Introduction
Oral squamous cell carcinoma (OSCC) is a common type
of cancer of the head and neck region, with a high mortality rate
and rising incidence (1,2). Oral carcinogenesis is a complex
multi-step process, undergoing sequential histopathological stages
including hyperplasia, oral epithelial dysplasia and final
malignant phenotypes (3,4). Despite significant advances in the
diagnosis and treatment of OSCC in recent years, 5-year survival
rate remains low due to late-stage diagnosis.
Oral submucous fibrosis (OSF), as a pre-cancerous
condition (5), is a chronic and
potentially malignant disorder, first described in the early 1950s
by Pindborg and Sirsat (6). OSF is
predominantly prevalent in Indian, mainland China (such as Hunan
and Hainan), Taiwan, Bangladesh and Pakistan (7). It is characterized by submucosal
fibrosis initially occurring in the oral cavity and then
progressively infringing the pharynx and upper esophagus (8). The malignant transformation rate of
OSF has been reported in the range of 7–30% based on the different
population (9). The etiology of OSF
is multifactorial and remains vague. Chewing of betel quid
containing areca nut is the strongest risk factor of OSF, and other
factors include tobacco use, alcohol drinking, as well as genetic
and immunologic predisposition (10–12).
Thus, elucidating the molecular events of OSF malignant progression
may be useful in identifying potential biomarkers for the early
diagnosis and prevention of OSCC.
Epigenetic modifications including the promoter CpG
methylation and histone modification have been recognized as being
crucial events in cancer development, such as genetic alterations
(13,14). Aberrant promoter methylation leads
to the transcriptional silencing of tumor-suppressor genes (TSGs),
involved in cell malignant transformation and tumorigenesis.
Detection of promoter CpG methylation in human DNA isolated from
primary tumor tissues and bodily fluids has become a promising
approach for non-invasive screening and early diagnosis of cancer
(15). Several TSGs have been
identified methylated in OSCC (16–19),
such as p16, p14arf, E-cadherin,
O-6-methylgua-nine-DNA methyltransferase (MGMT),
death-associated protein kinase (DAPK), runt-related
transcription factor 3 (RUNX3), as well as certain Wnt
antagonists.
The Wnt/β-catenin signaling pathway plays an
important role in human malignancies including OSCC (20,21).
Wnt inhibitory factor-1 (WIF1) as a secreted WNT antagonist
inhibits Wnt/β-catenin signaling by directly binding to Wnt
proteins (22). However, few
studies have focused on the epigenetic disruption of WIF1 in
OSF progression. In the present study, we investigated the
expression level and methylation status of WIF1, a key antagonist
in WNT signaling, in the carcinogenesis of OSF.
Materials and methods
Tissue specimens
OSCC (n=55), OSF (n=45) and normal oral mucosa
(n=15) tissue specimens were obtained at the time of surgical
resection at the Xiangya Second Hospital and Xiangya Hospital,
Central South University (Changsha, China) and Shanghai Ninth
People's Hospital, Shanghai Jiaotong University School of Medicine
(Shanghai, China) from Juanuary 2013 to June 2014. Informed consent
from the patients was obtained under a protocol reviewed and
approved by the Institutional Review Boards of the Xiangya School
of Medicine or Shanghai Jiaotong University School of Medicine. The
clinical diagnosis and pathological stage of OSF was determined in
terms of the Pindborg criteria (23). OSCC was diagnosed according to the
World Health Organization criteria of 2005. Fifteen normal
specimens were obtained from healthy oral mucosa. Forty-five cases
of OSF were incident, newly diagnosed without OSCC or neoplastic
disease. OSF was classified into 3 grades: early stage (E, n=15),
moderately advanced stage (M, n=15) and advanced stage (A, n=15).
The collected tissues were divided into two groups. Tissues in the
first group were frozen immediately after careful removal of the
tumor mass, OSF tissue in epithelium layer, and grossly
normal-looking epithelium, all at −80°C. Tissues in the second
group were fixed in 4% buffered formalin solution for pathological
diagnosis and immunohistochemical staining. Clinicopathological
staging of OSCC was determined by the TNM classification of the
International Union Against Cancer in 2009.
Immunohistochemistry
Immunohistochemical staining was performed on
4-µm serial sections from formalin-fixed paraffin-embedded
specimens. Following deparaffinization and hydration, the slides
were treated with endogenous peroxidase in 3%
H2O2 for 20 min, after which the sections
were blocked for 30 min at 37°C with 1.5% blocking serum in
phosphate-buffered saline (PBS) prior to reacting with WIF1
antibody (1:100 dilution, AP2723a; Abgent, San Diego, CA, USA) at
4°C in a moist chamber overnight. Negative control slides were
duplicate sections in the absence of primary antibodies. To
evaluate WIF1 expression, a scoring method was used. A mean
percentage of positive tumor cells was determined by the
examination of 500 cells in at least 5 areas at ×400 magnification.
The cells were assigned to 1 of the 5 following categories
according to percentage of positive cells (PP): 0) <5%, 1)
5–24%, 2) 25–49%, 3) 50–75%, and 4) >75%. The intensity of the
WIF1 staining (SI) then was scored as follows: 0) no, −; 1) weak,
+; 2) moderate, ++; and 3) intense, +++. The final immunoreactive
score (IRS=SI + PP) was as follows: −) 0, 1; +) 2, 3; ++) 4, 5; and
+++) 6, 7. The stained tissues were scored blindly with regard to
clinical patient data. Statistical analyses were performed with the
SPSS 17.0 software. Statistical significance was evaluated by the
Chi-square test (χ2).
Reverse transcription-polymerase chain
reaction
Total RNAs were extracted using TRIzol reagent
(Invitrogen Life Technologies, Karlsruhe, Germany) according to the
manufacturer's instructions. Reverse transcription polymerase chain
reaction (RT-PCR) was performed with the use of a kit from Promega
(Madison, WI, USA). Quantitative PCR was performed to detect
WIF1 expression, according to the manufacturer's
instructions (HT7500 system; Applied Biosystems, Foster City, CA,
USA). Primers for the amplification of WIF1 mRNA sequences
(accession no. U75285) were synthesized as previously described
(24,25). The 257-bp mRNA of WIF1 was
amplified by PCR using the primers: forward, 5′-TATGGA
TCGATGCTCACCAG and reverse, 5′-CAGAGGGACATT GACGGTTG. GAPDH
was used as an internal control. Primers used for GAPDH
were: forward, 5′-ATCTCTGCC CCCTCTGCTGA-3′ and the reverse,
5′-GATGACCTTGCC CACAGCCT-3′. PCR amplification was performed as
follows: denaturation at 94°C for 30 sec, annealing at 55°C for 30
sec, and extension at 72°C for 30 sec in 32 cycles. The PCR
products were visualized on 2% agarose gels under ultraviolet
transillumination.
Methylation-specific PCR and bisulfite
genomic sequencing
Bisulfite modification of DNA, methylation-specific
PCR (MSP) and bisulfite genomic sequencing (BGS) were performed as
previously described (24–27). Bisulfite-treated DNA was amplified
with the methylation-specific primer set WIF1-m3:
5′-TGTCGTTTTTATTTTCGTTCGC, WIF1-m4: 5′-CGTTTAAACGACTAAACGCG
or the unmethylation specific primer set WIF1-u3:
5′-TTTTTGTTGTTTTTATTT TTGTTTGT, WIF1-u4:
5′-TCCCATTTAAACAACTAAAC ACA. MSP was performed using AmpliTaq Gold
(methylation-specific primer: annealing temperature 60°C, 40
cycles, unmethylation-specific primer: annealing temperature 58°C,
40 cycles). For BGS, bisulfite-treated DNA was amplified using
primers BGS1: 5′-GTTTTAGGGGTTTTTGAGTGTT and BGS2:
5′-CAACTCCCTCAACCAAAACTA. The 463-bp PCR products were cloned into
the pCR4-TOPO vector (Invitrogen, Carlsbad, CA, USA), with 8–10
colonies being randomly selected and sequenced.
Results
Downregulation of WIF1 expression at the
protein level in the carcinogenesis of OSF
To evaluate WIF1 protein expression, we initially
performed immunohistochemical staining using WIF1-specific antibody
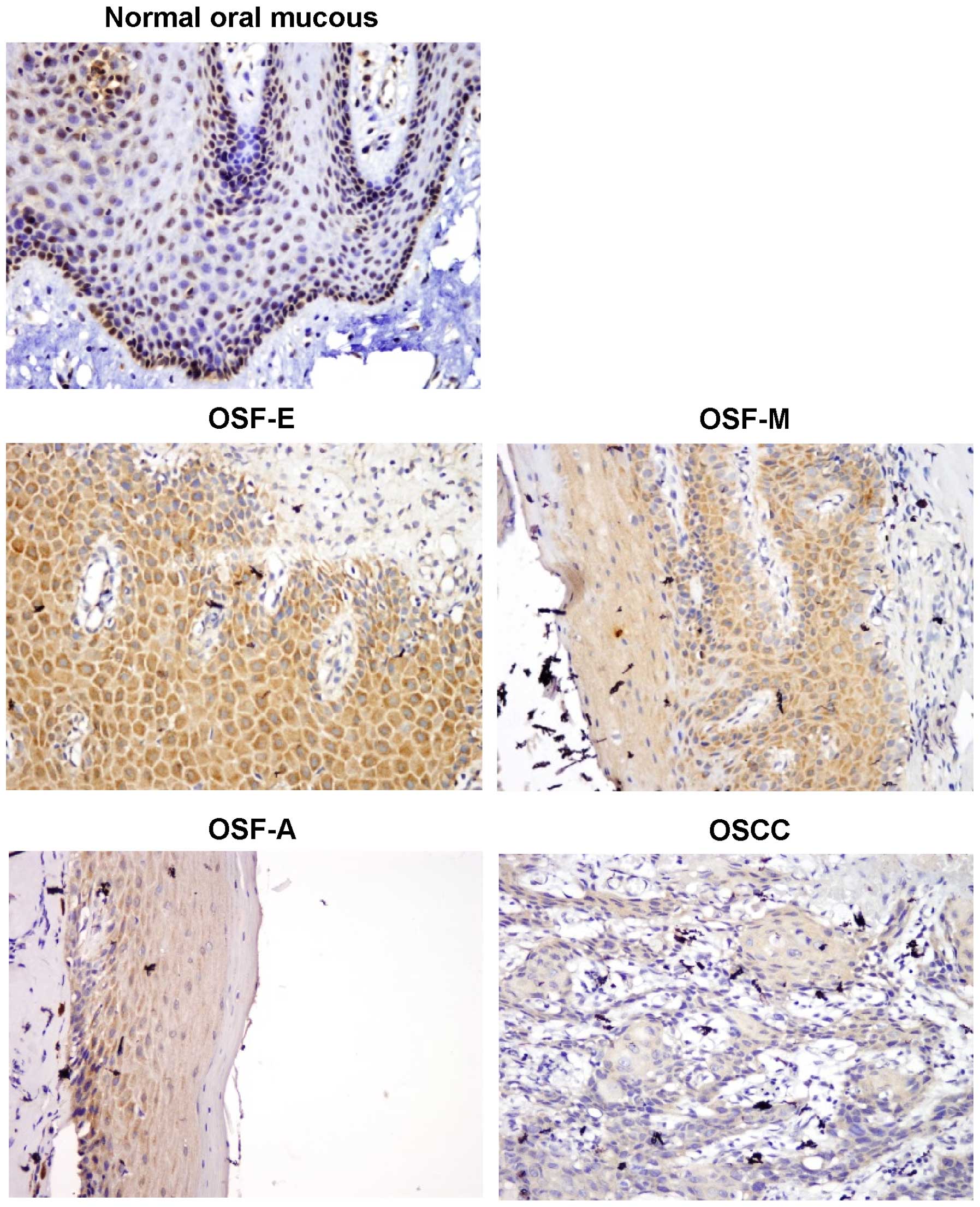
in normal oral mucous tissues, OSF and OSCC tissues. Fourteen of 15
(93.3%) normal oral mucous cases showed nuclear WIF1 positivity,
while 33 of 45 (73.3%) OSF tissues showed cytoplasmic WIF1
expression, including tissues from 13 of 15 (86.7%) early stage, 12
of 15 (80%) moderately advanced stage and 8 of 15 (53.3%) advanced
stage, as well as 20 of 55 (36.4%) OSCC (Fig. 1). The average values of WIF1
expression varied in different tissue samples, including mean score
of 5.37 in normal oral mucous tissues, 3.29 in OSF tissues, and
1.27 in OSCC tissues (Table I).
WIF1 expression was gradually reduced in normal oral tissues, OSF
and OSCC tissues (P=0.00001). However, no statistically significant
correlation was observed between WIF1 expression and the
clinicopathological characteristics of OSCC (P>0.05) (Table II). These results suggested that
WIF1 is downregulated at a protein level in the carcinogenesis of
OSF.
 | Table IWIF1 expression in the carcinogenesis
of oral submucous fibrosis (OSF). |
Table I
WIF1 expression in the carcinogenesis
of oral submucous fibrosis (OSF).
| Group | n | WIF1
| | Mean WIF1
|
|---|
| − | + | ++ | +++ | WIF1
expression | score |
|---|
| Normal | 15 | 1 | 0 | 5 | 9 | 93.3% | 5.73 |
| OSF | 45 | 12 | 10 | 17 | 6 | 73.3% | 3.29 |
| E | 15 | 2 | 2 | 8 | 3 | 86.7% | 4.13 |
| M | 15 | 3 | 3 | 7 | 2 | 80.0% | 3.60 |
| A | 15 | 7 | 5 | 2 | 1 | 53.3% | 2.13 |
| OSCC | 55 | 35 | 18 | 2 | 0 | 36.4% | 1.27 |
| Table IICorrelation between WIF1 expression
and clinicopathological characteristics in OSCC cases |
Table II
Correlation between WIF1 expression
and clinicopathological characteristics in OSCC cases
| Clinicopathological
characteristics | Total (n) | Results of
immunostaining, (n)
|
|---|
| WIF1 (+) | WIF1 (−) | P-value |
|---|
| Age (years) | | | | >0.05 |
| <50 | 43 | 18 | 25 | |
| ≥50 | 12 | 2 | 10 | |
| Gender | | | | >0.05 |
| Male | 52 | 20 | 32 | |
| Female | 3 | 0 | 3 | |
| Tumor site | | | | >0.05 |
| Tongue | 40 | 17 | 23 | |
| Others | 15 | 3 | 12 | |
| Primary tumor | | | | >0.05 |
| T1+T2 | 12 | 2 | 10 | |
| T3+T4 | 43 | 18 | 25 | |
| TNM stage |
| I+II | 12 | 7 | 5 | >0.05 |
| III+IV | 43 | 13 | 30 | |
| Differentiation
grade | | | | >0.05 |
| Well | 20 | 5 | 15 | |
|
Moderately-poorly | 35 | 15 | 20 | |
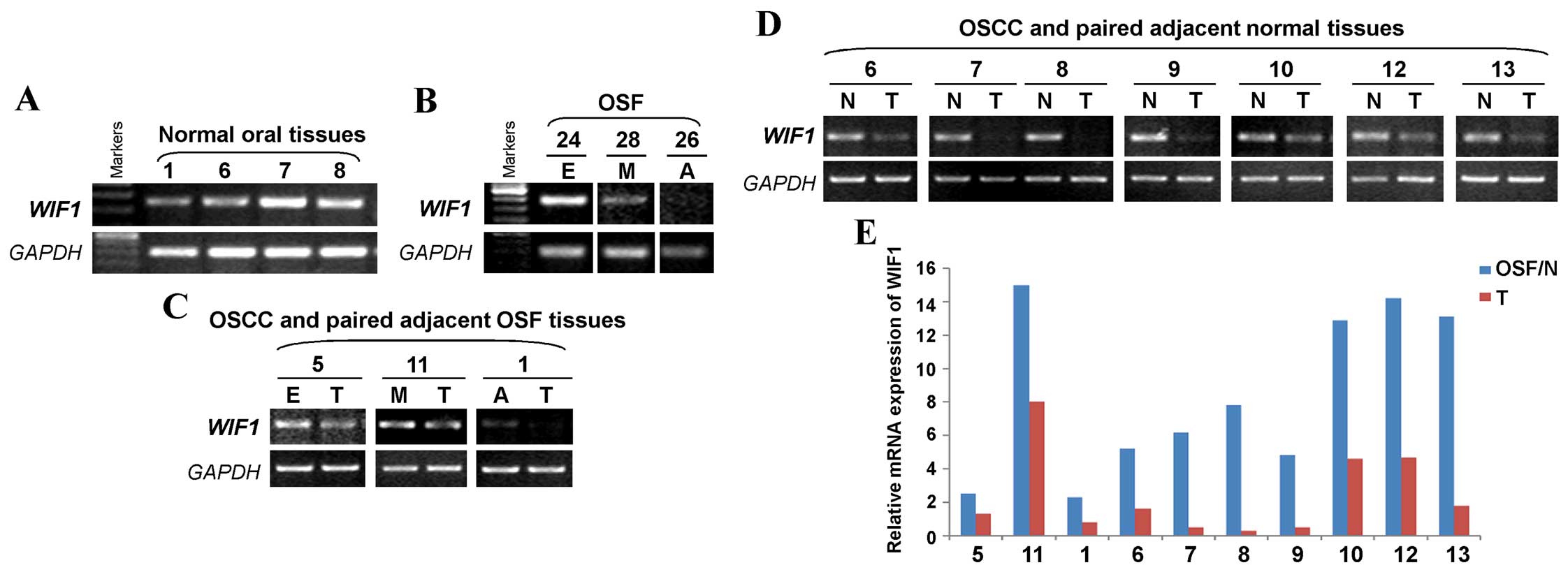
WIF1 mRNA expression is reduced in the
carcinogenesis of OSF
We also examined WIF1 expression at the mRNA
level in normal oral mucous tissues, OSF tissues, OSCC and their
paired adjacent tissues by semi-quantitative RT-PCR. The presence
of the WIF1 mRNA was shown by the 257-bp RT-PCR product. We
found that WIF1 was readily expressed in normal oral mucous
tissues (Fig. 2A) and OSF early
stage tissue, but decreased in OSF moderately advanced stage
tissue, while rarely expressed in OSF advanced stage tissue
(Fig. 2B). We also detected
WIF1 expression in OSCC and their adjacent OSF tissues. The
results showed that WIF1 was highly expressed in OSF early
stage and moderately advanced stage tissues, but markedly
downregulated in OSF advanced stage and OSCC tissues (Fig. 2C). We also found that WIF1
was barely detected in OSCC tissues, compared with their paired
adjacent normal tissues (Fig. 2D).
RT-qPCR also confirmed that a reduced expression of WIF1
mRNA in OSCC tissues, compared to the adjacent normal or OSF
tissues (Fig. 2E). Therefore,
WIF1 mRNA expression levels are decreased in the
carcinogenesis of OSF.
Promoter methylation of WIF1 in the
carcinogenesis of OSF
We investigated the possible regulatory mechanism of
WIF1 reduction in the carcinogenesis of OSF. As promoter
methylation mediates transcriptional repression of TSGs, we
initially examined the presence of CpG island (CGI) in the
WIF1 promoter and exon 1 by bioinformatics analysis. The
region spanning the WIF1 promoter and exon 1 fulfilled the
criteria of a CpG island (Gardiner-Garden and Frommer, 1987): GC
content, 58.8%; observed/expected CpG ratio, 0.73; and a total of
46 CpG sites in a 2026-bp region (Fig.
3), thus as a typical CGI.
We detected the promoter methylation of WIF1
in normal oral mucous and OSF tissues, OSCC and their paired
adjacent OSF or normal tissues. We found that WIF1
methylation was not detected in 10 normal oral tissues, 10 OSF
tissues from early stage, moderately advanced stage and advanced
stage (Fig. 4A and B). We also
found that WIF1 was frequently methylated in 16 of 20 (80%)
OSCC tumor tissues, but not any in their paired adjacent normal and
OSF tissues (Fig. 4C–E). These
results suggested that the promoter methylation of WIF1 is a
tumor-specific event in the carcinogenesis of OSF.
 | Figure 4Promoter methylation of WIF1
in normal oral mucosa, OSF and OSCC tissues. MSP was used to detect
WIF1 methylation in (A) normal oral mucosa tissues, (B) OSF
tissues, (C) OSCC and paired adjacent OSF tissues, (D) OSCC and
paired adjacent normal tissues, as well as (E) OSCC tissues. M,
methylated; U, unmethylated; E, early stage of OSF; M, moderately
advanced stage of OSF; A, advanced stage of OSF; N, normal tissue;
T, OSCC. |
Bisulfite genomic sequencing of the WIF1
promoter in OSF and OSCC tumor tissues
To confirm MSP data, high resolution methylation
analysis on every CpG site in the WIF1 promoter was carried
out by using bisulfite genomic sequencing (BGS). We found that no
or very few methylated CpG sites were detected in representative
normal oral mucous tissues, and OSF early stage and moderately
advanced stage tissues, while methylated alleles were detected in
OSCC tissues, but not in their paired normal and OSF tissues. The
results confirmed the MSP analysis (Fig. 5).
Discussion
Emerging evidence has shown that the epigenetic
silencing of cancer genes via promoter CpG methylation plays an
important role in OSCC pathogenesis. Several genes have been
identified to be aberrantly methylated in OSCC, some of which
exhibited a potential as biomarkers, for example, E-cadherin
promoter methylation is associated with the poor survival in
advanced OSCC (16), thus is an
ideal epigenetic biomarker for OSCC. Thus, identifying more
methylated genes may be useful to develop epigenetic biomarker for
the early detection of OSCC.
Wnt signaling pathways include the canonical Wnt,
the non-canonical planar cell polarity and the non-canonical
Wnt/calcium pathways (28). The
canonical Wnt pathway, also known as Wnt/β-catenin pathway,
accumulates cytoplasmic β-catenin in and eventual translocates into
the nucleus, thus acting as a transcriptional coactivator to
activate multiple oncogenic genes, leading to tumorigenesis
(29). The Wnt/β-catenin pathway is
deregulated in various common human cancers (30). Frequent mutations of the signaling
molecules in the Wnt/β-catenin pathway have been identified in
human cancers, including colon, hepatocellular, breast and prostate
carcinomas as well as glioblastoma. However, infrequent mutations
of APC, Axin1 and β-catenin genes were reported in OSCC (31), indicating that mechanisms such as
epigenetic modulation mediate inactivation of the Wnt/β-catenin
pathway in OSCC.
WIF1, a secreted Wnt inhibitor, is
downregulated and methy lated in various carcinomas including
hepatocellular (32),
nasopharyngeal (24,33), esophageal (5), prostate (34), breast (25), lung (35) and gastrointestinal (36) malignancies, and promotes tumor
development and progression by activating β-catenin. Notably,
WIF1 silencing may be an early event in tumorigenesis.
Although OSF is the precancerous lesion of OSCC, its molecular
mechanisms remain to be elucidated. In the present study, we found
that WIF1 either at the protein or mRNA level is highly expressed
in normal oral mucous tissues, and is gradually decreased in the
different stages of OSF and OSCC tissues. Additionally, WIF1
is frequently methylated in OSCC tissues, but not in normal oral
mucous tissues, and their paired adjacent normal or OSF tissues. We
also detected WIF1 methylation in a panel of OSCC patient
tissues with betel quid chewing habit, and observed WIF1
methylation. These results are consistent with those of other
studies on WIF1 methylation in OSCC in Western populations
(18). WIF1 methylation was
also reported to be correlated with shorter survival in oral cancer
patients (37). Thus, it may become
a tumor marker for the early detection of OSCC. Large-scale studies
focusing on OSF and OSCC samples should be conducted to confirm the
potential value of WIF1 methylation as an epigenetic
biomarker in OSCC in the Chinese population.
In summary, we provide evidence that WIF1 is
frequently methylated in OSCC in a Chinese population, but not in
normal oral mucous and OSF tissues, which is associated with its
reduced expression. The present study reveals a novel epigenetic
event in the carcinogenesis of OSF, which shed light on the
development of a valuable epigenetic biomarker that may be useful
for the early detection of OSCC.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (no. 81202133) and the Fifth
Outstanding youth Fund of Shanghai ninth People's Hospital,
Shanghai Jiaotong University School of Medicine.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Braakhuis BJ, Brakenhoff RH and Leemans
CR: Head and neck cancer: Molecular carcinogenesis. Ann Oncol.
16(Suppl 2): ii249–ii250. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Leemans CR, Braakhuis BJ and Brakenhoff
RH: The molecular biology of head and neck cancer. Nat Rev Cancer.
11:9–22. 2011. View
Article : Google Scholar
|
|
4
|
Choi S and Myers JN: Molecular
pathogenesis of oral squamous cell carcinoma: Implications for
therapy. J Dent Res. 87:14–32. 2008. View Article : Google Scholar
|
|
5
|
Pindborg JJ, Murti PR, Bhonsle RB, Gupta
PC, Daftary DK and Mehta FS: Oral submucous fibrosis as a
precancerous condition. Scand J Dent Res. 92:224–229.
1984.PubMed/NCBI
|
|
6
|
Pindborg JJ and Sirsat SM: Oral submucous
fibrosis. Oral Surg Oral Med Oral Pathol. 22:764–779. 1966.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tilakaratne WM, Klinikowski MF, Saku T,
Peters TJ and Warnakulasuriya S: Oral submucous fibrosis: Review on
aetiology and pathogenesis. Oral Oncol. 42:561–568. 2006.
View Article : Google Scholar
|
|
8
|
Wollina U, Verma SB, Ali FM and Patil K:
Oral submucous fibrosis: An update. Clin Cosmet Investig Dermatol.
8:193–204. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Murti PR, Bhonsle RB, Pindborg JJ, Daftary
DK, Gupta PC and Mehta FS: Malignant transformation rate in oral
submucous fibrosis over a 17-year period. Community Dent Oral
Epidemiol. 13:340–341. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang X and Reichart PA: A review of betel
quid chewing, oral cancer and precancer in Mainland China. Oral
Oncol. 43:424–430. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pillai R, Balaram P and Reddiar KS:
Pathogenesis of oral submucous fibrosis. Relationship to risk
factors associated with oral cancer. Cancer. 69:2011–2020. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jian XC, Liu SF, Shen ZH and Yang YH:
Histomorphology of oral submucous fibrosis. Report of 24 cases.
Chin Med J (Engl). 101:505–509. 1988.
|
|
13
|
Jones PA and Baylin SB: The fundamental
role of epigenetic events in cancer. Nat Rev Genet. 3:415–428.
2002.PubMed/NCBI
|
|
14
|
Jones PA and Baylin SB: The epigenomics of
cancer. Cell. 128:683–692. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Baylin SB and Jones PA: A decade of
exploring the cancer epigenome - biological and translational
implications. Nat Rev Cancer. 11:726–734. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Supić G, Kozomara R, Branković-Magić M,
Jović N and Magić Z: Gene hypermethylation in tumor tissue of
advanced oral squamous cell carcinoma patients. Oral Oncol.
45:1051–1057. 2009. View Article : Google Scholar
|
|
17
|
Towle R, Truong D, Hogg K, Robinson WP,
Poh CF and Garnis C: Global analysis of DNA methylation changes
during progression of oral cancer. Oral Oncol. 49:1033–1042. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pannone G, Bufo P, Santoro A, Franco R,
Aquino G, Longo F, Botti G, Serpico R, Cafarelli B, Abbruzzese A,
et al: WNT pathway in oral cancer: Epigenetic inactivation of
WNT-inhibitors. Oncol Rep. 24:1035–1041. 2010.PubMed/NCBI
|
|
19
|
Ha PK and Califano JA: Promoter
methylation and inactivation of tumour-suppressor genes in oral
squamous-cell carcinoma. Lancet Oncol. 7:77–82. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu F and Millar SE: Wnt/beta-catenin
signaling in oral tissue development and disease. J Dent Res.
89:318–330. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Noguti J, DE Moura CF, Hossaka TA, Franco
M, Oshima CT, Dedivitis RA and Ribeiro DA: The role of canonical
WNT signaling pathway in oral carcinogenesis: A comprehensive
review. Anticancer Res. 32:873–878. 2012.PubMed/NCBI
|
|
22
|
Hsieh JC, Kodjabachian L, Rebbert ML,
Rattner A, Smallwood PM, Samos CH, Nusse R, Dawid IB and Nathans J:
A new secreted protein that binds to Wnt proteins and inhibits
their activities. Nature. 398:431–436. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gupta PC, Sinor PN, Bhonsle RB, Pawar VS
and Mehta HC: Oral submucous fibrosis in India: A new epidemic?
Natl Med J India. 11:113–116. 1998.PubMed/NCBI
|
|
24
|
Chan SL, Cui Y, van Hasselt A, Li H,
Srivastava G, Jin H, Ng KM, Wang Y, Lee KY, Tsao GS, et al: The
tumor suppressor Wnt inhibitory factor 1 is frequently methylated
in nasopharyngeal and esophageal carcinomas. Lab Invest.
87:644–650. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ai L, Tao Q, Zhong S, Fields CR, Kim WJ,
Lee MW, Cui Y, Brown KD and Robertson KD: Inactivation of Wnt
inhibitory factor-1 (WIF1) expression by epigenetic silencing is a
common event in breast cancer. Carcinogenesis. 27:1341–1348. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Qiu GH, Tan LK, Loh KS, Lim CY, Srivastava
G, Tsai ST, Tsao SW and Tao Q: The candidate tumor suppressor gene
BLU, located at the commonly deleted region 3p21.3, is an
E2F-regulated, stress-responsive gene and inactivated by both
epigenetic and genetic mechanisms in nasopharyngeal carcinoma.
Oncogene. 23:4793–4806. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ying J, Li H, Seng TJ, Langford C,
Srivastava G, Tsao SW, Putti T, Murray P, Chan AT and Tao Q:
Functional epigenetics identifies a protocadherin PCDH10 as a
candidate tumor suppressor for nasopharyngeal, esophageal and
multiple other carcinomas with frequent methylation. Oncogene.
25:1070–1080. 2006. View Article : Google Scholar
|
|
28
|
Anastas JN and Moon RT: WNT signalling
pathways as therapeutic targets in cancer. Nat Rev Cancer.
13:11–26. 2013. View
Article : Google Scholar
|
|
29
|
Clevers H: Wnt/beta-catenin signaling in
development and disease. Cell. 127:469–480. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Polakis P: The many ways of Wnt in cancer.
Curr Opin Genet Dev. 17:45–51. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Iwai S, Katagiri W, Kong C, Amekawa S,
Nakazawa M and Yura Y: Mutations of the APC, beta-catenin, and axin
1 genes and cytoplasmic accumulation of beta-catenin in oral
squamous cell carcinoma. J Cancer Res Clin Oncol. 131:773–782.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ding Z, Qian YB, Zhu LX and Xiong QR:
Promoter methylation and mRNA expression of DKK-3 and WIF-1 in
hepatocellular carcinoma. World J Gastroenterol. 15:2595–2601.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lin YC, You L, Xu Z, He B, Mikami I, Thung
E, Chou J, Kuchenbecker K, Kim J, Raz D, et al: Wnt signaling
activation and WIF-1 silencing in nasopharyngeal cancer cell lines.
Biochem Biophys Res Commun. 341:635–640. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yee DS, Tang Y, Li X, Liu Z, Guo Y,
Ghaffar S, McQueen P, Atreya D, Xie J, Simoneau AR, et al: The Wnt
inhibitory factor 1 restoration in prostate cancer cells was
associated with reduced tumor growth, decreased capacity of cell
migration and invasion and a reversal of epithelial to mesenchymal
transition. Mol Cancer. 9:1622010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mazieres J, He B, You L, Xu Z, Lee AY,
Mikami I, Reguart N, Rosell R, McCormick F and Jablons DM: Wnt
inhibitory factor-1 is silenced by promoter hypermethylation in
human lung cancer. Cancer Res. 64:4717–4720. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Taniguchi H, Yamamoto H, Hirata T,
Miyamoto N, Oki M, Nosho K, Adachi Y, Endo T, Imai K and Shinomura
Y: Frequent epigenetic inactivation of Wnt inhibitory factor-1 in
human gastrointestinal cancers. Oncogene. 24:7946–7952. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Paluszczak J, Sarbak J,
Kostrzewska-Poczekaj M, Kiwerska K, Jarmuż-Szymczak M, Grenman R,
Mielcarek-Kuchta D and Baer-Dubowska W: The negative regulators of
Wnt pathway-DACH1, DKK1, and WIF1 are methylated in oral and
oropharyngeal cancer and WIF1 methylation predicts shorter
survival. Tumour Biol. 36:2855–2861. 2015. View Article : Google Scholar
|