Introduction
Cerebral ischemic stroke (IS) is a common
cerebrovascular disease in clinic (1). It is a major cause of disability and
the third leading cause of death after cancer and myocardial
infarction worldwide (2,3). Generally, there are two clinical
manifestations of stroke, IS and hemorrhagic stroke, and IS
accounts for ~87% of all cases (4,5). As
the only clinical medication approved by the US Food and Drug
Administration, tPA must be administered within 4.5 h at the latest
and therefore only few IS patients benefit from this drug (6,7).
Researches have reported that angiogenesis and newborn neurons are
crucial for the repair of cerebral ischemia and therefore are
acknowledged as a promising therapeutic strategy for IS (8,9).
Therefore, searching for a drug that facilitates angiogenesis and
the survival and function of neurons is now the focus of basic
research for IS treatment.
Saffron, as known as Crocus sativus L., is a
plant belonging to Iridaceae family. It is widely cultivated in a
great number of countries in the world, such as Turkey, Greece,
India, Iran and China (10,11).
In ancient times, saffron was used as the ingredient of energizing
drink and orange jam (12).
Nowadays, more and more pharmacological effects of saffron have
been revealed, such as antioxidant, anti-tumor, anti-depressant,
anticonvulsant, memory and learning enhancer and blood pressure
regulator (13,14). More importantly, some experimental
studies have demonstrated that saffron by oral administration can
suppress neuronal death and lead to a decrease of infarct size in
stroke rats (15,16). Safranal, a monoterpene aldehyde, is
an active ingredient of saffron and responsible for the
characteristic odor and aroma of this plant (17). The properties of safranal have been
reported to include anti-inflammatory (18), anti-oxidant (19), antidiabetic (20), anti-carcinogenic (21) and anti-hypertensive (22) effects. Its neuroprotective role in
neurological disorders has also been widely described, in
Alzheimer's disease (23),
epilepsy (24), spinal cord injury
(25) and Parkinson's disease
(26). Additionally, the
protective role of safranal in the onset and development of IS has
also been reported. For example, both Sadeghnia et al
(27) and Hosseinzadeh et
al (28) constructed a middle
cerebral artery occlusion/reperfusion (MCAO/R) model in rats
followed by administration with safranal via intraperitoneal
injection. They concluded that the hippocampal cell loss and
infarct volume of IS rats are markedly improved following safranal
treatment and these improvements are due to the antioxidant
activity of safranal (27,28). Forouzanfar et al (29) established an oxygen-glucose
deprivation/reoxygenation (OGD/R) model in PC12 cells and indicated
that safranal dramatically attenuates oxidative damage and
suppresses apoptosis in OGD/R-induced PC cells. However, the
detailed mechanism and the effects of safranal on angiogenesis and
the repair of nerve system are remain to be elucidated.
The present study not only constructed a MCAO/R rat
as in vivo model but also used OGD/R-induced rat brain
microvascular endothelial cells (RBMEC) as in vitro model.
The aim of the current study was to preliminarily ascertain the
neurorestorative effects of safranal against IS and provide a
theoretical basis the clinical therapy of IS patients.
Materials and methods
Reagents and cells
RBMEC and DMEM-H media containing 10% FBS were
procured from BeNa Culture Collection. MilliporeSigma provided
safranal (purity ≥90%). Bovine serum albumin (BSA) was purchased
from Beijing Solarbio Science & Technology Co., Ltd. Terminal
deoxynucleotidyl transferase dUTP nick end labeling (TUNEL)
Apoptosis Detection Kit was obtained from AtaGenix and
2,3,5-triphenyltetrazolium chloride (TTC) solution (2%) was
procured from Shanghai Macklin Biochemical Co., Ltd. Nissl staining
solution, DAPI, Rat TGF-β1 enzyme-linked immunosorbent assay
(ELISA) kit (cat. no. PT878), BCA protein assay kit, ECL detection
kit, RIPA lysis buffer and Triton X-100 were from Beyotime
Institute of Biotechnology. Rat VEGF ELISA kit (cat. no. ED-30908),
Rat basic fibroblast growth factor 2 (bFGF-2) ELISA kit (cat. no.
LCSCM30506) and Rat platelet-derived growth factor (PDGF) ELISA kit
(cat. no. ED-34434) were obtained from Xiamen Lun Changshuo
Biological Technology Co., Ltd. For western blotting, primary
antibodies against silent information regulator 1 (SIRT1),
brain-derived neurotrophic factor (BDNF), synaptophysin (SYN),
microtubule associated protein 2 (MAP-2), GAPDH and HRP-conjugated
secondary antibodies were from Proteintech Group, Inc.; nerve
growth factor (NGF), neurotrophin-4 (NT-4) and Tau-1 were from
Abcam; postsynaptic density protein 95 (PSD95) was from Cell
Signaling Technology, Inc. For immunofluorescence staining, primary
antibodies ionized calcium-binding adapter molecule 1 (Iba-1; cat.
no. 66827-1-Ig) and tyrosine hydroxylase (TH; cat. no. 66334-1-Ig)
were obtained from Proteintech Group, while primary antibody
syntaxin (cat. no. ab188583) and fluorescein
isothiocyanate-conjugated secondary antibody (cat. no. ab6717) were
from Abcam. RNA extraction agent TRIzol® reagent was
obtained from Thermo Fisher Scientific, Inc. Hifair® II
1st Strand cDNA Synthesis SuperMix and Hieff® qPCR SYBR
Green Master Mix were purchased from Shanghai Yeasen Biotechnology
Co., Ltd. Small interfering (siRNA SIRT1-1/-2/-3 and the negative
control (siRNA NC) were synthesized by Weizhen Biosciences.
Transfection reagent Lipofectamine® 3000 was obtained
from Invitrogen (Thermo Fisher Scientific, Inc.). Dojindo Molecular
Technologies, Inc. provided CCK-8 solution used for the measurement
of cell viability. Matrigel obtained from BD Biosciences was used
for tube formation assay.
Establishment for MCAO/R rat model and
neurological evaluation
A total of 30 healthy male Sprague-Dawley rats
(280±20 g; 10 weeks old) were purchased from Vital River Laboratory
Animal Technology (Pinghu, China) and housed in specific
pathogen-free environments with a temperature of 22±2˚C and a
humidity of 50-60%, under a 12 h light-dark cycle, with food and
water available ad libitum. Following acclimatization in
laboratory, rats were randomly divided into five groups: The sham,
model, safranal (10 mg/kg), safranal (20 mg/kg) and safranal (40
mg/kg) groups, with six rats in each group. The MCAO/R rat model
was established as previously described (30). Briefly, rats in each group were
anesthetized using isoflurane (2% for induction and 1.5% for
maintenance, in 80% N2O and 20% O2). MCAO/R
was performed as follows: A midline incision was made to expose the
right internal carotid artery and then a 6-0 nylon monofilament
suture was inserted into the right internal carotid artery of each
rat for artery occlusion for 2 h. Subsequently, the filament was
withdrawn carefully to restore blood reperfusion. Rats in the sham
group underwent the same surgical procedure apart from MCAO.
Different doses of safranal (10, 20 and 40 mg/kg) were used to
treat the model rats respectively via intraperitoneal injection for
28 consecutive days. Then, Zea-Longa and Ludmila Belayev scores
were used to assess the neurological functions of rats, in a
double-blind manner (31). The
present study was conducted after obtaining the approval of
Laboratory Animal Ethics Committee of Wenzhou Seventh People's
Hospital (Wenzhou, China; approval no. 19JCYBJC00323) and the
experimental procedures in rats were executed in compliance with
the NIH Guide for the Care and Use of Laboratory Animals (8th
edition; https://www.ncbi.nlm.nih.gov/books/NBK54050/).
TTC staining
The rats were euthanized by exposure to 4%
isoflurane followed by cervical dislocation, and death was
confirmed by a lack of response to tail clamping. The brain tissues
were harvested and cut into 2-mm-thick slices. The slices were
stained with TTC solution (2%) at 37˚C for 15 min in the dark and
then images were captured. Red represented the normal brain
tissues, while pale color represented the ischemic brain tissues.
The infarct size was assessed using ImageJ software (version 1.8;
National Institutes of Health).
Nissl staining
Brain paraffin sections were routinely dewaxed at
room temperature with xylene I and xylene II, for 10 min each, and
then gradient alcohol dehydration (100, 100, 95, 90, 80, 70 and
50%, each 5 min) was performed. Subsequently, the sections were
stained with Nissl staining solution for 40 min at 50-60˚C.
Sections were washed with deionized water, differentiated with
Nissl differentiation solution, dehydrated with anhydrous ethanol,
cleared with xylene and sealed with neutral gum. The stained
hippocampal neurons were observed under a light microscope (Olympus
Corporation; magnification, x400) and five areas were randomly
selected from each group for statistical analysis.
Immunofluorescence staining
Brain slices were deparaffinized with xylene
(changed twice, 10 min each) and rehydrated in a graded ethanol
series. After rinsing in PBS three times, the sections were
underwent antigen retrieval in 10 mM Citrate buffer for 20 min at
95˚C. Following washing by PBS for three times, the slices were
then permeabilized with 0.5% TritonX-100 in PBS at room temperature
for 1 h. The sections were blocked with 5% BSA at room temperature
for 1 h and were then incubated with primary antibodies Iba-1
(1:200; a marker of microglia), syntaxin (1:200; a marker of
α-synuclein) and TH (1:200; a marker of dopaminergic neuron)
overnight at 4˚C and then with the corresponding secondary
antibodies labeled with fluorescent dyes (1:200) at 37˚C for 30
min. Following staining with DAPI at 37˚C for 15 min, images were
captured using a fluorescence microscope (Olympus Corporation;
magnification, x400).
TUNEL assay
In accordance with the experimental operations in
the manufacturer's specifications of the kit, the apoptosis was
evaluated in brain tissues and RBMEC. The images were captured
using a fluorescence microscope (Olympus Corporation;
magnification, x400).
ELISA
According to the instructions, the concentrations of
VEGF, TGF-β1, bFGF-2 and PDGF in the serum and cortex of rats were
determined via the corresponding commercial kits.
Cell culture, OGD/R model
establishment and safranal treatment
DMEM-H containing 10% FBS was used for the
cultivation of RBMEC. The cells were maintained in a 5%
CO2 and 95% air incubator at 37˚C. As previously
reported (30), an in vitro
OGD/R model was established with slight modification. In brief,
RBMEC were cultured in a glucose-free DMEM-H medium for 6 h at 37˚C
in a gas mix of 95% N2 and 5% CO2 for
simulation of cerebral ischemia. Then, RBMEC were transferred into
a normoxic incubator (95% air and 5% CO2) and incubated
for 24 h to mimic reperfusion. RBMEC in the control group were
cultured in normoxic environment and normal medium. In
safranal-treated group, RBMEC subjected to ODG/R were treated with
50 µg/ml safranal for 12 h as previously described (32).
Cell transfection
Using Lipofectamine® 3000 (Invitrogen;
Thermo Fisher Scientific, Inc.) RBMEC were co-transfected with
siRNA SIRT1-1 (5'-CAGTTTCATAGAGCCATGAAGTATG-3'), siRNA SIRT1-2
(5'-CCAGTAGCACTAATTCCAAGTTCTA-3'), siRNA SIRT1-3
(5'-GCTACACTTGTAGACCAAACAATAA-3') or siRNA NC
(5'-GCTTTCAGATGACCAACAAACATAA-3') for 48 h at 37˚C. The
concentration of all siRNA agents was 50 nM. After that, 48 h after
transfection, RBMEC were gathered and the gene expression levels of
SIRT1 were detected through reverse transcription-quantitative
(RT-q) PCR.
Total RNA isolation and RT-qPCR
According to the manufacturer's protocols, total RNA
extraction from brain cortex and RBMEC (2x105) was
implemented using TRIzol®, followed by synthesizing cDNA
products with the aid of Hifair II 1st Strand cDNA Synthesis
SuperMix. Following the instructions of manufacturer, RT-qPCR
analysis was carried out using the Hieff qPCR SYBR Green Master
Mix. The following thermocycling conditions were used for the qPCR:
Initial denaturation at 95˚C for 3 min, followed by 40 cycles at
95˚C for 15 sec, annealing at 60˚C for 30 sec, elongation at 72˚C
for 1 min and a final extension at 72˚C for 5 min. GAPDH was used
for normalization. The mRNA expression levels were calculated by
applying the 2-ΔΔCq approach (33). The primers used are listed in
Table I.
 | Table IReal-time PCR primer synthesis
list. |
Table I
Real-time PCR primer synthesis
list.
| Gene | Sequences |
|---|
| SIRT1 | |
|
Forward |
5'-ATCTCCCAGATCCTCAAGCCA-3' |
|
Reverse |
5'-CTTCCACTGCACAGGCACAT-3' |
| BDNF | |
|
Forward |
5'-AATAATGTCTGACCCCAGTGCC-3' |
|
Reverse |
5'-ATTGTTGTCACGCTCCTGGT-3' |
| NGF | |
|
Forward |
5'-GAGCGCATCGCTCTCCTT-3' |
|
Reverse |
5'-GTGTGAGTCGTGGTGCAGTA-3' |
| NT-4 | |
|
Forward |
5'-AGGACCCTGACTTACCCTGG-3' |
|
Reverse |
5'-CCTAGCCCCAGCTCATACAT-3' |
| SYN | |
|
Forward |
5'-TACAGCCGTGTTCGCTTTCA-3' |
|
Reverse |
5'-GTGGCCATCTTCACATCGGA-3' |
| PSD95 | |
|
Forward |
5'-CCGCTACCAAGATGAAGACAC-3' |
|
Reverse |
5'-GTTCCATTCACCTGCAACTCA-3' |
| MAP-2 | |
|
Forward |
5'-CTGCACTGGAAGAAGCCTCG-3' |
|
Reverse |
5'-GAGGAACTAAGGCAGCGTGT-3' |
| Tau-1 | |
|
Forward |
5'-TCCTCGCCTCCTGTCGATTA-3' |
|
Reverse |
5'-AGCTTGGTCCTCCATGTTCG-3' |
| GAPDH | |
|
Forward |
5'-TCAAGAAGGTGGTGAAGCAGG-3' |
|
Reverse |
5'-TCAAAGGTGGAGGAGTGGGT-3' |
Western blotting
Cortex tissues or RBMEC were lysed with RIPA lysis
buffer on ice for 15 min. A BCA protein assay kit was used for
examining total protein content. After separating the protein
products (50 µg protein/lane) on 10% gels using SDS polyacrylamide
gel electrophoresis, they were transferred onto PVDF membranes,
blocked with 5% nonfat milk for 2 h at 25˚C. Subsequently, primary
antibodies including SIRT1 (1:20,000), BDNF (1:2,000), SYN
(1:20,000), MAP-2 (1:20,000), NGF (1:3,000), NT-4 (1:3,000) and
Tau-1 (1:1,000), PSD95 (1:1,000) and GAPDH (1:50,000) were added to
the membranes for incubation overnight at 4˚C. Then, the membrane
was placed in HRP-conjugated secondary antibodies (1:1,000), along
with incubation for 1 h at room temperature. The protein bands were
developed with an ECL detection Kit and analyzed with fully
automatic chemiluminescence imaging system (Tanon 5200Multi; Tanon
Science & Technology).
CCK-8 assay
RBMEC (5,000 cells/well) was plated onto a 96-well
plate. CCK-8 solution (10 µl) was mixed with the medium. Following
2 h of incubation, RBMEC were exposed to 10 µl stop buffer. Cell
viability was evaluated by examining the absorbance at 450 nm.
Wound healing assay
RBMEC were seeded in 6-well plates with a density of
5x104 cells/well. When the cells had grown to 100%
confluence, a scratch was conducted in the monolayer cells using a
pipette tip (10 µl) and RBMEC were then incubated in a serum-free
DMEM-H medium for 48 h. The width of scratches at 0 h or 48 h was
recorded under a microscope. The migrative potentials of RBMEC were
analyzed using an ImageTool software (UTHSCSA) with the following
formula: (the width at 0 h-the width at 48 h)/the width at 0 h
x100.
Tube formation experiments
RBMEC (5,000 cells/well) were plated into a 24-well
plate that pre-coated with Matrigel at 37˚C for 30 min, followed by
incubation in DMEM-H supplemented with 10% FBS at 37˚C for 24 h.
Tube formation was observed under a microscope. Tube length and
branch were analyzed using an ImageJ software (version 1.8;
National Institutes of Health).
Statistical analysis
In vitro experiments were performed in
triplicate, and each experiment was repeated three times. In
vivo experiments were performed using 6 rats per group. The
variations among the data were evaluated by one-way ANOVA, followed
by Tukey's multiple comparison test. Data analysis was performed
with SPSS software v22.0 (IBM Corp.). Measurement data were
displayed as the mean ± standard deviation. P<0.05 was
considered to indicate a statistically significant difference.
Results
Safranal ameliorates neuronal injury
in a MCAO/R rat model
Zea-Longa 5-point method and Ludmila Belayev
12-point method were used to assess the neurological functions of
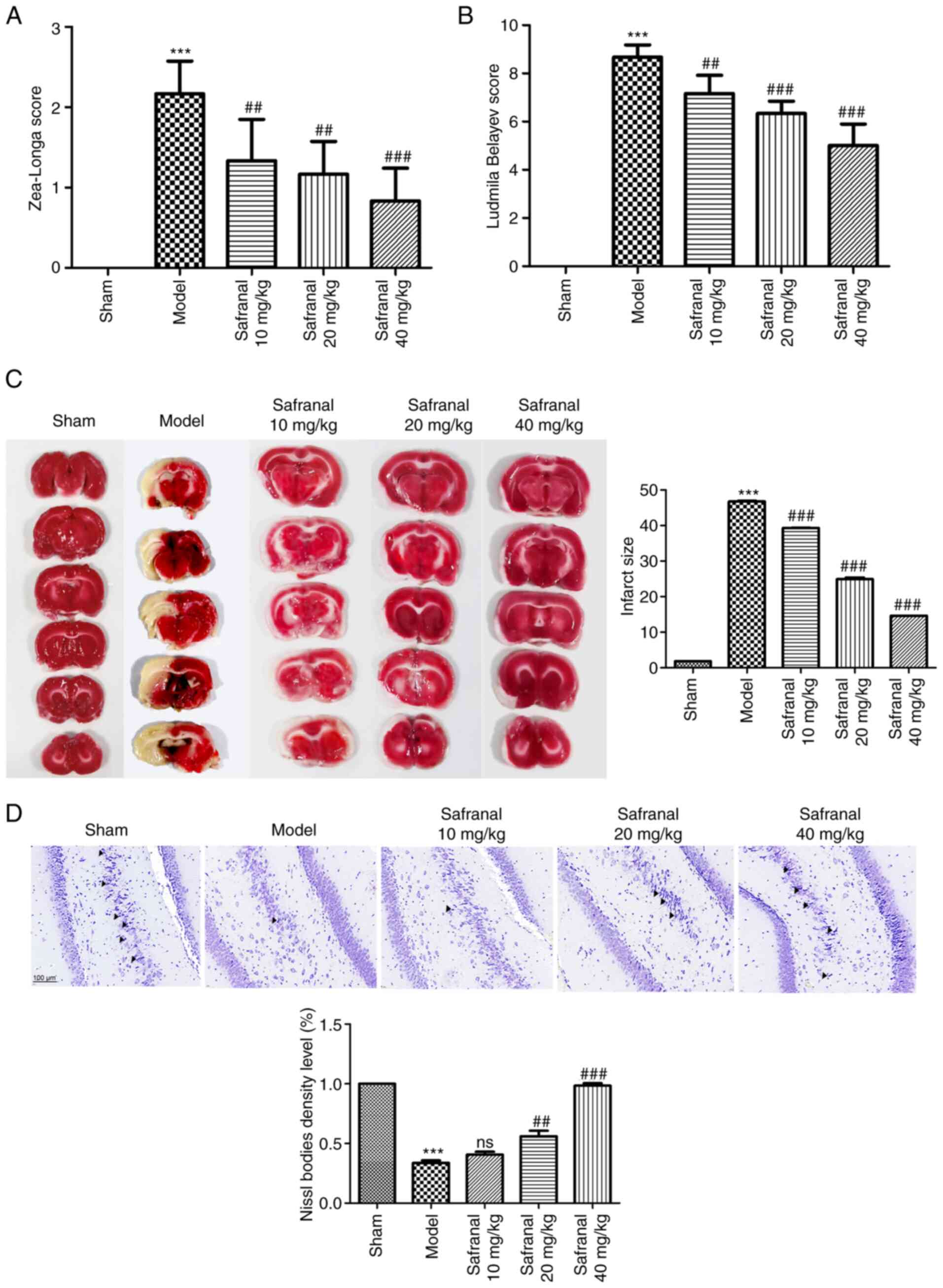
MCAO/R models. As shown in Fig. 1A
and B, compared with the sham
group, the Zea-Longa score and Ludmila Belayev score in the model
group were markedly increased (P<0.001). However, the
neurological functions of rats in the MCAO/R models were notably
improved following safranal treatment (P<0.01). The cerebral
infarct size was then evaluated via TTC staining. A larger infarct
size was observed in the model group as compared with the sham
group (Fig. 1C; P<0.001).
Safranal treatment could reduce infarct size remarkably, especially
for 40 mg/kg of safranal (P<0.001). As illustrated in Fig. 1D, Nissl staining showed that the
neurons in the sham group were arranged neatly and densely, with
abundant Nissl bodies. The arrangement of neurons in the model
group was loose, with a large number of vacuolar structures and a
significant decrease of Nissl bodies. In contrast to the model
group, the loss of Nissl bodies was significantly improved and the
activity of neuron was enhanced in safranal treatment group.
Overall, high dose of safranal (40 mg/kg) seemed to decrease the
Zea-Longa score and Ludmila Belayev score, reduce infarct size and
improve the loss of Nissl bodies more effectively compared with
lower doses. These results implied that high dose of safranal (40
mg/kg) was more helpful for ameliorating neuronal injury in a
MCAO/R rat model.
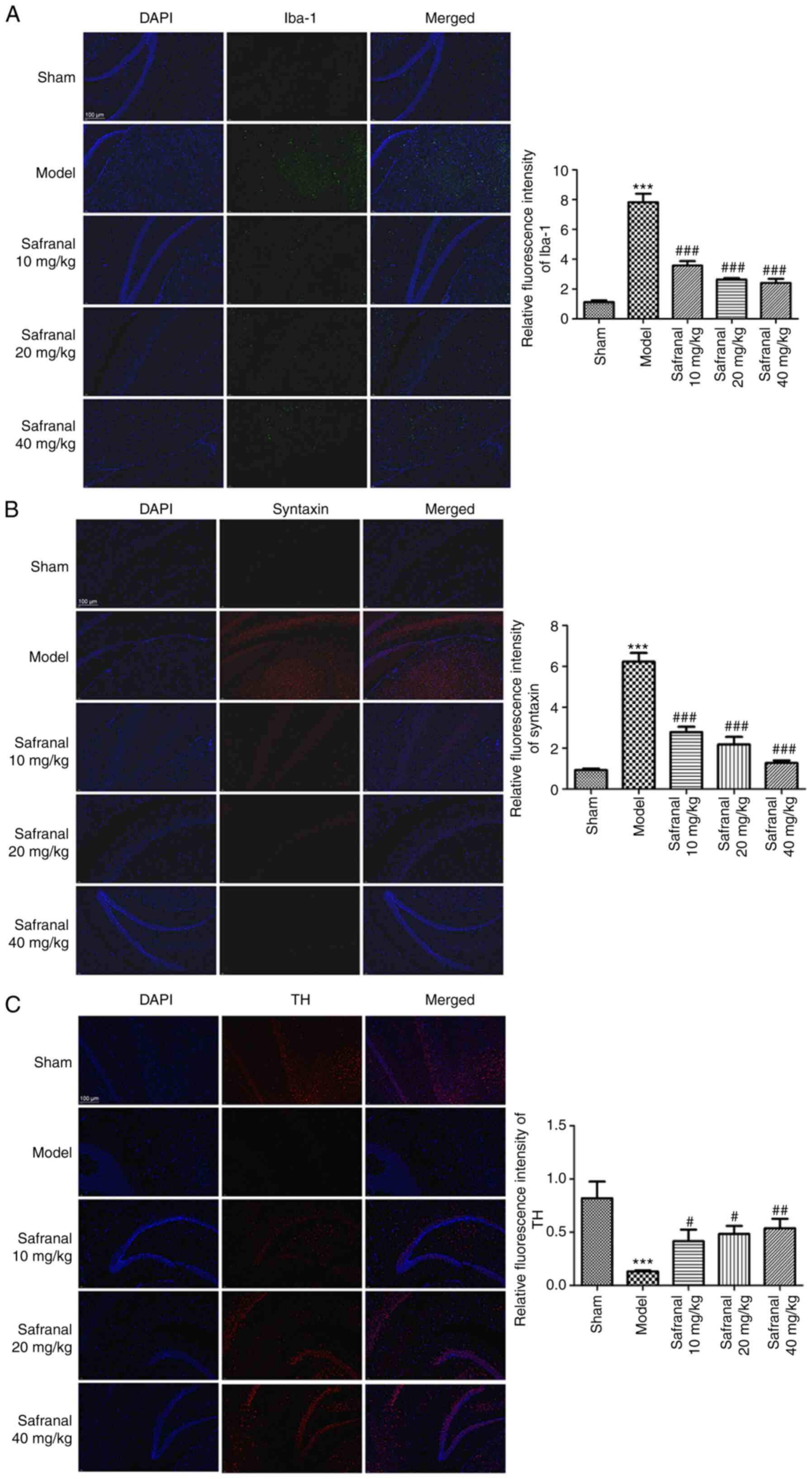
Safranal represses microglia
over-activation, synuclein aggregation and dopaminergic
neurodegeneration
Microglia over-activation, synuclein aggregation and
dopaminergic neuron loss are considered important pathological
features in neuronal injury-related diseases including, IS
(34,35). As shown in Fig. 2A and B, the results of immunofluorescence
demonstrated that the relative fluorescence intensity of Iba-1 and
synuclein in the model group was markedly increased compared with
that of the sham group (P<0.001). However, in different safranal
treatment groups, the relative fluorescence intensity of Iba-1 and
synuclein was significantly weaker (P<0.001). The opposite
results were observed in the relative fluorescence intensity of TH
(Fig. 2C; P<0.05).
Additionally, it was found that 40 mg/kg of safranal seemed to have
relatively stronger suppressive effect on the levels of Iba-1 and
synuclein, and relatively stronger alleviative effect on TH level.
These data suggested that high dose of safranal (40 mg/kg) can
significantly repress microglia over-activation, synuclein
aggregation and dopaminergic neurodegeneration.
Safranal suppresses neuron apoptosis
and promotes angiogenesis in a MCAO/R rat model
As the survival of neurons in brain tissues is
neuroprotective in IS, the effect of safranal on neuron apoptosis
was determined via TUNEL assay. TUNEL-positive cells in the model
group were significantly increased, whereas were clearly reduced
following safranal treatment (Fig.
3A; P<0.001). Angiogenic factors play an important role in
neovascularization after nerve injury. Safranal treatment,
especially for 40 mg/kg of safranal, markedly increased the levels
of VEGF, TGF-β1, bFGF-2 and PDGF in the serum and cortex of MCAO/R
rats (Fig. 3B and C; P<0.05). The above results indicated
that high dose of safranal (40 mg/kg) can suppress neuron apoptosis
and promotes angiogenesis effectively in a MCAO/R rat model.
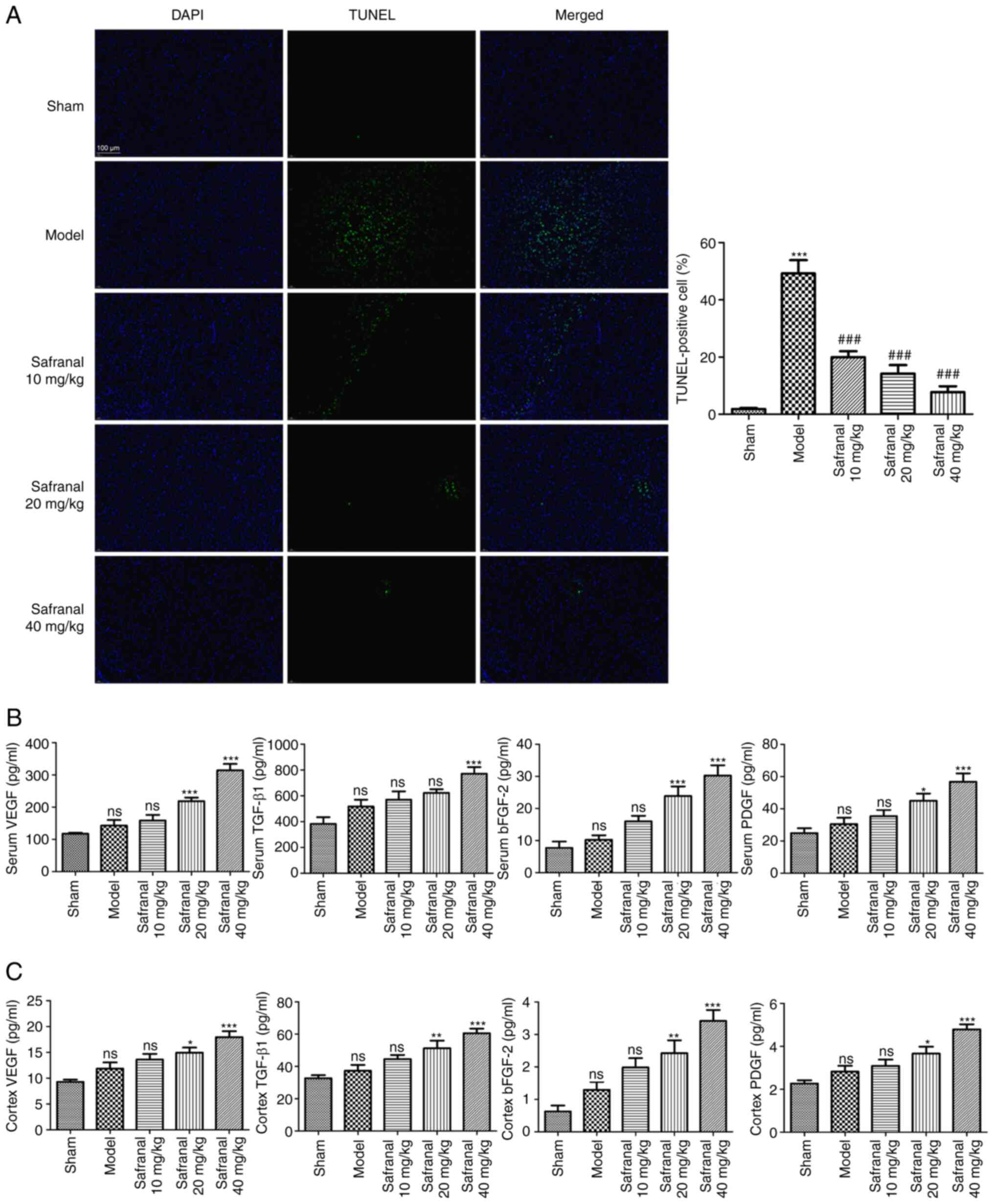 | Figure 3Safranal suppresses neuron apoptosis
and promotes angiogenesis in a MCAO/R rat model. (A) TUNEL-positive
cells in different groups were determined by TUNEL assay;
magnification x400. ***P<0.001 vs. sham.
###P<0.001 vs. model. (B) The concentrations of VEGF,
TGF-β1, bFGF-2 and PDGF in the serum of rats were measured by
ELISA. (C) The concentrations of VEGF, TGF-β1, bFGF-2 and PDGF in
the cortex of rats were measured by ELISA. *P<0.05,
**P<0.01, ***P<0.001 vs. model; ns, no
significance. In vivo experiments were performed using six
rats per group. MCAO/R, middle cerebral artery
occlusion/reperfusion; TUNEL, terminal deoxynucleotidyl transferase
dUTP nick end labeling; bFGF-2, basic fibroblast growth factor 2;
PDGF, platelet-derived growth factor; ELISA, enzyme-linked
immunosorbent assay. |
Safranal repairs nervous system
through upregulating SIRT1 expression
Activation of SIRT1 has been confirmed to exert
neuroprotection against cerebral IS (36). Neurotrophins such as BDNF, NGF and
NT-4 have been demonstrated to promote neurogenesis after cerebral
ischemia (37). Additionally, a
previous study demonstrated that dendritic remodeling and
synaptogenesis are important for neural restoration (38). The present study therefore explored
the effects of safranal treatment on the restoration of neural
function. As shown in Fig. 4A, the
mRNA expression of SIRT1, BDNF, NGF, NT-4, SYN, PSD95, MAP-2 (a
somato-dendritic marker) and Tau-1 (an axonal marker) was decreased
in the model group in contrast to the sham group (P<0.001). High
dose of safranal (40 mg/kg) significantly increased the mRNA
expression of SIRT1, BDNF, NGF, NT-4, SYN, PSD95, MAP-2 and Tau-1
(P<0.001). Similarly, the protein levels of these factors were
also markedly elevated following safranal treatment (Fig. 4B; P<0.001). These results
implied that high dose of safranal (40 mg/kg) can repair nervous
system by upregulating SIRT1 expression.
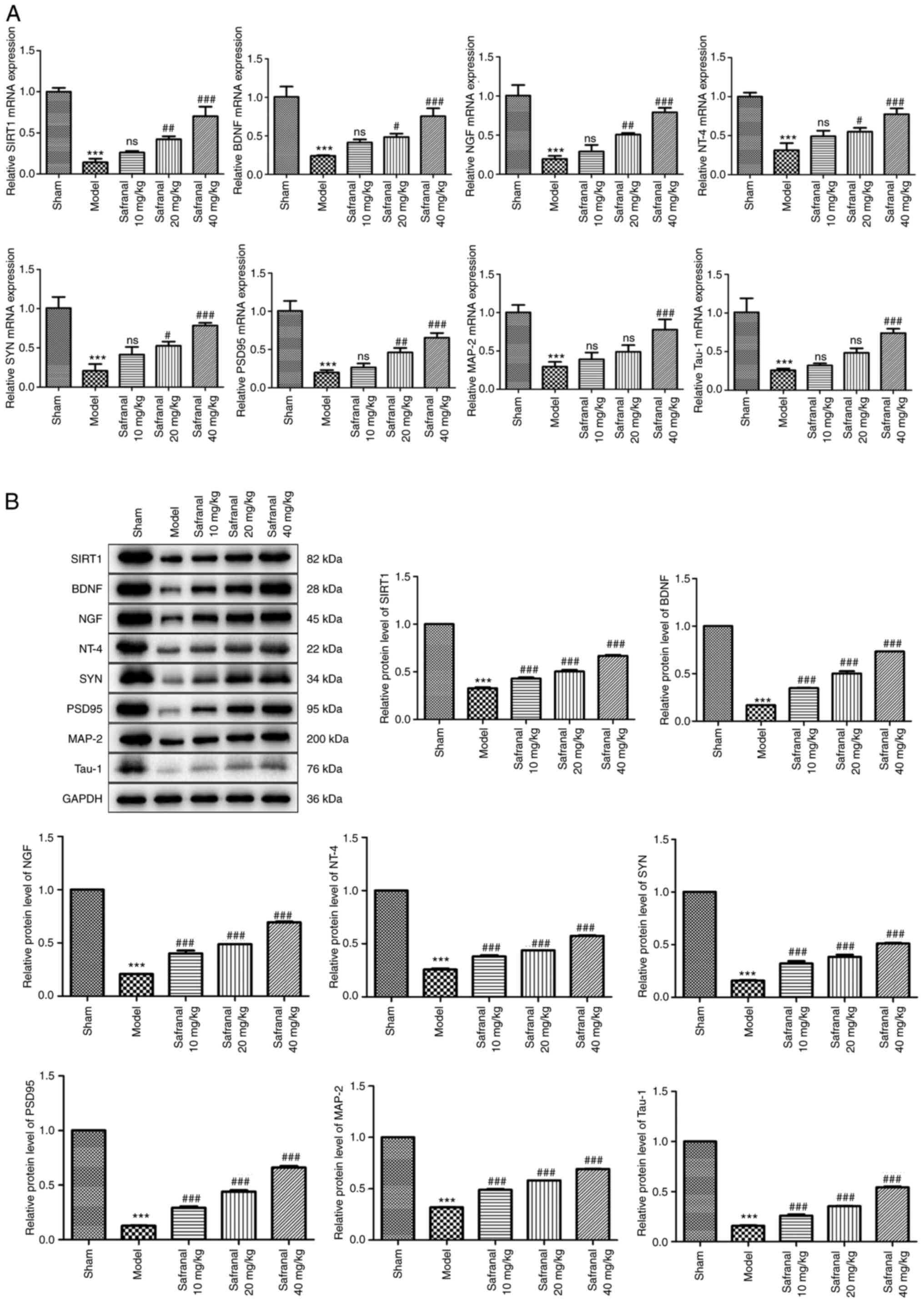 | Figure 4Safranal repairs nervous system
through upregulating SIRT1 expression. (A) The mRNA expression of
SIRT1, BDNF, NGF, NT-4, SYN, PSD95, MAP-2 and Tau-1 in different
groups was measured by reverse transcription-quantitative PCR. (B)
The protein levels of SIRT1, BDNF, NGF, NT-4, SYN, PSD95, MAP-2 and
Tau-1 in different groups were determined by western blotting.
***P<0.001 vs. sham. #P<0.05,
##P<0.01, ###P<0.001 vs. model; ns, no
significance. In vivo experiments were performed using six
rats per group. SIRT1, silent information regulator 1; BDNF,
brain-derived neurotrophic factor; NGF, nerve growth factor; NT-4,
neurotrophin-4; SYN, synaptophysin; PSD95, postsynaptic density
protein 95; MAP-2, microtubule associated protein 2; NGF, nerve
growth factor; NT-4, neurotrophin-4. |
Silencing of SIRT1 reverses the
effects of safranal on cell growth, migration, apoptosis and tube
formation in OGD/R-induced RBMEC
The interaction of safranal with SIRT1 was
ascertained in OGD/R-induced RBMEC. First, siRNA SIRT1-1/-2/-3 or
siRNA NC was introduced into RBMEC. As shown in Fig. 5A, SIRT1 had the lowest expression
level in RBMEC when transfected with siRNA SIRT1-1 (P<0.001).
Therefore, siRNA SIRT1-1 was selected for the succeeding tests.
CCK-8 and wound healing assays demonstrated that the viability and
migrative potential of RBMEC could be suppressed by OGD/R challenge
(Fig. 5B-D; P<0.001), but were
markedly enhanced following safranal treatment (P<0.05).
Notably, transfection of siRNA SIRT1 notably reversed the promoting
effects of safranal on the viability and migration of OGD/R-induced
RBMEC (P<0.05). For the effect of safranal interaction with
SIRT1 on the apoptosis of OGD/R-induced RBMEC, a large number of
TUNEL-positive cells were found in the OGD/R group as compared with
the control group (Fig. 5E;
P<0.001). In the OGD/R + safranal + siRNA NC group, the number
of TUNEL-positive cells was significantly reduced (P<0.01).
Moreover, knocking down SIRT1 attenuated the repressive effect of
safranal on the number of TUNEL-positive cells (P<0.01). The
angiogenesis of RBMEC was detected by tube formation assay. It was
demonstrated that safranal treatment promoted the tube formation of
RBMEC, with visual representation of increased total tube length
and branches (Fig. 5F; P<0.01)
and at the same time, siRNA SIRT1 transfection in OGD/R-induced
RBMEC in presence of safranal suppressed total tube and branch
length (P<0.01).
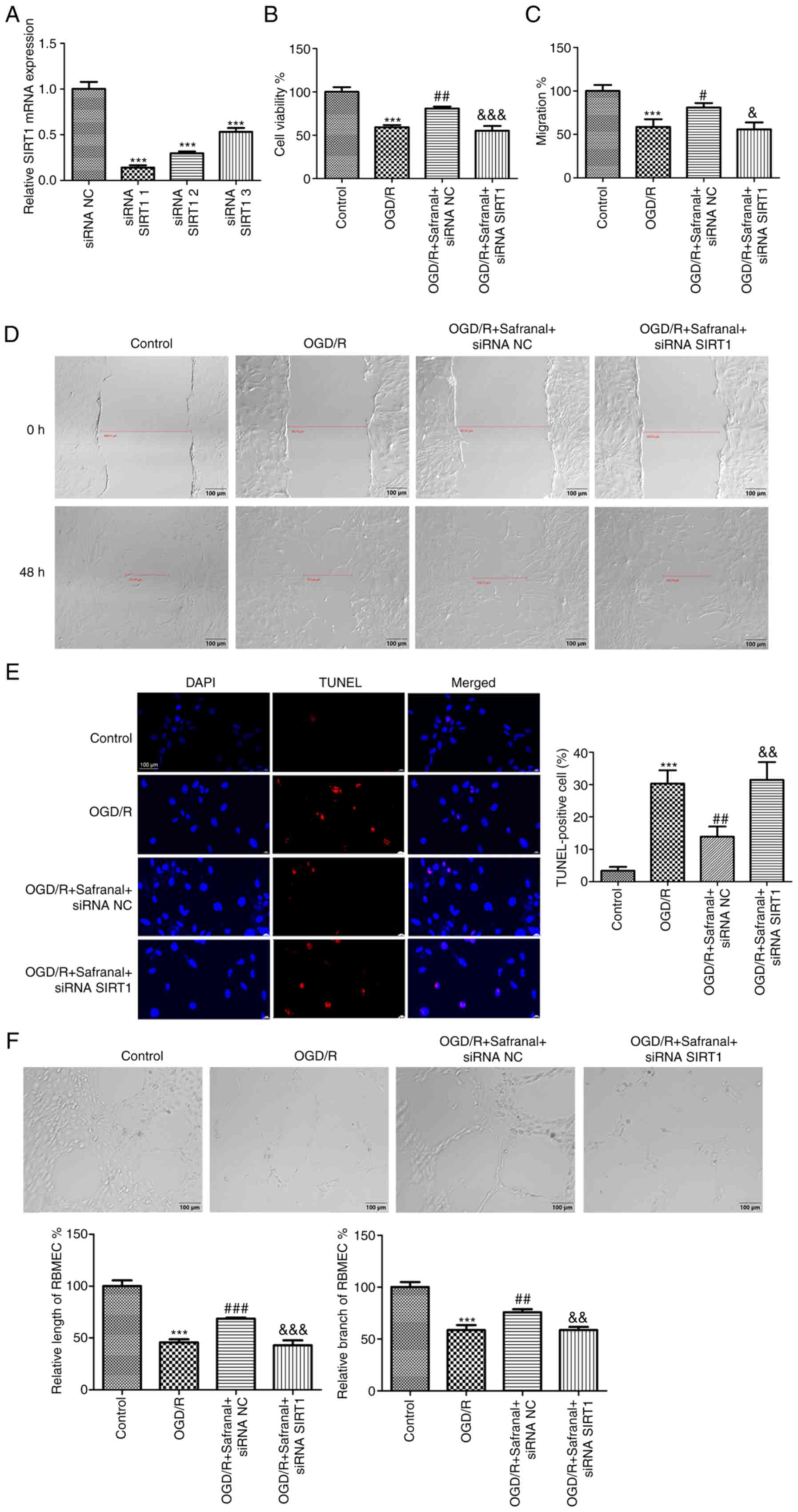 | Figure 5Silencing of SIRT1 reverses the
effects of safranal on cell growth, migration, apoptosis and tube
formation in OGD/R-induced RBMEC. (A) The mRNA expression of SIRT1
in RBMEC transfected with siRNA NC or siRNA SIRT1-1/-2/-3 was
measured by reverse transcription-quantitative PCR.
***P<0.001 vs. siRNA NC. (B) The viability of
OGD/R-induced RBMEC in different groups was calculated via CCK-8
assay. (C and D) The migrative potentials of OGD/R-induced RBMEC
were assessed through wound healing assay; scale bar, 100 µm. (E)
TUNEL-positive cells in different groups were determined by TUNEL
assay; magnification x 400. (F) Tube formation assay in
OGD/R-induced RBMEC; scale bar, 100 µm. ***P<0.001
vs. control. #P<0.05, ##P<0.01,
###P<0.001 vs. OGD/R. &P<0.05,
&&P<0.01,
&&&P<0.001 vs. OGD/R + safranal + siRNA
NC. In vitro experiments were performed in triplicate, and
each experiment was repeated three times. SIRT1, silent information
regulator 1; OGD/R, oxygen-glucose deprivation/reoxygenation;
RBMEC, rat brain microvascular endothelial cells; si, short
interfering; NC, negative control; TUNEL, terminal deoxynucleotidyl
transferase dUTP nick end labeling. |
Effects of interaction of safranal
with SIRT1 on the levels of neurotrophins and
synaptogenesis-related factors
As the effects of safranal on the restoration of
neural function had been ascertained in MCAO/R rat model as
described earlier, the effects of interaction of safranal with
SIRT1 on the levels of neurotrophins and synaptogenesis-related
factors in OGD/R-induced RBMEC needed to be further validated. As
shown in Fig. 6A and B, the mRNA expression and protein levels
of SIRT1, BDNF, NGF, NT-4, SYN, PSD95, MAP-2 and Tau-1 in the OGD/R
group were markedly decreased relative to those in the control
group (P<0.001). As expected, the mRNA expression and protein
levels of these factors were restored to some extent in the OGD/R +
safranal + siRNA NC group (P<0.05), but were further reduced in
OGD/R + safranal + siRNA SIRT1 group (P<0.05).
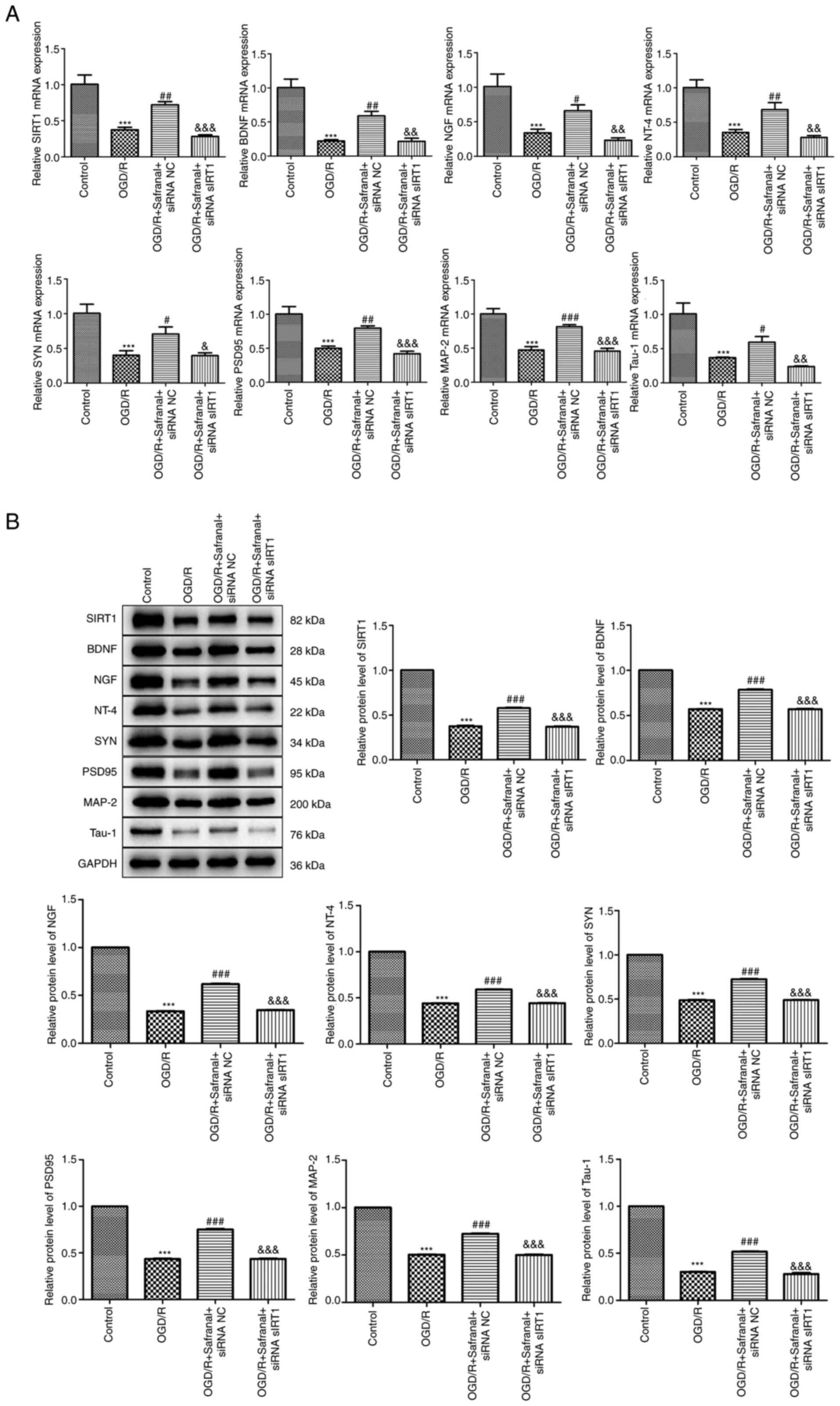 | Figure 6Effects of interaction of safranal
with SIRT1 on the levels of neurotrophins and
synaptogenesis-related factors. (A) The mRNA expression of SIRT1,
BDNF, NGF, NT-4, SYN, PSD95, MAP-2 and Tau-1 in OGD/R-induced RBMEC
was measured by RT-qPCR. (B) The protein levels of SIRT1, BDNF,
NGF, NT-4, SYN, PSD95, MAP-2 and Tau-1 in OGD/R-induced RBMEC were
determined by western blotting. ***P<0.001 vs.
control. #P<0.05, ##P<0.01,
###P<0.001 vs. OGD/R. &P<0.05,
&&P<0.01,
&&&P<0.001 vs. OGD/R + safranal + siRNA
NC. In vitro experiments were performed in triplicate, and
each experiment was repeated three times. SIRT1, silent information
regulator 1; BDNF, brain-derived neurotrophic factor; NGF, nerve
growth factor; NT-4, neurotrophin-4; SYN, synaptophysin; PSD95,
postsynaptic density protein 95; MAP-2, microtubule associated
protein 2 OGD/R, oxygen-glucose deprivation/reoxygenation; RBMEC,
rat brain microvascular endothelial cells; si, short interfering;
NC, negative control. |
Discussion
IS is a serious medical problem globally and
previous researches have reported that there are 15 million
patients suffered from IS every year, with ~6.5 million mortalities
and ~1/3 of patients suffered permanent disability due to neuronal
damage (39-41).
Therefore, IS has become an enormous burden to the healthcare
system worldwide. Although tPA is currently the best available
option for IS patients, the short therapeutic window and some
adverse effects such as intracranial hemorrhage have limited its
clinical application (42). In the
ischemic penumbra, neuron death and low cerebrovascular density are
the main features of IS patients (43,44).
However, little success in the therapeutics that promote neural
repair due to the complex mechanisms of IS process (45). The current study identified the
neurorestorative role and mechanism of safranal in IS, indicating
that safranal can promoting neuron survival, angiogenesis and
neurogenesis to attenuate IS progression via regulating SIRT1
expression.
MCAO/R is the most frequently used method to mimic
IS in murine models. In the present study, based on the
experimental results that rats in the model group exhibited
significantly higher neurological scores, Nissl body loss and
increased infarct size, a MCAO/R rat model was successfully
established. Additionally, it was found that high dose of safranal
(40 mg/kg) can significantly attenuate these symptoms, which was in
line with the data from Sadeghnia et al (27) indicating that safranal has
protective effects on MCAO/R rats. A previous study indicated that
angiogenesis is conducive to the recovery of blood supply in
ischemic penumbra, thus promoting the activation of neurons
(46). Angiogenesis is a process
that enhances the proliferative and migrative potentials of
vascular endothelial cells and the reconstruction of new vascular
networks (47). It is a complex
process regulated by pro-angiogenesis and anti-angiogenesis
cytokines. VEGF is considered as the most indispensable cytokine to
promote angiogenesis (43). TGF-β1
and bFGF-2 are both pro-angiogenesis and are involved in the
migration and proliferation of endothelial cells (46). PDGF combined with its receptor are
responsible for recruiting brain pericytes into newly formed
vessels during the progression of angiogenesis (48). Therefore, it was hypothesized that
safranal may exert a neuroprotective role by inhibiting endothelial
cell apoptosis and promoting angiogenesis. As expected, it was
found that safranal treatment, especially at 40 mg/kg, markedly
elevated the levels of VEGF, TGF-β1, bFGF-2 and PDGF in both serum
and cortex. In addition, it also suppressed the percentage of
TUNEL-positive cell to a great extent. These results corroborated
the hypothesis.
Furthermore, the over-activation of microglia,
α-synuclein aggregation and dopaminergic neurodegeneration are
considered as the important pathological features in neuronal
injury-related diseases including IS (34,35).
A previous study revealed that safranal can drive the inhibition
and disaggregation of synuclein fibrils (49). It was considered that safranal
treatment may suppress microglia over-activation, synuclein
aggregation and dopaminergic neuron loss in MCAO/R rats. The
results that high dose of safranal (40 mg/kg) distinctly attenuated
the fluorescence intensities of Iba-1 and syntaxin and enhanced TH
fluorescence intensity validated this hypothesis. Dendrites and
axons are two main structures in mature neurons. It is reported
that dendritic remodeling and synaptogenesis can promote the
neurological activity in the cerebral cortex (38). Moreover, neurons can release
neurotransmitters to regulate the populations of neurons (50). In the results of the present study,
a high dose of safranal (40 mg/kg) not only significantly increased
the levels of SYN and PSD95, but also elevated the levels of axon
and dendrite markers Tau-1 and MAP-2. These results implied that
safranal may be beneficial for the differentiation of neuron. In
addition, neurotrophins such as BDNF, NGF and NT-4 have been
reported to be related to neurogenesis (37). BDNF has been shown to reduce neuron
death and promote neuroplasticity and neurite growth (41). NGF can stimulate the formation of
axonal sprouting and induce axonal functional reconnection
(51). NT-4 is involved in the
differentiation and regeneration of neurons (52). The present study therefore further
ascertained the effects of safranal on these neurotrophins. As
expected, safranal at 40 mg/kg also clearly increased the levels of
BDNF, NGF and NT-4. All the above results confirmed the
neurorehabilitation role of safranal in IS development.
SIRT1 is a type of histone deacetylase belong to
sirtuin family (53). An
anatomical study conducted by Zakhary et al (54) indicated that SIRT1 is mainly
distributed in basal ganglia, prefrontal cortex and hippocampus of
the rodent and human nervous systems. A large number of researches
has demonstrated that SIRT1 can be activated by some compounds to
alleviate IS through several mechanisms, namely antioxidative,
antiapoptotic and anti-inflammatory effects. For example, α-lipoic
acid can alleviate the damage to the ischemic brain by reducing
oxidative damage through activation of SIRT1(55). Salvianolic acid B treatment
increases the expression of SIRT1 and then leads to a decrease of
Bax and an increase of Bcl-2, finally attenuating IS development
via inhibiting neuron apoptosis (56). Another study indicated that
resveratrol acts as an activator of SIRT1 to exert neuroprotective
effect in MCAO/R rats (57). Based
on the protective role of safranal in MCAO/R rats obtained in the
present study, it was hypothesized that safranal may promote
angiogenesis and neurogenesis by upregulating SIRT1 expression.
First, the present study found that safranal upregulated the
expression of SIRT1 both in vivo and in vitro. This
finding implied that SIRT1 was indeed a molecular target of
safranal. The interaction between safranal and SIRT1 was then
explored in OGD/R-induced RBMEC. The present study demonstrated
that silencing of SIRT1 reversed the promoting effects of safranal
on the viability and migration of OGD/R-induced RBMEC and the
inhibiting effect on apoptosis. Knocking down SIRT1 also suppressed
RBMEC tube formation caused by safranal. These results confirmed
the antiapoptotic and angiogenesis roles of safranal in IS
progression. Additionally, in OGD/R-induced RBMEC, it was further
indicated that silencing of SIRT1 also reversed the enabling
effects of safranal on the levels of BDNF, NGF, NT-4, SYN, PSD95,
MAP-2 and Tau-1. It was considered that safranal can induce
neurogenesis through upregulating SIRT1 expression.
Briefly, the present study mainly analyzed the
neurorestorative role of safranal in MCAO/R rats and OGD/R-induced
RBMEC and indicated that safranal can promote neuron survival,
angiogenesis and neurogenesis to attenuate IS progression by
upregulating SIRT1 expression. This may contribute more information
and insights on the underlying action mechanism of safranal for IS
treatment in clinical practice.
Acknowledgements
Not applicable.
Funding
Funding: The present study was supported by The Medical and
Health Technology Plan Project of Zhejiang Province (grant no.
2023KY1163) and The Basic scientific research projects of Wenzhou
City (grant no. Y20220230).
Availability of data and materials
The datasets analyzed during the current study are
available from the corresponding author on reasonable request.
Authors' contributions
JY and GH made substantial contributions to the
conception and design of the work. FH and CC made substantial
contributions to the acquisition, analysis and interpretation of
data for the work, and drafted the manuscript. YW, SW, SL and JJ
revised the manuscript critically for important intellectual
content. FH and CC confirm the authenticity of all the raw data.
All authors agreed to be accountable for all aspects of the work in
ensuring that questions related to the accuracy, and all authors
read and approved the final manuscript.
Ethics approval and consent to
participate
Experiments for animals were conducted in compliance
with the NIH Guide for the Care and Use of Laboratory Animals, and
approved by the ethical committee of Wenzhou Seventh People's
Hospital (Wenzhou, China; approval no. 19JCYBJC00323).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chen Z, Jiang B, Ru X, Sun H, Sun D, Liu
X, Li Y, Li D, Guo X and Wang W: Mortality of stroke and its
subtypes in China: Results from a nationwide population-based
survey. Neuroepidemiology. 48:95–102. 2017.PubMed/NCBI View Article : Google Scholar
|
|
2
|
Žitňanová I, Šiarnik P, Kollár B, Chomová
M, Pazderová P, Andrezálová L, Ježovičová M, Koňariková K,
Laubertová L, Krivošíková Z, et al: Oxidative stress markers and
their dynamic changes in patients after acute ischemic stroke. Oxid
Med Cell Longev. 2016(9761697)2016.PubMed/NCBI View Article : Google Scholar
|
|
3
|
Katan M and Luft A: Global burden of
stroke. Semin Neurol. 38:208–211. 2018.PubMed/NCBI View Article : Google Scholar
|
|
4
|
Donkor ES: Stroke in the 21st century: A
snapshot of the burden, epidemiology, and quality of life. Stroke
Res Treat. 2018(3238165)2018.PubMed/NCBI View Article : Google Scholar
|
|
5
|
Boehme AK, Esenwa C and Elkind MS: Stroke
Risk factors, genetics, and prevention. Circ Res. 120:472–495.
2017.PubMed/NCBI View Article : Google Scholar
|
|
6
|
Ma H, Campbell BCV, Parsons MW, Churilov
L, Levi CR, Hsu C, Kleinig TJ, Wijeratne T, Curtze S, Dewey HM, et
al: Thrombolysis guided by perfusion imaging up to 9 h after onset
of stroke. N Engl J Med. 380:1795–1803. 2019.PubMed/NCBI View Article : Google Scholar
|
|
7
|
Campbell BCV, Ma H, Ringleb PA, Parsons
MW, Churilov L, Bendszus M, Levi CR, Hsu C, Kleinig TJ, Fatar M, et
al: Extending thrombolysis to 4·5-9 h and wake-up stroke using
perfusion imaging: A systematic review and meta-analysis of
individual patient data. Lancet. 394:139–147. 2019.PubMed/NCBI View Article : Google Scholar
|
|
8
|
Li Y, Zhang X, Cui L, Chen R, Zhang Y,
Zhang C, Zhu X, He T, Shen Z, Dong L, et al: Salvianolic acids
enhance cerebral angiogenesis and neurological recovery by
activating JAK2/STAT3 signaling pathway after ischemic stroke in
mice. J Neurochem. 143:87–99. 2017.PubMed/NCBI View Article : Google Scholar
|
|
9
|
Oshikawa M, Okada K, Kaneko N, Sawamoto K
and Ajioka I: Affinity-immobilization of VEGF on laminin porous
sponge enhances angiogenesis in the ischemic brain. Adv Healthc
Mater. 6:2017.PubMed/NCBI View Article : Google Scholar
|
|
10
|
Javadi B, Sahebkar A and Emami SA: A
survey on saffron in major islamic traditional medicine books. Iran
J Basic Med Sci. 16:1–11. 2013.PubMed/NCBI
|
|
11
|
Khazdair MR, Boskabady MH, Hosseini M,
Rezaee R and M Tsatsakis A: The effects of Crocus sativus
(saffron) and its constituents on nervous system: A review.
Avicenna J Phytomed. 5:376–391. 2015.PubMed/NCBI
|
|
12
|
Nasrallah N: Ingredients used in foods and
medicinal preparations: Herbs, spices, aromatics, minerals, food
colors, and seasoning sauces. Treasure Trove of Benefits and
Variety at the Table: A Fourteenth-Century Egyptian Cookbook.
Brill, pp529-575, 2017.
|
|
13
|
Asalgoo S, Tat M, Sahraei H and Pirzad
Jahromi G: The psychoactive agent crocin can regulate
hypothalamic-pituitary-adrenal axis activity. Front Neurosci.
11(668)2017.PubMed/NCBI View Article : Google Scholar
|
|
14
|
Mokhtari Hashtjini M, Pirzad Jahromi G,
Meftahi GH, Esmaeili D and Javidnazar D: Aqueous extract of saffron
administration along with amygdala deep brain stimulation promoted
alleviation of symptoms in post-traumatic stress disorder (PTSD) in
rats. Avicenna J Phytomed. 8:358–369. 2018.PubMed/NCBI
|
|
15
|
Saleem S, Ahmad M, Ahmad AS, Yousuf S,
Ansari MA, Khan MB, Ishrat T and Islam F: Effect of saffron
(Crocus sativus) on neurobehavioral and neurochemical
changes in cerebral ischemia in rats. J Med Food. 9:246–253.
2006.PubMed/NCBI View Article : Google Scholar
|
|
16
|
Vakili A, Einali MR and Bandegi AR:
Protective effect of crocin against cerebral ischemia in a
dose-dependent manner in a rat model of ischemic stroke. J Stroke
Cerebrovasc Dis. 23:106–113. 2014.PubMed/NCBI View Article : Google Scholar
|
|
17
|
Tarantilis PA, Tsoupras G and Polissiou M:
Determination of saffron (Crocus sativus L.) components in
crude plant extract using high-performance liquid
chromatography-UV-visible photodiode-array detection-mass
spectrometry. J Chromatogr A. 699:107–118. 1995.PubMed/NCBI View Article : Google Scholar
|
|
18
|
Hosseinzadeh H and Younesi HM:
Antinociceptive and anti-inflammatory effects of Crocus sativus
L stigma and petal extracts in mice. BMC Pharmacol.
2(7)2002.PubMed/NCBI View Article : Google Scholar
|
|
19
|
Farahmand SK, Samini F, Samini M and
Samarghandian S: Safranal ameliorates antioxidant enzymes and
suppresses lipid peroxidation and nitric oxide formation in aged
male rat liver. Biogerontology. 14:63–71. 2013.PubMed/NCBI View Article : Google Scholar
|
|
20
|
Samarghandian S, Borji A, Delkhosh MB and
Samini F: Safranal treatment improves hyperglycemia, hyperlipidemia
and oxidative stress in streptozotocin-induced diabetic rats. J
Pharm Pharm Sci. 16:352–362. 2013.PubMed/NCBI View
Article : Google Scholar
|
|
21
|
Malaekeh-Nikouei B, Mousavi SH, Shahsavand
S, Mehri S, Nassirli H and Moallem SA: Assessment of cytotoxic
properties of safranal and nanoliposomal safranal in various cancer
cell lines. Phytother Res. 27:1868–1873. 2013.PubMed/NCBI View
Article : Google Scholar
|
|
22
|
Imenshahidi M, Hosseinzadeh H and
Javadpour Y: Hypotensive effect of aqueous saffron extract
(Crocus sativus L.) and its constituents, safranal and
crocin, in normotensive and hypertensive rats. Phytother Res.
24:990–994. 2010.PubMed/NCBI View
Article : Google Scholar
|
|
23
|
Baluchnejadmojarad T, Mohamadi-Zarch SM
and Roghani M: Safranal, an active ingredient of saffron,
attenuates cognitive deficits in amyloid β-induced rat model of
Alzheimer's disease: Underlying mechanisms. Metab Brain Dis.
34:1747–1759. 2019.PubMed/NCBI View Article : Google Scholar
|
|
24
|
Bo-Qiang L, Si-Tong Z, Zu-Yuan L, Wan-Yun
N, Bin C, Yuan L, Xuyun L, Liangen M, You-Chao C, Xin-Zhen Y, et
al: Safranal carried by nanostructured lipid vehicles inhibits
generalized epilepsy in mice. Pharmazie. 73:207–212.
2018.PubMed/NCBI View Article : Google Scholar
|
|
25
|
Zhang C, Ma J, Fan L, Zou Y, Dang X, Wang
K and Song J: Neuroprotective effects of safranal in a rat model of
traumatic injury to the spinal cord by anti-apoptotic,
anti-inflammatory and edema-attenuating. Tissue Cell. 47:291–300.
2015.PubMed/NCBI View Article : Google Scholar
|
|
26
|
Pan PK, Qiao LY and Wen XN: Safranal
prevents rotenone-induced oxidative stress and apoptosis in an in
vitro model of Parkinson's disease through regulating Keap1/Nrf2
signaling pathway. Cell Mol Biol (Noisy-le-grand). 62:11–17.
2016.PubMed/NCBI
|
|
27
|
Sadeghnia HR, Shaterzadeh H, Forouzanfar F
and Hosseinzadeh H: Neuroprotective effect of safranal, an active
ingredient of Crocus sativus, in a rat model of transient
cerebral ischemia. Folia Neuropathol. 55:206–213. 2017.PubMed/NCBI View Article : Google Scholar
|
|
28
|
Hosseinzadeh H and Sadeghnia HR: Safranal,
a constituent of Crocus sativus (saffron), attenuated
cerebral ischemia induced oxidative damage in rat hippocampus. J
Pharm Pharm Sci. 8:394–399. 2005.PubMed/NCBI
|
|
29
|
Forouzanfar F, Asadpour E, Hosseinzadeh H,
Boroushaki MT, Adab A, Dastpeiman SH and Sadeghnia HR: Safranal
protects against ischemia-induced PC12 cell injury through
inhibiting oxidative stress and apoptosis. Naunyn Schmiedebergs
Arch Pharmacol. 394:707–716. 2021.PubMed/NCBI View Article : Google Scholar
|
|
30
|
Meng X, Wang M, Wang X, Sun G, Ye J, Xu H
and Sun X: Suppression of NADPH oxidase- and mitochondrion-derived
superoxide by Notoginsenoside R1 protects against cerebral
ischemia-reperfusion injury through estrogen receptor-dependent
activation of Akt/Nrf2 pathways. Free Radic Res. 48:823–838.
2014.PubMed/NCBI View Article : Google Scholar
|
|
31
|
Belayev L, Alonso OF, Busto R, Zhao W and
Ginsberg MD: Middle cerebral artery occlusion in the rat by
intraluminal suture. Neurological and pathological evaluation of an
improved model. Stroke. 27:1616–1623. 1996.PubMed/NCBI View Article : Google Scholar
|
|
32
|
Alinejad B, Ghorbani A and Sadeghnia HR:
Effects of combinations of curcumin, linalool, rutin, safranal, and
thymoquinone on glucose/serum deprivation-induced cell death.
Avicenna J Phytomed. 3:321–328. 2013.PubMed/NCBI
|
|
33
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408.
2001.PubMed/NCBI View Article : Google Scholar
|
|
34
|
Patel AR, Ritzel R, McCullough LD and Liu
F: Microglia and ischemic stroke: A double-edged sword. Int J
Physiol Pathophysiol Pharmacol. 5:73–90. 2013.PubMed/NCBI
|
|
35
|
Zhu J, Dou S, Jiang Y, Chen J, Wang C and
Cheng B: Apelin-13 protects dopaminergic neurons in MPTP-induced
Parkinson's disease model mice through inhibiting endoplasmic
reticulum stress and promoting autophagy. Brain Res. 1715:203–212.
2019.PubMed/NCBI View Article : Google Scholar
|
|
36
|
Herskovits AZ and Guarente L: SIRT1 in
neurodevelopment and brain senescence. Neuron. 81:471–483.
2014.PubMed/NCBI View Article : Google Scholar
|
|
37
|
Edelbrock AN, Àlvarez Z, Simkin D, Fyrner
T, Chin SM, Sato K, Kiskinis E and Stupp SI: Supramolecular
nanostructure activates TrkB receptor signaling of neuronal cells
by mimicking brain-derived neurotrophic factor. Nano Lett.
18:6237–6247. 2018.PubMed/NCBI View Article : Google Scholar
|
|
38
|
Dimyan MA and Cohen LG: Neuroplasticity in
the context of motor rehabilitation after stroke. Nat Rev Neurol.
7:76–85. 2011.PubMed/NCBI View Article : Google Scholar
|
|
39
|
Liu Y, Li C, Wang J, Fang Y, Sun H, Tao X,
Zhou XF and Liao H: Nafamostat mesilate improves neurological
outcome and axonal regeneration after stroke in rats. Mol
Neurobiol. 54:4217–4231. 2017.PubMed/NCBI View Article : Google Scholar
|
|
40
|
Virani SS, Alonso A, Aparicio HJ, Benjamin
EJ, Bittencourt MS, Callaway CW, Carson AP, Chamberlain AM, Cheng
S, Delling FN, et al: Heart disease and stroke statistics-2021
update: A report from the american heart association. Circulation.
143:e254–e743. 2021.PubMed/NCBI View Article : Google Scholar
|
|
41
|
Liu W, Wang X, O'Connor M, Wang G and Han
F: Brain-derived neurotrophic factor and its potential therapeutic
role in stroke comorbidities. Neural Plast.
2020(1969482)2020.PubMed/NCBI View Article : Google Scholar
|
|
42
|
Fonarow GC, Smith EE, Saver JL, Reeves MJ,
Bhatt DL, Grau-Sepulveda MV, Olson DM, Hernandez AF, Peterson ED
and Schwamm LH: Timeliness of tissue-type plasminogen activator
therapy in acute ischemic stroke: Patient characteristics, hospital
factors, and outcomes associated with door-to-needle times within
60 min. Circulation. 123:750–758. 2011.PubMed/NCBI View Article : Google Scholar
|
|
43
|
Yang S, Jin H, Zhu Y, Wan Y, Opoku EN, Zhu
L and Hu B: Diverse functions and mechanisms of pericytes in
ischemic stroke. Curr Neuropharmacol. 15:892–905. 2017.PubMed/NCBI View Article : Google Scholar
|
|
44
|
Zhu T, Wang L, Xie W, Meng X, Feng Y, Sun
G and Sun X: Notoginsenoside R1 improves cerebral
ischemia/reperfusion injury by promoting neurogenesis via the
BDNF/Akt/CREB pathway. Front Pharmacol. 12(615998)2021.PubMed/NCBI View Article : Google Scholar
|
|
45
|
Herpich F and Rincon F: Management of
acute ischemic stroke. Crit Care Med. 48:1654–1663. 2020.PubMed/NCBI View Article : Google Scholar
|
|
46
|
Xu S, Yang J, Wan H, Yu L and He Y:
Combination of radix astragali and safflower promotes angiogenesis
in rats with ischemic stroke via silencing PTGS2. Int J Mol Sci.
24(2126)2023.PubMed/NCBI View Article : Google Scholar
|
|
47
|
Wen HC, Huo YN, Chou CM and Lee WS: PMA
inhibits endothelial cell migration through activating the
PKC-δ/Syk/NF-κB-mediated up-regulation of Thy-1. Sci Rep.
8(16247)2018.PubMed/NCBI View Article : Google Scholar
|
|
48
|
Yao Y: Basement membrane and stroke. J
Cereb Blood Flow Metab. 39:3–19. 2019.PubMed/NCBI View Article : Google Scholar
|
|
49
|
Save SS, Rachineni K, Hosur RV and
Choudhary S: Natural compound safranal driven inhibition and
dis-aggregation of α-synuclein fibrils. Int J Biol Macromol.
141:585–595. 2019.PubMed/NCBI View Article : Google Scholar
|
|
50
|
Vogt N: Sensing neurotransmitters. Nat
Methods. 16(17)2019.PubMed/NCBI View Article : Google Scholar
|
|
51
|
Ramer MS, Priestley JV and McMahon SB:
Functional regeneration of sensory axons into the adult spinal
cord. Nature. 403:312–316. 2000.PubMed/NCBI View Article : Google Scholar
|
|
52
|
Wang T, Zhang J, Li P, Ding Y, Tang J,
Chen G and Zhang JH: NT-4 attenuates neuroinflammation via
TrkB/PI3K/FoxO1 pathway after germinal matrix hemorrhage in
neonatal rats. J Neuroinflammation. 17(158)2020.PubMed/NCBI View Article : Google Scholar
|
|
53
|
Michan S and Sinclair D: Sirtuins in
mammals: Insights into their biological function. Biochem J.
404:1–13. 2007.PubMed/NCBI View Article : Google Scholar
|
|
54
|
Zakhary SM, Ayubcha D, Dileo JN, Jose R,
Leheste JR, Horowitz JM and Torres G: Distribution analysis of
deacetylase SIRT1 in rodent and human nervous systems. Anat Rec
(Hoboken). 293:1024–1032. 2010.PubMed/NCBI View Article : Google Scholar
|
|
55
|
Fu B, Zhang J, Zhang X, Zhang C, Li Y,
Zhang Y, He T, Li P, Zhu X, Zhao Y, et al: Alpha-lipoic acid
upregulates SIRT1-dependent PGC-1α expression and protects mouse
brain against focal ischemia. Neuroscience. 281:251–257.
2014.PubMed/NCBI View Article : Google Scholar
|
|
56
|
Lv H, Wang L, Shen J, Hao S, Ming A, Wang
X, Su F and Zhang Z: Salvianolic acid B attenuates apoptosis and
inflammation via SIRT1 activation in experimental stroke rats.
Brain Res Bull. 115:30–36. 2015.PubMed/NCBI View Article : Google Scholar
|
|
57
|
Wan D, Zhou Y, Wang K, Hou Y, Hou R and Ye
X: Resveratrol provides neuroprotection by inhibiting
phosphodiesterases and regulating the cAMP/AMPK/SIRT1 pathway after
stroke in rats. Brain Res Bull. 121:255–262. 2016.PubMed/NCBI View Article : Google Scholar
|