Introduction
Cutaneous malignant melanoma (CMM) is a highly
malignant and invasive skin tumor that has a high incidence rate in
Japan, Europe and other countries (1). CMM occurs in the surface of the skin
or in mucous membranes, including the leptomeninges and the choroid
(2). According to the origin and
site of melanoma, CMM includes four pathological types, namely,
superficial spreading type melanoma, acral lentiginous melanoma,
nodular melanoma and lentigo malignant melanoma (3). Clinical data demonstrate that melanoma
is the fifth most common cancer in males and fifth most common
cancer in females in the US (4).
Even though the incidence rates are lower in Asian countries, the
incidence rate of melanoma has reached to 1/100,000 in China
(5).
The occurrence of malignant melanoma is a complex
process with multiple factors and steps (6). A previous study has reported that the
cause of malignant melanoma includes sun exposure, ultraviolet
light (UV) exposure, ionizing radiation and genetic factors
(7). UV rays can penetrate the
skin's surface and cause the destruction of collagen and elastic
proteins in the skin, leading to malignant melanoma. Furthermore,
sex, age, skin color and race are factors that are associated with
incidence of malignant melanoma (8). For Caucasians, superficial diffuse
melanoma is the most frequent melanoma and accounts for ~90% of all
malignant melanoma cases. For persons of Asian and African descent,
freckle-like melanoma accounts for 50–70% of all patients with
malignant melanoma (9,10). In addition, malignant melanoma may
be inherited, and it has been suggested that 10% of patients with
melanoma have a family history (11).
To date, vemurafenib, ipilimumab, sibutramine and
other targeted drugs have been recommended for the treatment of
malignant melanoma. However, malignant melanoma is not completely
cured by these agents (12).
Metastasis may occur during the early stages of cutaneous malignant
melanoma and prognosis is often poor. Metastasis is also an
important cause of high mortality. In addition, multidrug
resistance has been reported in the chemotherapy of melanoma,
leading to a very poor prognosis (13); thus, there is a need to identify
novel therapeutic drugs and drug targets for melanoma.
Bufotalin, a compound of the traditional Chinese
medicine Venenum bufonis, is isolated from the dried secretions of
the auricular and skin glands of Bufo gargarizans Cantor
(14). Venenum bufonis has been
used to treat various conditions, including cardiac illness, pain
and cancer, in China and other Asian countries. Venenum bufonis has
been historically used as an oral or topical anti-neoplastic drug
in China (15). A previous study
has shown that bufotalin is a bufadienolide, a C-24 steroidal
compound, and one of the main active components of Venenum bufonis
(16). In vitro
investigations have demonstrated that bufotalin can effectively
inhibit the proliferation of hepatoma cells, gastric cancer cells,
and sarcoma cells (17). Bufotalin
also inhibits the growth of liver cancer and sarcoma in animal
models of transplanted tumors, and without obvious side-effects;
however, the antitumor effect of bufotalin on malignant melanoma
has not been investigated to date.
The present study examined the in vitro
effects of bufotalin on human malignant melanoma A375 cells and
explored its underlying molecular mechanism. We found that
bufotalin remarkably induced cell cycle arrest at the G2/M phase
and cell apoptosis, which in turn inhibited cellular proliferation.
These findings suggest that bufotalin may be a potential
chemotherapeutic agent for malignant melanoma treatment.
Materials and methods
Cell culture
Human malignant melanoma A375 cells (cat. no.
SCSP-533) were purchased from Cell Bank, Typical Culture
Preservation Commission, Chinese Academy of Sciences (Shanghai,
China). The normal human keratinocyte cell line HaCaT (cat. no.
CL-0090; STR profiling authentication) was purchased from Procell
Life Science & Technology Co., Ltd. (Wuhan, China). The cells
were cultured with Dulbecco's modified Eagles' medium
(DMEM)/high-glucose basal medium (HyClone; GE Healthcare Life
Sciences, Logan, UT, USA), containing 10% fetal calf serum (Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA), 1% sodium
pyruvate (Beijing Solarbio Science & Technology Co., Ltd.,
Beijing, China), 0.1 U/l penicillin, and 0.1 µg/l streptomycin
(Beijing Solarbio Science & Technology Co., Ltd.) and incubated
in a 5% CO2 incubator at 37°C. Fetal calf serum was
inactivated in a 56°C water bath for 30 min before use.
MTT assay
The effect of bufotalin on the viability of A375 and
HaCaT cells was determined by an MTT assay. The cells in suspension
(1×104 cells in 100 µl DMEM) were seeded into 96-well
plates. After 24 h of incubation, different concentrations of
bufotalin (cat. no. ASB-00002466-005; J&K Scientific Ltd.,
Beijing, China) were added, and the cells were further cultured for
24 and 48 h. Then, 10 µl MTT solution (5 mg/ml) was added into each
well, and the cells were incubated for an additional 2 h. The
resulting formazan crystals were dissolved using 150 µl dimethyl
sulfoxide (DMSO). Absorbance was measured at a wavelength of 490 nm
using a microplate reader (Bio-Rad Laboratories, Inc., Hercules,
CA, USA). The absorbance of the cells treated with DMEM medium
containing 0.1% DMSO was regarded as the control group (survival
rate 100%). IC50 represents the drug concentration
causing a 50% decrease in survival rate.
Colony formation assay
The A375 cells were trypsinized, collected, and
diluted into 300 cells in 1 ml DMEM medium. The cell suspension was
seeded into a 6-well plate at 300 cells/well. After culturing for
24 h, the cells were treated with different concentrations of
bufotalin for 24 h. Then, the treated cells were incubated with
DMEM complete medium at 37°C for 14 days. Finally, cells were fixed
in 4% paraformaldehyde solution for 15 min at room temperature, and
the cells were stained with Giemsa solution at room temperature for
10 min, then examined and images under a light microscope. The
colonies containing >50 cells were counted and used to calculate
the colony formation rate.
Hoechst 33258 staining
The A375 cells (6×105 cells) were seeded
onto sterile cover slips and placed into 6-well plates and cultured
for 24 h. Then, the cells were treated with different
concentrations of bufotalin. After 12 h of treatment, the cells
were fixed in 4% paraformaldehyde solution for 15 min at room
temperature, washed twice with PBS, and then the cells were stained
with Hoechst 33258 (cat. no. C1018; Beyotime Institute of
Biotechnology, Haimen, China) at room temperature for 10 min.
Stained nuclei were observed, and images were captured using a
fluorescence microscope. Apoptotic cells (cells exhibiting nuclear
fragmentation and/or with distinct apoptotic bodies) were counted,
and the percentage of apoptotic cells (number of apoptotic
cells/the total number of cells × 100) was calculated.
Cell cycle analysis
Cell cycle distribution of the bufotalin-treated
A375 cells was assessed by flow cytometry. Briefly, the
bufotalin-treated A375 cells (~8×105 cells/group) were
collected and fixed in 70% ethanol at 4°C overnight. After
fixation, the cells were centrifuged at 800 × g for 5 min to remove
the ethanol. Then, the cells were suspended in 500 µl PBS
containing 0.02 mg/ml propidium iodide (PI) and 0.1 mg/ml
ribonuclease A (RNase A) in darkness at 37°C for 30 min. The
fluorescence emitted by the PI-DNA complexes was measured using an
Epics XL flow cytometry (Beckman Coulter, Inc., Brea, CA, USA).
Cell distribution in different phases of the cell cycle was
analyzed using WinMDI 2.8 software (The Scripps Research Institute,
La Jolla, CA, USA).
Annexin V-fluorescein isothiocyanate
(FITC)/PI double staining assay
Annexin V-FITC/PI double staining assay (Annexin
V-FITC Apoptosis Detection Kit; cat. no. CA1020; Beijing Solarbio
Science & Technology Co., Ltd.) was performed according to the
manufacturer's instructions. Briefly, A375 cells (~8×105
cells/group) were harvested at 24 h after bufotalin treatment and
then incubated in 100 µl labeling solution, which consisted of 5 µl
Annexin V-FITC, 10 µl PI, 10 µl 10X binding buffer and 75 µl
H2O in darkness at room temperature for 15 min. Then,
400 µl 1X binding buffer were added to stop the staining reaction.
The stained cells were detected using FL1 and FL3 fluorescence
channels on a FACSort flow cytometry system (FACSCanto II; Becton,
Dickinson and Company, Franklin Lakes, NJ, USA), and the
percentages of apoptotic cells in each group were analyzed using BD
FACSDiva software (version 6.1.3; BD Biosciences, Becton, Dickinson
and Company).
Western blot analysis
A375 cells (1×106/well) were seeded in
100-mm culture dishes and treated with different concentrations of
bufotalin for 24 h. Following treatment, cells were collected and
incubated in cell lysis buffer (20 mM Tris-HCl, 150 mM NaCl, 1 mM
EDTA, 1% Triton X-100, 2.5 mM sodium pyrophosphate, 0.1 mM
phenylmethanesulfonylfluoride, 0.1 mM sodium orthovanadate, 0.5 mM
dithiothreitol and 10X protease inhibitor; pH 6.8) on ice for 30
min. Cells were vortexed for 45 sec and then centrifuged at 10,000
× g at 4°C for 10 min. The supernatant was collected and stored at
−20°C. Protein concentrations were determined using a bicinchoninic
acid protein assay kit (Beijing Solarbio Science & Technology
Co., Ltd.). Cell lysates with a protein content of 40 mg were mixed
with an equal volume of sodium dodecyl sulfate (SDS) loading dye
(2% SDS, 10% sucrose, 0.002% bromophenol blue, 5% 2-mercaptoethanol
and 625 mM Tris; pH 6.8) and subsequently separated by SDS-PAGE on
12.5% gels along with a rainbow-colored protein molecular marker
(Beijing Solarbio Science & Technology Co., Ltd.). After
running for 2 h at 110V, the proteins were transferred to
polyvinylidene fluoride membranes (EMD Millipore, Billerica, MA,
USA) using a Trans-blots SD semi-dry transfer cell (Bio-Rad
Laboratories, Inc.). The membranes were blocked with 5% bovine
serum albumin (BSA; cat. no. A8010; Beijing Solarbio Science &
Technology Co., Ltd.) in Tris-buffered saline containing 0.1%
Tween-20 (TBS-T) for 60 min before being probed with primary
antibodies [protein kinase B (AKT), cat. no. ab38449, 1:500;
phosphorylated (p)-AKT, cat. no. ab38449, 1:500; ATM
serine/threonine kinase (ATM), cat. no. ab78, 1:1,000; checkpoint
kinase 2 (Chk2), cat. no. ab47433, 1:1,000; cell division cycle 25C
(CDC25C), cat. no. ab32444, 1:1,000; cyclin-dependent kinase 1
(CDK1), cat. no. ab18, 1:1,000; cyclin B, cat. no. ab181593,
1:1,000; BCL2-associated X, apoptosis regulator (BAX), cat. no.
ab32503, 1:1,000; B cell lymphoma-2 (BCL-2), cat. no. ab32124,
1:1,000; caspase-3, cat. no. ab13585, 1:1,000; caspase-9, cat. no.
ab202068, 1:1,000; all Abcam, Cambridge, UK] containing 5% BSA at
4°C overnight. Subsequently, the membranes were washed with TBS-T
buffer for 45 min and then probed with appropriate secondary
antibodies (goat anti-mouse IgG H&L, cat. no. ab205719,
1:20,000; or goat anti-rabbit IgG H&L, cat. no. ab6721,
1:20,000; Abcam) conjugated with horseradish peroxidase for 1 h at
room temperature. Immunoreactive bands were visualized with
enhanced chemiluminescence detection reagents (Invitrogen; Thermo
Fisher Scientific, Inc.) using a film processor (Kodak, Rochester,
NY, USA), and the gray-scale values of each image were analyzed
using Image-Pro Plus 6.0 (IPP6) software.
Statistical analysis
The experiments were conducted at least three times.
All the data are expressed as the mean ± standard deviation.
Analyses were performed using the SPSS 21.0 software package
(version 21.0; IBM Corp., Armonk, NY, USA). Student's t-test, a
one-way analysis of variance (ANOVA) or a two-way ANOVA was used to
calculate statistical differences, respectively. Tukey's post hoc
tests were undertaken following a significant one-way ANOVA, and
Bonferroni's post hoc tests were undertaken following a significant
two-way ANOVA. P<0.05 was considered to indicate a statistically
significant difference.
Results
In vitro anti-melanoma effects of
bufotalin
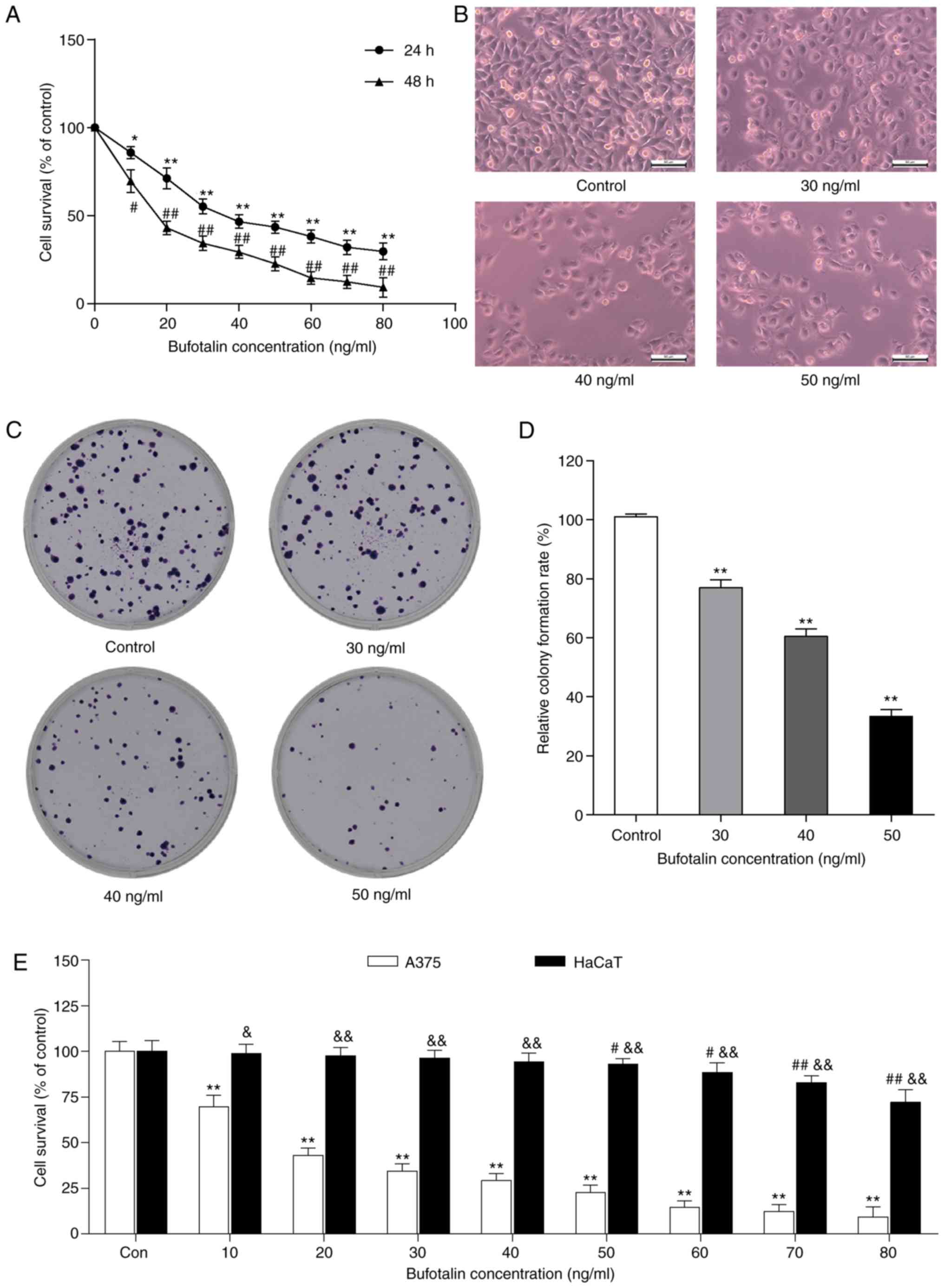
To examine the antiproliferative effect of bufotalin
on human malignant melanoma A375 cells, the cells were treated with
or without various concentrations of bufotalin for 24 and 48 h,
then the viability of bufotalin-treated cells was determined by the
MTT assay. Bufotalin (10–80 ng/ml) significantly inhibited A375
cell proliferation, with an IC50 value of 40 ng/ml after
24 h treatment (Fig. 1A).
Furthermore, the proliferation rate was significantly lower in A375
cells after 48 h treatment with bufotalin than that in A375 cells
after 24 h treatment with the same concentration (10–80 ng/ml) of
bufotalin (Fig. 1A). Subsequently,
according to the IC50 value of bufotalin (40 ng/ml), we
selected 30, 40 and 50 ng/ml as the concentrations of bufotalin
used in the next studies. Additionally, light microscopy indicated
changes in cell number and cell morphology. There was a marked
decrease in the number of A375 cells after treatment with
bufotalin, and some cells began to shrink and deform, and even
detach from the dish surface (Fig.
1B). In addition, bufotalin significantly inhibited A375 cell
colony formation at concentrations of 30, 40 and 50 ng/ml (Fig. 1C and D). Bufotalin also effectively
inhibited HaCaT cell proliferation at 50–80 ng/ml after 24 h
treatment, whereas the cytotoxicity of bufotalin on HaCaT cells is
significantly lower than that on A375 cells (Fig. 1E).
Bufotalin induces cell cycle arrest at
the G2/M phase in A375 cells
To explore the mechanism of action underlying the
bufotalin-mediated antiproliferative effect, the cell cycle
distribution profile of bufotalin-treated A375 cells was
investigated. After exposure of A375 cells to bufotalin for 24 h, a
marked accumulation of cells at the G2/M phase compared with the
control group was observed (Fig. 2A and
B). The protein levels of G2/M phase regulatory proteins, CDK1
and cyclin B, were subsequently analyzed by western blot analysis.
The results demonstrated that the CDK1 protein expression levels in
bufotalin-treated A375 cells significantly decreased compared to
the control group, whereas no change in cyclin B protein expression
levels in bufotalin-treated A375 cells was detected (Fig. 2C and D). These results indicated
that bufotalin effectively inhibited the expression of CDK1, but
not cyclin B.
Effects of bufotalin on
ATM/Chk2/CDC25C signaling
Considering the cell cycle arrest at the G2/M phase
and the decreased ratio of CDK1/cyclin B in bufotalin-treated A375
cells, changes in the levels of the components of the
ATM/Chk2/CDC25C signaling pathway, which regulates cell
transmission through the G2/M phase (18), were examined by western blot
analysis. The results demonstrated that the protein levels of ATM
and Ckh2 were significantly higher, and that of CDC25C was
significantly lower in bufotalin-treated A375 cells compared to the
control group (Fig. 3A and B).
Bufotalin promotes cell apoptosis in
A375 cells
In the above experiment, bufotalin effectively
inhibited A375 cell proliferation through G2/M cell cycle arrest.
Additionally, the changes in A375 cell morphology that were induced
by bufotalin were similar to those observed during apoptosis. To
explore whether this proliferation inhibition is associated with
cell apoptosis, the bufotalin-treated A375 cells were stained with
Hoechst 33258 and examined by fluorescence microscopy. The typical
apoptosis characteristics, nuclear shrinkage, DNA condensation, and
apoptotic bodies, were observed in bufotalin-treated A375 cells
(Fig. 4A and B). Furthermore, the
apoptotic rate of bufotalin-treated A375 cells was determined using
an Annexin V FITC/PI double staining assay and flow cytometry
analysis. The percentage of apoptotic cells was significantly
increased with increasing bufotalin concentration (Fig. 4C and D). These results suggested
that bufotalin induced cell apoptosis in A375 cells, leading to an
antiproliferative effect.
Effects of bufotalin on
apoptosis-associated signaling pathways
The levels of cell apoptosis-associated proteins,
BCL-2, BAX, caspase-3 and −9, were detected in A375 cells by
western blot analysis. The level of BCL-2 was significantly
downregulated and the levels of BAX, caspase-3 and −9 were markedly
upregulated in bufotalin-treated A375 cells compared with the
control group (Fig. 5A and B).
Additionally, considering the regulatory effect of the activation
of AKT on caspase-3/-9, the levels of AKT and p-AKT in
bufotalin-treated A375 cells. Bufotalin markedly decreased p-AKT
levels and the ratio of p-AKT/AKT, whereas no significant change in
AKT expression levels was observed in A375 cells following
bufotalin treatment (Fig. 5C and
D).
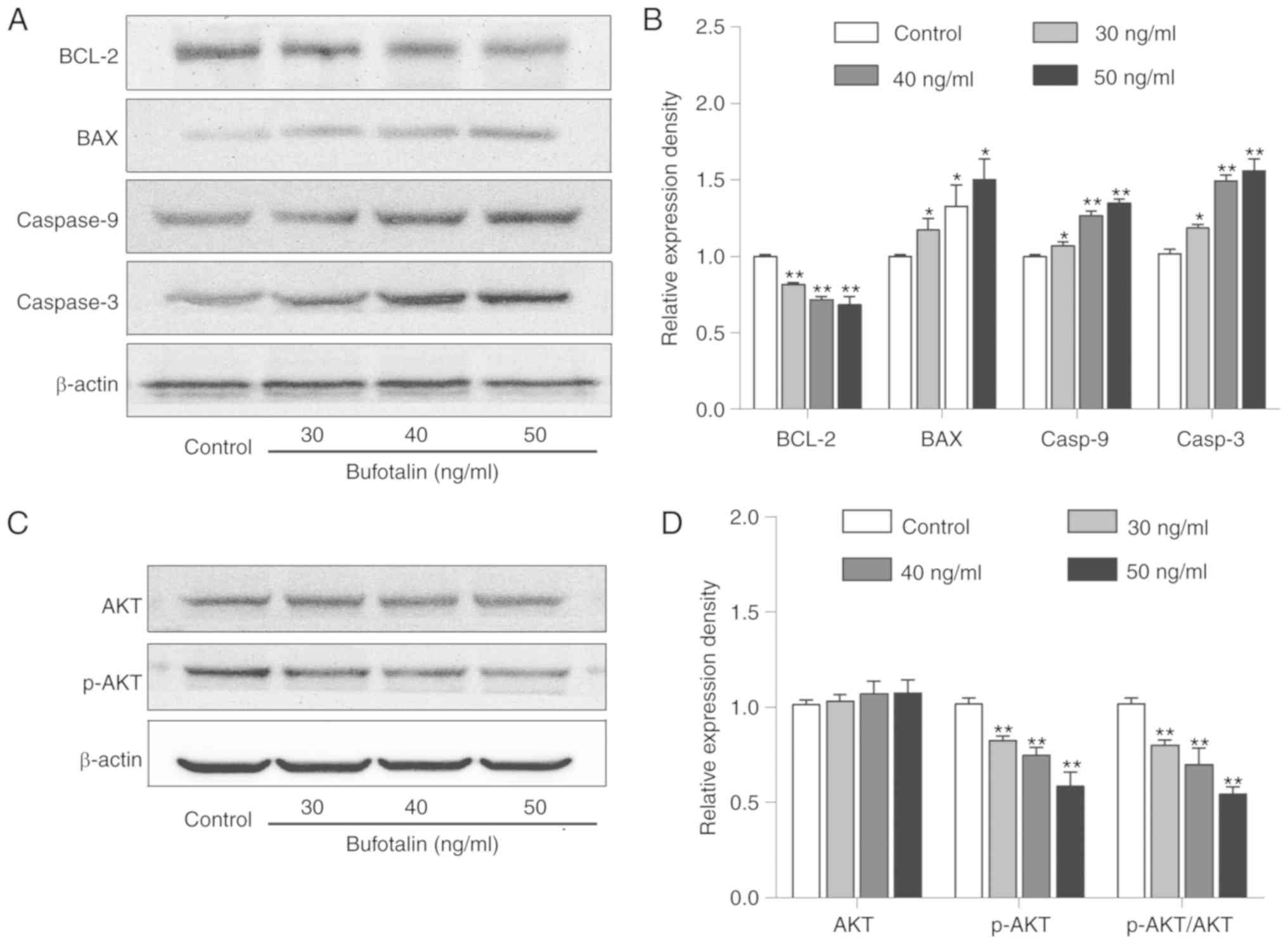 | Figure 5.Effects of bufotalin on the protein
levels of apoptosis-associated signaling molecules. (A) The protein
levels of BAX, BCL-2, caspase-3 and −9 in bufotalin-treated A375
cells were assessed by western blotting. (B) Statistical analysis
of the relative protein levels of BAX, BCL-2, caspase-3 and −9 in
bufotalin-treated A375 cells. (C) Western blot analysis for the
protein levels of AKT and p-AKT in bufotalin-treated A375 cells.
(D) Statistical analysis of the relative protein levels of AKT and
p-AKT. All data are presented as the mean ± standard deviation of
three independent experiments. *P<0.05, **P<0.01 vs. control
group. BAX, BCL2-associated X, apoptosis regulator; BCL-2, B cell
lymphoma-2; AKT, protein kinase B; p-, phosphorylated. |
Discussion
It has been reported that bufotalin, a
bufadienolide, has strong antitumor efficacy in several tumor cell
lines, including gastric cancer, liver cancer, hepatoma and sarcoma
cells (17,19). The present study explored the effect
of bufotalin on human malignant melanoma A375 cell proliferation
and its associated mechanism. Bufotalin effectively suppressed the
proliferation of melanoma A375 cells by arresting them at the G2/M
phase and inducing cell apoptosis.
The cell cycle, which is a complex cell division
process, has an important role in cell proliferation. Cyclins and
CDKs are the direct regulatory factors of the cell cycle, which
have important roles in normal and tumor cells (20). Cyclin B, in complex with CDK1,
triggers mitotic entry and is a key modulator of the G2/M
checkpoint (21). It has been
reported that coffee oil-algae oil nanoemulsions upregulated the
protein level of cyclin B and downregulated the protein level of
CDK1, subsequently causing cell cycle arrest of B16-F10 cells at
the G2/M phase (22). By contrast,
immature colon carcinoma transcript-1 knockdown downregulated the
levels of cyclin B and CDK1, arresting human breast cancer ZR-75-30
cells at G2/M phase (23). Thus, it
is uncertain whether the level of cyclin B is
upregulated/downregulated in G2/M phase-arrested cells. However,
several studies showed that the downregulation of CDK1/cyclin B
complex/activity is associated with cell cycle arrest at G2/M phase
(24–26). Consistent with these studies,
bufotalin decreased the level of CDK1, which could downregulate the
level of CDK1/cyclin B complex, leading to cell cycle arrest at
G2/M phase in bufotalin-treated A375 cells in the present
study.
Several studies have demonstrated that cells with
DNA damage do not pass through G2/M phase, resulting in an increase
in the number of cells arrested at the G2/M phase (27). Thus, the appearance of G2/M cell
cycle arrest in bufotalin-treated A375 cells suggests that the
cells may have incurred DNA damage and repair. It has been reported
that the signal pathway mediated by the ATM/ATR serine/threonine
kinase protein repairs damaged DNA (28). DNA damage caused by various causes
can activate ATM (29), which
subsequently upregulates Chk2 and downregulates CDC25C, leading to
cell cycle arrest at the G2/M phase (30). The present data is consistent with
the above studies, wherein bufotalin upregulates ATM in A375 cells,
followed by an increase in Chk2 and decrease in CDC25C expression,
leading to cell cycle arrest at the G2-M phase.
It has been reported that bufotalin can induce cell
apoptosis in HeLa cells and drug-resistant HepG2 cells by
regulating the expression of caspase family proteins (17). In the current study, the levels of
caspase-3 and −9 were significantly increased in A375 cells after
bufotalin treatment for 24 h. Furthermore, changes in mitochondrial
apoptosis-associated proteins, BCL-2 and BAX were observed in
bufotalin-treated A375 cells, indicating that bufotalin may induce
apoptosis via the mitochondrial apoptosis pathway. Additionally, it
has been reported that the upregulation of AKT, an anti-apoptotic
protein (31), in tumor cells leads
to rapid growth of tumor cells, and activated AKT is also an
inhibitor of caspase-3 and −9 (32). Bufotalin effectively inhibited the
phosphorylation of AKT in A375 cells. This result indicated that
bufotalin inhibits AKT phosphorylation and subsequently increases
the levels of downstream executioner caspases (caspase-3 and −9),
leading to A375 cell apoptosis.
Notably, there are some safety concerns regarding
the treatment of melanoma using natural compounds, even when these
traditional medicines have been used for thousands of years. The
data of the present study demonstrated that bufotalin could inhibit
HaCaT cell proliferation at 50–80 ng/ml, but bufotalin
significantly inhibited A375 cell proliferation at a lower dose (10
ng/ml). It is suggested that bufotalin has a certain degree of
selectivity against malignant melanoma compared with normal skin
cells.
Additionally, there are certain limitations to this
study. The anti-melanoma effect of bufotalin was only evaluated in
one human melanoma cell line, A375, and thus other skin cancer
cells (e.g. mouse malignant melanoma B16 cells) will be using to
further explore the therapeutic effect of bufotalin in subsequent
studies. Furthermore, the effect of bufotalin in an
A375-xenografted model in nude mice has not been determined.
In conclusion, the results demonstrated that
bufotalin simultaneously induces cell cycle arrest and apoptosis in
A375 cells. Further studies revealed that bufotalin could induce
cell cycle arrest at G2/M phase by upregulating ATM and Chk2, and
downregulating Cdc25C. Bufotalin also induces cell apoptosis via
the mitochondrial apoptosis pathway and inhibition of AKT
phosphorylation. The results provide a strong basis for developing
bufotalin into a potential therapeutic agent for the treatment of
cutaneous malignant melanoma.
Acknowledgements
Not applicable.
Funding
The National Natural Science Foundation of China
(grant nos. 81872162 and 81602556 to DL; grant no. 31870338 to QZ),
the Natural Science Foundation of Shandong Province (grant no.
ZR2017JL030 to DL), Taishan Scholars Construction Engineering of
Shandong Province (to DL), Yantai High-End Talent Introduction Plan
‘Double Hundred’ (to DL), the Scientific Research Foundation of
Binzhou Medical University (grant no. BY2016KYQD01 to DL) and the
Dominant Disciplines' Talent Team Development Scheme of Higher
Education of Shandong Province (to DL) supported this study.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
ZP, DL and QZ designed the study, acquired the data
and wrote the manuscript; CQ, YC, XC and XL collected cell samples
for Hoechst 33258 staining, cell cycle and western blot analyses;
WH, WX, LY and PL interpreted and analyzed the data; DL and QZ
revised and approved the final version of the manuscript. All
authors read and approved the manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Saida T: Melanoma and Non-melanoma skin
cancers. Gan To Kagaku Ryoho. 45:612–613. 2018.(In Japanese).
PubMed/NCBI
|
|
2
|
Gallego Y, Mendicute J, Ruiz M, Ruiz I and
Ubeda M: Melanocytoma of the ciliary body. Arch Soc Esp Oftalmol.
80:109–112. 2005.(In Spanish). View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lang J and MacKie RM: Prevalence of exon
15 BRAF mutations in primary melanoma of the superficial
spreading, nodular, acral, and lentigo maligna subtypes. J Invest
Dermatol. 125:575–579. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nakamura Y II: Diagnosis and treatment for
nail Apparatus (Subungual) Melanoma]. Gan to Kagaku Ryoho.
45:619–621. 2018.(In Japanese). PubMed/NCBI
|
|
5
|
Chen F, Zhang X, Ma K, Madajewski B,
Benezra M, Zhang L, Phillips E, Turker MZ, Gallazzi F,
Penate-Medina O, et al: Melanocortin-1 receptor-targeting
ultrasmall silica nanoparticles for dual-modality human melanoma
imaging. ACS Appl Mater Interfaces. 10:4379–4393. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Guo W, Wang H, Yang Y, Guo S, Zhang W, Liu
Y, Yi X, Ma J, Zhao T, Liu L, et al: Down-regulated miR-23a
contributes to the metastasis of cutaneous melanoma by promoting
autophagy. Theranostics. 7:2231–2249. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ye Z, Dong H, Li Y, Ma T, Huang H, Leong
HS, Eckel-Passow J, Kocher JA, Liang H and Wang L: Prevalent
homozygous deletions of type I interferon and defensin genes in
human cancers associate with immunotherapy resistance. Clin Cancer
Res. 24:3299–3308. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen X, Chen P, Chen SS, Ma T, Shi G, Zhou
Y, Li J and Sheng L: miR-106b-5p promotes cell cycle
progression of malignant melanoma by targeting PTEN. Oncol
Rep. 39:331–337. 2018.PubMed/NCBI
|
|
9
|
Jiang L, Tu Y, Hu X, Bao A, Chen H, Ma X,
Doyle T, Shi H and Cheng Z: Pilot study of 64Cu(I) for
PET imaging of melanoma. Sci Rep. 7:25742017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yu J, Wu X, Yan J, Yu H, Xu L, Chi Z,
Sheng X, Si L, Cui C, Dai J, et al: Anti-GD2/4-1BB chimeric antigen
receptor T cell therapy for the treatment of Chinese melanoma
patients. J Hematol Oncol. 11:12018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sonoda Y, Kijima M and Uchida H: Melanoma
of the choroid. Nihon Ganka Kiyo. 19:828–831. 1968.(In Japanese).
PubMed/NCBI
|
|
12
|
Liu SM, Lin CH, Lu J, Lin IY, Tsai MS,
Chen MH and Ma N: miR-596 modulates melanoma growth by regulating
cell survival and death. J Invest Dermatol. 138:911–921. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ma Y, Cheng X, Wang F, Pan J, Liu J, Chen
H, Wang Y and Cai L: ING4 inhibits proliferation and induces
apoptosis in human melanoma A375 cells via the Fas/Caspase-8
apoptosis pathway. Dermatology. 232:265–272. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lin S, Lv J, Peng P, Cai C, Deng J, Deng
H, Li X and Tang X: Bufadienolides induce p53-mediated apoptosis in
esophageal squamous cell carcinoma cells in vitro and in
vivo. Oncol Lett. 15:1566–1572. 2018.PubMed/NCBI
|
|
15
|
Raymond-Hamet, . Toxicity of orally
administered bufotalin. C R Seances Soc Biol Fil. 152:571–574.
1958.(In French). PubMed/NCBI
|
|
16
|
Pettit GR and Kamano Y: Bufadienolides.
21. Synthesis of cinobufagin from bufotalin. J Org Chem.
37:4040–4044. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang DM, Liu JS, Tang MK, Yiu A, Cao HH,
Jiang L, Chan JY, Tian HY, Fung KP and Ye WC: Bufotalin from
Venenum bufonis inhibits growth of multidrug resistant HepG2 cells
through G2/M cell cycle arrest and apoptosis. Eur J
Pharmacol. 692:19–28. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wen L, Liu L, Wen L, Yu T and Wei F:
Artesunate promotes G2/M cell cycle arrest in MCF7 breast cancer
cells through ATM activation. Breast Cancer. 25:681–686. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yin PH, Liu X, Qiu YY, Cai JF, Qin JM, Zhu
HR and Li Q: Antitumor activity and apoptosis-regulation mechanisms
of bufalin in various cancers: New hope for cancer patients. Asian
Pac J Cancer Prev. 13:5339–5343. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Malumbres M and Barbacid M: Cell cycle,
CDKs and cancer: A changing paradigm. Nat Rev Cancer. 9:153–166.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ren Y, Zhang SW, Xie ZH, Xu XM, Chen LL,
Lou ZG, Weng GB and Yao XP: Cantharidin induces G2/M
arrest and triggers apoptosis in renal cell carcinoma. Mol Med Rep.
14:5614–5618. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang CC, Hung CF and Chen BH: Preparation
of coffee oil-algae oil-based nanoemulsions and the study of their
inhibition effect on UVA-induced skin damage in mice and melanoma
cell growth. Int J Nanomedicine. 12:6559–6580. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang C, Liang C, Feng W, Xia X, Chen F,
Qiao E, Zhang X, Chen D, Ling Z and Yang H: ICT1 knockdown
inhibits breast cancer cell growth via induction of cell cycle
arrest and apoptosis. Int J Mol Med. 39:1037–1045. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang WW, Tsai SC, Peng SF, Lin MW, Chiang
JH, Chiu YJ, Fushiya S, Tseng MT and Yang JS: Kaempferol induces
autophagy through AMPK and AKT signaling molecules and causes
G2/M arrest via downregulation of CDK1/cyclin B in
SK-HEP-1 human hepatic cancer cells. Int J Oncol. 42:2069–2077.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li P, Ding N, Zhang W and Chen L: COPS2
antagonizes OCT4 to accelerate the G2/M transition of mouse
embryonic stem cells. Stem Cell Reports. 11:317–324. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Anger M, Stein P and Schultz RM: CDC6
requirement for spindle formation during maturation of mouse
oocytes. Biol Reprod. 72:188–194. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li S and Guo L: Pseudolaric acid B induces
G2/M arrest and inhibits invasion and migration in HepG2 hepatoma
cells. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi. 34:59–64. 2018.(In
Chinese). PubMed/NCBI
|
|
28
|
She H and Mao Z: Study of ATM
phosphorylation by Cdk5 in neuronal cells. Methods Mol Biol.
1599:363–374. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Blackford AN and Jackson SP: ATM, ATR, and
DNA-PK: The trinity at the heart of the DNA damage response. Mol
Cell. 66:801–817. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ashley AK and Kemp CJ: DNA-PK, ATM, and
ATR: PIKKing on p53. Cell Cycle. 17:275–276. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hennessy BT, Smith DL, Ram PT, Lu Y and
Mills GB: Exploiting the PI3K/AKT pathway for cancer drug
discovery. Nat Rev Drug Discov. 4:988–1004. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xu QG, Yu J, Guo XG, Hou GJ, Yuan SX, Yang
Y, Yang Y, Liu H, Pan ZY, Yang F, et al: IL-17A promotes the
invasion-metastasis cascade via the AKT pathway in hepatocellular
carcinoma. Mol Oncol. 12:936–952. 2018. View Article : Google Scholar : PubMed/NCBI
|