1. Introduction
Cervical cancer (CC) is the third most common
malignancy among females worldwide, with a global incidence of
>500,000 diagnosed new cases and ~260,000 mortalities each year
(1). Over 85% of the CC global
burden occurs in developing countries, where it represents ~13% of
all types of cancer of female patients. Higher incidence regions
include Eastern and Western Africa (>30 cases/100,000
inhabitants), Southern Africa (26.8/100,000 inhabitants),
South-Central Asia (24.6/100,000 inhabitants), and South America
and Middle Africa (23.9 and 23.0/100,000 inhabitants,
respectively). Cervical cancer remains the most common cancer among
females only in Eastern Africa, South-Central Asia and Melanesia
(1,2). In Latin America, the mortality rates
in the region are seven-fold greater than those in North America,
with an estimated 72,000 new cases of cervical cancer and 33,000
mortalities that occur annually (3).
Conventional treatment of CC patients involves
surgery and chemoradiotherapy, although radical hysterectomy is
employed in early stages. Patients with locally advanced cervical
cancer (LACC) are usually treated with radiotherapy in combination
with cis-platinum (4,5). Nevertheless, ~50% of patients present
with recurrent or persistent disease, which may be explained by the
presence of therapy-resistant cells inside the tumor mass.
Despite advances in our understanding of this
neoplasm, the absence of predictive conventional treatment response
markers and the lack of alternative treatments hamper CC
personalized treatment. Thus, it is imperative to understand the
biology of this tumor, by analyzing its molecular dynamics as well
as the clinical characteristics of CC patients in order to improve
the outcome.
MicroRNAs or miRNAs are small (21–23 nucleotide
long) endogenous, non-coding, single-stranded RNAs that control
gene expression by binding to the 3′ untranslated region (3′UTR) of
messenger RNA (mRNA), leading to mRNA degradation or protein
translation inhibition (6,7). Over 1,000 miRNAs are present in the
human genome, each of which potentially regulates hundreds of
mRNAs. Approximately 60% of all protein-coding genes are
potentially regulated by miRNAs, which confers them a fundamental
role in the modulation of numerous cell processes (8,9).
MicroRNAs with altered expression patterns have been found to have
oncogenic (oncomirs) or tumor-suppressing (anti-oncomirs) functions
present in the pathogenesis of most malignancies. In the canonical
model, oncomirs are upregulated and anti-oncomirs are
downregulated, which has been attributed to amplification, deletion
and/or mutation of miRNA loci, dysregulation of transcription
factors and epigenetic silencing. Globally, miRNA expression
alterations lead to the disturbance of several onco-genic or
tumor-suppressor protein levels, which in turn alter cell growth by
favoring tumor malignancy (10,11).
Over the last decade, the widespread deregulation of
miRNAs in virtually all types of cancer has been clearly
established, as have their implications during each stage of
disease initiation, progression and development (12,13).
Consequently, miRNAs are solid diagnostic and prognostic biomarker
candidates and viable CC therapeutic targets (14,15).
2. miRNAs are associated with cervical
carcinogenesis
Several factors are required for CC development,
including the interaction of viral, environmental and
host-dependent factors, which trigger tumor growth, invasion and
metastasis. In addition, evidence of the importance of epigenetic
regulation mechanisms has steadily increased over the last two
decades, and has focused on the dysregulation of oncogenes and
tumor-suppressor genes as the main generator of the malignant
phenotype. In this regard, miRNAs have an important role as
regulators of cell processes such as apoptosis, cell cycle
progression, metastases and both chemo- and radioresistance
(16). In an effort to understand
the important role of several miRNAs during cervical
carcinogenesis, we conducted an extensive literature review and
found 60 research articles published between 2009 and 2014. These
studies describe >40 deregulated miRNAs that target genes and
are likely to be involved in the development and progression of CC,
as well as in the chemoradiotherapy resistance mechanism (Table I).
 | Table IDifferential expression of miRNAS in
CC vs. normal samples. |
Table I
Differential expression of miRNAS in
CC vs. normal samples.
| miRNA | Status | Target gene | Refs. | miRNA | Status | Target gene | Refs. |
|---|
| miR-155 | U | LKB1 | (22) | miR-944 | U | HECW2/S100PB | (44) |
| miR-196a | U | HOXC8 | (45) | miR-497 | D | IGF-1R | (46) |
| miR-31 | U | ARID1A | (33) | miR-214 | D | BCL2L2 | (37) |
| miR-130a | U | Dicer | (47) | miR-155 | D | EGF | (48) |
| miR-215 | U | Not identified | (49) | miR-303-367 | D | AKT1 | (50) |
| miR-99a/99b | D | mTOR | (51) | miR-424 | D | CHK1 | (52) |
| miR-506 | D | GLI3 | (53) | miR-205 | U | CYR61/CGF | (54) |
| miR-135a | U | SIAH1 | (55) | miR-17-5p | D | TP53INP1 | (56) |
| miR-129-5p | D | SP1 | (57) | miR-10a | U | CHL1 | (58) |
| miR-590 | U | CHL1 | (59) | miR-19a/19b | U | CUL5 | (60) |
| miR-181a | U | PRKCD | (40) | miR-125b | D | PIK3CD | (61) |
| miR-99 | D | TRIB2 | (62) | miR-20a | U | TNKS2 | (63) |
| miR-196a | U | NTN4 | (64) | miR-214 | D | GALNT7 | (65) |
| miR-125b | U | BAK1 | (66) | miR-143 | D | BCL2 | (20) |
| miR-203 | D | VEGFA | (21) | miR-133b | U |
MST2/CDC42/RHOA | (67) |
| miR-196b | D | HOXB7 | (68) | miR-21 | U | CCL20 | (17) |
| miR-7 | D | XIAP | (69) | miR-1 | D | PLK1 | (24) |
| miR-218 | D | Not identified | (70) | miR-375 | D | SP1 | (15) |
| miR-886-5p | U | BAX | (19) | miR-372 | D | CDK2/cyclin A1 | (71) |
| miR-29 | D | YY1/CDK6 | (25) | miR-34a | D | Notch | (18) |
| miR-23b | D | uPA | (72) | miR-519 | D | HUR | (73) |
| miR-214 | D | Plexin-B1 | (74) | miR-21 | U | PDCD4 | (75) |
Evidence supporting the correlation between miRNA
expression and CC-related processes was initially described in 2009
(17). Findings of that study
demonstrated that the expression of miR-21 promoted cell
proliferation in HeLa CC cells, while its inhibition suppressed
cell proliferation by overexpression of the tumor-suppressor gene
PDCD4, a related programmed cell death protein. It also provided
direct evidence that PDCD4 3′UTR is a functional target of this
miRNA. Subsequently, it was demonstrated that miR-21 is a major
oncomir, overexpressed in a wide variety of cancers including CC
(15).
miR-34a, a small molecule, has been described in
studies on cervical carcinogenesis. Pang et al, demonstrated
that it is downregulated in different CC cell lines such as HeLa,
SiHa, C4I, C33a and CaSki, while the induction of its expression in
HeLa cells reduced the invasiveness, affecting Notch and Jagged1
proteins as well as Notch downstream signaling by regulating
urokinase plasminogen activator (uPA) expression. The binding of
miR-34a to the 3′UTR of Notch and Jagged1 was determined using
siRNA functional assays (18).
MicroRNA expression profiling in squamous CC and
adjacent non-tumor tissues showed that miR-886-5p was overexpressed
in CC tissues. Subsequent in vitro assays in human H8
cervical squamous epithelial cell line revealed that this miRNA
depressed BAX protein levels, reducing apoptosis and promoting cell
proliferation. Conversely, knocking down miR-886-5p increased its
pro-apoptotic protein target, BAX, inducing apoptotic cell death
(19).
The same expression profile revealed miR-143
downregu-lation. Functional characterization of this miRNA through
its overexpression in HeLa cells significantly inhibited cell
proliferation and promoted apoptosis, while the co-expression of
anti-miR-143 reestablished the tumor phenotype. Additionally, HeLa
cells transfected with pre-miR-143 were injected subcutaneously
into the flanks of female athymic mice and miR-143 upregulation
suppressed tumor formation. The functional target of this miRNA is
Bcl-2, as shown through functional assays in which miR-143
suppressed the activity of a luciferase reporter carrying the 3′UTR
of the Bcl-2 mRNA. Bcl-2 expression levels in CC tissues were
inversely proportional to the levels of miR-143 (20).
By contrast, miR-203 was consistently downregulated
in both biopsies and tumor cell lines due to hypermethylation of
its promoter region. Functional assays revealed that this miRNA
suppresses proliferation, tumor growth and angiogenesis (21). Lao et al (22) recently demonstrated that miR-155 was
upregulated in CC tissues compared to adjacent non-neoplasic
tissues, and its overexpression in HeLa and SiHa cells promoted
proliferation. By contrast, its downregulation, inhibited cell
cycle progression, promoted apoptosis and induced cell cycle arrest
in those cell lines by directly targeting the LKB1 gene,
which codes for a primary upstream kinase involved in AMPK
activation, cell growth and proliferation suppression. Moreover,
LKB1 was significantly downregulated in CC tissues, suggesting that
this miRNA promoted CC cell prolife ration by regulating LKB1
expression (22).
The aforementioned findings demonstrate that miRNAs
bear an important role in CC tumorigenesis. The main
miRNA-regulated targets participate in mechanisms such as cell
growth, apoptosis, cell cycle arrest and angiogenesis, while it is
widely accepted that deregulation of these cell processes are the
major hallmarks of CC.
3. miRNAs involved in cervical cancer
progression
A key process during CC progression is the
proliferation of differentiating epithelial cells, and the
participation of several miRNAs therein has been investigated. We
conducted an extensive search of the literature, which yielded
information concerning 17 miRNAs associated with the progression of
premalignant lesions to invasive cancer, as identified in 20
articles (Table II).
| Table IImiRNAs involved in cervical cancer
progression. |
Table II
miRNAs involved in cervical cancer
progression.
| miRNA | Status | Cellular
process | Target gene | Clinical
background | Refs. |
|---|
| miR-34a | D | p53-dependent
pathway (cell cycle progression, cellular senescence) | NOTCH, P18Ink4c,
CDK4, CDK6, cyclin A, E2, E2F1, BCL2, BIRC3 | ↓CIN I, ↓↓CIN II,
↓↓↓CIN III | (26,76) |
| miR-218 | D | Focal adhesion | LAMB3 | ↓CIN III,
↓↓↓CaCu | (77) |
| miR-200a,
miR-205 | NC | Metastases (inhibit
the EMT) | ZEB1, ZEB2 and
Sip1 | CaCu → CaCu
metastasis | (78) |
| miR-372 | D | Cell growth
(induces arrest in the S/G2 phases of cell cycle | CDK2, cyclin
A1 | Cervical normal
tissue → cervical cancer tissue | (71) |
| miR-203 | D | Keratinocyte
differentiation/maintenance HPV episomes | p-63-family | Normal epithelia →
HPV-infected epithelia | (79) |
| miR-143 | D | Cell growth and
proliferation | PPAR signaling | ↑Normal, ↓↓CIN,
↓↓CIN III, ↓↓carcinoma | (23) |
| miR-145 | D | Cell motility | IGF-1 | ↑Normal, ↓CIN, ↓CIN
III, ↓carcinoma | (23) |
| miR-99a, miR-513,
miR-29a | D | Cell death, tissue
development | IGF-1, BCL2L2,
VEGFA and CDK6 | ↑Normal, ↓CIN, ↓CIN
III, ↓carcinoma | (23) |
| miR-148a | U | Tumor suppressor
genes | PTEN, P53INP1 and
TP53INP2 | ↑Normal, ↑↑CIN,
↑↑CIN III, ↑↑↑carcinoma | (23) |
| miR-10a, miR-96a,
miR-132 | U | Cell transformation
and progression | HOX genes | ↑Normal, ↑↑↑CIN,
↑↑↑CIN III, ↑↑↑carcinoma | (23) |
| miR-886-5p | U | Cell transformation
and B progression | AX | ↑ANTT, ↑↑↑CSCC | (19) |
| miR-100 | D | Growth, cell cycle
and apoptosis | PLK1 | ↑Normal, ↓CIN,
↓↓carcinoma | (24) |
The differential miRNA expression pattern associated
with CC progression using normal tissues, moderate/severe dysplasia
and invasive squamous cell carcinoma infected with HPV16 sequences
has been identified. The deregulated expression of 9 miRNAs was
detected in cancer and dysplasia compared with normal cervical
tissues. Significantly over-expressed miRNAs were miR-148a,
miR-10a, miR-196a and miR-132; while significantly underexpressed
miRNAs were miR143, miR145, miR-99a, miR-513 and miR-29a. The
findings suggest that these miRNAs can potentially be used to
recognize cervical cancer dysplasia from normal tissue (23).
An interesting study performed by Li et al
(24) evidenced that, miR-100
downregulation plays an important role through loss of inhibition
of its target gene PLK1, a kinase involved in the G2/M transition
during CC development. Authors of that study examined the
expression of miR-100 by RT-qPCR and the PLK1 mRNA and protein by
RT-qPCR and immuno blotting, respectively, in 125 cervical tissues
including normal cervical epithelia, cervical intraepithelial
neoplasia (CIN) and cervical cancer, as well as in five CC cell
lines. miR-100 expression gradually decreased from low-grade to
high-grade CIN and cervical cancer tissues, which correlated with
the upregulated expression of PLK1 in CIN3 and cervical tissues.
Findings of that study also showed that modulating the expression
of miR-100 increased cell proliferation, deregulated the cell cycle
and decreased apoptosis (24).
To determine the relationship of HPV 16-infection
and miRNA expression in CC progression, Li et al (25), employed samples from 18 tissues
obtained from normal, CIN 2–3 and squamous cell carcinoma biopsies
to perform miRNAs microarray analysis; they covered 875 human
miRNAs. Notably, the findings showed 31 unique miRNAs with a
significantly deregulated expression from normal to CC (17 up- and
14 downregulated). Among these, miR-218 was the most significantly
downregulated in the course from normal tissue to CC, while miR-29
was the most overex-pressed. Furthermore, miR-29 showed an
important negative correlation between YY1 and CDK6 expression. The
findings of that suggested that the expression level of miR-29 was
regulated by HR-HPV E6/E7 (25).
Wang et al carried out in vitro assays
that revealed a specific seed match between miR-34a and the 5′UTR
of p18Ink4c, an important modulator of cell cycle progression,
which suppresses its expression. Cervical pre-cancer lesions and
cervical cancer showed an increase of p18Ink4c protein.
Consistently, the immunohistochemical staining of cervical tissue
arrays showed an increased p18Ink4c expression of 4.8% in normal
cervical tissues, of 8.3% in chronic cervical inflammation and of
68% in cervical cancer. Those findings suggested that miR-34a was
downregulated in infected cervical tissues, which is critical to
persistent p18Ink4c activation (26).
Concerning invasion and metastasis, Lee et al
(27) analyzed the expression level
of 157 human mature miRNAs by means of TaqMan quantitative PCR
array. In that study, 10 tumor biopsies staged as primary invasive
squamous cell carcinomas (ISCC) and 10 normal tissues were used.
The results identified significant evidence of 70 miRNAs being
differentially expressed, 68 of which were upregulated and two
downregulated. Of these, 10 miRNAs were significantly
overexpressed, i.e., miR-199-s, miR-9, miR199a*,
miR-199a, miR-199b, miR-145, miR-133a, miR-133b, miR-214 and
miR-127 and only two were underexpressed, i.e., miR-149 and
miR-203. Of note, the expression of miR-127 was significantly
associated with lymph node metastasis. In vitro assays
showed that blocking miR-199a expression exhibited an important
reduction in cell growth (27).
Fig. 1 summarizes the principal
miRNAs involved in cervical carcinogenesis.
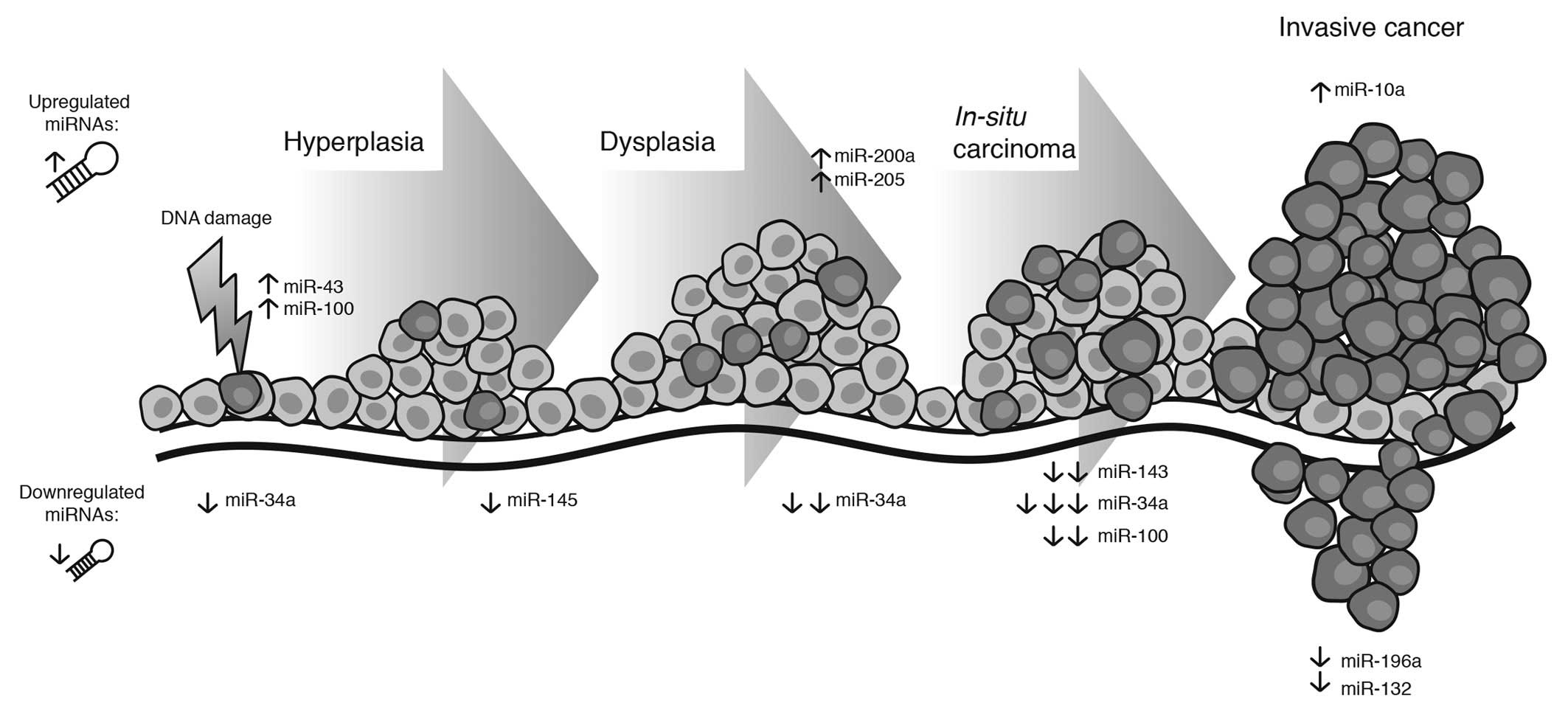 | Figure 1Expression of principal miRNAs
associated with different clinical stages, from normal tissue,
hyperplasia and in situ cancer to invasive cancer. There is
evidence of a number of miRNAs that reduce their expression from
normal tissue to in situ cancer, such as miR-143 and
miR-100, while miR-10a, miR-196a and miR-132 are overexpressed in
invasive cancer. Notably, miR-34 maintains its level through the
evolution of CC. The result of this deregulation alters cell
mechanisms such as cell cycle progression, focal adhesion,
metastasis, cell growth, apoptosis and cell motility. CC, cervical
carcinoma. |
miRNAs associated with complex CC progression have
been investigated. However, known CC-associated miRNAs target
important, well-studied regulators of cell metabolism,
strengthening the significance of their participation in the
evolution of this pathology.
4. miRNAs involved in cervical cancer
clinical outcome
The conventional treatment for LACC patients is
radiotherapy concomitant with cisplatin. However, ~50% of LACC
patients that receive radiotherapy exhibit recurrence or persistent
disease, which may be explained by the presence of radioresistant
cells within the tumor mass. Therefore, radio- and chemoresistance
are major obstacles for an efficient cervical cancer treatment
(4,5). Nonetheless, a meta-analysis showed
that CC patients, treated with radiotherapy alone or in combination
with different chemotherapeutic agents had a 40–70% 5-year survival
rate (28).
In the last years, several miRNAs have been
associated with survival and prognosis of CC patients. Thus, the
expression and regulation of their targets have become molecular
markers with clinical relevance prediction. In this section, we
summarized data from an extensive bibliographical investigation
regarding miRNAs associated with clinical outcome (Table III).
| Table IIImiRNAs involved in cervical cancer
clinical outcome. |
Table III
miRNAs involved in cervical cancer
clinical outcome.
| miRNA | Status | Target gen | Outcome | Function | CC cell type | Refs. |
|---|
| miR-9 | D | Not defined | Poor survival | Invasion and cell
motility | Squamous and
adenocarcinoma cells | (29) |
| miR-93,
miR-200a | U | RECK | Poor survival | Invasion and
lymphatic metastases | Invasive
carcinoma | (32) |
| miR-31 | U | ARID1A | Poor survival | Node metastases,
stromal invasion | Invasive
carcinoma | (33) |
| miR-26a | D | PRL-1 | Poor survival | Inhibits cell
proliferation and invasion | Invasive
carcinoma | (80) |
| miR-224 | U | Not defined | Poor survival | Aggressive
progression | Invasive
carcinoma | (31) |
| Let-7c | D | HMGA2 | Poor survival | Lymph node
metastases | Small cell
carcinoma | (30) |
| miR-100 | D | RSP3, PLK1 | Poor survival | Lymph node
metastases | Small cell
carcinoma | (30) |
| miR-125b | D | BAK1 | Poor survival | Lymph node
metastases | Small cell
carcinoma | (30) |
| miR-143 | D | BCL2, KRAS,
DNMT3A | Poor survival | Lymph node
metastases | Small cell
carcinoma | (30) |
| miR-145 | D | BNIP3, IRS, STAT1,
C-MYC | Poor survival | Lymph node
metastases and poor survival | Small cell
carcinoma | (30) |
| miR-199a-5p | D | SWI, SNF, PAK4 | Poor survival | Lymph node
metastases and poor survival | Small cell
carcinoma | (30) |
For instance, the expression profile of 96
cancer-related miRNAs from 102 CC tumor biopsies was analyzed
(29). Through a mathematical
algorithm, the authors identified that miR-200a and miR-9 were
significantly associated with overall survival (OS). These miRNAS
were individually transfected into HeLa cells and the expression
profile of transfected cells was analyzed. Gene set enrichment and
gene ontology-based analyses showed that miR-200a regulated
ZEB1, ZEB2, TGFB2 and EXOC5 genes
involved in the metastatic potential of cancer cells. Genes
regulated by miR-9 were involved in metabolic processes, explaining
the maintenance of a high metabolic rate by tumor cells, an
important trait of the rapid proliferation of cervical cancer cells
(29).
A similar study published in 2012, analyzed the
expression profiles of 30 miRNAs associated with tumor metastasis
from the formalin-fixed paraffin-embedded samples of 44 SCCC
patients who underwent radical hysterectomy. Seven miRNAs, i.e.,
let-7c, miR-10b, miR-100, miR-125b, miR-143, miR-145 and
miR-199a-5p, were significantly downregulated in the advanced stage
SCCC patients (FIGO IB2-IV) compared to the early stage SCCC
patients (FIGO IB1). Downregulation of these miRNAS, with the
exception of miR-10b, were significantly associated with lymph node
metastasis and with reduced survival in SCCC. Through a survival
analysis, the authors identified that SCCC patients with a low
expression of miR-100 and miR-125b projected a significant tendency
towards a poorer prognosis (30).
Another study reported that the expression of
miR-224 was significantly upregulated in cancer tissues of advanced
FIGO stage cervical cancer patients, in lymph node
metastasis-positive patients and in less-differentiated tumors
(31). Kaplan-Meier analysis showed
that patients with a higher miR-224 expression exhibited a shorter
OS. Additionally, it was associated with aggressive progression and
poor prognosis and was employed as an independent marker for
predicting the clinical outcome (31).
Subsequently, the expression of miR-93 and miR-200a
was retrospectively evaluated in 116 patients with invasive CC and
100 patients undergoing hysterectomy for benign lesions in order to
determine their clinical significance. The levels of miR-93 and
miR-200a were measured by RT-qPCR, and the proteins RECK, MMP2 and
MMP9 were assessed by immunohistochemical staining. Results showed
the upregulation of miR-93, miR-200a, and MMP2 and MMP9 proteins,
but the downregulation of RECK in CC tissues compared to benign
lesion tissues. Notably, patients with a strong RECK expression had
a 5-year survival rate, significantly higher than that of patients
with lower RECK-expressing tumors. In addition, a significant
inverse correlation was identified between RECK downregulation with
invasion and lymphatic metastases. Thus, the expression of miRNAs
and RECK, served as potential prognostic markers for the long-term
survival of CC patients (32).
The role of miR-31 as an independent prognosis
factor was determined and it was demonstrated that it is
upregulated in CC cell lines as well as clinical samples. The high
miR-31 level was significantly correlated with higher FIGO stages,
node metastases, vascular involvement and deep stromal invasion and
with poorer OS. Those results showed that, the downregulation of
miR-31 impaired cell proliferation, colony formation, in
vitro cell migration and invasion, and inhibited in vivo
xenograft tumor growth. The authors verified that ARID1A was a
direct target of miR-31, which was further confirmed by the
inversely correlated expression of miR-31 and ARID1A in patient
specimens. The authors of that study concluded that the
miR-31/ARID1A pathway provides insight into the progression and
clinical outcome of CC (33).
The abovementioned findings suggested that miRNAs
may be used as biomarkers of OS, as they regulate genes involved in
cell processes such as invasion, migration, growth and metastases.
In this sense and to attempt to integrate and comprehend the
complexity of the deregulation and interaction of miRNAs and the
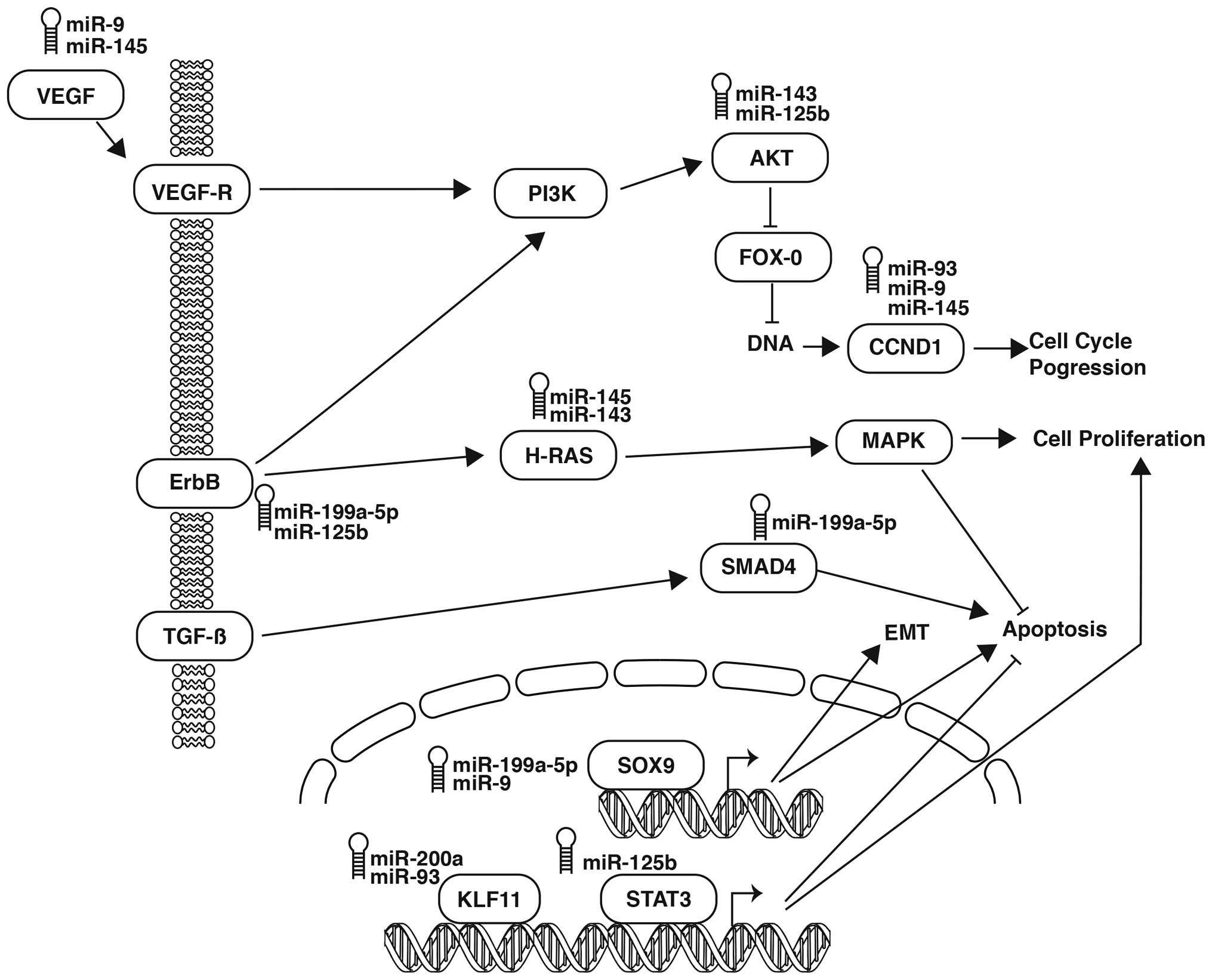
cell processes involved in the clinical outcome, we sketched a
network with the miRNAs and their targets that shows miRNA-mRNA
interaction using 7 of the 12 miRNAs involved in the clinical
outcome, i.e., miR-199a-5p, miR125b, miR-143, miR-145, miR-9, miR93
and miR-200a (Fig. 2). These 7
miRNAs have a direct or indirect association with the regulation of
their target. For example, miR-9 and miR-199a-5p have an indirect
interaction with SOX9, a transcription factor associated with
epithelial-mesenchymal transition (EMT), proliferation and
regulation of apoptosis, while miR-199a-5p and miR-125b putatively
target ERBB, a tyrosine kinase that activated the AKT/PI3K pathway,
indirectly. Another instance is miR-125b, which interacts
indirectly with miR-143 through the regulation of AKT1 and its
migration and cell proliferation pathway. Furthermore, miR-143 and
miR-145 inhibit HRAS secondarily, leading to activation of the MAPK
cascade, progression of the cell cycle and suppression of
apoptosis. VEGF-A, in turn, is downregulated by miR-145 and miR-9,
while miR-9 also acts together with miR-93 and deregulates the
expression of CCND1, a cyclin that activates the cell cycle.
Notably, miR-93 interrelates with another 3 miRNAs: with miR-200a
through the regulation of KLF11, a transcription factor that
participates in cell cycle progression, proliferation and the
inhibition of apoptosis; miR-125b, which possibly targets STAT3, a
transcription factor involved in proliferation and suppression of
apoptosis; and miR-199a-5p, which activates apoptosis through the
underexpression of the tumor suppressor SMAD4.
5. miRNAs involved in the improvement of
cervical cancer treatment response
miRNAs have become important regulators of treatment
response and the understanding of their mechanisms represent
potentially personalized molecular markers for the prediction of
individual clinical outcomes. In addition, miRNAs were employed as
novel therapeutic targets (34,35).
Although they have been studied in different types of cancer, it
has been established that 7 miRNAs were involved in CC treatment
response, two of which were associated with chemoresistance, one
with chemosistance and radioresistance, and the remaining miRNAs
only with radioresistance.
The first study that established the association
between miRNAs with treatment response was published by Shi et
al (36). The aim of their
study was to demonstrate a novel mechanism by which glucocorticoids
modulate p53-dependent miR-145 expression in HPV-positive CC cells
through the induction of E6 proteins. miR-145 expression was
reduced in CC tissues by the action of cortisol, which
simultaneously induced HPV-E6 expression and p53 suppression in CC
cells. Expression of miR-145 in CC cells was wild-type and
p53-dependent, and the cortisol-induced downregulation of miR-145
prevented chemotherapy-induced apoptosis, whereas its
overexpression enhanced sensitivity to mitomycin and reversed the
chemoresistance induced by glucocorticoids. Furthermore, miR-145
enhanced the effects of p53 by suppressing its inhibitors in CC
cells, suggesting that miR-145 plays a role in p53 tumor
suppression and inhibition in the motility and invasion of CC
cells. These findings identified a novel pathway by which the
neuroendocrine macro-environment affected cervical tumor growth,
invasion and therapy resistance, while showing that miR-145 served
as a target for CC therapy (36).
miR-214 is another miRNA that inhibits cell growth,
migration and invasion. Wang et al (37) analyzed its role and function
focusing on demonstrating that increasing levels of miR-214 reduced
cell survival and enhanced cisplatin-induced cytotoxicity in CC
cells. Concordantly, the authors of that study showed that the
ectopic expression of this miRNA reduced the cell survival rate,
induced apoptosis and enhanced sensitivity to cisplatin by directly
inhibiting BCL2L2 expression in HeLa and C-33A cervical cancer
cells. Further analysis revealed that apoptosis was correlated with
increased expression of Bax, caspase-3, -8 and -9. Collectively,
those findings suggested that miR-214 is a potential target for the
development of novel therapeutic strategies (37).
The mechanisms responsible for CC radioresistance
regulated by miRNAs are largely unexplored, nevertheless, Zhang
et al aimed to identify specific miRNAs involved (38). In order to find a specific miRNA
signature, they established radioresistant CC cell variants by
repeated radiation selection. The miRNA profiles of radioresistant
cells and their corresponding controls were analyzed and compared
using microarrays. Among the differentially expressed profiles, 20
miRNAs showed similar patterns of alteration: 14 miRNAs were
overexpressed, while 6 were underexpressed in all three
radioresistant CC cell variants compared to their controls.
miR-630, miR-1246, miR-1290 and miR-3138, exhibited a >5-fold
increase in radioresistant cells. Subsequent analysis revealed that
the four miRNAs could be upregulated in CC cells by radiation
treatment in time- and dose-dependent manners. Ectopic expression
of each miRNA was demonstrated to markedly increase the survival
fraction of irradiated CC cells. Notably, inhibition of miR-630, a
miRNA of the specific signature reversed radioresistance of CC
cells (38).
Ke et al demonstrated that miR-181a had an
important role in radiation therapy (39). The aim of that study was to describe
the roles of miR-181a as a regulator in CC-radioresistant
phenotype, to explore the underlying mechanism and to evaluate the
potential of miR-181a as a radio-sensitivity biomarker. miRNA
profiles of CC tumor specimens were analyzed. The specimens were
sourced from 18 patients with a histological diagnosis of squamous
CC (clinical stage IIIB). The patients had not received any
chemotherapy prior to radiation therapy. A higher expression of
miR-181a was observed in radiation-insensitive CC specimens and
cell lines, when compared to sensitive cancer specimens.
Furthermore, miR-181a negatively regulated the expression of PRKCD,
a pro-apoptotic protein kinase, through interaction with its 3′UTR
messenger, thereby resulting in the inhibition of
irradiation-induced apoptosis and decreasing G2/M blockade. The
role of miR-181a in radioresistance was validated in cell culture
and mouse tumor xenograft models. Cells bearing anti-miRNA were
unable to resist radiation therapy, in contrast to cells expressing
a miR181a mimic. Thus, the expression of miR-181a in CC constitutes
a potential biomarker of sensitivity or response to radiation
therapy and target miR-181a represented a new approach to sensitize
CC to radiation treatment (39).
In addition to its role in radioresistance, miR-181
also confers chemoresistance in CC, according to studies of its
significant upregulation in clinical biopsies from patients that do
not respond to conventional cisplatin treatment (40). To clarify the role of miR-181a in
regulating the chemoresistance of CC, its overexpression was
induced in human cervical squamous cancer cell lines, SiHa and
Me180, which resulted in enhanced chemoresistance to cisplatin
through apoptosis reversion. In a nude mouse xenograft model, the
overexpres-sion of miR-181a markedly inhibited the therapeutic
response to cisplatin in a PRKCD-mediated manner. Additionally,
PRKCD silencing yielded a similar effect to that of miR-181a
upregulation, inhibiting apoptosis in CC cells. Thus, miR-181a may
be used as a biomarker to predict chemosensitivity to cisplatin in
patients with cervical squamous cancer (40).
The studies associated with miRNAs involved clinical
outcome in CC patients. However, few may be applied as biomarkers
of therapy response and as co-adjuvant therapeutic targets. As
therapeutic agents, miRNAs were employed to enhance
chemoradiosensitivity with the downregulation of its
anti-apoptotic, DNA damage repair and cell cycle progression
targets (41–43).
5. Conclusions
Complex diseases such as CC require the conjunction
of different factors. Thus, there are a number of events that lead
an HPV-infected cell to form an invading, chemo- and radioresistant
tumor mass that eventually takes the life of its host. It has been
long established that many of these events can be genetic mutations
that arise due to genome instability and persist due to selection.
In this regard, many deregulated genes have been studied, the
majority of which are associated with apoptosis, cell cycle
control, migration, genetic instability, cell adhesion and
metastasis.
Since miRNA profiles can be used to describe the
chemo-radioresistance of tumors prior to treatment delivery and to
monitor the response through the treatment, miRNA profiles can be
useful in the selection of intensification strategies and predict
final response to therapy and risks of recurrence or metastases. By
contrast, individual miRNAs present an important opportunity to
enhance treatment efficacy, as is the case of miR-181a. This miRNA
was proven to be a chemosensitizer and radiosensitizer, and in both
cases, it downregulates PRKCD, a kinase involved in the regulation
of apoptosis and inhibition of cell growth, making it a reasonable
candidate for miRNA-based therapy enhancement.
The evidence reviewed in the present study showed
that miRNAs affect various biological pathways associated with the
development, progression (CIN 1, 2, 3 and cancer), clinical outcome
and treatment response improvement in CC and reinforce their
relatively recent role as key players in carcino genesis.
Consequently, their use in cervical cancer for diagnosis as well as
clinical outcome prediction and therapy improvement may be
utilized.
Acknowledgments
The present study was submitted by V. G.-Q. in
partial fulfillment of the requirements for the degree of Doctor of
Philosophy (Doctorado en Ciencias Biomedicas), Universidad Nacional
Autonoma de Mexico. It was financially supported in part by the
Universidad Nacional Autonoma de Mexico (UNAM; grant PAPCA-2014-6)
and by Consejo Nacional de Ciencia y Tecnologia (CONACyT; grant
SALUD-2010-01-141907). V. G.-Q. received a CONACyT fellowship.
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics for Hispanics/Latinos, 2012. CA Cancer J Clin.
62:283–298. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar
|
|
3
|
Serrano B, Alemany L, Ruiz PA, Tous S,
Lima MA, Bruni L, Jain A, Clifford GM, Qiao YL, Weiss T, et al:
Potential impact of a 9-valent HPV vaccine in HPV-related cervical
disease in 4 emerging countries (Brazil, Mexico, India and China).
Cancer Epidemiol. 38:748–756. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Landoni F, Maneo A, Colombo A, Placa F,
Milani R, Perego P, Favini G, Ferri L and Mangioni C: Randomised
study of radical surgery versus radiotherapy for stage Ib-IIa
cervical cancer. Lancet. 350:535–540. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Quinn MA, Benedet JL, Odicino F,
Maisonneuve P, Beller U, Creasman WT, Heintz AP, Ngan HY and
Pecorelli S: Carcinoma of the cervix uteri. FIGO 26th Annual Report
on the Results of Treatment in Gynecological Cancer. Int J Gynaecol
Obstet. 95(Suppl 1): S43–S103. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kong YW, Ferland-McCollough D, Jackson TJ
and Bushell M: microRNAs in cancer management. Lancet Oncol.
13:e249–e258. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Suzuki H, Maruyama R, Yamamoto E and Kai
M: Epigenetic alteration and microRNA dysregulation in cancer.
Front Genet. 4:2582013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Friedman RC, Farh KKH, Burge CB and Bartel
DP: Most mammalian mRNAs are conserved targets of microRNAs. Genome
Res. 19:92–105. 2009. View Article : Google Scholar :
|
|
9
|
Ma R, Jiang T and Kang X: Circulating
microRNAs in cancer: Origin, function and application. J Exp Clin
Cancer Res. 31:382012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang B, Pan X, Cobb GP and Anderson TA:
microRNAs as oncogenes and tumor suppressors. Dev Biol. 302:1–12.
2007. View Article : Google Scholar
|
|
11
|
Iorio MV and Croce CM: Causes and
consequences of microRNA dysregulation. Cancer J. 18:215–222. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Agarwal SM, Raghav D, Singh H and Raghava
GPS: CCDB: A curated database of genes involved in cervix cancer.
Nucleic Acids Res. 39:D975–D979. 2011. View Article : Google Scholar :
|
|
14
|
Juan L, Tong HL, Zhang P, Guo G, Wang Z,
Wen X, Dong Z and Tian YP: Identification and characterization of
novel serum microRNA candidates from deep sequencing in cervical
cancer patients. Sci Rep. 4:62772014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang F, Li Y, Zhou J, Xu J, Peng C, Ye F,
Shen Y, Lu W, Wan X and Xie X: miR-375 is down-regulated in
squamous cervical cancer and inhibits cell migration and invasion
via targeting transcription factor SP1. Am J Pathol. 179:2580–2588.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kumar MS, Lu J, Mercer KL, Golub TR and
Jacks T: Impaired microRNA processing enhances cellular
transformation and tumorigenesis. Nat Genet. 39:673–677. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yao T and Lin Z: MiR-21 is involved in
cervical squamous cell tumorigenesis and regulates CCL20. Biochim
Biophys Acta. 1822:248–260. 2012. View Article : Google Scholar
|
|
18
|
Pang RTK, Leung CON, Ye TM, Liu W, Chiu
PC, Lam KK, Lee KF and Yeung WS: MicroRNA-34a suppresses invasion
through downregulation of Notch1 and Jagged1 in cervical carcinoma
and choriocarcinoma cells. Carcinogenesis. 31:1037–1044. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li JH, Xiao X, Zhang YN, Wang YM, Feng LM,
Wu YM and Zhang YX: MicroRNA miR-886-5p inhibits apoptosis by
down-regulating Bax expression in human cervical carcinoma cells.
Gynecol Oncol. 120:145–151. 2011. View Article : Google Scholar
|
|
20
|
Liu L, Yu X, Guo X, Tian Z, Su M, Long Y,
Huang C, Zhou F, Liu M, Wu X, et al: miR-143 is downregulated in
cervical cancer and promotes apoptosis and inhibits tumor formation
by targeting Bcl-2. Mol Med Rep. 5:753–760. 2012.
|
|
21
|
Zhu X, Er K, Mao C, Yan Q, Xu H, Zhang Y,
Zhu J, Cui F, Zhao W and Shi H: miR-203 suppresses tumor growth and
angiogenesis by targeting VEGFA in cervical cancer. Cell Physiol
Biochem. 32:64–73. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lao G, Liu P, Wu Q, Zhang W, Liu Y, Yang L
and Ma C: Mir-155 promotes cervical cancer cell proliferation
through suppression of its target gene LKB1. Tumour Biol.
35:11933–11939. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pereira PM, Marques JP, Soares AR, Carreto
L and Santos MA: MicroRNA expression variability in human cervical
tissues. PLoS One. 5:e117802010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li BH, Zhou JS, Ye F, Cheng XD, Zhou CY,
Lu WG and Xie X: Reduced miR-100 expression in cervical cancer and
precursors and its carcinogenic effect through targeting PLK1
protein. Eur J Cancer. 47:2166–2174. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li Y, Wang F, Xu J, Ye F, Shen Y, Zhou J,
Lu W, Wan X, Ma D and Xie X: Progressive miRNA expression profiles
in cervical carcinogenesis and identification of HPV-related target
genes for miR-29. J Pathol. 224:484–495. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang X, Meyers C, Guo M and Zheng ZM:
Upregulation of p18Ink4c expression by oncogenic HPV E6 via
p53-miR-34a pathway. Int J Cancer. 129:1362–1372. 2011. View Article : Google Scholar :
|
|
27
|
Lee JW, Choi CH, Choi JJ, Park YA, Kim SJ,
Hwang SY, Kim WY, Kim TJ, Lee JH, Kim BG, et al: Altered MicroRNA
expression in cervical carcinomas. Clin Cancer Res. 14:2535–2542.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chemoradiotherapy for Cervical Cancer
Meta-Analysis Collaboration: Reducing uncertainties about the
effects of chemoradiotherapy for cervical cancer: A systematic
review and meta-analysis of individual patient data from 18
randomized trials. J Clin Oncol. 26:5802–5812. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hu X, Schwarz JK, Lewis JS Jr, Huettner
PC, Rader JS, Deasy JO, Grigsby PW and Wang X: A microRNA
expression signature for cervical cancer prognosis. Cancer Res.
70:1441–1448. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Huang L, Lin JX, Yu YH, Zhang MY, Wang HY
and Zheng M: Downregulation of six microRNAs is associated with
advanced stage, lymph node metastasis and poor prognosis in small
cell carcinoma of the cervix. PLoS One. 7:e337622012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shen SN, Wang LF, Jia YF, Hao YQ, Zhang L
and Wang H: Upregulation of microRNA-224 is associated with
aggressive progression and poor prognosis in human cervical cancer.
Diagn Pathol. 8:692013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang L, Wang Q, Li HL and Han LY:
Expression of MiR200a, miR93, metastasis-related gene RECK and
MMP2/MMP9 in human cervical carcinoma - relationship with
prognosis. Asian Pac J Cancer Prev. 14:2113–2118. 2013. View Article : Google Scholar
|
|
33
|
Wang N, Zhou Y, Zheng L and Li H: MiR-31
is an independent prognostic factor and functions as an oncomir in
cervical cancer via targeting ARID1A. Gynecol Oncol. 134:129–137.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hu A, Huang JJ, Xu WH, Jin XJ, Li JP, Tang
YJ, Huang XF, Cui HJ and Sun GB: miR-21 and miR-375 microRNAs as
candidate diagnostic biomarkers in squamous cell carcinoma of the
larynx: Association with patient survival. Am J Transl Res.
6:604–613. 2014.PubMed/NCBI
|
|
35
|
Miller P, Clarke J, Koru-Sengul T,
Brinkman J and El-Ashry D: A novel MAPK-microRNA signature is
predictive of hormone-therapy resistance and poor outcome in
ER-positive breast cancer. Clin Cancer Res. 21:373–385. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shi M, Du L, Liu D, Qian L, Hu M, Yu M,
Yang Z, Zhao M, Chen C, Guo L, et al: Glucocorticoid regulation of
a novel HPV-E6-p53-miR-145 pathway modulates invasion and therapy
resistance of cervical cancer cells. J Pathol. 228:148–157. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang F, Liu M, Li X and Tang H: MiR-214
reduces cell survival and enhances cisplatin-induced cytotoxicity
via down-regulation of Bcl2l2 in cervical cancer cells. FEBS Lett.
587:488–495. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang B, Chen J, Ren Z, Chen Y, Li J, Miao
X, Song Y, Zhao T, Li Y, Shi Y, et al: A specific miRNA signature
promotes radioresistance of human cervical cancer cells. Cancer
Cell Int. 13:1182013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ke G, Liang L, Yang JM, Huang X, Han D,
Huang S, Zhao Y, Zha R, He X and Wu X: MiR-181a confers resistance
of cervical cancer to radiation therapy through targeting the
pro-apoptotic PRKCD gene. Oncogene. 32:3019–3027. 2013. View Article : Google Scholar
|
|
40
|
Chen Y, Ke G, Han D, Liang S, Yang G and
Wu X: MicroRNA-181a enhances the chemoresistance of human cervical
squamous cell carcinoma to cisplatin by targeting PRKCD. Exp Cell
Res. 320:12–20. 2014. View Article : Google Scholar
|
|
41
|
Fang L, Li H, Wang L, Hu J, Jin T, Wang J
and Yang BB: MicroRNA-17-5p promotes chemotherapeutic drug
resistance and tumour metastasis of colorectal cancer by repressing
PTEN expression. Oncotarget. 5:2974–2987. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lee KM, Choi EJ and Kim IA: microRNA-7
increases radiosen-sitivity of human cancer cells with activated
EGFR-associated signaling. Radiother Oncol. 101:171–176. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yang SM, Huang C, Li XF, Yu MZ, He Y and
Li J: miR-21 confers cisplatin resistance in gastric cancer cells
by regulating PTEN. Toxicology. 306:162–168. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Xie H, Lee L, Scicluna P, Kavak E, Larsson
C, Sandberg R and Lui WO: Novel functions and targets of miR-944 in
human cervical cancer cells. Int J Cancer. 136:E230–E241. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Villegas-Ruiz V, Juárez-Méndez S,
Pérez-González OA, Arreola H, Paniagua-García L, Parra-Melquiadez
M, Peralta-Rodríguez R, López-Romero R, Monroy-García A,
Mantilla-Morales A, et al: Heterogeneity of microRNAs expression in
cervical cancer cells: Over-expression of miR-196a. Int J Clin Exp
Pathol. 7:1389–1401. 2014.PubMed/NCBI
|
|
46
|
Luo M, Shen D, Zhou X, Chen X and Wang W:
MicroRNA-497 is a potential prognostic marker in human cervical
cancer and functions as a tumor suppressor by targeting the
insulin-like growth factor 1 receptor. Surgery. 153:836–847. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
He L, Wang HY, Zhang L, Huang L, Li JD,
Xiong Y, Zhang MY, Jia WH, Yun JP, Luo RZ, et al: Prognostic
significance of low DICER expression regulated by miR-130a in
cervical cancer. Cell Death Dis. 5:e12052014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lei C, Wang Y, Huang Y, Yu H, Huang Y, Wu
L and Huang L: Up-regulated miR155 reverses the
epithelial-mesenchymal transition induced by EGF and increases
chemo-sensitivity to cisplatin in human Caski cervical cancer
cells. PLoS One. 7:e523102012. View Article : Google Scholar
|
|
49
|
Liang H, Li Y, Luo RY and Shen FJ:
MicroRNA-215 is a potential prognostic marker for cervical cancer.
J Huazhong Univ Sci Technolog Med Sci. 34:207–212. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Cai N, Wang YD and Zheng PS: The
microRNA-302-367 cluster suppresses the proliferation of cervical
carcinoma cells through the novel target AKT1. RNA. 19:85–95. 2013.
View Article : Google Scholar :
|
|
51
|
Wang L, Chang L, Li Z, Gao Q, Cai D, Tian
Y, Zeng L and Li M: miR-99a and -99b inhibit cervical cancer cell
proliferation and invasion by targeting mTOR signaling pathway. Med
Oncol. 31:9342014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Xu J, Li Y, Wang F, Wang X, Cheng B, Ye F,
Xie X, Zhou C and Lu W: Suppressed miR-424 expression via
upregulation of target gene Chk1 contributes to the progression of
cervical cancer. Oncogene. 32:976–987. 2013. View Article : Google Scholar
|
|
53
|
Wen SY, Lin Y, Yu YQ, Cao SJ, Zhang R,
Yang XM, Li J, Zhang YL, Wang YH, Ma MZ, et al: miR-506 acts as a
tumor suppressor by directly targeting the hedgehog pathway
transcription factor Gli3 in human cervical cancer. Oncogene.
34:717–725. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Xie H, Zhao Y, Caramuta S, Larsson C and
Lui WO: miR-205 expression promotes cell proliferation and
migration of human cervical cancer cells. PLoS One. 7:e469902012.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Leung CON, Deng W, Ye T-M, Ngan HY, Tsao
SW, Cheung AN, Pang RT and Yeung WS: miR-135a leads to cervical
cancer cell transformation through regulation of β-catenin via a
SIAH1-dependent ubiquitin proteosomal pathway. Carcinogenesis.
35:1931–1940. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Wei Q, Li Y-X, Liu M, Li X and Tang H:
MiR-17-5p targets TP53INP1 and regulates cell proliferation and
apoptosis of cervical cancer cells. IUBMB Life. 64:697–704. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhang J, Li S, Yan Q, Chen X, Yang Y, Liu
X and Wan X: Interferon-β induced microRNA-129-5p down-regulates
HPV-18 E6 and E7 viral gene expression by targeting SP1 in cervical
cancer cells. PLoS One. 8:e813662013. View Article : Google Scholar
|
|
58
|
Long MJ, Wu FX, Li P, Liu M, Li X and Tang
H: MicroRNA-10a targets CHL1 and promotes cell growth, migration
and invasion in human cervical cancer cells. Cancer Lett.
324:186–196. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Chu Y, Ouyang Y, Wang F, Zheng A, Bai L,
Han L, Chen Y and Wang H: MicroRNA-590 promotes cervical cancer
cell growth and invasion by targeting CHL1. J Cell Biochem.
115:847–853. 2014. View Article : Google Scholar
|
|
60
|
Xu XM, Wang XB, Chen MM, Liu T, Li YX, Jia
WH, Liu M, Li X and Tang H: MicroRNA-19a and -19b regulate cervical
carcinoma cell proliferation and invasion by targeting CUL5. Cancer
Lett. 322:148–158. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Cui F, Li X, Zhu X, Huang L, Huang Y, Mao
C, Yan Q, Zhu J, Zhao W and Shi H: MiR-125b inhibits tumor growth
and promotes apoptosis of cervical cancer cells by targeting
phosphoinositide 3-kinase catalytic subunit delta. Cell Physiol
Biochem. 30:1310–1318. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Xin JX, Yue Z, Zhang S, Jiang ZH, Wang PY,
Li YJ, Pang M and Xie SY: miR-99 inhibits cervical carcinoma cell
proliferation by targeting TRIB2. Oncol Lett. 6:1025–1030.
2013.PubMed/NCBI
|
|
63
|
Kang HW, Wang F, Wei Q, Zhao YF, Liu M, Li
X and Tang H: miR-20a promotes migration and invasion by regulating
TNKS2 in human cervical cancer cells. FEBS Lett. 586:897–904. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Zhang J, Zheng F, Yu G, Yin Y and Lu Q:
miR-196a targets netrin 4 and regulates cell proliferation and
migration of cervical cancer cells. Biochem Biophys Res Commun.
440:582–588. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Peng RQ, Wan HY, Li HF, Liu M, Li X and
Tang H: MicroRNA-214 suppresses growth and invasiveness of cervical
cancer cells by targeting
UDP-N-acetyl-α-D-galactosamine:polypeptide
N-acetylgalactosaminyltransferase 7. J Biol Chem. 287:14301–14309.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Wang YD, Cai N, Wu XL, Cao H-Z, Xie LL and
Zheng PS: OCT4 promotes tumorigenesis and inhibits apoptosis of
cervical cancer cells by miR-125b/BAK1 pathway. Cell Death Dis.
4:e7602013. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Qin W, Dong P, Ma C, Mitchelson K, Deng T,
Zhang L, Sun Y, Feng X, Ding Y, Lu X, et al: MicroRNA-133b is a key
promoter of cervical carcinoma development through the activation
of the ERK and AKT1 pathways. Oncogene. 31:4067–4075. 2012.
View Article : Google Scholar
|
|
68
|
How C, Hui ABY, Alajez NM, Shi W, Boutros
PC, Clarke BA, Yan R, Pintilie M, Fyles A, Hedley DW, et al:
MicroRNA-196b regulates the homeobox B7-vascular endothelial growth
factor axis in cervical cancer. PLoS One. 8:e678462013. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Liu S, Zhang P, Chen Z, Liu M, Li X and
Tang H: MicroRNA-7 downregulates XIAP expression to suppress cell
growth and promote apoptosis in cervical cancer cells. FEBS Lett.
587:2247–2253. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Yamamoto N, Kinoshita T, Nohata N, Itesako
T, Yoshino H, Enokida H, Nakagawa M, Shozu M and Seki N: Tumor
suppressive microRNA-218 inhibits cancer cell migration and
invasion by targeting focal adhesion pathways in cervical squamous
cell carcinoma. Int J Oncol. 42:1523–1532. 2013.PubMed/NCBI
|
|
71
|
Tian RQ, Wang XH, Hou LJ, Jia WH, Yang Q,
Li YX, Liu M, Li X and Tang H: MicroRNA-372 is down-regulated and
targets cyclin-dependent kinase 2 (CDK2) and cyclin A1 in human
cervical cancer, which may contribute to tumorigenesis. J Biol
Chem. 286:25556–25563. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Au Yeung CL, Tsang TY, Yau PL and Kwok TT:
Human papillomavirus type 16 E6 induces cervical cancer cell
migration through the p53/microRNA-23b/urokinase-type plasminogen
activator pathway. Oncogene. 30:2401–2410. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Abdelmohsen K, Kim MM, Srikantan S,
Mercken EM, Brennan SE, Wilson GM, Cabo R and Gorospe M: miR-519
suppresses tumor growth by reducing HuR levels. Cell Cycle.
9:1354–1359. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Qiang R, Wang F, Shi LY, Liu M, Chen S,
Wan HY, Li YX, Li X, Gao SY and Sun BC: Plexin-B1 is a target of
miR-214 in cervical cancer and promotes the growth and invasion of
HeLa cells. Int J Biochem Cell Biol. 43:632–641. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Yao Q, Xu H, Zhang QQ, Zhou H and Qu LH:
MicroRNA-21 promotes cell proliferation and down-regulates the
expression of programmed cell death 4 (PDCD4) in HeLa cervical
carcinoma cells. Biochem Biophys Res Commun. 388:539–542. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Li B, Hu Y, Ye F, Li Y, Lv W and Xie X:
Reduced miR-34a expression in normal cervical tissues and cervical
lesions with high-risk human papillomavirus infection. Int J
Gynecol Cancer. 20:597–604. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Martinez I, Gardiner AS, Board KF, Monzon
FA, Edwards RP and Khan SA: Human papillomavirus type 16 reduces
the expression of microRNA-218 in cervical carcinoma cells.
Oncogene. 27:2575–2582. 2008. View Article : Google Scholar :
|
|
78
|
Gregory PA, Bert AG, Paterson EL, Barry
SC, Tsykin A, Farshid G, Vadas MA, Khew-Goodall Y and Goodall GJ:
The miR-200 family and miR-205 regulate epithelial to mesenchymal
transition by targeting ZEB1 and SIP1. Nat Cell Biol. 10:593–601.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Melar-New M and Laimins LA: Human
papillomaviruses modulate expression of microRNA 203 upon
epithelial differentiation to control levels of p63 proteins. J
Virol. 84:5212–5221. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Dong J, Sui L, Wang Q, Chen M and Sun H:
MicroRNA-26a inhibits cell proliferation and invasion of cervical
cancer cells by targeting protein tyrosine phosphatase type IVA 1.
Mol Med Rep. 10:1426–1432. 2014.PubMed/NCBI
|