Introduction
Recently, energy metabolism has been considered an
attractive target for antitumoral therapies, since metabolic
changes are a common feature of cancerous tissues. Multiple
molecular mechanisms, both intrinsic and extrinsic, converge to
alter cellular metabolism. The best characterized metabolic
phenotype observed in tumor cells is the Warburg effect, which
involves ATP generation through glycolysis instead of oxidative
phosphorylation (OXPHOS), even under normal oxygen concentrations
(1). However, metabolic adaptation
in tumors extends beyond the Warburg effect. In fact, the classical
theory on the metabolism of cancer cells (increased glycolytic
activity and downregulation of OXPHOS) is still under investigation
since different studies have shown that cancer cells can live in a
wide spectrum of states ranging from the predominance of the
glycolytic phenotype to a phosphorylative phenotype (2). Complete catabolism of pyruvate to
CO2 may be counterproductive in a dividing cell as it
may limit the availability of precursors necessary for cell
replication, e.g., sugars for nucleotide biosynthesis.
In normal cells, glucose is catabolized to pyruvate,
which can be later converted to acetyl-CoA to fuel the
tricarboxylic acid cycle. On the other hand, the manner by which
tumor cells process pyruvate, the end product of glycolysis, is
also different from normal cells, since pyruvate-to-lactate is a
preferred reaction in tumor cells (3). Therefore, in cancer cells, the
glycolytic flux may be blocked at pyruvate production by pyruvate
kinase. In this case, pyruvate is no longer oxidized in
mitochondria and is metabolized by cytosolic lactate dehydrogenase.
Neverthess, a portion of pyruvate is imported into mitochondria and
it still might be metabolized through a mitochondrial oxidative
phosphorylative pathway (OXPHOS), creating the partial OXPHOS
phenotype.
Differences in OXPHOS status originate from
variability of metabolic reprogramming among cancer cells and from
the relative contributions of different oncogenes, tumor
environment, and mitochondrial function. Disruption of cancer cell
metabolism represents an elegant approach to induce tumoral cell
death; indeed cell death and metabolic alteration are closely
related. The strategy to target tumor metabolism requires an
integrated vision of this process since it is not a single
metabolic step that is altered; but the entire energetic metabolism
working in a pattern is profoundly affected with respect to normal
cells. In addition, the metabolic alterations and adaptations of
cancer cells create a particular phenotype essential for tumor cell
growth and survival that is particular for each type of cancer.
Therefore, it may help to understand how, step by step, the
metabolic pathways are arranged in comparison with normal
metabolism to characterize a cancer metabolic phenotype.
Several drugs could be referred to as mitocans,
metabo-cans, or aberrocans and many are being developed at present
(4). Recently, several studies have
documented the ability of chemopreventive phytochemicals to
increase the sensitivity of tumoral cells to anticancer drugs
(5,6). Bergapten (5-methoxypsoralen, Bg) is a
psoralen or taxol (also known as furocoumarins) found in bergamot
essential oil and in other citrus essential oils. It belongs to the
flavonoid class, which has been found to exhibit a variety of
biological activities, usually associated with low toxicity.
Flavonoids have attracted considerable interest because of their
diverse pharmacological properties. Bg activity against different
types of tumor including prostate, ovarian, lung cancer and breast
cancer has been observed (7).
It is generally believed that the antitumor effect
of Bg is due to its interference with the normal function of
microtubule-inducing arrest of the cell cycle (7). However, studies have shown that
taxol-induced cell death is via multiple signaling pathways
(8–11). For example, taxol-induced apoptosis
in breast cancer cells requires downregulation of IκBα, which in
turn activates NF-κB (10).
Activation of MAPK pathways by taxol in HeLa cells was found to
lead to a time-dependent increase in poly(ADP-ribose) polymerase
cleavage and apoptosis (11). A
study of differential gene expression in mouse cells treated with
taxol identified a number of genes that modulate apoptosis
(9). We previously demonstrated
that in breast cancer cells, bergapten (Bg) treatment affects the
PI3K/AKT survival pathway, inducing apoptosis and lowering cell
proliferation (12). In addition,
we found that Bg enhanced p53 gene expression inducing apoptosis in
breast cancer cells (13).
Herein, we report a new anticancer effect of Bg on
human breast cancer, since it was able to interfere in cancer
reprogrammed metabolism. We characterized the basal metabolic
profile of MCF7 and ZR75 cells and evaluated the effect of Bg on
different metabolic pathways. Moreover, we present various
quantitative aspects of human breast cancer cell metabolism which
are limited by the lack of available data at the level of
metabolite amounts and enzymatic activities. The quantification can
yield multiple insights into the organization of the cancer
metabolic network.
Materials and methods
Materials
Methoxypsoralen or Bg, aprotinin, leupeptin,
phenylmethylsulfonyl fluoride (PMFS), sodium orthovanadate,
Tris-Cl, MnCl2, NADP+, isocitrate and malate
were obtained from Sigma-Aldrich (Milan, Italy). Antibodies used in
this study were from Santa Cruz Biotechnology (Santa Cruz, CA,
USA). MitoProfile® Total OXPHOS WB Antibody Cocktail was
from Abcam (Milan, Italy). Triglycerides, lipase activity,
glucose-6-phosphate dehydrogenase (G6PDH) activity, LDH activity
and glucose assay kits were from Inter-Medical (Biogemina Italia
Srl, Catania, Italy). Molecular Probes' ATP Determination kit
(A22066) was from Invitrogen (Milan, Italy). Bg was dissolved in
ethanol (0.02% final concentration) which was used as a solvent
control and did not induce any positive results in all in
vitro assays (data not shown).
Cell culture
Human breast cancer MCF7 and ZR75-1 (ZR75) cells
(American Type Culture Collection, ATCC) were stored according to
supplier's instructions. Every 4 months, cells were authenticated
by single tandem repeat analysis at our Sequencing Core;
morphology, doubling times, estrogen sensitivity, and mycoplasma
negativity were tested (MycoAlert; Lonza). Both cell lines were
maintained in DMEM/F-12 medium containing 5% fetal calf serum
(FCS), 1% L-glutamine, 1% Eagle's non-essential amino acids and 1
mg/ml penicillin/streptomycin in a 5% CO2 humidified
atmosphere. Cells cultured in phenol red and serum-free medium for
24 h, were treated with Bg in medium containing 5% charcoal-treated
FCS. The Bg concentrations were chosen on the basis of our previous
studies (12,13). In addition, we found that Bg induces
apoptosis at 24 h, therefore we hypothesized that a metabolic
disruption may occur in a shorter time period, thus we treated the
cells for 6 and 16 h (12).
Western blotting
Total protein extracts were obtained as previously
described (14). Proteins were
resolved on a 10% sodium dodecyl sulfate-polyacrylamide gel,
transferred to a nitrocellulose membrane, and probed overnight at
4°C with the indicated antibodies. β-actin was used as loading
control.
Glucose assay
Glucose oxidase catalyzes the oxidation of glucose
to gluconic acid. The formed hydrogen peroxide is detected by a
chromogenic oxygen acceptor, phenol, 4-amino-phenazone in the
presence of peroxidase. The intensity of the color formed is
proportional to the glucose concentration in the sample (14). Data are presented as nM/mg
protein.
LDH assay
LDH catalyzes the interconversion of pyruvate and
lactate with concomitant interconversion of NADH and
NAD+. After the treatments, LDH assay was performed on
cell lysates as previously described (13). Data are presented as absorbance
change at 340 nm.
Triglyceride assay
Triglycerides were measured in duplicate by a
GPO-POD enzymatic colorimetric method according to the
manufacturer's instructions in cell lysates and as previously
described (16). Data are presented
as nM/mg protein.
Lipase activity assay
Lipase activity was evaluated by the method of
Panteghini et al (17) based
on the use of 1,2-o-dilauryl-rac-glycero-3-glutaric
acid-(6′-methylresorufin) ester as substrate, as previously
described (18). Data are presented
as nM/min/mg protein.
Assay of the G6PDH activity
The conversion of NADP+ to NADPH,
catalyzed by G6PDH, was measured by the increase in absorbance at
340 nm as previously described (16,18).
Data are presented as nM/min/mg protein.
ATP assay
A bioluminescence assay for quantitative
determination of ATP with recombinant firefly luciferase and its
substrate D-luciferin (light emission at 560 nm at pH 7.8), was
performed as previously described (19). Data are presented as nM/mg
protein.
Isocitrate dehydrogenase (ICD) activity
assay
ICD activity was measured as in Gnoni and
Paglialonga (20). The reaction was
performed on the cell lysate using a final 1-ml reaction volume
containing 0.2 mg protein, 50 mM Tris-Cl pH 7.4, 5 mM
MnCl2, 0.25 mM NADP+ and 0.25 mM isocitrate,
at 37°C. Reaction progress was monitored at 340 nm for 2 min; the
NADPH production was calculated using an NADPH extinction
coefficient of 6.26103 M−1 cm−1. Data are
presented as nM/min/mg protein.
Malic enzyme (ME) activity assay
ME activity was measured in the cell lysates as in
Gnoni and Paglialonga (20). The
reaction was performed using a final 1-ml reaction volume
containing 0.3 mg protein, 50 mM Tris-HCl pH 7.4, 5 mM
MnCl2, 0.1 mM NADP+ and 5 mM malate, at room
temperature. Reaction progress was monitored at 340 nm for 2 min;
the NADPH production was calculated using an NADPH extinction
coefficient of 6.26103 M−1 cm−1. Data are
presented as nM/min/mg protein.
Trypan blue assay
Cells were treated as indicated and the loss of
survival was assessed by the trypan blue assay as previously
described (14).
Statistical analysis
Each datum point represents the mean ± SEM of the
experimental values as indicated in the figure legends. Data were
analyzed by the Student's t-test using the GraphPad Prism 4
software program. P<0.05 was considered to indicate a
statistically significant result.
Results
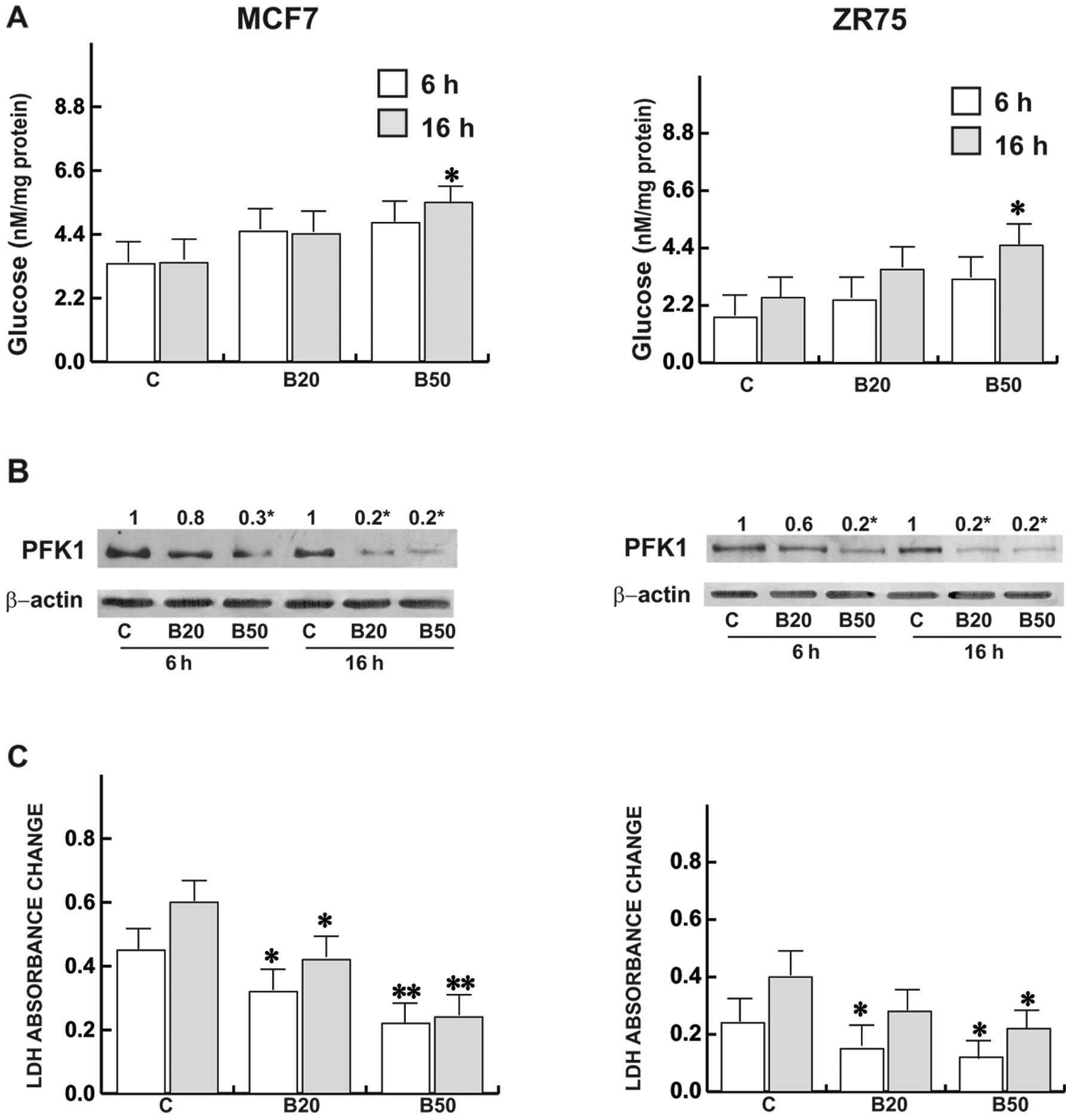
Bg interferes with the glycolysis in MCF7
and ZR75 cells
To characterize the basal metabolic phenotype of
both breast cancer cell lines as well as following Bg treatment, we
first determined the glucose content and analyzed various key
enzymes involved in the carbohydrate metabolism:
6-phos-phofructo-1-kinase (PFK1) expression and the LDH activity.
Bg produced a slight glucose accumulation although it was
significant following treatment with Bg at 50 µM at 16 h in
both cell types (Fig. 1A). The
cells had decreased glucose utilization after 16 h, and this
commensurated with inhibition of cell viability as reported in
previous studies (12,13); while a drastic reduction in the
glycolytic enzyme PFK1 was observed following treatment with Bg at
20 and 50 µM (Fig. 1B). In
the same experimental conditions, lactate production significantly
decreased (Fig. 1C), both at 6 and
16 h. Noteworthy, ZR75 cells possessed lower basal LDH activity as
well as glucose content with respect to the MCF7 cells.
Effects of Bg on biosynthetic
contributions in MCF7 and ZR75 cells
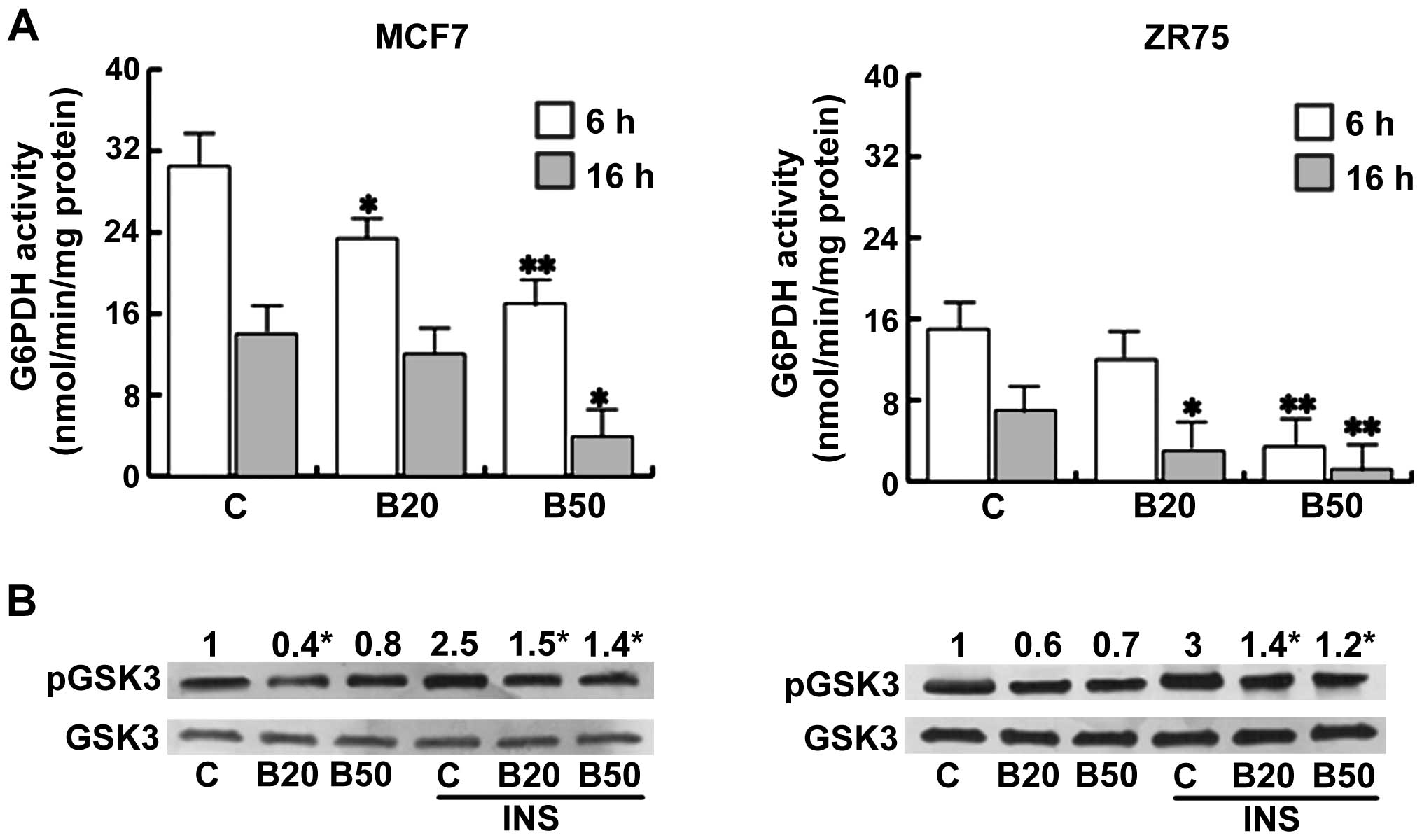
Another manner of glucose utilization is via the
pentose phosphate cycle (PPP), through G6PDH activity. Bg treatment
induced a significant reduction in the enzymatic activity (Fig. 2A), indicating how the decrease in
G6PDH in treated cells contributes to the decrease in glucose
utilization. Interestingly, ZR75 cells possessed a lower basal
G6PDH activity with respect to the MCF7 cells. Glycogen represents
an alternative energy source in tumors expressing active glycogen
synthase (20). Phosphorylation of
glycogen synthase by GSK3 phosphorylated on Ser9 inhibits glycogen
synthesis. In the present study, Bg was able to reduce this GSK3
phosphorylation, implying a blockade on the glycogen synthase
activity. GSK3 has been initially identified as a key regulator of
insulin-dependent glycogen synthesis and we found that Bg
antagonized the upregulatory effect elicited by insulin on this
enzyme (Fig. 2B).
Therefore, we decided to further investigate the
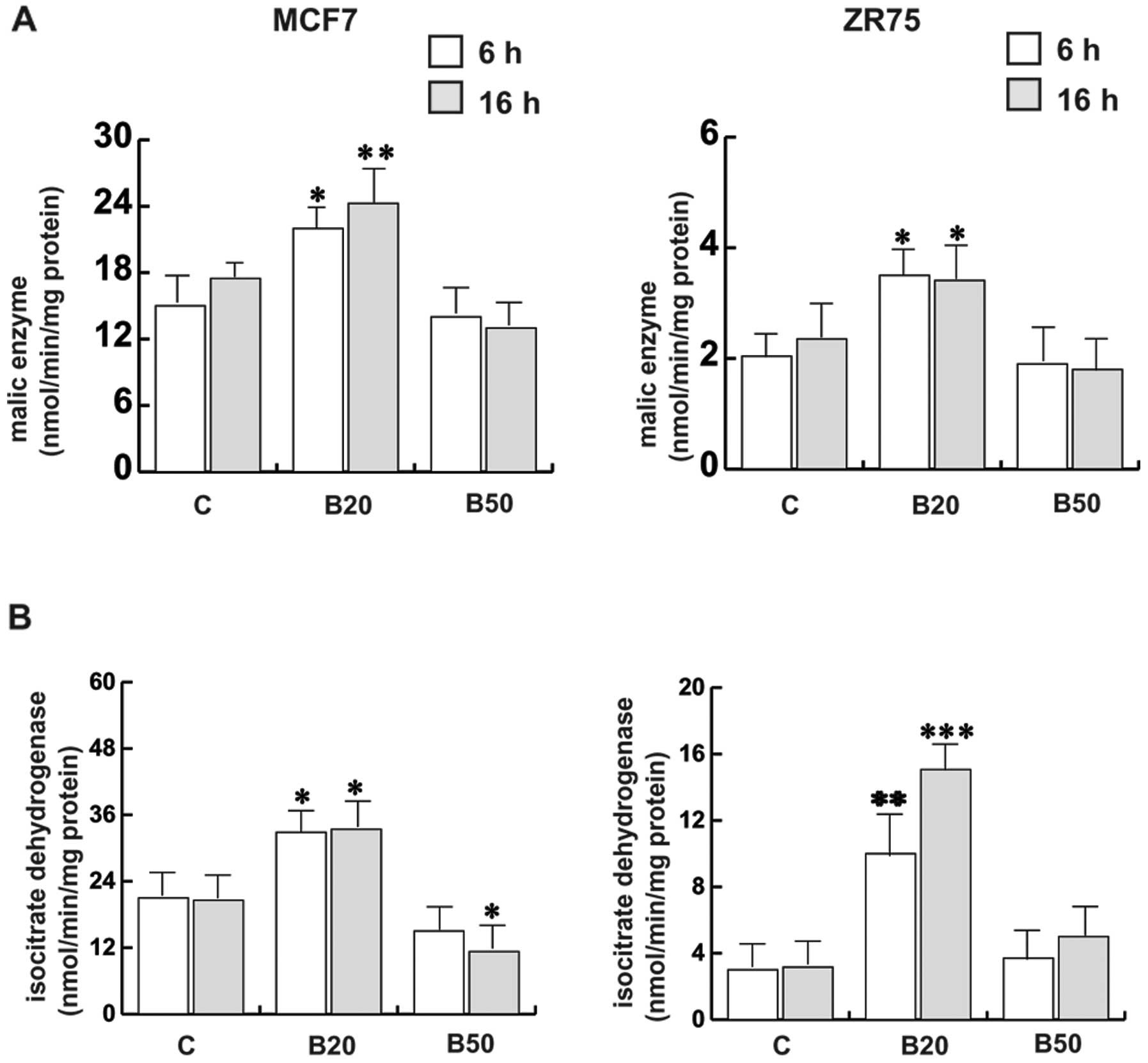
effect of Bg on the bioenergetic balance of breast cancer cells.
Two enzymes involved in the production of NADPH other than PPP, ME
and ICDH, were studied. In MCF7 cells, (Fig. 3A and B), treatment of Bg at 20
µM was able to induce the activities of both enzymes, while
treatment with Bg at 50 µM reduced ME activity although not
significantly; the extension of treatment with Bg at 50 µM
for up to 16 h induced a significant reduction in ICDH activity. A
similar pattern of response was obtained in ZR75 cells upon
treatment of Bg at 20 µM; however, following treatment with
Bg at 50 µM, the enzymatic activities were close to that of
the controls. Notably, ZR75 cells possessed lower basal enzymatic
activities with respect to the MCF7 cells.
Bg modulates bioenergetic requirements in
MCF7 and ZR75 cells
Successively, we focused our investigation on
OXPHOS, by analyzing the mitochondrial OXPHOS components, the ATP
content and the AMP-activated protein kinase (AMPK) expression.
Some of the OXPHOS components were not affected by
Bg treatment in the MCF7 cells, except for Complex (C) IV that
resulted in its upregulation, while CI was reduced particularly at
16 h (Fig. 4A and B). These data,
as expected, resulted in a decrease in ATP production (Fig. 4C). Concerning ZR75 cells, the OXPHOS
protein pattern at basal conditions and under Bg was expressed to a
greater extent with respect to the MCF7 cells, although starting
from the same protein loading. Even when the CIV component was
poorly expressed with respect to the other components in ZR75
cells, it ensured the flow of electrons along the other components,
that were overexpressed (including the CII) maintaining a high ATP
synthesis, as shown in Fig. 4C. The
CIV component of the mitochondrial respiratory chain was expressed
to a lesser extent in the ZR75 cells with respect to the MCF7
cells.
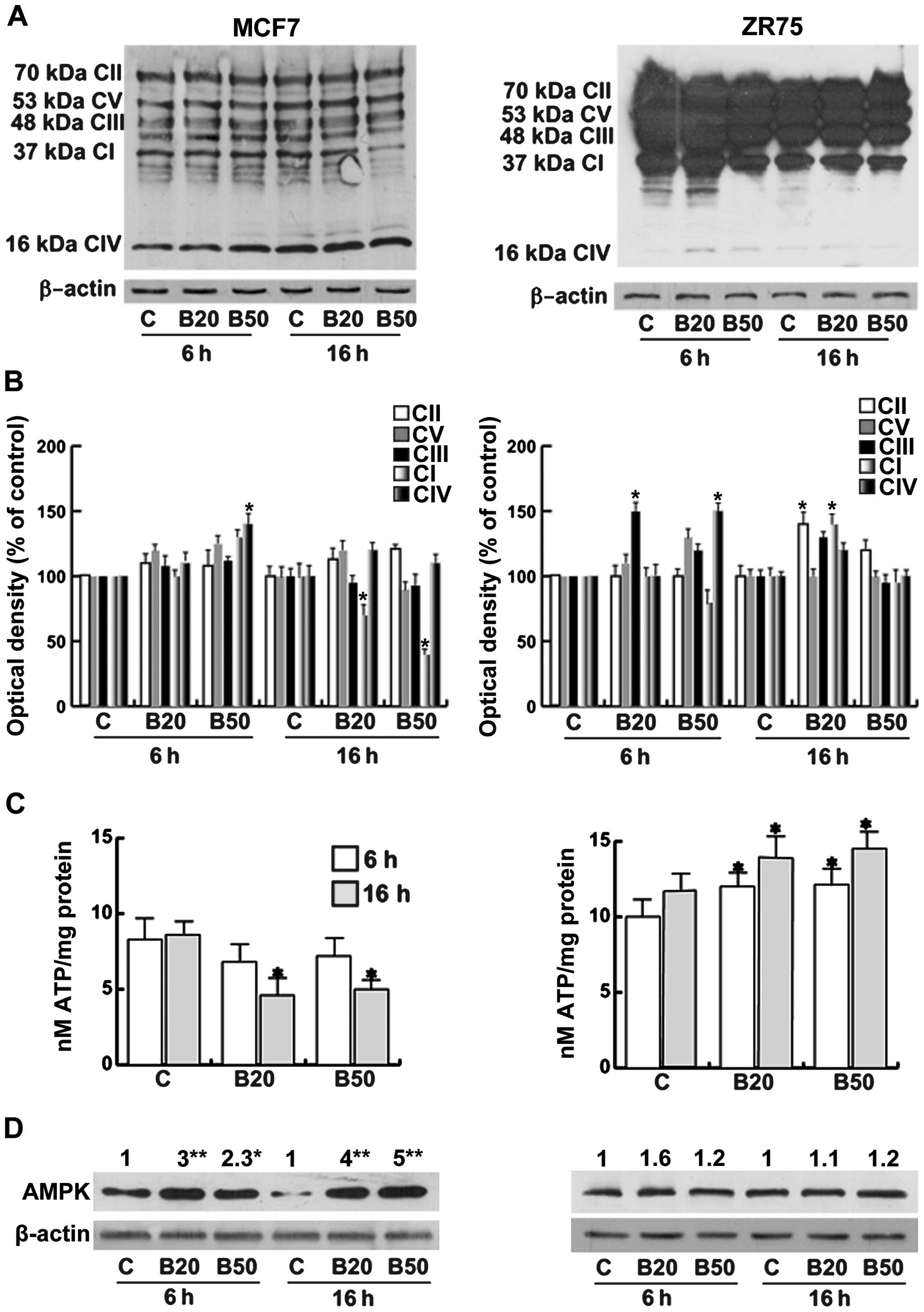 | Figure 4Bg alters the bioenergetics in breast
cancer cells. (A) Western blot assay of OXPHOS in both cell lines
in the absence (C) or with increasing Bg concentrations (B20
µM and B50 µM) at 6 and 16 h. The autoradiographs
show the results of one representative experiment repeated at least
3 times. β-actin was used as a control. (B) Quantitative
representation after densitometry of the OXPHOS components. The
columns are the mean of three independent experiments in which band
intensities were evaluated in terms of optical density arbitrary
units and expressed as a percentage with respect to the respective
time controls, which were assumed to be 100%. *P=0.05,
*P<0.02. CIV, CI, CIII, CV, CII indicate OXPHOS
components. (C) ATP content was assessed as reported in Materials
and methods. Columns represent the mean ± SEM of 20 independent
experiments performed in duplicate. *P<0.05 vs. the
control (C) at the same time; 6 and 16 h.. (D) Western blot assay
of AMPK in both cell lines in the absence (C) or with increasing Bg
concentrations (B20 µM and B50 µM) at the indicated
times. β-actin was used as a control. The autoradiographs show the
results of one representative experiment, and the numbers on the
top of the blot are mean of 3 independent experiments in which band
intensities were evaluated in terms of optical density arbitrary
units and expressed as the fold over each control (C), which was
assumed to be 1. *P<0.02, **P<0.01 vs.
the control (C) at the same time. Bg, bergapten. |
AMPK is activated by metabolic stresses that
interfere with ATP production (2).
In basal conditions the levels of ATP appeared to be higher in the
ZR75 cells, although not significantly different from that observed
in the MCF7 cells Fig. 4D.
Following treatment with Bg, our data correlated well with the
increased AMPK expression in the MCF7 cells accordingly to the
reduced ATP production, while AMPK expression was unchanged in the
ZR75 cells, that produced a major amount of ATP.
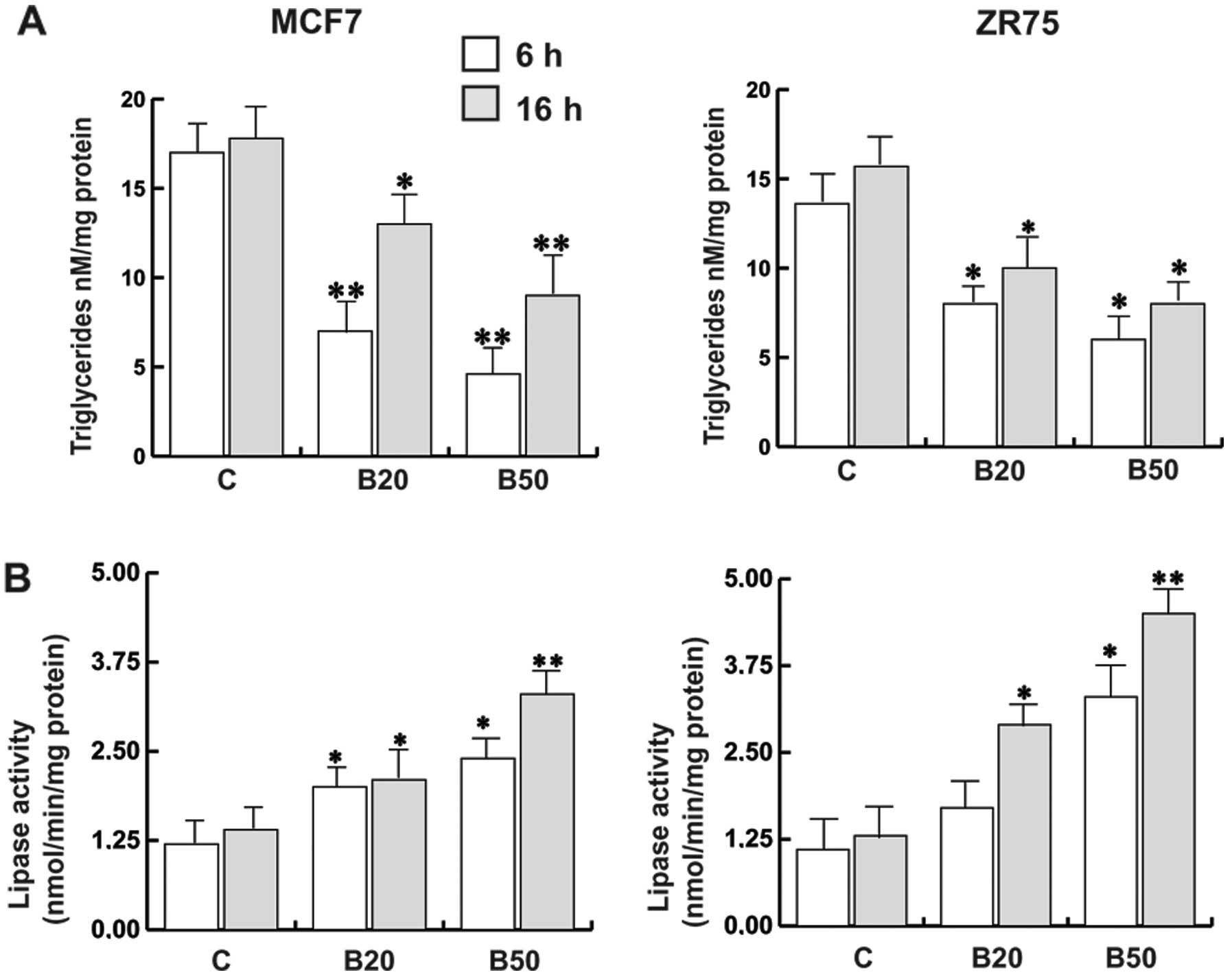
Bg promotes a lipid-lowering effect in
breast cancer cells
The bioenergetic and biosynthetic requirements of
cancer cells are balanced by regulating the flux of pathways that
metabolize fatty acids other than glucose. Increased fatty acid
synthesis has been linked to poor prognosis in breast cancer
(22). Bg-treated cells exhibited a
reduced triglyceride level compared to the untreated cells,
accordingly to the increased lipase activity observed in both cell
lines in a time- and concentration-dependent manner (Fig. 5A and B).
Bg-induced metabolic reprogramming
produces cell death in breast cancer
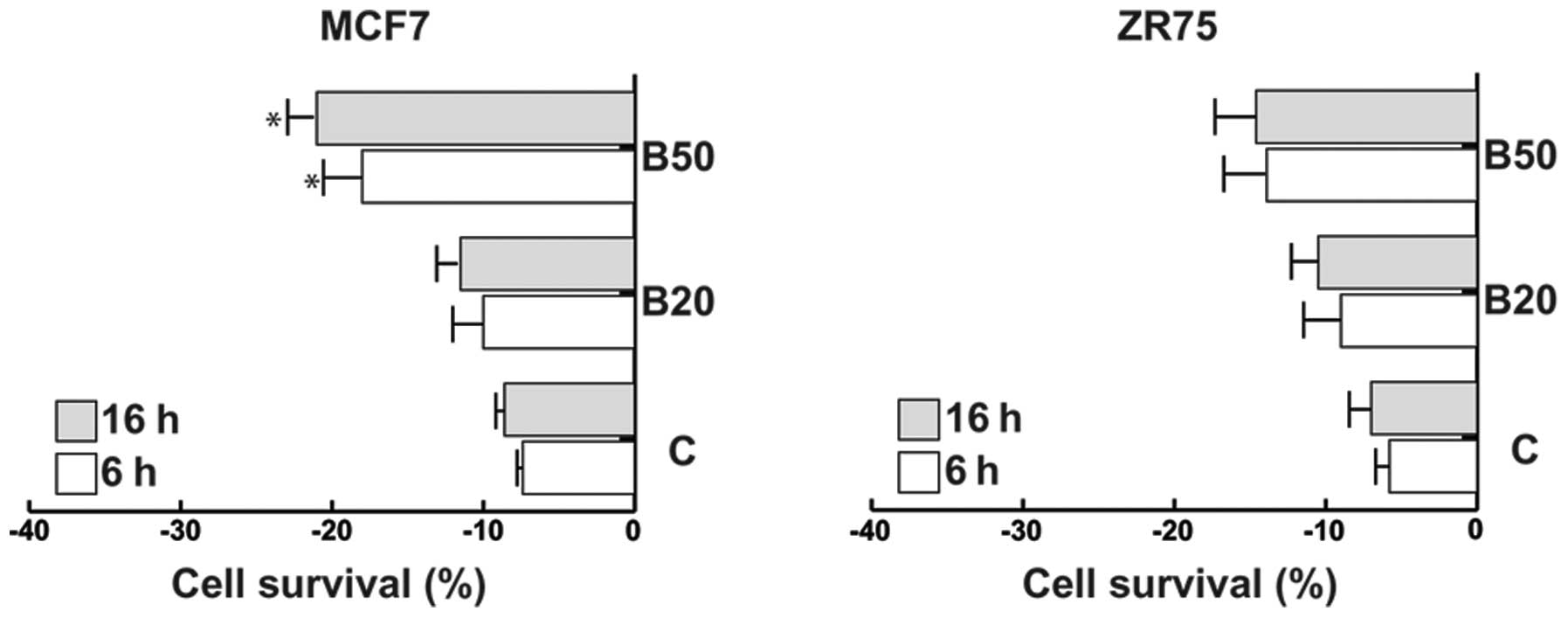
As shown in Fig. 6
both MCF7 and ZR75 cells exposed to Bg exhibited a loss of cell
survival, although the effect was higher in the MCF7 than that in
the ZR75 cells.
Discussion
It has now been clearly established that cancer
cells depend on an altered cellular metabolism (23). Cancers are extremely heterogeneous
diseases and each cancer has its individual metabolic features. The
literature indicates that the Warburg phenotype is not exclusive
and that a decrease in mitochondrial function is not a general
feature of cancer cells. In fact, even in glycolytic tumors, OXPHOS
is not completely shut down. Herein, we characterized the metabolic
profile of MCF7 and ZR75 cells evaluating various features of tumor
metabolism. We found that Bg, a natural product, alters both
glucose and lipid metabolism in a way and extent that cannot be
counterbalanced by breast cancer cells as they die.
Energy metabolism represents an additional hallmark
of cancer cells and a new promising target for its treatment.
Multiple molecular mechanisms converge to alter core cellular
metabolism. Malignant cells have the ability to adopt an altered
metabolic profile that fulfills biosynthetic and bioenergetic
requirements for rapid and uncontrolled growth. Thus, its
disruption may then resolve the malignant process, independently of
its origin.
In the present study, detailed attention was given
to evaluate the effect of Bg on breast cancer metabolism.
Glycolysis is the metabolic program of choice for cells that are
actively engaged in cell growth and mitogenesis. Because of this
fact, we first analyzed the glucose content and utilization in both
MCF7 and ZR75 cells. The basal glucose content was higher in MCF7
than in ZR75 cells and it increased upon Bg treatment in both cell
lines, while the PFK1 expression and the lactate production rates
were strongly decreased. Elevated production of lactate has been
previously correlated with the activation of PFK1, the major
regulatory glycolytic enzyme (24).
It has been proposed that inhibition of LDH and PFK1 may represent
alternative strategies toward the development of
anti-glycolytic-based therapies for cancer and in this scenario,
the data obtained upon Bg treatment was well-fit. The LDH activity
may be reduced since PFK1 was in turn lowered, however it was not a
rate-limiting step in our breast tumor cell lines. It is known that
inhibition of PFK-1 results in the accumulation of
fructose-6-phosphate, which is then isomerized to
glucose-6-phosphate and in turn is diverted into PPP, an important
pathway of glucose catabolism representing another feature of the
cancer metabolic phenotype. PPP is the primary cellular source of
NADPH, the principal intracellular reductant and crucial to fatty
acid and amino acid biosynthesis. It has been reported that the
enhanced activation of PPP has a number of pro-oncogenic effects
(1) and it is known to be
hyperactive in cancer (25,26). In this respect, our data showed that
Bg was able to affect the PPP pathway reducing G6PDH activity. The
Bg-induced lowering of LDH and G6PDH activities may explain the
higher glucose levels upon treatment as were obtained. The basal
levels of G6PDH activity were lower at 16 h, which may be due to
the longer absence of growth factor in the medium used (27). The different LDH and G6PDH basal
activities between MCF7 and ZR75 cells that we found, may suggest
that the MCF7 cells have a marked glycolytic phenotype when
compared with the ZR75 cells. Moreover, the reduction in these
enzymatic activities by Bg indicates that it is able to derange
glucose metabolism in breast cancer cells. It appears that Bg
shares similarities with other classes of molecules found in nature
within polyphenols, that are widely known as enzyme inhibitors and
particularly of glucose metabolism. The repression of the PI3K/Akt
activity as we previously demonstrated (12), also impaired anabolic glucose
metabolism supporting our new data. Furthermore, Bg also
antagonized the upregulatory effect of insulin on pGSK3S9
expression, an essential control point in glycogen synthesis. It is
clear that the absence of energetic reservoirs does not favors
cancer cell survival.
Since oncogenic metabolic programs support
bioenergetics, we focused our study on NAPDH production, oxidative
mitochondrial activity as well as ATP content. Other than PPP, two
enzymes are involved in the production of NADPH, ME and ICDH. Bg
was able to affect both ME and ICDH activities, by inducing an
increase at 20 µM, while at a higher dose after 16 h of
incubation the level decreased significantly in the MCF7 cells. A
similar pattern of response was obtained in ZR75 cells at 6 h;
however, treatment with Bg at 50 µM was not able to reduce
both enzymatic activities. Of consequence, it appears that Bg acts
on NADPH production without biasing these two metabolic steps.
Furthermore, as observed for LDH and G6PDH activities, both basal
ME and ICDH activities were higher in the MCF7 cells when compred
with the activities in the ZR75 cells.
It is well known that oxidation of NADH mainly
occurs via mitochondrial OXPHOS, where electron transport is
coupled along 4 enzyme complexes (CI-CIV), and ATP is synthesized
at CV (ATP synthase) (28,29). From our data, it is evident that
treatment with Bg in MCF7 cells tended to reduce the expression of
CI, and then the entrance of the electrons in the mitochondria,
although levels of the other downstream components were maintained.
This would translate into an internal imbalance that is unable to
supply energy to the cells, accordingly to the minor amount of ATP
produced. In contrast, in ZR75 cells, the amount of OXPHOS in
untreated and Bg-treated cells was higher compared to that observed
in the MCF7 cells. This finding supports the increased ATP
production in ZR75 cells where a reduction in complex IV of the
respiratory chain was noted. Deregulation of these processes is a
major pathological consequence of cancer progression.
AMPK is a highly conserved sensor of increased
levels of AMP and ADP originating from ATP depletion (30,31).
It appears to be involved in cancer because of its ability to act
as a tumor-suppressor. From our data, upon Bg treatment, the AMPK
content increased in MCF7 cells acting as an energy sensor allowing
the cells to meet the reduced energy supply, while in ZR75 cells
its expression did not change, in agreement with the higher ATP
levels obtained in these cells. Summarizing the role of Bg on
glucose utilization and bioenergetics: in MCF7 cells, upon Bg
treatment, glycolysis and PPP rates were significantly reduced,
OXPHOS components were scarcely impaired and the ATP production was
reduced. In ZR75 cells, the psoralen induced similar effects on
glycolysis, while an increased OXPHOS as well as ATP content were
observed. These data suggest that the two cell lines exhibit
different bioenergetic phenotypes. MCF7 cells express prevalently a
glycolytic phenotype only partially oxidative, while ZR75 cells
mainly express an oxidative phenotype. This may explain why ZR75
cells are more resistant to B-induced cell death vs. MCF7 cells, as
we previously reported (12). This
assumption is supported by a recent finding demonstrating how ZR75
cells selectively use genes for energy, sugar metabolism and other
pathways differently from MCF7 cells, imparting more aggressiveness
to ZR75 cells (24).
The bioenergetic and biosynthetic requirements of
cancer cells are balanced by regulating the flux of pathways that
metabolize fatty acids other than glucose. Proliferating cells can
synthesize fatty acids and cholesterol de novo from glucose
and increased fatty acid synthesis has been linked to poor
prognosis in breast cancer (32).
The dependence of tumor cells on deregulated lipid metabolism
suggests that pathways involved in this process are additional
attractive targets for cancer treatment. In our study, Bg greatly
impacted this feature of cancer cell metabolism since it induced a
general lipid-lowering effect. Triglyceride levels decreased
concomitantly with an increase in lipase activity. All together,
the metabolic reprogramming by Bg was acutely efficacious to reduce
cancer cell survival according to previously reported data.
We report, for the first time, that Bg treatment
interferes with breast cancer cell metabolism disrupting different
features of this process. The outcomes emerged from an extensive
characterization of the different biochemical pathways involved in
cell metabolism. On the basis of these and previous data, the
activity of Bg against breast cancer cells is therefore
orchestrated by several components rather than being the result of
the effect on a single molecular target and/or signaling
pathway.
Future clinical data describing the metabolic
profiles of human tumors will be required to determine which
metabolic alterations are most prevalent in specific tumor types.
In conclusion, bergapten, on the basis of its metabolic targeting,
can be used in combination with other forms of targeted
chemotherapy to improve cancer treatment outcomes.
Acknowledgments
The present study was supported by MIUR Ex 60%-2014
and Associazione Italiana Ricerca sul Cancro (AIRC) (grant no.
IG15738). Our special thanks to Dr Vincenzo Cunsolo (Biogemina
Italia Srl, Catania, Italy); and Serena Gervasi and Maria Clelia
Gervasi for the English language revision of the manuscript.
Abbreviations:
|
Bg
|
bergapten
|
|
LDH
|
lactate dehydrogenase
|
|
G6PDH
|
glucose-6-phosphate dehydrogenase
|
|
OXPHOS
|
oxidative phosphorylation
|
References
|
1
|
Barger JF and Plas DR: Balancing
biosynthesis and bioenergetics: Metabolic programs in oncogenesis.
Endocr Relat Cancer. 17:R287–R304. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Smolková K, Plecitá-Hlavatá L, Bellance N,
Benard G, Rossignol R and Ježek P: Waves of gene regulation
suppress and then restore oxidative phosphorylation in cancer
cells. Int J Biochem Cell Biol. 43:950–968. 2011. View Article : Google Scholar
|
|
3
|
Phan LM, Yeung S-C and Lee M-H: Cancer
metabolic reprogramming: importance, main features, and potentials
for precise targeted anti-cancer therapies. Cancer Biol Med.
11:1–19. 2014.PubMed/NCBI
|
|
4
|
Vamecq J, Colet JM, Vanden Eynde JJ,
Briand G, Porchet N and Rocchi S: PPARs: Interference with Warburg'
effect and clinical anticancer trials. PPAR Res. 2012:3047602012.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nabekura T: Overcoming multidrug
resistance in human cancer cells by natural compounds. Toxins
(Basel). 2:1207–1224. 2010. View Article : Google Scholar
|
|
6
|
da Rocha AB, Lopes RM and Schwartsmann G:
Natural products in anticancer therapy. Curr Opin Pharmacol.
1:364–369. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rowinsky EK: Paclitaxel pharmacology and
other tumor types. Semin Oncol. 2:S19-1–S19-12. 1997.
|
|
8
|
Jordan MA, Toso RJ, Thrower D and Wilson
L: Mechanism of mitotic block and inhibition of cell proliferation
by Taxol at low concentrations. Proc Natl Acad Sci USA.
90:9552–9556. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Moos PJ and Fitzpatrick FA: Taxanes
propagate apoptosis via two cell populations with distinctive
cytological and molecular traits. Cell Growth Differ. 9:687–697.
1998.PubMed/NCBI
|
|
10
|
Huang Y, Johnson KR, Norris JS and Fan W:
Nuclear factor-kappaB/IkappaB signaling pathway may contribute to
the mediation of paclitaxel-induced apoptosis in solid tumor cells.
Cancer Res. 60:4426–4432. 2000.PubMed/NCBI
|
|
11
|
McDaid HM and Horwitz SB: Selective
potentiation of paclitaxel (Taxol)-induced cell death by
mitogen-activated protein kinase kinase inhibition in human cancer
cell lines. Mol Pharmacol. 60:290–301. 2001.PubMed/NCBI
|
|
12
|
Panno ML, Giordano F, Mastroianni F, Palma
MG, Bartella V, Carpino A, Aquila S and Andò S: Breast cancer cell
survival signal is affected by bergapten combined with an
ultraviolet irradiation. FEBS Lett. 584:2321–2326. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Panno ML, Giordano F, Palma MG, Bartella
V, Rago V, Maggiolini M, Sisci D, Lanzino M, De Amicis F and Andò
S: Evidence that bergapten, independently of its photoactivation,
enhances p53 gene expression and induces apoptosis in human breast
cancer cells. Curr Cancer Drug Targets. 9:469–481. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Guido C, Panza S, Santoro M, Avena P,
Panno ML, Perrotta I, Giordano F, Casaburi I, Catalano S, De Amicis
F, et al: Estrogen receptor beta (ERβ) produces autophagy and
necroptosis in human seminoma cell line through the binding of the
Sp1 on the phosphatase and tensin homolog deleted from chromosome
10 (PTEN) promoter gene. Cell Cycle. 11:2911–2921. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Saifer A and Gerstenfeld S: The
photometric microdetermi-nation of blood glucose with glucose
oxidase. J Lab Clin Med. 51:448–460. 1958.PubMed/NCBI
|
|
16
|
Guido C, Perrotta I, Panza S, Middea E,
Avena P, Santoro M, Marsico S, Imbrogno P, Andò S and Aquila S:
Human sperm physiology: Estrogen receptor alpha (ERα) and estrogen
receptor beta (ERβ) influence sperm metabolism and may be involved
in the pathophysiology of varicocele-associated male infertility. J
Cell Physiol. 226:3403–3412. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Panteghini M, Bonora R and Pagani F:
Measurement of pancreatic lipase activity in serum by a kinetic
colorimetric assay using a new chromogenic substrate. Ann Clin
Biochem. 38:365–370. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Aquila S, Bonofiglio D, Gentile M, Middea
E, Gabriele S, Belmonte M, Catalano S, Pellegrino M and Andò S:
Peroxisome proliferator-activated receptor (PPAR)gamma is expressed
by human spermatozoa: Its potential role on the sperm physiology. J
Cell Physiol. 209:977–986. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pingitore A, Cione E, Senatore V and
Genchi G: Adrenal glands and testes as steroidogenic tissue are
affected by retinoylation reaction. J Bioenerg Biomembr.
41:215–221. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gnoni GV and Paglialonga G: Resveratrol
inhibits fatty acid and triacylglycerol synthesis in rat
hepatocytes. Eur J Clin Invest. 39:211–218. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Luo J: Glycogen synthase kinase 3beta
(GSK3beta) in tumorigenesis and cancer chemotherapy. Cancer Lett.
273:194–200. 2009. View Article : Google Scholar
|
|
22
|
Zhang F and Du G: Dysregulated lipid
metabolism in cancer. World J Biol Chem. 3:167–174. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Phan LM, Yeung SC and Lee MH: Cancer
metabolic reprogramming: Importance, main features, and potentials
for precise targeted anti-cancer therapies. Cancer Biol Med.
11:1–19. 2014.PubMed/NCBI
|
|
24
|
Mandal S and Davie JR: An integrated
analysis of genes and pathways exhibiting metabolic differences
between estrogen receptor positive breast cancer cells. BMC Cancer.
7:1812007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Richardson AD, Yang C, Osterman A and
Smith JW: Central carbon metabolism in the progression of mammary
carcinoma. Breast Cancer Res Treat. 110:297–307. 2008. View Article : Google Scholar :
|
|
26
|
Meadows AL, Kong B, Berdichevsky M, Roy S,
Rosiva R, Blanch HW and Clark DS: Metabolic and morphological
differences between rapidly proliferating cancerous and normal
breast epithelial cells. Biotechnol Prog. 24:334–341. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tian WN, Braunstein LD, Pang J, Stuhlmeier
KM, Xi QC, Tian X and Stanton RC: Importance of glucose-6-phosphate
dehydrogenase activity for cell growth. J Biol Chem.
273:10609–10617. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sun X, Wang JF, Tseng M and Young LT:
Downregulation in components of the mitochondrial electron
transport chain in the postmortem frontal cortex of subjects with
bipolar disorder. J Psychiatry Neurosci. 31:189–196.
2006.PubMed/NCBI
|
|
29
|
Reinecke F, Smeitink JA and van der
Westhuizen FH: OXPHOS gene expression and control in mitochondrial
disorders. Biochim Biophys Acta. 1792:1113–1121. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hardie DG: AMP-activated protein kinase:
An energy sensor that regulates all aspects of cell function. Genes
Dev. 25:1895–1908. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mihaylova MM and Shaw RJ: The AMPK
signalling pathway coordinates cell growth, autophagy and
metabolism. Nat Cell Biol. 13:1016–1023. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shurbaji MS, Kalbfleisch JH and Thurmond
TS: Immunohistochemical detection of a fatty acid synthase (OA-519)
as a predictor of progression of prostate cancer. Hum Pathol.
27:917–921. 1996. View Article : Google Scholar : PubMed/NCBI
|