Introduction
Ovarian cancer is one of the deadliest gynecological
cancers worldwide. The most recent update from the Cancer
Statistics of America stated that ovarian cancer is the fifth most
common cause of cancer-related deaths. In 2017, the number of newly
diagnosed cases of ovarian cancer was projected to be ~22,440, and
the number of deaths was predicted to be 14,080 (1). In 2015, the number of new cases of
ovarian cancer in China was ~52,100, whereas the number of deaths
was ~22,500; this places ovarian cancer as the seventh leading
cause of cancer-related death among Chinese women (2). Among the diverse histological subtypes
of ovarian cancer, epithelial ovarian cancer (EOC) accounts for
~85–90% of all types of ovarian carcinoma, and up to 75% of
patients with EOC were diagnosed with advanced stage disease and
were characterized by rapid progression, poor prognosis and a high
frequency of TP53 mutations (3–5). The
cause of EOC is still unclear, but in recent years, accumulating
studies have demonstrated that inflammation plays an important role
in ovarian cancer tumorigenesis and progression (6–9).
Formyl peptide receptor 2 (FPR2) has been identified
as a member of the G protein-coupled chemoattractant receptor
(GPCR) family. It is a seven-transmembrane receptor with 351 amino
acids and is located on human chromosome 19q13.3-q13.4 (10). It was first detected on the cell
membrane of macrophages and neutrophils and is activated by
bacterial formylated chemotactic peptides. FPR2 was later reported
to be expressed in a wide range of cells, tissues and organs,
including inflammatory, microglial and astrocytoma and
neuroblastoma cells, hepatocytes, microvascular endothelial cells,
fibroblasts, spleen, lung, testes, ovaries, placenta, brain and
bone marrow (11,12). FPR2 is a multi-functional receptor
that is associated with diverse pathophysiologic processes, such as
inflammation, cancer, amyloidosis and neurodegenerative diseases,
wound healing, diabetes and obesity and AIDS (13). There is also a close relationship
between FPR2 and cancer, as FPR2 has been detected via flow
cytometry in seven ovarian cancer cell lines and the FPR2
agonist-antimicrobial peptide LL-37 was suggested to stimulate the
invasiveness of ovarian cancer cells by interacting with FPR2
(14). FPR2 has been proposed as a
bridge between inflammation and cancer. Serum amyloid A (SAA) is an
acute phase protein that can be stimulated by inflammation,
infection, trauma or tumorigenesis. In a study of Urieli-Shoval
et al, SAA was found to be strongly expressed in ovarian
cancer tissues by IHC, ISH and RT-PCR (15). SAA is one of the primary agonists of
FPR2, and previous studies have suggested that SAA-induced
activation of FPR2 stimulates inflammation, cell migration,
adhesion and infiltration (16).
The SAA-FPR2 interaction also contributes to inflammation-mediated
neovascularization in the cornea (17). Thus, we hypothesized that SAA may be
expressed by ovarian cancer cells and interacts with FPR2 to
stimulate ovarian cancer invasion and migration. In the present
study, we explored the effects of FPR2 in the presence or absence
of SAA on EOC progression.
Materials and methods
Quantitative real-time PCR
Total RNA was extracted from either tissues or cells
using TRIzol reagents (Pufei Biotechnology, Shanghai, China).
Reverse transcription was performed using M-MLV reverse
transcriptase following the manufacturer's instructions (Promega,
Madison, WI, USA). We used 2 µg of total RNA in a sterile
RNase-free microcentrifuge tube, and then added 0.5 µg of the
primer or per microgram of the total RNA sample in a total volume
of 15 µl in water. The tube was heated to 70°C for 5 min to melt
secondary structure within the template. The tube was cooled
immediately on ice to prevent secondary structure from reforming,
and then spinned briefly to collect the solution at the bottom of
the tube. Then, the following components were added to the annealed
primer/template: M-MLV 5X reaction buffer 5 µl; dATP, 10 mM 1.25
µl; dCTP, 10 mM 1.25 µl; dGTP, 10 mM 1.25 µl; dTTP, 10 mM 1.25 µl;
recombinant RNasin ribonuclease inhibitor 25 units; M-MLV RT 200
units; nuclease-free water to a final volume of 25 µl. Gentle
mixing was performed by flicking the tube, and incubation was
carried out for 60 min at 42°C for FPR2 primers. The extension
temperature was 37°C. Quantitative PCR was performed using
SYBR-Green Real-Time PCR Master Mix (Toyobo, Osaka, Japan)
according to the manufacturer's protocol. The primer sequences were
as follows: FPR2 forward, 5′-AGTCTGCTGGCTACACTGTTC-3′ and reverse,
5′-TGGTAATGTGGCCGTGAAAGA-3′; STAT3 forward,
5′-AGAAGGACATCAGCGGTAAG-3′ and reverse,
5′-CCTTGGGAATGTCAGGATAGAG-3′; NF-κB forward,
5′-AGGATTTCGTTTCCGTTATGT-3′ and reverse,
5′-CCTGAGGGTAAGACTTCTTGTTC-3′; MAPK1 forward,
5′-GTCGCCATCAAGAAAATCAGC-3′ and reverse,
5′-GGAAGGTTTGAGGTCACGGT-3′; Notch3 forward,
5′-TGTGGACGAGTGCTCTATCG-3′ and reverse, 5′-AATGTCCACTCGCAATAGG-3′;
GAPDH forward, 5′-TGACTTCAACAGCGACACCCA-3′ and reverse,
5′-CACCCTGTTGCTGTAGCCAAA-3′.
Immunohistochemistry (IHC)
The ovarian cancer tissues were fixed in 10%
formalin and paraffin-embedded to form tissue blocks from which
sections (4-µm thickness) could be sliced for IHC studies. The
tissue sections were deparaffinized in xylene and rehydrated in
graded concentrations of alcohol, and then were placed in a
microwave for 30 min for antigen retrieval. After the slides were
dewaxed, 3% H2O2 was added for 10 min to
inhibit any endogenous peroxidase activity. The sections were
incubated with the FPR2 antibody (13448–1-AP; 1:50 dilution;
Proteintech, San Diego, CA, USA) at 4°C overnight and subsequently
incubated with a streptavidin-peroxidase system (Zhongshan
Goldenbridge Biotechnology, Beijing, China) according to the
manufacturer's instructions.
Cell culture
The human ovarian cancer cell lines SKOV3, OVCAR3
and the HUVECs were obtained from the America Type Culture
Collection (ATCC; Manassas, VA, USA), and the HO-8910 and HO-8910PM
cell lines were obtained from the China Center for Type Culture
Collection (CCTCC). The cell lines were cultured in either
Dulbeccos modified Eagles medium (DMEM) or RPMI-1640 medium
supplemented with 10% fetal bovine serum (FBS) and 100 UI/ml of
penicillin and 100 mg/ml of streptromycin (all from Biological
Industries, Cromwell, CT, USA) in a 37°C incubator containing 5%
CO2.
Vector construction and
transfection
The FPR2 knockdown and control lentiviruses were
purchased from GeneChem (Shanghai, China). The shRNA sequences for
FPR2 knockdown were as follows: shRNA-1, ccggAATCATTGACAT
CCTGGTTAActcgagTTAACCAGGATGTCAATGATTtttttg; shRNA-2,
ccggGGCCAAGACTTCCGAGAGAGActcgagTC TCTCTCGGAAGTCTTGGCCtttttg;
shRNA-3, ccggGTCC TATGAGTCTGCTGGCTActcgagTAGCCAGCAGACTCAT
AGGACtttttg. The shRNA-2 sequence exhibited the best interference
efficiency.
Wound-healing assay
Cells (3×104/well) were seeded on 96-well
plates and grown to 90% confluence, after which a scratch was made
in the monolayer using a 10-µl pipette tip. Then, the cells were
incubated at 37°C in 5% CO2 for another 4 h according to
the result of the pre-experiment, and the images were obtained at
the time points of 0, 2 and 4 h. Each experiment was performed
three times.
Transwell assay
The assay was performed using a pre-coated cell
invasion kit (pore size, 8.0 µm; Corning Inc., Corning, NY, USA)
and Matrigel (BD Biosciences, Bedford, MA, USA) was inserted in the
upper chambers. Approximately 1×105 cells in 100 µl
serum-free medium were placed into the upper chambers, the cells
were cultured in 5% CO2 at 37°C for 16 h (according to
the pre-experiment). The lower chambers contained 30% FBS, thus,
the cells migrated to the lower chambers. The cells remaining in
the upper chambers were removed with a cotton swab and the cells
that migrated through the membrane to the lower surface were
stained with Giemsa's staining for 3–5 min at room temperature. The
number of cells that migrated through the lower membrane of the
inserts was counted under a light microscope. Each experiment was
performed three times.
Tube formation assay
HUVECs (4×104 cells/well) were seeded on
Matrigel-coated 96-well plates and incubated at 37°C. Serum-free
supernatant from SKOV3, shFPR2 SKOV3, SAA+shFPR2 SKOV3 and
SAA+SKOV3 cells were collected and centrifuged. The supernatants
were then incubated with the HUVECs at 37°C for 24 h. Afterwards,
the cells were fixed with 4% paraformaldehyde at 37°C for 15 min
and the capillary-like structures were obtained using Cellomics
(CCX7C1115; Thermo Fisher Scientific, Waltham, MA, USA). Branching
points and lengths were measured using ImageJ software [National
Institutes of Health (NIH), Bethesda, MD, USA]. Each experiment was
performed three times.
Simple Western (WES)
Proteins from ovarian cancer cells were extracted in
RIPA lysis buffer (P0013B; Beyotime Biotechnology, Shanghai,
China.). The protein concentrations were determined using a BCA
protein assay (Bio-Rad Laboratories, Inc., Hercules, CA, USA). The
samples and reagent were subsequently loaded into an assay plate
and placed in a ProteinSimple WES system (WS-2471; ProteinSimple,
San Jose, CA, USA.) according to the standard protocol. Finally,
the resulting chemiluminescent signal was detected and quantitated
by ProteinSimple Compass software. The following antibodies were
used: FPR2 (13448-1-AP; 1:20 dilution; Proteintech), VEGF
(Ab183100, 1:20 dilution, Abcam, Cambridge, MA, USA), STAT3
antibodies (9139; 1:20 dilution), MAPK1 (4376; 1:20 dilution),
Notch3 (2889; 1:20 dilution) [all from Cell Signaling Technology
(CST; Beverly, MA, USA)], and β-actin (69879; 1:50; Santa Cruz
Biotechnology, Inc., Santa Cruz, CA, USA).
Statistical analysis
Statistical analysis was performed using IBM SPSS
Statistics 24.0 (IBM SPSS, Armonk, NY, USA). Statistics of
continuous data were made using AVOVA or Student's t-test, while
Chi-square test was used for categorical data. At least three
independent experiments for each group were conducted, and
differences in mean values were assessed by analysis of variance
and Student's t-test. The survival curves were generated using the
Kaplan-Meier method and calculated using the log-rank test.
Prognostic factors were evaluated using Cox regression analysis.
P-value <0.05 was considered to indicate a statistically
significant result.
Results
Increased expression of FPR2 in
EOC
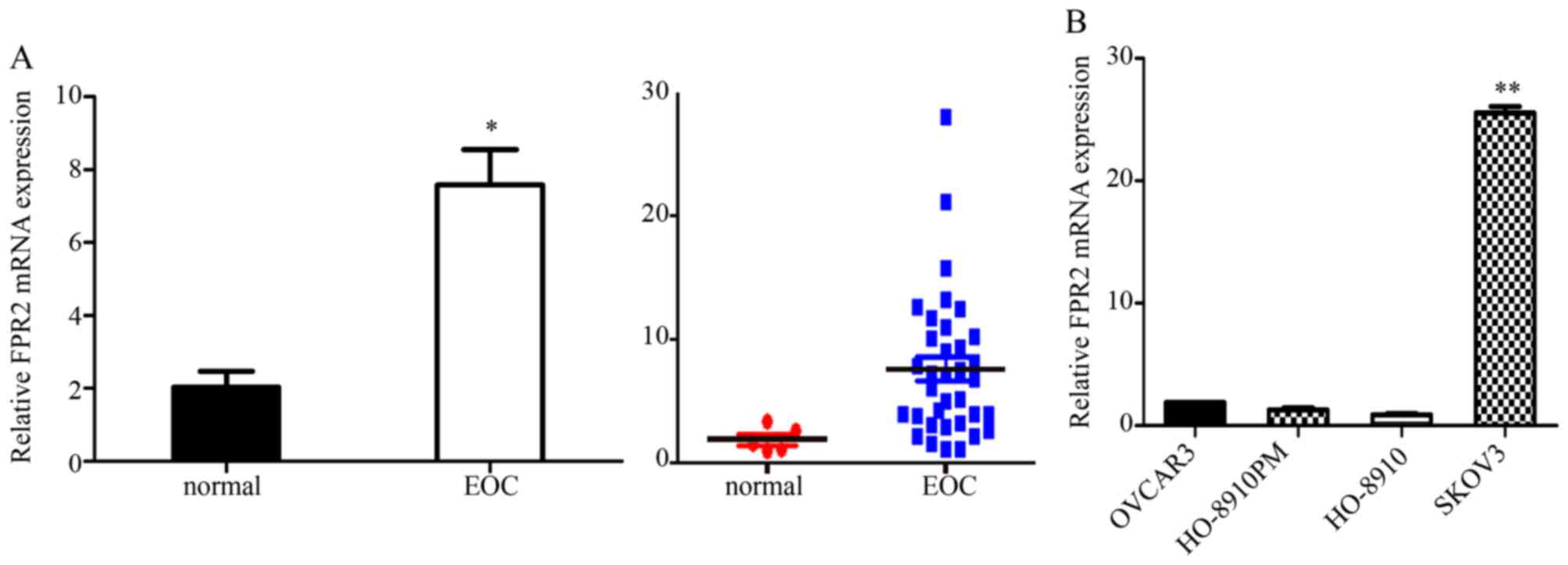
We used RT-qPCR to analyze the FPR2 expression in
both EOC tissues and cells. We collected 35 cases of EOC tissues
and five normal ovarian tissues samples for RT-qPCR assay. The
results indicated that FPR2 mRNA expression was frequently higher
in the ovarian cancer samples than that noted in the control group.
Then, we detected FPR2 mRNA expression in four ovarian cancer cell
lines including SKOV3, OVCAR3, HO-8910 and HO-8910PM. The results
showed that FPR2 mRNA expression was significantly higher in the
SKOV3 cell line than that in the other three ovarian cancer cell
lines (Fig. 1); thus, we selected
the SKOV3 cell line as a target cell line for FPR2 knockdown to
perform the further cell functional experiments. Subsequently, we
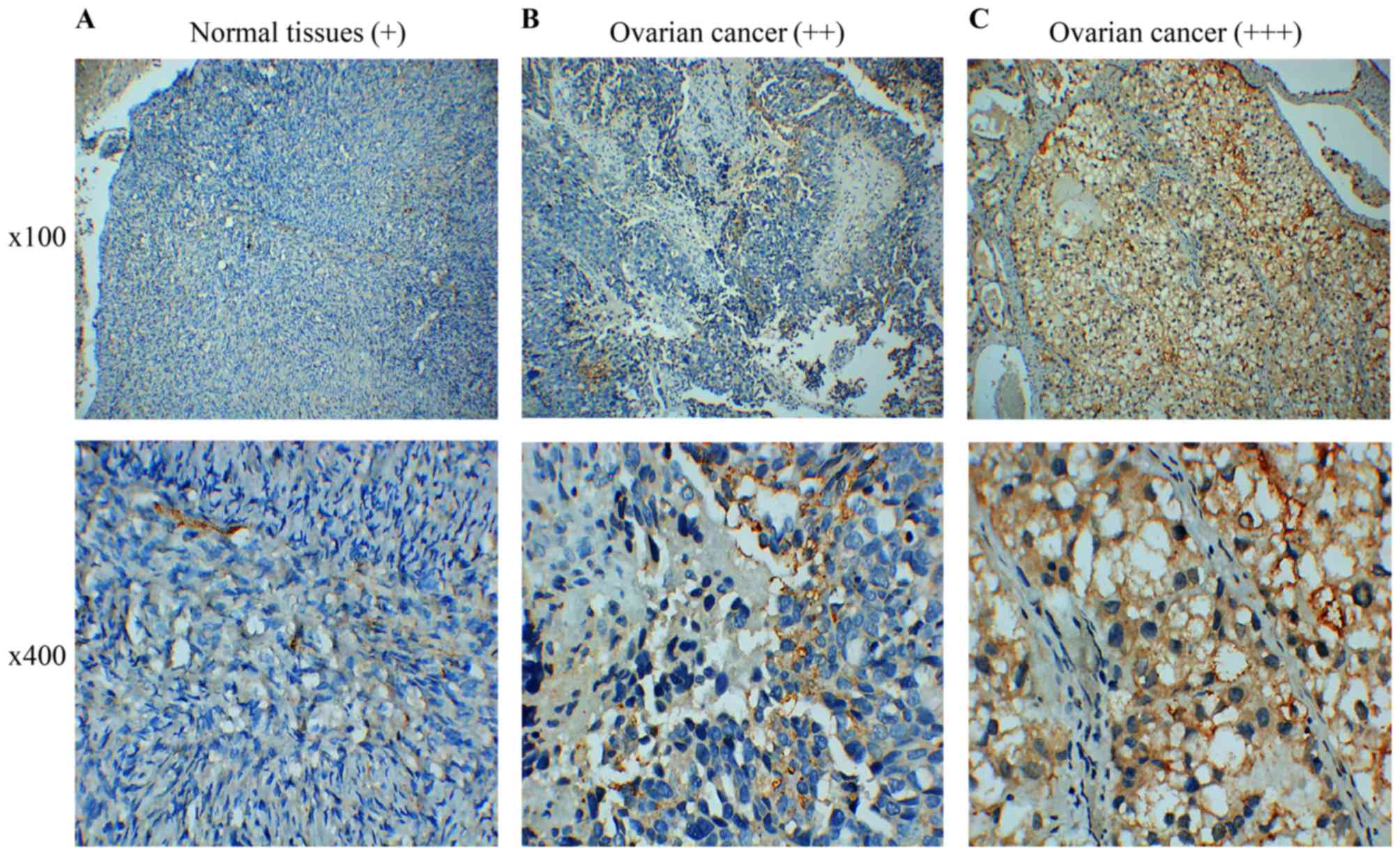
collected 60 cases of paraffin-embedded EOC specimens and 5 cases
of normal ovarian tissue specimens for IHC assay. The results
indicated that FPR2 protein expression was frequently higher in the
ovarian cancer samples than that in the control group as shown by
IHC. FPR2 expressed on the cytomembrane or cytoplasm of EOC cells
was considered as a positive result. The staining of the entire
slide was scored according to intensity: +, weak; ++, moderate; and
+++, strong). The score was calculated using the following formula:
(3 × percentage of strong staining) + (2 × percentage of moderate
staining) + (1 × percentage of weak staining), with the possible
scores ranging from 0 to 300. FPR2 expression was classified into
two groups according to a cut-off score of 100 (0–99 = low/negative
expression; 100–300 = high/positive expression) (Fig. 2) (18).
FPR2 expression level is correlated
with the clinicopathological characteristics of EOC
Analysis of the correlation between FPR2 expression
and the clinicopathological characteristics of the EOC patients
showed that high levels of FPR2 expression indicated a significant
positive correlation with International Federation of Gynecology
and Obstetrics (FIGO) stage, histological grade and ovarian cancer
type (19). (Tables I and II).
 | Table I.Correlation between FPR2 mRNA
expression and clinicopathological characteristics of the EOC
cases. |
Table I.
Correlation between FPR2 mRNA
expression and clinicopathological characteristics of the EOC
cases.
| Variables | Cases | FPR2 mRNA
expression (mean ± SD) | P-value |
|---|
| Age (years) |
|
| 0.613 |
|
≥50 | 26 | 7.94±5.71 |
|
|
<50 | 9 | 6.77±6.43 |
|
| Histological type
(carcinoma) |
|
| 0.025a |
|
Serous | 25 | 9.28±6.13 |
|
|
Mucinous | 7 | 3.80±1.43 |
|
|
Endometrial + clear cell | 3 | 2.92±1.24 |
|
| FIGO stage |
|
| 0.005b |
|
I+II | 9 | 3.59±1.78 |
|
|
III+IV | 26 | 9.04±6.10 |
|
| Histological
grade |
|
| 0.021a |
|
I+II | 13 | 4.72±3.60 |
|
|
III | 22 | 9.36±6.27 |
|
| CA125 (U/ml) |
|
| 0.274 |
|
>35 | 32 | 7.97±5.99 |
|
|
≤35 | 3 | 4.07±0.20 |
|
| Type |
|
| 0.002b |
| I | 9 | 3.35±1.31 |
|
| II | 26 | 9.12±6.06 |
|
| Table II.Correlation between FPR2 protein
expression and clinicopathological characteristics of the EOC
cases. |
Table II.
Correlation between FPR2 protein
expression and clinicopathological characteristics of the EOC
cases.
|
|
| FPR2
expression |
|
|---|
|
|
|
|
|
|---|
| Variables | Cases | Positive/high
(37) | Negative/low
(23) | P-value |
|---|
| Age (years) |
|
|
|
|
|
≥50 | 37 | 24 | 13 | 0.590 |
|
<50 | 23 | 13 | 10 |
|
| Histological type
(carcinoma) |
|
|
| 0.079 |
|
Serous | 36 | 27 | 9 |
|
|
Mucinous | 10 | 4 | 6 |
|
|
Endometrial | 7 | 3 | 4 |
|
| Clear
cell | 7 | 3 | 4 |
|
| FIGO stage |
|
|
| 0.000b |
|
I+II | 18 | 3 | 15 |
|
|
III+IV | 42 | 34 | 8 |
|
| Histological
grade |
|
|
| 0.000b |
|
I+II | 26 | 9 | 17 |
|
|
III | 34 | 28 | 6 |
|
| CA125 (U/ml) |
|
|
| 0.233 |
|
>35 | 53 | 31 | 22 |
|
|
≤35 | 7 | 6 | 1 |
|
| Type |
|
|
| 0.01a |
| I | 19 | 7 | 12 |
|
| II | 41 | 30 | 11 |
|
FPR2 overexpression is associated with
poor prognosis in EOC
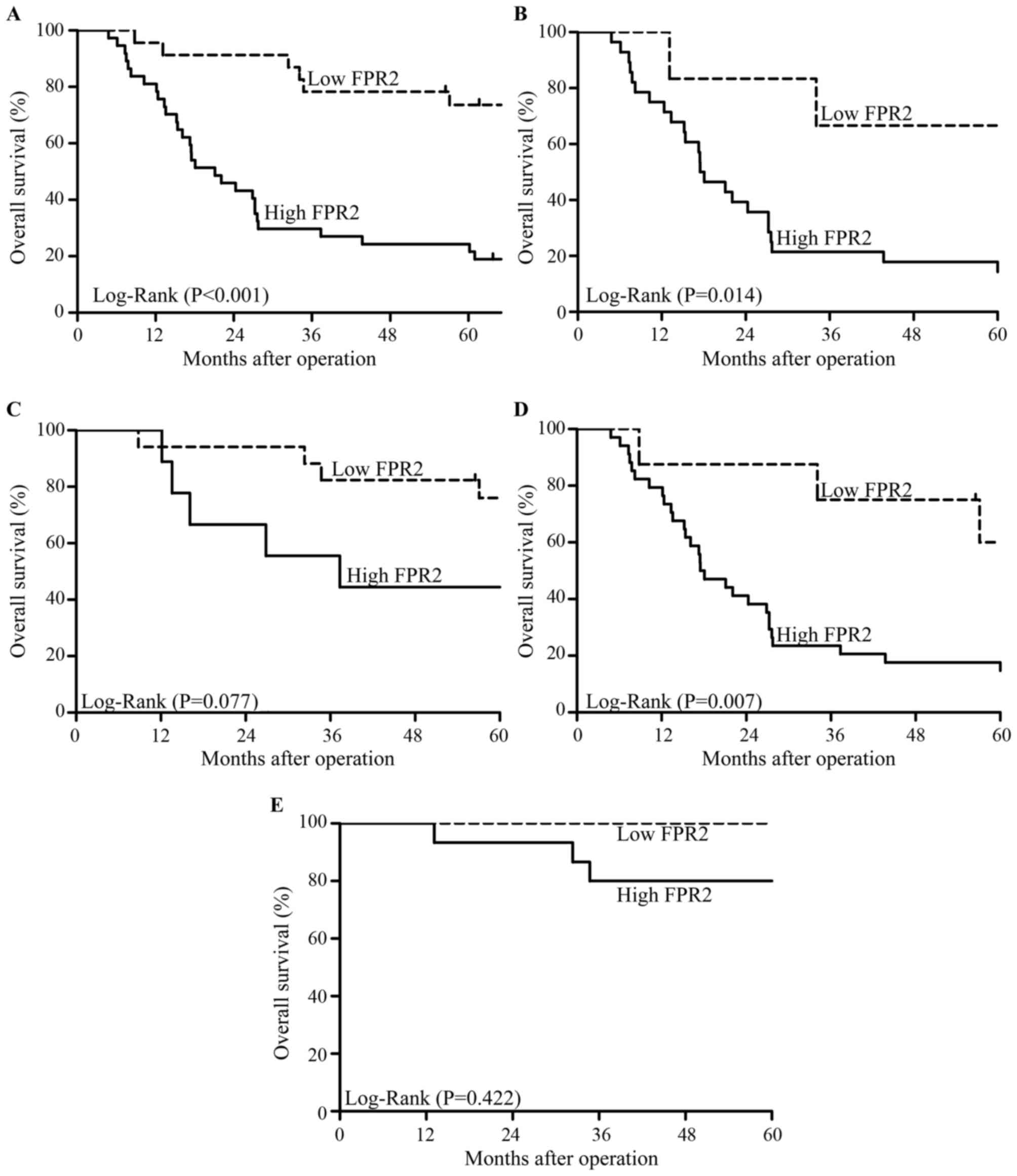
We followed up 60 cases of EOC patients for 5 years
after surgery. A total of 36 deaths occurred during follow-up and
all cases died due to cancer or cancer-related diseases. The
Kaplan-Meier survival analysis showed that patients with high FPR2
expression had a lower overall survival (OS) than those with low
FPR2 expression (Fig. 3A). In
addition, in high-grade EOC patients, high FPR2 expression
indicated a lower OS than a low FPR2 expression; whereas, OS showed
no significant difference among low-grade EOC patients regardless
of the expression of FPR2 (Fig. 3B and
C). Moreover, in advanced FIGO stage EOC patients, high FPR2
expression was associated with a lower OS compared with a low FPR2
expression; whereas, OS showed no significant difference among
patients in early FIGO stage of EOC regardless of the expression of
FPR2 (Fig. 3D and E). Univariate
and multivariate analyses showed that high FPR2 expression was a
risk factor for EOC patients (Table
III).
| Table III.Univariate and multivariate analyses
of prognostic factors for EOC. |
Table III.
Univariate and multivariate analyses
of prognostic factors for EOC.
|
|
| Univariate
analysisa | Multivariate
analysisb |
|---|
|
|
|
|
|
|---|
| Variables | n | Mean survival
(months) | P-value | HR (95% CI) | P-value |
|---|
| FIGO stage |
|
|
| 4.922
(1.389–17.446) | 0.014c |
|
I+II | 18 | 90.479±6.784 | 0.000d |
|
|
|
III+IV | 42 | 36.968±4.901 |
|
|
|
| Histological
grade |
|
|
| 1.556
(0.679–3.564) | 0.486 |
|
I+II | 26 | 76.586±7.362 | 0.000d |
|
|
|
III | 34 | 35.803±5.442 |
|
|
|
| CA125 (U/ml) |
|
|
| 0.528
(0.178–1.569) | 0.250 |
|
>35 | 53 | 54.549±5.642 | 0.898 |
|
|
|
≤35 | 7 | 41.190±9.796 |
|
|
|
| Type |
|
|
| 2.014
(0.793–5.114) | 0.141 |
| I | 19 | 83.010±7.389 | 0.001d |
|
|
| II | 41 | 38.836±5.265 |
|
|
|
| FPR2
expression |
|
|
| 3.063
(1.193–7.864) | 0.020c |
|
Low | 23 | 84.015±6.932 | 0.000d |
|
|
|
High | 37 | 34.492±5.069 |
|
|
|
FPR2 knockdown reduces the invasion
and migration of SKOV3 cells
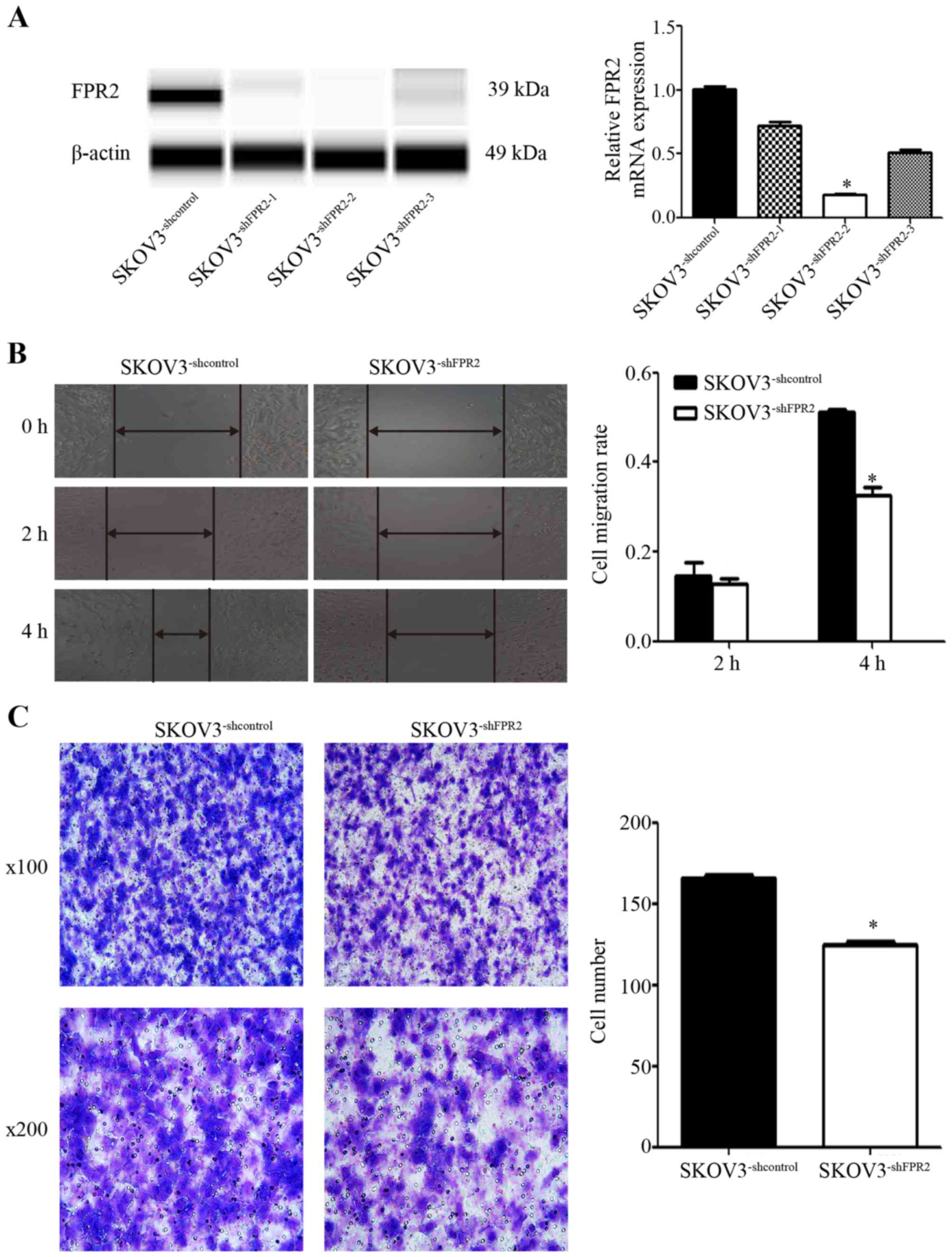
To study the role of FPR2 in ovarian cancer
migration, we stably transduced FPR2shRNA into SKOV3
(SKOV3−shFPR2) cells and verified FPR2 expression using
RT-qPCR and WES (Fig. 4A). The
wound healing and Transwell assays were used to detect the
metastatic potential of SKOV3−shFPR2 cells, with more
active motility of the tumor cells corresponding to increased
metastatic potential. The results of wound healing assay showed
that 4 h after scraping, the average migration rate of the
SKOV3−shFPR2 cells was significantly decreased (Fig. 4B). The Transwell assay also
indicated that compared to the control cells, fewer
SKOV3−shFPR2 cells penetrated the Matrigel (Fig. 4C).
SAA promotes the motility of ovarian
cancer cells via FPR2
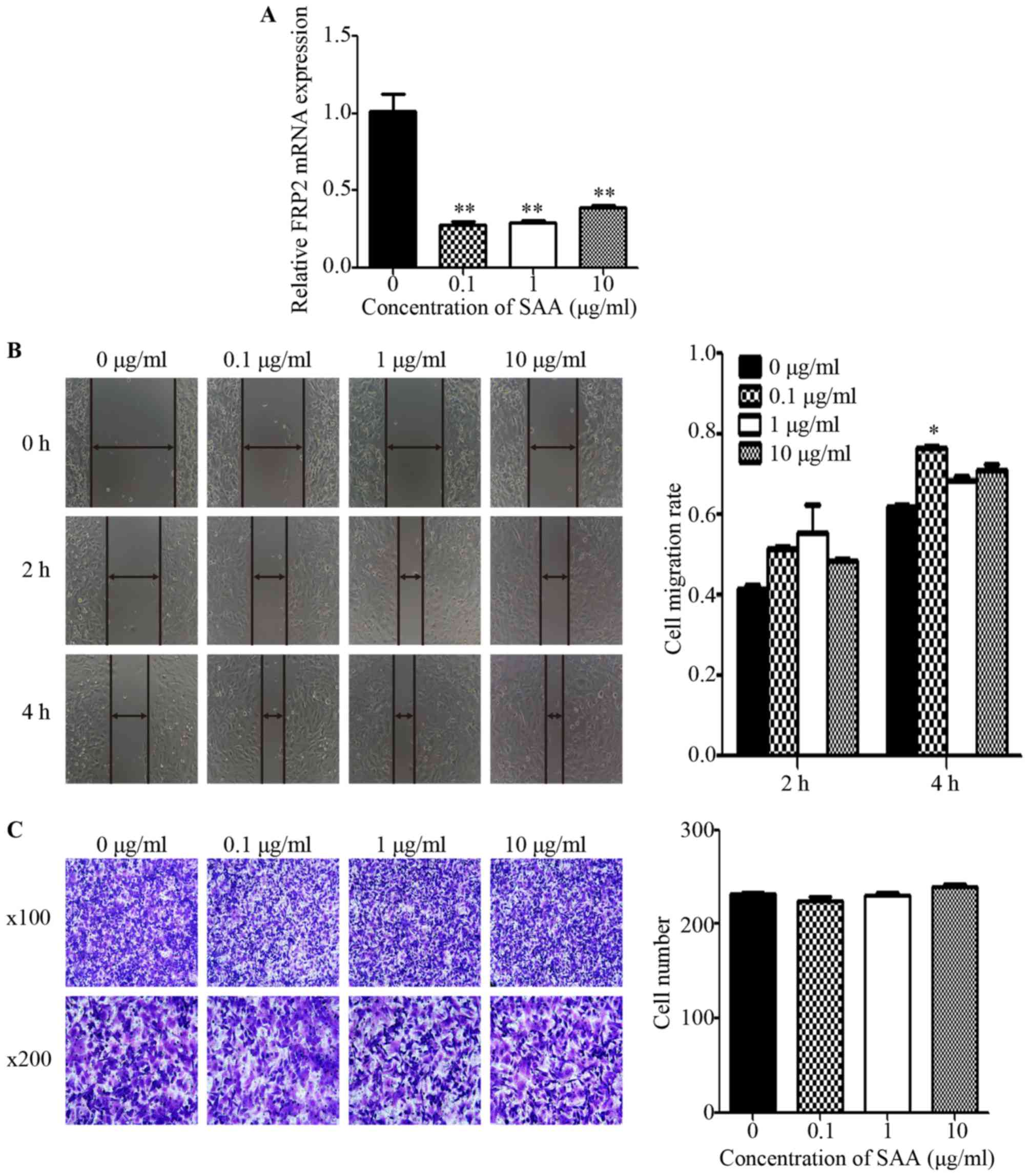
In SKOV3 cells treated with SAA (0.1, 1 and 10
µg/ml) for 48 h, the mRNA expression of FPR2 was markedly decreased
compared to the untreated control group (Fig. 5A). Then, we performed wound healing
and Transwell assays to detect cell motility. The wound healing
assay revealed that the average migration rate of the SKOV3 cells
was significantly increased after treatment with 0.1 µg/ml SAA at 4
h after scraping (Fig. 5B).
However, the Transwell assay showed no significant difference among
the SAA treatment groups regarding the number of cells that
penetrated the Matrigel when compared to the untreated control
group (Fig. 5C). To investigate
whether SAA mediates ovarian cancer cell migration through FPR2,
0.1 µg/ml SAA was added to SKOV3−shFPR2 and
SKOV3−shcontrol cells for 48 h, and the wound healing
and Transwell assays were performed. The results showed that the
average migration rate of the SAA+SKOV3−shFPR2 cells was
significantly decreased (Fig. 5D),
and Transwell assay indicate that compared to the control cells,
fewer SAA+SKOV3−shFPR2 cells penetrated the Matrigel
(Fig. 5E).
FPR2 knockdown reduces the
angiogenesis of SKOV3 cells
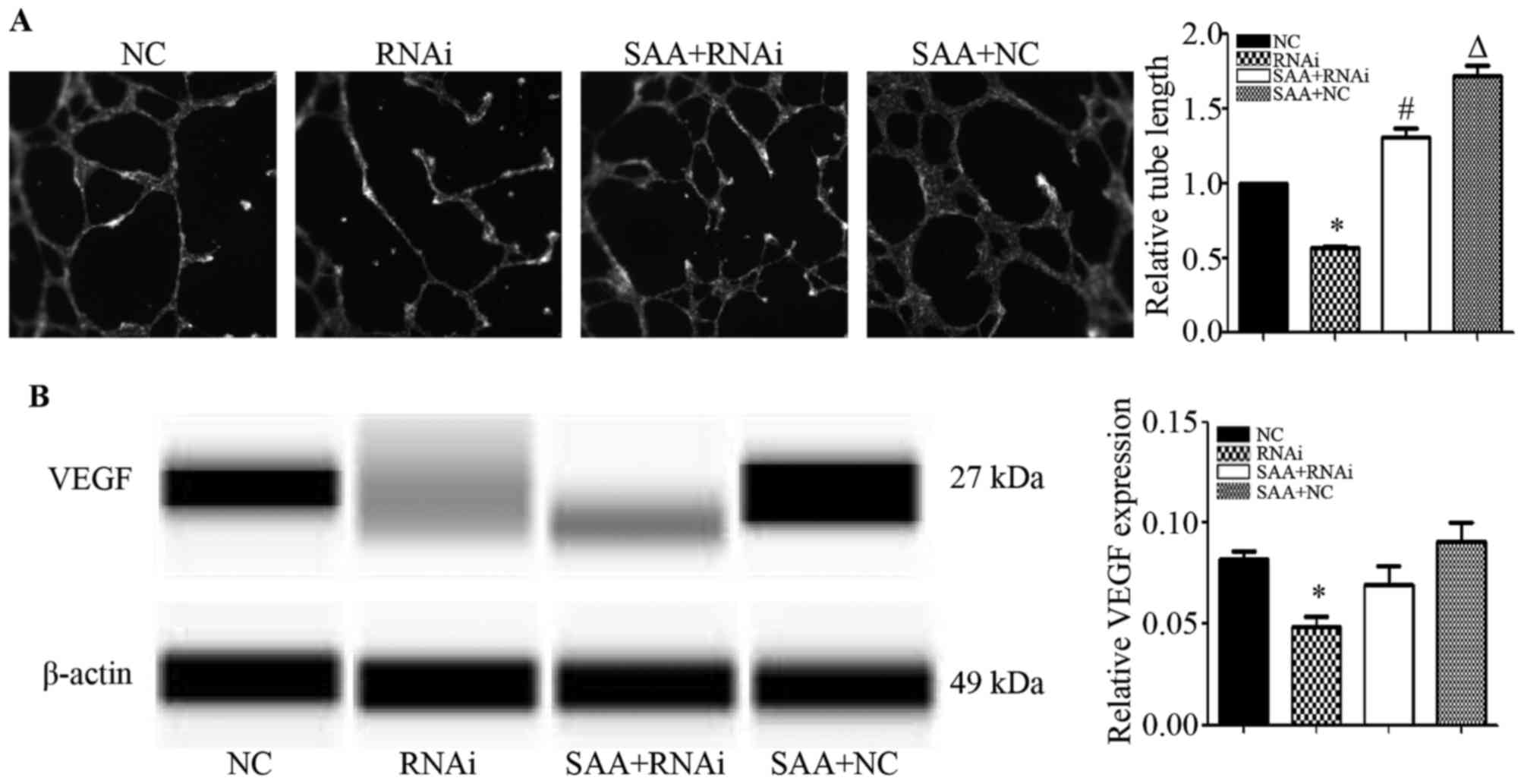
To study the effect of FPR2 on the angiogenesis of
ovarian cancer cells, we treated HUVECs with the cell culture media
from each cell type for 24 h. The results showed that compared to
the SKOV3−shcontrol (NC) group, the FPR2-knockdown
(RNAi) group exhibited significantly shorter tube lengths.
Additionally, when compared to the SKOV3−shFPR2 (RNAi)
group, the SAA+SKOV3−shFPR2 (SAA+RNAi) group markedly
increased the angiogenic ability of the HUVECs. Compared to the
SKOV3−shcontrol (NC) group, the
SAA+SKOV3−shcontrol (SAA+NC) group markedly increased
the angiogenic ability of the HUVECs (Fig. 6A). The WES results showed that the
FPR2-knockdown group exhibited significantly decreased VEGF
expression compared to that of the NC group (Fig. 6B).
The signaling pathways activated by
FPR2 in SKOV3 cells
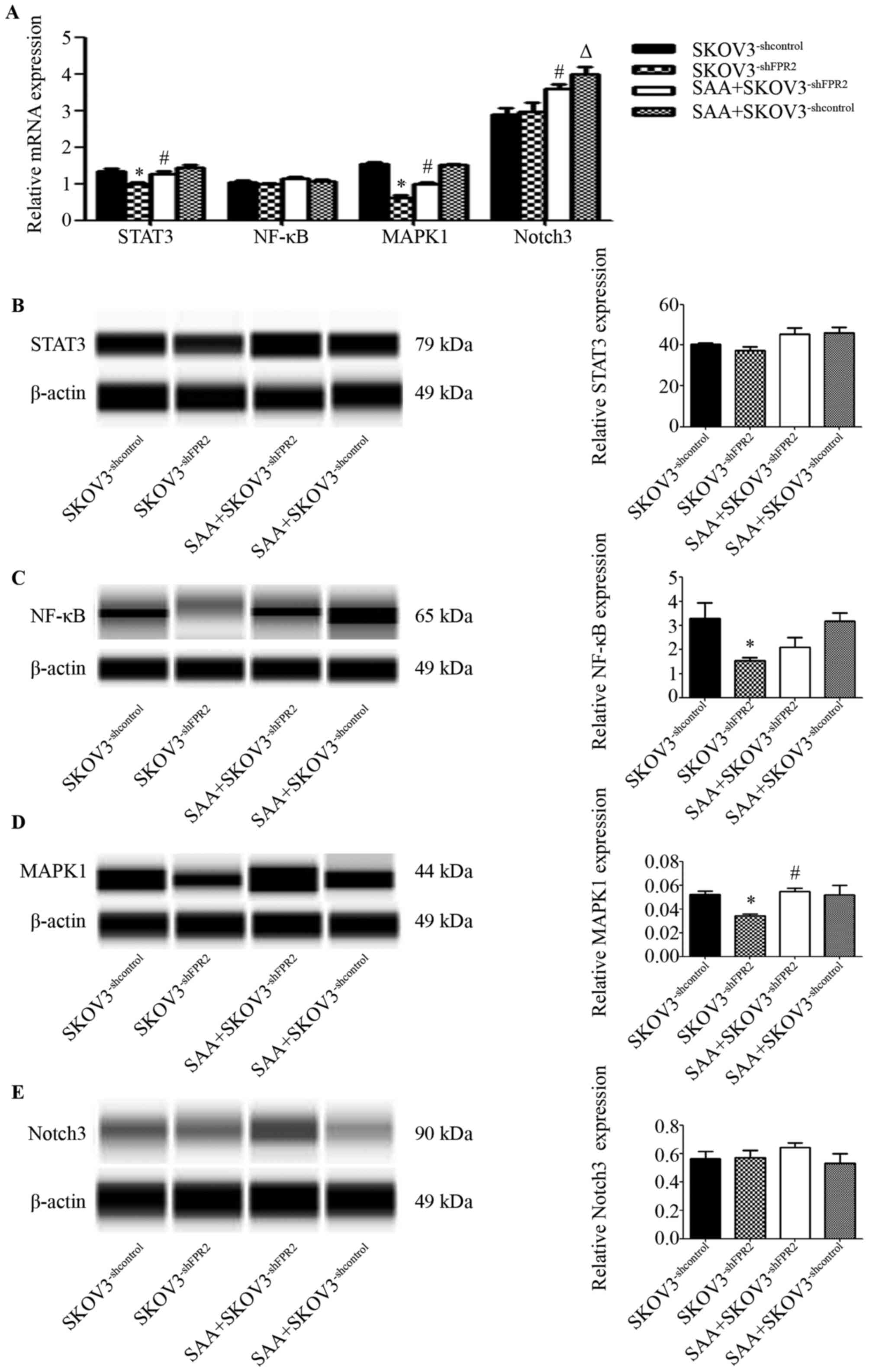
To study the potential mechanism of FPR2 regulation
in SKOV3 cells, we used RT-qPCR and WES to detected several
reported genes that are correlated with inflammation and ovarian
cancer. These genes included STAT3, NF-κB, MAPK1 and Notch3. The
RT-qPCR results showed that mRNA expression of STAT3 and MAPK1 was
evidently downregulated in the SKOV3−shFPR2 cells
compared to those in the SKOV3−shcontrol cells (Fig. 7A). The WES results indicated that
protein levels of MAPK1 and NF-κB were evidently downregulated in
the SKOV3−shFPR2 cells compared to those in the
SKOV3−shcontrol cells (Fig.
7C and D). Expression of STAT3 protein and Notch3 protein
showed no significant difference between groups (P>0.05)
(Fig. 7E)
Discussion
In the present study, we first demonstrated that
FPR2 was upregulated in ovarian cancer tissues. The literature
describes the initial discovery of FPR2 in inflammatory cells, but
additional studies have stated that this protein is expressed in a
variety of cells (including cancer cells) and binds ligands
produced under inflammatory or tumor conditions (10–12).
Oldekamp et al reported that among mice infected with
pneumococcus, mFPR1-knockout and mFPR2-transgenic mice presented
increased bacterial burden, elevated neutrophil infiltration and
high mortality compared to wild-type mice. This suggests that FPR1
and FPR2 play significant roles in the innate immune response
(20). Coffelt et al
demonstrated that ovarian cancer cell lines express FPR2 to varying
degrees. Human cathelicidin LL-37 stimulates the invasion of
ovarian cancer cells via FPR2, and oncogenes such as c5, coll8al
and mmp2 are upregulated in ovarian cancer cells (14). FPR2 has also been shown to act as a
promoter in other types of malignancies. In a study by Xiang et
al, FPR2 was detected both in vivo and in vitro
in human colon cancer. Additionally, FPR2 was demonstrated to be
highly expressed in progressive colon cancer, correlated with a
worse patient prognosis and play a role in stimulating
tumorigenesis and invasion in colon cancer cells (21). Khau et al showed that FPR2
protein could be detected in both epithelial and stromal cells of
breast cancer tissues and was shown to promote mitogens in breast
cancer cells (22). In the present
study, we showed that FPR2 is highly expressed in ovarian cancer
tissues using both IHC and RT-qPCR. Subsequently, upon analyzing
the correlation of FPR2 expression with clinicopathological
characteristics, we discovered that FPR2 expression was correlated
with FIGO stage, histological grade and ovarian cancer type, which
suggests that FPR2 may be associated with the progression of
ovarian cancer. In survival analysis, we found that FPR2
overexpression indicated a poorer prognosis of epithelial ovarian
cancer (EOC) patients and suggested that FPR2 may be an independent
risk factor for EOC, which has not been reported before. As shown
in cell experiments, the knockdown of FPR2 resulted in inhibition
of ovarian cancer cell movement, indicating that FPR2 may
contribute to the metastasis of ovarian cancer. In the future, we
intend to enlarge the sample size and add a validation cohort study
as well as more experiments may be performed to demonstrate the
role that FPR2 plays in ovarian cancer.
The acute phase protein SAA is a biomarker for
ovarian tumorigenesis and prognosis (15,23).
Liang et al reported that SAA acts as an agonist for FPR2.
In mouse neutrophils, SAA binds FPR2 to induce calcium flux and
chemotaxis (16). As reported by
Sodin-Semrl et al, FPR2 was shown to participate in
SAA-induced release of IL-8, IL-6, and MMP-3 proteins as well as
upregulation of NF-κB and AP-1 DNA binding activity in
fibroblast-like synoviocytes (24).
SAA and FPR2 mediate the migration, adhesion and tissue
infiltration of inflammatory cells, and these physiological
processes can be adapted for tumor development in the study of
Badolato et al (25). In the
present study, we found that SAA reduced the FPR2 mRNA expression
levels as assessed by RT-qPCR, whereas the results of the wound
healing assay revealed that SAA may stimulate the migration of
SKOV3 cells. However, the migratory potential was significantly
decreased upon FPR2 knockdown; thus, we suggest SAA may utilize
FPR2 molecules expressed on the membranes of ovarian cancer cells.
We did not note any other studies similar to ours; however, there
are similar studies that we may use to illustrate the results. Aβ4
is also a chemotactic agonist for FPR2. According to Yazawa et
al, when FPR2/293 cells were co-cultured with Aβ4, Aβ4/FPR2
complexes could be detected in the cytoplasmic region and no FPR2
could be detected on the cell surface (26). It is a common feature for G
protein-coupled receptors (GPCRs) to undergo internalization after
ligand binding, which may involve different pathways. GPCR
expression levels are a balance of three highly regulated, dynamic
intracellular trafficking processes, called export, internalization
and degradation (27). Thus, we
hypothesized that incubation of ovarian cancer cells with continued
SAA may induce the internalization of FPR2 in association with SAA
and subsequently result in the degradation of FPR2. Actually, we
focused on only the first small step in this hypothesis, and there
is further research that is required.
Next, we studied the effect of FPR2 on the
angiogenesis of ovarian cancer cells. Angiogenesis occurs during
both inflammation and tumorigenesis, as sprouting new vessels
provide oxygen and nutrition for sites of wound healing and tumor
development. It is considered an essential process for oncogenesis.
According to Byrne et al, in ovarian cancer, angiogenesis is
associated with tumor growth, peritoneal implants and ascites
formation (28). In the present
study, the tube formation potential was decreased after FPR2
knockdown, which suggests that FPR2 contributes to angiogenesis to
some extent, whereas SAA could stimulate tube formation regardless
of the FPR2 expression levels. This finding may be attributed to
FPR2 expression on the HUVEC plasma membrane. Tumor angiogenesis is
regulated by angiogenic stimulators such as the VEGF family and
fibroblast growth factors. Perren et al demonstrated that
anti-angiogenesis therapy is considered a new strategy for treating
ovarian cancer. Bevacizumab, a humanized anti-VEGF monoclonal
antibody, is the most widely studied therapy that was shown to
prolong the progression-free survival (PFS) of ovarian cancer
patients (29). In inflamed corneas
of mice, the SAA/FPR2/MMP pathway was reported to stimulate corneal
neovascularization (17). According
to Lu et al, SAA induced VEGFR2 expression and angiogenesis
via FPR2 on HUVECs and activated downstream MAPKs (30). In the present study, we found that
VEGF expression was downregulated upon FPR2 knockdown, which is
consistent with the results of the above-mentioned studies.
Our subsequent experiments indicated that STAT3,
NF-κB and MAPK1 may be mediated by FPR2 to promote SKOV3 cell
migration. It was suggested that pro-inflammatory cytokines such as
tumor necrosis factor and interleukins trigger signaling cascades
that directly or indirectly activate key transcription factors,
including AP-1, NF-κB, STAT3, YAP and Notch, that control tumor
promotion and progression, cell cycle, cell death,
dedifferentiation, stemness, motility and migration (31,32).
We found STAT3 mRNA was downregulated in FPR2-knockdown cells.
According to Kim et al, STAT3 is a transcription factor
located in the cytoplasm that transduces extracellular signals to
the nucleus. STAT3 modulates the transcription of a variety of
genes to regulate important biological functions, including cell
proliferation, differentiation, survival, angiogenesis and the
immune response (33), and plays a
prominent role in tumor growth and invasion. In the study of Cai
et al, STAT3 knockdown downregulated the expression of
oncogenes such as cyclin D1, survivin and VEGF, which led to tumor
suppression and apoptosis in ovarian cancer (34). Cattaneo et al suggested that
activation of FPR2 induces the phosphorylation of the
Y1313/Y1349/Y1356 residues of c-Met and triggers some of the
molecular responses elicited by c-Met/HGF binding, including the
STAT3, PLC-γ1/PKCα and PI3K/Akt pathways (35). These studies are in accordance with
our results to some extent. Our Simple Western results showed that
NF-κB protein was downregulated in FPR2-knockdown SKOV3 cells.
NF-κB is a nuclear transcription factor that plays an essential
role in inflammation, innate immunity and cancer. According to Kam
et al, FPR2 can activate NF-κB signaling via inhibitor-κB
kinase (IKK) phosphorylation, and they suggested that NF-κB may be
a potential therapeutic target for FPR2-related diseases (36). These data combined with our results
indicate that FPR2 may promote ovarian cancer cell migration
through NF-κB. The present study showed that MAPK1 expression was
downregulated in FPR2-knockdown SKOV3 cells as shown by both
RT-qPCR and Simple Western. MAPKs act as key regulators in multiple
biological processes such as cell proliferation, death,
differentiation, migration and invasion. Previous studies have
reported that the MAPK pathway is activated in ovarian cancer.
Manzano et al confirmed that downregulation of CL100, which
is an endogenous dual-specificity phosphatase known to inhibit
MAPK, could stimulate human ovarian cancer progression by promoting
the MAPK pathway (37).
Additionally, the expression of receptors, including FPR2, BLTR and
CXCR1, on neutrophils was downregulated when p38MAPK was blocked,
which suggests that p38MAPK plays an essential role in neutrophil
chemotaxis (38). In the present
study, SAA exerted no obvious effects on stimulating MAPK
expression via FPR2. LL-37, another agonist peptide of FPR2, was
also demonstrated to activate the MAPK signaling pathway in ovarian
cancer cells despite inhibiting FPR2 (14). Previous research showed that Notch3
was preferentially upregulated and prevalent; furthermore, this
upregulation was significantly correlated with the poor clinical
outcomes of ovarian cancer patients (39). There is no research that indicates
the correlation between FPR2 and the Notch signaling pathway. In
the present study, we did not observe any evidence to prove the
relationship between SAA/FPR2 activation and Notch signaling.
In conclusion, our results demonstrated that FPR2
was significantly overexpressed in EOC and was positively
correlated with clinicopathological features including FIGO stage,
histological grade and high grade ovarian cancer. High FPR2
expression also indicated the poor prognosis of EOC patients.
Additionally, FPR2 knockdown decreased the migratory ability of
SKOV3 cells, which indicates that FPR2 is essential for invasion
and metastasis of EOC. We also observed that STAT3, NF-κB and MAPK1
expression may be affected by FPR2 function. In the future,
xenograft models may be used for further research and the
mechanisms of signaling molecules that are involved in the efffects
of FPR2 on ovarian cancer also require more detailed research.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen W: Cancer statistics: Updated cancer
burden in China. Chin J Cancer Res. 27:12015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Herzog TJ and Pothuri B: Ovarian cancer: A
focus on management of recurrent disease. Nat Clin Pract Oncol.
3:604–611. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Malpica A, Deavers MT, Lu K, Bodurka DC,
Atkinson EN, Gershenson DM and Silva EG: Grading ovarian serous
carcinoma using a two-tier system. Am J Surg Pathol. 28:496–504.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kurman RJ and Shih IeM: The origin and
pathogenesis of epithelial ovarian cancer: A proposed unifying
theory. Am J Surg Pathol. 34:433–443. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hunn J and Rodriguez GC: Ovarian cancer:
Etiology, risk factors, and epidemiology. Clin Obstet Gynecol.
55:3–23. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lin HW, Tu YY, Lin SY, Su WJ, Lin WL, Lin
WZ, Wu SC and Lai YL: Risk of ovarian cancer in women with pelvic
inflammatory disease: A population-based study. Lancet Oncol.
12:900–904. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Risch HA and Howe GR: Pelvic inflammatory
disease and the risk of epithelial ovarian cancer. Cancer Epidemiol
Biomarkers Prev. 4:447–451. 1995.PubMed/NCBI
|
|
9
|
Seidman JD, Sherman ME, Bell KA, Katabuchi
H, O'Leary TJ and Kurman RJ: Salpingitis, salpingoliths, and serous
tumors of the ovaries: Is there a connection? Int J Gynecol Pathol.
21:101–107. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ye RD, Boulay F, Wang JM, Dahlgren C,
Gerard C, Parmentier M, Serhan CN and Murphy PM: International
Union of Basic and Clinical Pharmacology. LXXIII. Nomenclature for
the formyl peptide receptor (FPR) family. Pharmacol Rev.
61:119–161. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Becker EL, Forouhar FA, Grunnet ML, Boulay
F, Tardif M, Bormann BJ, Sodja D, Ye RD, Woska JR Jr and Murphy PM:
Broad immunocytochemical localization of the formylpeptide receptor
in human organs, tissues, and cells. Cell Tissue Res. 292:129–135.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Migeotte I, Communi D and Parmentier M:
Formyl peptide receptors: A promiscuous subfamily of G
protein-coupled receptors controlling immune responses. Cytokine
Growth Factor Rev. 17:501–519. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li Y and Ye D: Molecular biology for
formyl peptide receptors in human diseases. J Mol Med. 91:781–789.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Coffelt SB, Tomchuck SL, Zwezdaryk KJ,
Danka ES and Scandurro AB: Leucine leucine-37 uses formyl peptide
receptor-like 1 to activate signal transduction pathways, stimulate
oncogenic gene expression, and enhance the invasiveness of ovarian
cancer cells. Mol Cancer Res. 7:907–915. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Urieli-Shoval S, Finci-Yeheskel Z, Dishon
S, Galinsky D, Linke RP, Ariel I, Levin M, Ben-Shachar I and Prus
D: Expression of serum amyloid a in human ovarian epithelial
tumors: Implication for a role in ovarian tumorigenesis. J
Histochem Cytochem. 58:1015–1023. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liang TS, Wang JM, Murphy PM and Gao JL:
Serum amyloid A is a chemotactic agonist at FPR2, a low-affinity
N-formylpeptide receptor on mouse neutrophils. Biochem
Biophys Res Commun. 270:331–335. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ren SW, Qi X, Jia CK and Wang YQ: Serum
amyloid A and pairing formyl peptide receptor 2 are expressed in
corneas and involved in inflammation-mediated neovascularization.
Int J Ophthalmol. 7:187–193. 2014.PubMed/NCBI
|
|
18
|
Yang M, Liu F and Higuchi K, Sawashita J,
Fu X, Zhang L, Zhang L, Fu L, Tong Z and Higuchi K: Serum amyloid A
expression in the breast cancer tissue is associated with poor
prognosis. Oncotarget. 7:35843–35852. 2016.PubMed/NCBI
|
|
19
|
Romero I and Bast RC Jr: Minireview: Human
ovarian cancer: Biology, current management, and paths to
personalizing therapy. Endocrinology. 153:1593–1602. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Oldekamp S, Pscheidl S, Kress E, Soehnlein
O, Jansen S, Pufe T, Wang JM, Tauber SC and Brandenburg LO: Lack of
formyl peptide receptor 1 and 2 leads to more severe inflammation
and higher mortality in mice with of pneumococcal meningitis.
Immunology. 143:447–461. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xiang Y, Yao X, Chen K, Wang X, Zhou J,
Gong W, Yoshimura T, Huang J, Wang R, Wu Y, et al: The G-protein
coupled chemoattractant receptor FPR2 promotes malignant phenotype
of human colon cancer cells. Am J Cancer Res. 6:2599–2610.
2016.PubMed/NCBI
|
|
22
|
Khau T, Langenbach SY, Schuliga M, Harris
T, Johnstone CN, Anderson RL and Stewart AG: Annexin-1 signals
mitogen-stimulated breast tumor cell proliferation by activation of
the formyl peptide receptors (FPRs) 1 and 2. FASEB J. 25:483–496.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang J, Sharma A, Ghamande SA, Bush S,
Ferris D, Zhi W, He M, Wang M, Wang X, Miller E, et al: Serum
protein profile at remission can accurately assess therapeutic
outcomes and survival for serous ovarian cancer. PLoS One.
8:e783932013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sodin-Semrl S, Spagnolo A, Mikus R,
Barbaro B, Varga J and Fiore S: Opposing regulation of
interleukin-8 and NF-kappaB responses by lipoxin A4 and serum
amyloid A via the common lipoxin A receptor. Int J Immunopathol
Pharmacol. 17:145–156. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Badolato R, Wang JM, Murphy WJ, Lloyd AR,
Michiel DF, Bausserman LL, Kelvin DJ and Oppenheim JJ: Serum
amyloid A is a chemoattractant: Induction of migration, adhesion,
and tissue infiltration of monocytes and polymorphonuclear
leukocytes. J Exp Med. 180:203–209. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yazawa H, Yu ZX, Takeda, Le Y, Gong W,
Ferrans VJ, Oppenheim JJ, Li CC and Wang JM: β amyloid peptide
(Aβ42) is internalized via the G-protein-coupled receptor FPRL1 and
forms fibrillar aggregates in macrophages. FASEB J. 15:2454–2462.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Duvernay MT, Filipeanu CM and Wu G: The
regulatory mechanisms of export trafficking of G protein-coupled
receptors. Cell Signal. 17:1457–1465. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Byrne AT, Ross L, Holash J, Nakanishi M,
Hu L, Hofmann JI, Yancopoulos GD and Jaffe RB: Vascular endothelial
growth factor-trap decreases tumor burden, inhibits ascites, and
causes dramatic vascular remodeling in an ovarian cancer model.
Clin Cancer Res. 9:5721–5728. 2003.PubMed/NCBI
|
|
29
|
Perren TJ, Swart AM, Pfisterer J,
Ledermann JA, Pujade-Lauraine E, Kristensen G, Carey MS, Beale P,
Cervantes A, Kurzeder C, et al: ICON7 Investigators: A phase 3
trial of bevacizumab in ovarian cancer. N Engl J Med.
365:2484–2496. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lu Q, Quan W, Wu J, Zhang X, Ma W, Pang L
and Li D: Effect of antibacterial peptide hCAP18/LL-37 on ovarian
cancer microenvironment and the regulatory mechanism of its
expression. Zhonghua Zhong Liu Za Zhi. 37:725–730. 2015.(In
Chinese). PubMed/NCBI
|
|
31
|
Pesic M and Greten FR: Inflammation and
cancer: Tissue regeneration gone awry. Curr Opin Cell Biol.
43:55–61. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Karin M and Clevers H: Reparative
inflammation takes charge of tissue regeneration. Nature.
529:307–315. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kim BH, Yi EH and Ye SK: Signal transducer
and activator of transcription 3 as a therapeutic target for cancer
and the tumor microenvironment. Arch Pharm Res. 39:1085–1099. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cai L, Zhang G, Tong X, You Q, An Y, Wang
Y, Guo L, Wang T, Zhu D and Zheng J: Growth inhibition of human
ovarian cancer cells by blocking STAT3 activation with small
interfering RNA. Eur J Obstet Gynecol Reprod Biol. 148:73–80. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cattaneo F, Parisi M and Ammendola R:
WKYMVm-induced cross-talk between FPR2 and HGF receptor in human
prostate epithelial cell line PNT1A. FEBS Lett. 587:1536–1542.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kam AY, Liu AM and Wong YH: Formyl
peptide-receptor like-1 requires lipid raft and extracellular
signal-regulated protein kinase to activate inhibitor-kappa B
kinase in human U87 astrocytoma cells. J Neurochem. 103:1553–1566.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Manzano RG, Montuenga LM, Dayton M, Dent
P, Kinoshita I, Vicent S, Gardner GJ, Nguyen P, Choi YH, Trepel J,
et al: CL100 expression is down-regulated in advanced epithelial
ovarian cancer and its re-expression decreases its malignant
potential. Oncogene. 21:4435–4447. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kim D and Haynes CL: The role of p38 MAPK
in neutrophil functions: Single cell chemotaxis and surface marker
expression. Analyst. 138:6826–6833. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Park JT, Li M, Nakayama K, Mao TL,
Davidson B, Zhang Z, Kurman RJ, Eberhart CG, Shih IeM and Wang TL:
Notch3 gene amplification in ovarian cancer. Cancer Res.
66:6312–6318. 2006. View Article : Google Scholar : PubMed/NCBI
|