exhibited the opposite effect (30). Jiang et al reported that
miR-138 inhibited cell migration and invasion in tongue squamous
cell carcinoma through the downregulation of RhoC and ROCK2, which
constitute the pivotal genes of the Rho GTPase pathway (32). Additionally, G protein α inhibiting
activity polypeptide 2 was validated as a direct target gene of
miR-138 in tongue squamous cell carcinoma (33). In the present study, we have
demonstrated that miR-138 directly regulates YAP1 in OSCC cells. We
report that the overexpression of miR-138 effectively decreased the
mRNA and protein expression in OSCC cells as well as cell
proliferation and tumor growth. By contrast, the downregulation of
miR-138 exhibited the opposite effect.
YAP1 has been proposed as an oncogene in many human
types of cancers (23–25). Overexpression of YAP1 has been
suggested to be associated with tumor cell proliferation and
growth, cell metastasis and poor prognosis of cancer cells
(26,27,34,35).
In OSCC, YAP1 has been found to be amplified and overexpressed
(28), suggesting the important
role of YAP1 in OSCC. Moreover, recurrent amplifications of YAP1
have been reported in gingivo-buccal OSCC (36). Zhang et al reported that YAP1
is overexpressed in OSCC cell lines and promotes cell proliferation
by activating Fos-related activator 1 (37). Thus, the therapeutic targeting of
YAP1 is a promising method for repressing cancer. Notably, multiple
miRNAs have been reported to regulate YAP1 expression in human
types of cancers. Recent findings have shown that targeting and
inhibiting YAP1 by miR-141 suppress cell growth and colony
formation and elevate cell apoptosis in pancreatic ductal
adenocarcinoma cells (38). Zhang
et al have delineated that YAP1 is highly expressed in
pancreatic progenitor cells, and miR-375 targets the 3′-UTR of YAP1
and decreases its mRNA and protein expression (39). miR-375 was also reported to regulate
YAP1 expression in avian leukosis (40). In colorectal cancer, miR-375
overexpression accelerates cell apoptosis by directly targeting
YAP1 and its downstream anti-apoptotic targets (41). We have demonstrated that miR-138, a
tumor-suppressor gene, also effectively targets YAP1 and is capable
of suppressing YAP1, mRNA and protein expression levels in OSCC
cells. The downregulated expression of miR-138 in OSCC tissues and
cell lines may account for the increased expression of YAP1.
Furthermore, we found that overexpression of miR-138 significantly
inhibited cell proliferation and tumor growth in vitro and
in vivo. Our results demonstrate a direct interaction
between miR-138 and YAP1 and their critical role in OSCC, and
confirm that the targeting and inhibition of YAP1 by miRNA is a
feasible therapeutic treatment of various types of cancer.
In the present study, we have identified a
significant role of miR-138 in OSCC that suppressed cell
proliferation and growth of OSCC by targeting and inhibiting YAP1
expression. Our results suggest that miR-138 is a tumor suppressor
miRNA in OSCC by targeting and inhibiting YAP1, which serves as a
promising therapeutic target for the treatment of OSCC.
Abbreviations:
|
OSCC
|
oral squamous cell carcinoma
|
|
miR-138
|
microRNA-138
|
|
UTR
|
untranslated region
|
|
YAP1
|
Yes-associated protein 1
|
|
mRNA
|
messenger RNA
|
References
|
1
|
Warnakulasuriya S: Global epidemiology of
oral and oropharyngeal cancer. Oral Oncol. 45:309–316. 2009.
View Article : Google Scholar
|
|
2
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Haddad RI and Shin DM: Recent advances in
head and neck cancer. N Engl J Med. 359:1143–1154. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li Y and Kowdley KV: MicroRNAs in common
human diseases. Genomics Proteomics Bioinformatics. 10:246–253.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jansson MD and Lund AH: MicroRNA and
cancer. Mol Oncol. 6:590–610. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mendell JT and Olson EN: MicroRNAs in
stress signaling and human disease. Cell. 148:1172–1187. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ranganathan K and Sivasankar V: MicroRNAs
- Biology and clinical applications. J Oral Maxillofac Pathol.
18:229–234. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gomes CC, de Sousa SF and Gomez RS:
MicroRNAs: Small molecules with a potentially role in oral squamous
cell carcinoma. Curr Pharm Des. 19:1285–1291. 2013.
|
|
10
|
Manikandan M, Deva Magendhra Rao AK,
Rajkumar KS, Rajaraman R and Munirajan AK: Altered levels of
miR-21, miR-125b-2*, miR-138, miR-155, miR-184, and miR-205 in oral
squamous cell carcinoma and association with clinicopathological
characteristics. J Oral Pathol Med. Dec 8–2014.Epub ahead of print.
View Article : Google Scholar
|
|
11
|
Yoon AJ, Wang S, Shen J, Robine N,
Philipone E, Oster MW, Nam A and Santella RM: Prognostic value of
miR-375 and miR-214-3p in early stage oral squamous cell carcinoma.
Am J Transl Res. 6:580–592. 2014.PubMed/NCBI
|
|
12
|
Shiah SG, Hsiao JR, Chang WM, Chen YW, Jin
YT, Wong TY, Huang JS, Tsai ST, Hsu YM, Chou ST, et al:
Downregulated miR329 and miR410 promote the proliferation and
invasion of oral squamous cell carcinoma by targeting Wnt-7b.
Cancer Res. 74:7560–7572. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tiwari A, Shivananda S, Gopinath KS and
Kumar A: Micro-RNA-125a reduces proliferation and invasion of oral
squamous cell carcinoma cells by targeting estrogen-related
receptor α: Implications for cancer therapeutics. J Biol Chem.
289:32276–32290. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Seike M, Goto A, Okano T, Bowman ED,
Schetter AJ, Horikawa I, Mathe EA, Jen J, Yang P, Sugimura H, et
al: MiR-21 is an EGFR-regulated anti-apoptotic factor in lung
cancer in never-smokers. Proc Natl Acad Sci USA. 106:12085–12090.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang H, Zhang H, Zhao M, Lv Z, Zhang X,
Qin X, Wang H, Wang S, Su J, Lv X, et al: MiR-138 inhibits tumor
growth through repression of EZH2 in non-small cell lung cancer.
Cell Physiol Biochem. 31:56–65. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ye XW, Yu H, Jin YK, Jing XT, Xu M, Wan ZF
and Zhang XY: miR-138 inhibits proliferation by targeting
3-phosphoinositide-dependent protein kinase-1 in non-small cell
lung cancer cells. Clin Respir J. 9:27–33. 2015. View Article : Google Scholar
|
|
17
|
Mitomo S, Maesawa C, Ogasawara S, Iwaya T,
Shibazaki M, Yashima-Abo A, Kotani K, Oikawa H, Sakurai E, Izutsu
N, et al: Downregulation of miR-138 is associated with
overexpression of human telomerase reverse transcriptase protein in
human anaplastic thyroid carcinoma cell lines. Cancer Sci.
99:280–286. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang W, Zhao LJ, Tan YX, Ren H and Qi ZT:
MiR-138 induces cell cycle arrest by targeting cyclin D3 in
hepatocellular carcinoma. Carcinogenesis. 33:1113–1120. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Long L, Huang G, Zhu H, Guo Y, Liu Y and
Huo J: Downregulation of miR-138 promotes colorectal cancer
metastasis via directly targeting TWIST2. J Transl Med. 11:2752013.
View Article : Google Scholar
|
|
20
|
Chen P, Zeng M, Zhao Y and Fang X:
Upregulation of Limk1 caused by microRNA-138 loss aggravates the
metastasis of ovarian cancer by activation of Limk1/cofilin
signaling. Oncol Rep. 32:2070–2076. 2014.PubMed/NCBI
|
|
21
|
Hogg RP, Honorio S, Martinez A,
Agathanggelou A, Dallol A, Fullwood P, Weichselbaum R, Kuo MJ,
Maher ER and Latif F: Frequent 3p allele loss and epigenetic
inactivation of the RASSF1A tumour suppressor gene from region
3p21.3 in head and neck squamous cell carcinoma. Eur J Cancer.
38:1585–1592. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Piccinin S, Gasparotto D, Vukosavljevic T,
Barzan L, Sulfaro S, Maestro R and Boiocchi M: Microsatellite
instability in squamous cell carcinomas of the head and neck
related to field cancerization phenomena. Br J Cancer.
78:1147–1151. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Harvey KF, Pfleger CM and Hariharan IK:
The Drosophila Mst ortholog, hippo, restricts growth and cell
proliferation and promotes apoptosis. Cell. 114:457–467. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Edgar BA: From cell structure to
transcription: Hippo forges a new path. Cell. 124:267–273. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Overholtzer M, Zhang J, Smolen GA, Muir B,
Li W, Sgroi DC, Deng CX, Brugge JS and Haber DA: Transforming
properties of YAP, a candidate oncogene on the chromosome 11q22
amplicon. Proc Natl Acad Sci USA. 103:12405–12410. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhao B, Wei X, Li W, Udan RS, Yang Q, Kim
J, Xie J, Ikenoue T, Yu J, Li L, et al: Inactivation of YAP
oncoprotein by the Hippo pathway is involved in cell contact
inhibition and tissue growth control. Genes Dev. 21:2747–2761.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hamaratoglu F, Gajewski K, Sansores-Garcia
L, Morrison C, Tao C and Halder G: The Hippo tumor-suppressor
pathway regulates apical-domain size in parallel to tissue growth.
J Cell Sci. 122:2351–2359. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Snijders AM, Schmidt BL, Fridlyand J,
Dekker N, Pinkel D, Jordan RC and Albertson DG: Rare amplicons
implicate frequent deregulation of cell fate specification pathways
in oral squamous cell carcinoma. Oncogene. 24:4232–4242. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Seton-Rogers S: Oncogenes: All eyes on
YAP1. Nat Rev Cancer. 14:514–515. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu X, Jiang L, Wang A, Yu J, Shi F and
Zhou X: MicroRNA-138 suppresses invasion and promotes apoptosis in
head and neck squamous cell carcinoma cell lines. Cancer Lett.
286:217–222. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Han L, Zhang G, Zhang N, Li H, Liu Y, Fu A
and Zheng Y: Prognostic potential of microRNA-138 and its target
mRNA PDK1 in sera for patients with non-small cell lung cancer. Med
Oncol. 31:1292014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jiang L, Liu X, Kolokythas A, Yu J, Wang
A, Heidbreder CE, Shi F and Zhou X: Downregulation of the Rho
GTPase signaling pathway is involved in the microRNA-138-mediated
inhibition of cell migration and invasion in tongue squamous cell
carcinoma. Int J Cancer. 127:505–512. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jiang L, Dai Y, Liu X, Wang C, Wang A,
Chen Z, Heidbreder CE, Kolokythas A and Zhou X: Identification and
experimental validation of G protein alpha inhibiting activity
polypeptide 2 (GNAI2) as a microRNA-138 target in tongue squamous
cell carcinoma. Hum Genet. 129:189–197. 2011. View Article : Google Scholar :
|
|
34
|
Liu R, Huang S, Lei Y, Zhang T, Wang K,
Liu B, Nice EC, Xiang R, Xie K, Li J, et al: FGF8 promotes
colorectal cancer growth and metastasis by activating YAP1.
Oncotarget. 6:935–952. 2015.
|
|
35
|
Lee KW, Lee SS, Kim SB, Sohn BH, Lee HS,
Jang HJ, Park YY, Kopetz S, Kim SS, Oh SC, et al: Significant
association of oncogene YAP1 with poor prognosis and cetuximab
resistance in colorectal cancer patients. Clin Cancer Res.
21:357–364. 2015. View Article : Google Scholar :
|
|
36
|
India Project Team of the International
Cancer Genome Consortium: Mutational landscape of gingivo-buccal
oral squamous cell carcinoma reveals new recurrently-mutated genes
and molecular subgroups. Nat Commun. 4:28732013.PubMed/NCBI
|
|
37
|
Zhang L, Ye DX, Pan HY, Wei KJ, Wang LZ,
Wang XD, Shen GF and Zhang ZY: Yes-associated protein promotes cell
proliferation by activating Fos Related Activator-1 in oral
squamous cell carcinoma. Oral Oncol. 47:693–697. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhu ZM, Xu YF, Su QJ, Du JD, Tan XL, Tu
YL, Tan JW and Jiao HB: Prognostic significance of microRNA-141
expression and its tumor suppressor function in human pancreatic
ductal adenocarcinoma. Mol Cell Biochem. 388:39–49. 2014.
View Article : Google Scholar
|
|
39
|
Zhang ZW, Men T, Feng RC, Li YC, Zhou D
and Teng CB: miR-375 inhibits proliferation of mouse pancreatic
progenitor cells by targeting YAP1. Cell Physiol Biochem.
32:1808–1817. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li H, Shang H, Shu D, Zhang H, Ji J, Sun
B, Li H and Xie Q: gga-miR-375 plays a key role in tumorigenesis
post subgroup J avian leukosis virus infection. PLoS One.
9:e908782014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Christensen LL, Holm A, Rantala J,
Kallioniemi O, Rasmussen MH, Ostenfeld MS, Dagnaes-Hansen F, Øster
B, Schepeler T, Tobiasen H, et al: Functional screening identifies
miRNAs influencing apoptosis and proliferation in colorectal
cancer. PLoS One. 9:e967672014. View Article : Google Scholar : PubMed/NCBI
|