AD is a common neurodegenerative disorder
characterized by gradual cognitive decline, memory loss, and
behavioral changes (1). AD is
mainly characterized by the accumulation of extracellular amyloid β
(Aβ), which forms senile plaques, and intracellular
hyperphosphorylated tau, which binds to microtubules and leads to
the development of neurofibrillary tangles (2). The disease is becoming increasingly
prevalent, with projections estimating a global population of 115
million patients with AD by 2050 (3). With a growing aging population, the
management of AD is becoming increasingly critical.
The pathogenesis of AD is multifactorial and
involves a number of hypotheses, including the cholinergic theory,
the amyloid cascade theory, the oxidative stress theory, the tau
protein hypothesis and the neuroinflammation hypothesis (4). Evidence supports the
neuroinflammation as a crucial factor in the development of AD
(5,6). Neuroinflammation (7-9) is
present in the majority of patients with AD (10) and animal models (11), particularly in the cerebral cortex
and hippocampus (12,13). Elevated levels of inflammatory
factors and increased activation of microglia around senile plaques
observed in patients with AD further support this hypothesis
(14). In addition, whole-genome
studies of post-mortem brain samples from patients with AD have
shown upregulation of inflammation-related genes and significant
downregulation of anti-inflammatory molecules (15). Activated microglia, responding to
Aβ (16), demonstrate a
significant inflammatory response highly correlated with the
severity of AD (17). Taken
together, these findings suggest that neuroinflammatory responses
mediated by microglial cell activation may play a central role in
the pathogenesis of AD.
Under normal circumstances, highly active microglia
cells efficiently monitor the entire brain in real time (18), detecting abnormalities such as
pathogens and cellular debris (19,20) and providing essential support to
maintain optimal brain function (21). However, when the brain is exposed
to abnormal conditions, microglia become activated and switch to a
transforming state, migrating towards the site of injury to remove
pathogens, cellular debris and degenerated cells (22). Depending on their activation state
and environmental stimuli, microglia cells can be classified as
either the pro-inflammatory M1 type or the anti-inflammatory M2
type (23). In the early stages
of AD, microglia play a crucial role in maintaining a dynamic
balance of amyloid protein in the brain by engulfing and clearing
excess Aβ, thereby helping to delay disease progression. However,
as the disease progresses, excessive accumulation of Aβ can lead to
overactivation of microglia cells, causing them to adopt a
pro-inflammatory M1 type (24).
In the central nervous system (CNS), activated microglia are the
primary source of inflammatory molecules, such as cytokines,
chemokines, neurotransmitters, reactive oxygen species (ROS) and
nitric oxide (NO) (25).
Inflammatory molecules trigger a positive feedback mechanism that
activates more microglia and thus further exacerbating the
neuroinflammatory response (25,26). As a result, secreted inflammatory
mediators facilitate the migration of monocytes and lymphocytes to
the site of inflammation, where they penetrate the blood-brain
barrier (BBB), exacerbating CNS inflammation and leading to
sustained neuronal damage (27),
ultimately culminating in cognitive decline. Several inhibitors,
drugs and their active ingredients can exert an
anti-neuroinflammatory effects, with different drugs acting via
single or multiple signaling pathways. Therefore, it is essential
to consolidate research findings to identify potential drug
candidates for the prevention and treatment of AD.
Neuroinflammation is a critical factor and even a
core event in the pathogenesis of AD (17,28). Microglia, as the primary immune
cells in brain tissue, play an essential role in neuroinflammation
through multiple targets and signaling pathways. Therefore, the
development of drugs or inhibitors that target microglia could
alleviate neuroinflammation, which could have a positive effect on
both the prevention and treatment of AD. The present study
conducted a literature search using keywords the 'inhibitors',
'microglia', 'inflammation' and 'Alzheimer's disease' in PubMed
between 2012 and 2022 to comprehensively review the major signaling
pathways involved in microglia activation and the ways in which
drugs exert anti-neuroinflammatory effects by targeting these
pathways. Out of the 327 articles retrieved, 35 were excluded,
including reviews, commentaries, retractions, or unavailability
online. Also excluded were 201 articles that did not involve
signaling pathways. Finally, 96 references were included. In
addition, 'medicine' and 'drugs' were added as keywords to the
search to further identify promising drug candidates for AD
prevention.
NF-κB is widely expressed in brain tissue and plays
a critical regulatory role in various target genes within the CNS.
Its regulatory scope encompasses oxidative stress,
neuroinflammation and microglia activation (31). In particular, excessive activation
of NF-κB has been implicated in the neuropathological features of
AD. Multiple studies have identified increased activation of NF-κB
in the brains of patients with AD (31,37), particularly in the most affected
brain regions (38-40). Additionally, the activation of
NF-κB by Aβ leads to further production of Aβ, exacerbating the
pathology of AD (41,42). Moreover, NF-κB not only acts
downstream of tau but also seems to directly mediate its cognitive
toxicity (43). This increased
DNA-binding activity of NF-κB leads to aggravated oxidative stress,
which exacerbates neurotoxicity. In addition, downstream
pro-inflammatory mediators are activated, thereby affecting
neuronal function (44,45). Above all, activation of glial
cells via the NF-κB pathway serves as a critical link in the
neuroinflammatory response (46),
further amplifying neuroinflammation and worsening AD pathology
(47,48). As such, modulation of the NF-κB
signaling pathway in microglia may represent a promising new
approach to the prevention and treatment of AD.
Studies have shown that certain compounds found in
traditional Chinese herbal medicine possess the capacity to inhibit
NF-κB activation and exert anti-inflammatory effects. Rutin, a
natural flavonoid glycoside with anti-inflammatory and antioxidant
properties (49), is a promising
neuroprotective agent for neurodegenerative diseases (50). A recent study has revealed that
treatment with Rutin can reduce NF-κB activation in the Tau-P301S
mouse, resulting in lower levels of IL-1 and TNF-α in brain tissue,
thereby counteracting neuroinflammation (51). Results consistent with in
vivo findings were also observed in microglia induced with tau
oligomers (51). Similarly,
piperlongumine, an alkaloid amide from Piper longum, was
found to be neuroprotective effects (52) against lipopolysaccharide
(LPS)-induced neuroinflammation by inhibiting the NF-κB pathway and
reducing the expression of key pro-inflammatory mediators such as
cyclooxygenase-2 (COX-2), inducible nitric oxide synthase (iNOS),
TNF-α, IL-1β, and IL-6. Thus, these compounds show therapeutic
potential for the treatment of neuroinflammatory disorders by
modulating the NF-κB signaling pathway in microglia (53). Bee venom, which contains various
peptides, enzymes, and biogenic amines, has been shown to be
effective in the treatment of diseases such as arthritis,
rheumatism and cancer (54). A
study has highlighted its potential for treating AD by inhibiting
the expression of neuroinflammatory proteins such as β-site amyloid
precursor protein cleaving enzyme 1 (BACE1), COX-2, iNOS, glial
fibrillary acidic protein (GFAP), and ionized calcium binding
adaptor molecule 1, in vitro and in vivo, through
inactivation of the NF-κB pathway, resulting in a reduction in
LPS-induced memory impairment (55). Punicalagin, a polyphenol sourced
from pomegranate fruit, has antioxidant, anti-proliferative and
anti-inflammatory properties (56). It has been shown to bind directly
to NF-κB, impede IκB degradation and prevent the nuclear
translocation of p50 and p65, thereby inhibiting the production of
ROS, NO, TNF-α and IL-1β in LPS-induced BV-2 microglia (57). Similarly, tenuifolin, a valuable
neuroprotective compound extracted from Polygala tenuifolia
Willd, can block the activation of the NF-κB pathway and
subsequently improve cognitive impairment symptoms in AD (58). Piperine, a crystalline alkaloid
extracted from pepper, has several properties such as
anticarcinogenic, stimulatory, anti-inflammatory and antiulcer
activities (59). Furthermore,
piperine derivatives, such as (2E,4E)-5-(benzo[d][1,3]dioxo
l-5-yl)-N-[4-(hydroxymethyl) phenyl] penta-2,4-dienamide (D4) have
demonstrated anti-neuroinflammatory effects (60) by inhibiting the translocation of
NF-κB and suppressing the expression of iNOS and the secretion of
NO, TNF-α, and IL-1β in LPS-induced human microglia clone 3. In
addition, an in silico study showed excellent D4
bioavailability after oral administration (61). Bupleurum falcatum L. (BF)
is a traditional oriental medicine commonly used in the treatment
of chronic hepatitis and autoimmune diseases (62). It has been demonstrated that the
ethanol extract of BF (BFE) can inhibit the expression of
pro-inflammatory genes and NF-κB p65/RELA mRNA in BV2 microglia
that have been activated with LPS. This suggests that NF-κB is a
molecular target of BFE (63). In
addition, BFE has been shown to inhibit the activation of microglia
in the hippocampus and substantia nigra of LPS-treated mice
(63), suggesting its potential
as a treatment for AD. Similarly, macasiamenene F (MF), a compound
extracted from Macaranga siamensis S. J. Davies
(Euphorbiaceae), has also been shown to have promising potential in
the treatment of neuroinflammatory responses. MF treatment
significantly suppresses NF-κB activity and TNF-α expression in
LPS-induced human monocytes (64), and similar responses may occur in
microglia of brain given their phenotypic similarity. Miconazole
(MCZ) is an azole drug commonly used as an antifungal agent that
can cross the BBB and exhibits neuroprotective effects (64,65). MCZ can reduce the expression of
ionized calcium binding adaptor molecule 1 (Iba-1) reactive cells
and downregulate the expression of GFAP, Iba-1, and COX-2 in the
hippocampus by inhibiting the NF-κB signaling pathway in a mouse
model of Aβ1-42-induced memory impairment. This
anti-inflammatory effect of MCZ was further confirmed in an
LPS-induced BV2 microglia model (66).
Several drugs have been developed to target specific
components of the body and exert anti-neuroinflammatory effects by
inhibiting NF-κB (67-70). Among these, LD55, a resveratrol
analogue, is widely used as a novel inhibitor of NF-κB activation
(71). A study has shown that
dietary supplementation with LD55 can effectively suppress the
activation of microglia in transgenic amyloid-β
protein/presenilin-1 (APP/PS1) mice, diminish the density of Aβ
plaques in the brain and notably reduce them by 2-15 times in the
hippocampal region. These findings suggest that LD55 may provide
some relief from the burden of Aβ plaques and neuroinflammation in
AD models (67). Additionally,
glucocorticoid-induced leucine zipper (GILZ), which functions as a
transcriptional regulatory protein, has the ability to impede the
activity of NF-κB (72,73). A small molecule GILZ analogue, GA,
was found to inhibit the levels of NF-κB p65 in the brains of 5XFAD
(familial Alzheimer's disease) mice. Furthermore, GA can
downregulate the expression of inflammatory factors while hindering
the proliferation and activation of hippocampal microglia (68). Consequently, this leads to the
suppression of neuroinflammation. Chitinase-3 like-protein-1
(CHI3L1) is a secreted, inflammatory glycoprotein that is expressed
in a number of chronic neuroinflammatory diseases including AD,
making it a potential biomarker for AD diagnosis (74). Conversely, CHI3L1 deficiency has
been shown to attenuate microglia-mediated inflammation and inhibit
the progression of AD (75,76). Study has shown that the CHI3L1
inhibitor, K284-6111, can suppress NF-κB activation and the
expression of related inflammatory factors in AD animal models
following intracerebroventricular infusion of Aβ1-42 and
in LPS-induced BV-2 microglia cells (69). Furthermore, the
anti-neuroinflammatory effects of K284-6111 are also observed in a
Tg2576 mouse model and in Aβ-induced BV2 microglia, implicating the
extracellular signal-regulated kinases (ERK)-mediated pentraxin 3
and NF-κB pathways (16). DL0410,
an acetylcholinesterase (AChE) inhibitor, has been shown to
suppress the receptor for advanced glycation end products
(RAGE)/NF-κB signaling pathway, resulting in inhibition of
D-galactose-induced microglia activation. This results in the
downregulation of COX2 and iNOS expression, ultimately suppressing
inflammation in the cortex and hippocampus of the brain (70).
Toll-like receptors (TLRs) are essential pattern
recognition receptors in the immune and inflammatory responses,
with TLR4 being highly expressed on microglia (77). However, excessive activation of
TLRs can initiate a cascade of events, leading to activation of
NF-κB in the brain, resulting in the synthesis and release of
various inflammatory mediators that contribute to neuronal damage
(78,79). Therefore, targeting the TLR/NF-κB
pathway may prove beneficial in the treatment of AD. Several
studies have illustrated that natural compounds can reduce
neuroinflammation by inhibiting the TLR4/NF-κB pathway (80-82). One such compound is
epigallocatechin-3-gallate (EGCG), a polyphenol found in green tea
that has been extensively studied for its neuroprotective effects
(83). EGCG is known to suppress
the activation of both classical NOD-like receptor thermal protein
domain associated protein 3 (NLRP3) inflammasomes and
caspase-11-mediated non-classical inflammasomes via the TLR4/NF-κB
pathway, thereby effectively exerting its anti-inflammatory
properties (80). Genistein
(Gen), a compound derived from Soybean isoflavone (SIF) (84), has been shown to improve memory
abilities in patients with AD and to attenuate inflammation in
Aβ25-35-induced BV-2 microglia through inhibition of the
TLR4/NF-κB signaling pathway. These findings suggest that a diet
rich in plant-derived Gen may be beneficial in reducing the risk of
AD by alleviating inflammation (81). In addition, oxysophoridine
extracted from Sophora alopecuroides L. seeds (85,86) was found to downregulate the
expression of TNF-α and IL-1β in Aβ-induced BV-2 cells, with
therapeutic effects comparable to those of the TLR4 inhibitor
TAK-242. These results demonstrate the promising
anti-neuroinflammatory properties of oxysophoridine (82). The initial interaction between
CD14 and TLR4 is a crucial step in the activation of
neuroinflammatory signals induced by LPS (87). A study has identified a novel
biphenyl compound, called Protosappanin A (PTA), derived from
Caesalpinia sappan L., which effectively inhibits
neuroinflammation in vitro (88). PTA achieves this by disrupting the
CD14-TLR4 interaction in BV-2 microglia that are stimulated by LPS,
thereby inhibiting the NF-κB signaling pathway (88). Similarly, resveratrol, a natural
neuroprotectant agent, has been shown to significantly reduce
microglia-mediated neuroinflammation (89). Oral administration of resveratrol
to APP/PS1 mice significantly reduced the number of activated
microglia around amyloid plaques (90). Further in vitro research
revealed that resveratrol's mechanism of action involves disruption
of TLR4 oligomerization to attenuate the TLR4/NF-κB/STAT signaling
pathway, ultimately leading to a reduction in TNF-α and IL-6
production (90).
Upon activation, TLR4 recruits the adaptor myeloid
differentiation factor 88 (MyD88), which initiates downstream
activation of the transcription factor NF-κB (91). Certain active compounds in some
traditional Chinese medicines have been found to interfere with
this pathway and exert anti-neuroinflammatory effects. For example,
Icariside II (ICS II), an active component of Epimedium, has
been shown to have multiple pharmacological activities, including
anti-inflammatory, anticancer and anti-aging (92,93). In an LPS-induced SD rat model of
neuroinflammation, ICS II demonstrated potent anti-inflammatory
effects by reducing the expression of the microglia marker Iba-1
and downregulating related pro-inflammatory cytokine proteins by
intervening in the TLR4/MyD88/NF-κB pathway (94). Similarly, DL0410 is a dual
inhibitor of both AChE and butyrylcholinesterase with a unique
structural scaffold (95). This
compound has been shown to improve memory when administered with
Aβ1-42 and scopolamine administration (96), as well as cognitive impairment
when administered with D-galactose. It holds significant potential
as a therapeutic agent for AD by inhibiting the
TLR4-mediated/MyD88/NF-κB signaling pathway and reducing
pro-inflammatory cytokines (such as TNF, IL-1 and IL-6), while
increasing the anti-inflammatory cytokine IL-10 to combat
neuroinflammation (97). ATP50-3
is a purified product that is extracted from crude polysaccharides
obtained from the traditional Chinese medicine Acorus
tatarinowii (98,99). In vitro study has shown
that it effectively inhibits the activation of NF-κB and the
expression of TLR4, MyD88, phosphorylated (p)-PI3K
(phosphoinositide 3-kinase), p-Akt (p-, phosphorylated), and
inflammatory mediators in LPS-induced BV2 cells (100). Moreover, its anti-inflammatory
efficacy is further enhanced by the TLR4 inhibitor TAK242 and the
PI3K inhibitor LY294002, suggesting that its neuroprotective
effects against neuroinflammation are due to the regulation of the
TLR4/MyD88/NF-κB and PI3K/Akt signaling pathways (100). Another natural compound,
dihydromyricetin (DHM) from Ampelopsis grossedentata, has
also been found to exhibit promising anti-inflammatory effects
(101) and is being considered
as a potential treatment for AD. In an LPS-induced inflammation
model of BV-2 microglia, DHM was found to downregulate
pro-inflammatory cytokine mRNA expression by inhibiting TLR4 and
MyD88 expression, and activation of the NF-кB pathway induced by
LPS (102). These results
strongly suggest that DHM exerts anti-inflammatory effects through
inhibition of the TLR4/MyD88/NF-кB signaling pathway (102). GX-50, a compound derived from
Sichuan pepper, exhibits promising anti-inflammatory and AD
therapeutic effects (103).
Research has shown that GX-50 effectively inhibits Aβ-induced TLR4
activation, preventing the recruitment of MyD88 and TNF receptor
associated factor 6. This ultimately suppresses the NF-κB and MAPK
signaling pathways, demonstrating potent anti-inflammatory activity
(104). WD repeat and FYVE
domain-containing 1 (WDFY1), a pivotal adaptor molecule in the
TLR3/TLR4 signaling pathway, facilitates the recruitment of the
downstream molecule TRIF found on intracellular vesicles, leading
to a pro-inflammatory effect (105,106). Forsythoside B (FTS-B), a
phenylethanoid glycoside derived from Forsythiae fructus,
has been found to possess significant anti-inflammatory properties
and exhibit neuroprotective benefits in AD (107). In vivo study has revealed
that FTS-B can ameliorate cognitive impairment, mitigate
pathological changes and decrease the production of
pro-inflammatory cytokines in mice with AD (108). Consistent with these findings,
FTS-B has been shown to suppress the inflammatory response of
LPS-induced BV-2 microglia and hippocampal HT22 cells in
vitro by blocking the WDFY1/TLR3/NF-κB signaling pathway
(108).
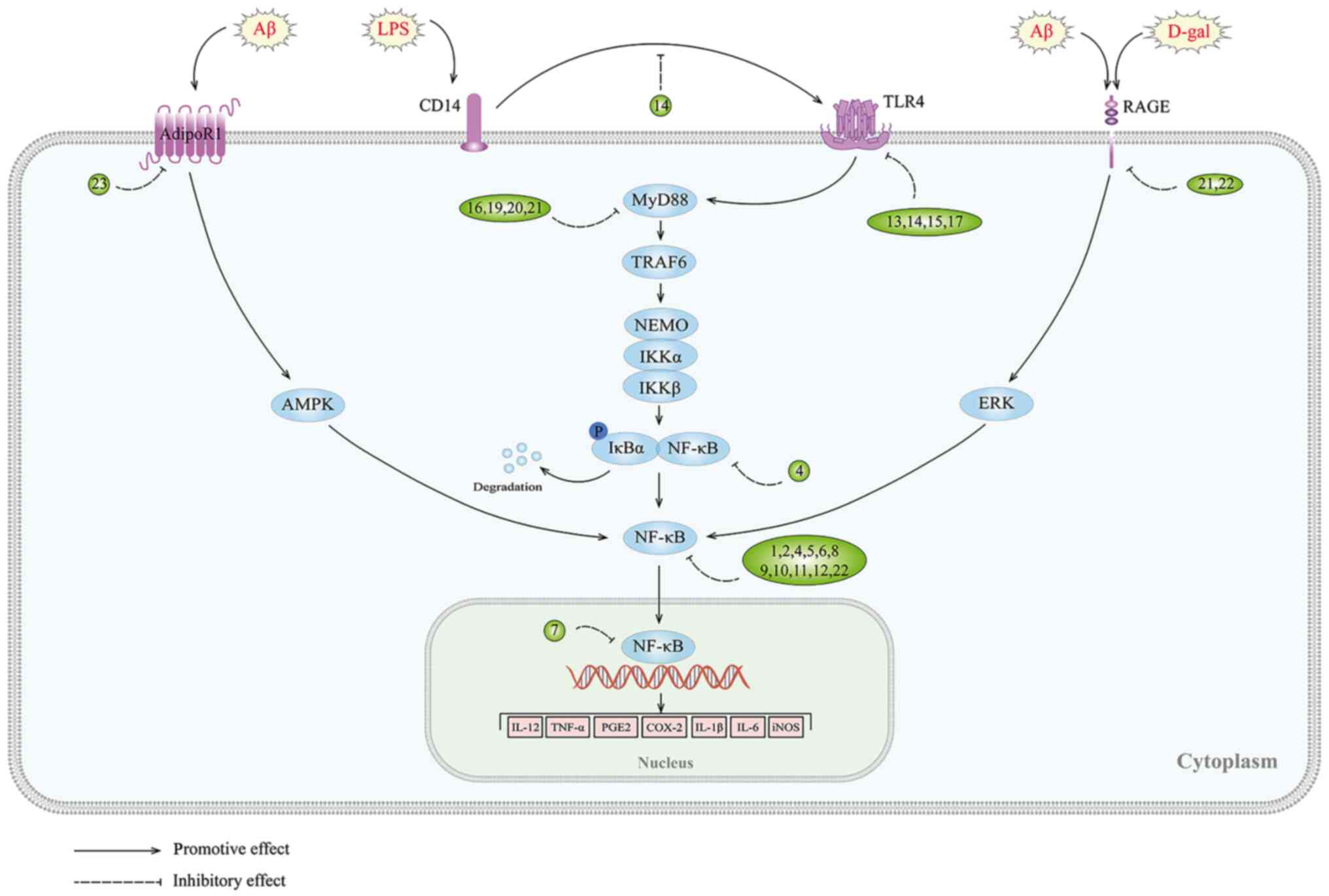
Adiponectin (APN) is an adipokine that is produced
by adipocytes that binds to the AdipoR1 and AdipoR2 receptors
(117). In aged mice, chronic
deficiency of APN has been associated with cognitive impairment and
the development of AD-like symptoms (118). It has also been revealed that
APN deficiency exacerbates microglia activation and
neuroinflammation in APN 5XFAD mice (119). Pre-treatment with APN can
inhibit the release of TNFα and IL-1β in AβO-induced BV2 cells by
activating the AdipoR1/Adenosine 5′-monophosphate (AMP)-activated
protein kinase (AMPK)/NF-κB signaling pathway, thereby ameliorating
neuroinflammation (120). This
research highlights the potential therapeutic benefits of APN in
the prevention and treatment of AD (Fig. 1 and Table I).
It is widely recognized that MAPKs, which include
p38 MAPK, ERK, and c-Jun NH2-terminal kinases (JNK), as well as
their isoforms (121), play a
critical role in the regulation of various biological processes,
including proliferation, differentiation, apoptosis and
inflammation in mammalian cells (122). The MAPK signaling cascade
comprises a MAPKK kinase, a MAPK kinase, and a MAP kinase (123) that respond to both internal and
external stimuli, such as growth factors, cytokines, oxidation, and
endoplasmic reticulum stress. Activation of the MAPK signaling
pathway has been observed in the brains of patients with AD
(124,125) and animal models (126). In vitro studies have
shown that stimulation of Aβ induces the activation of this pathway
in glial cell cultures, indicating its involvement in the
development of AD (127-129). Inhibition of tau kinases, such
as p38 MAPK, has been shown to improve cognitive deficits and
reduce tau pathology in AD (130). Furthermore, blocking the ERK
pathway can reverse mitochondrial dysfunction in AD (131,132), while specific JNK inhibitors can
enhance synaptic function (133). Of note, the MAPK signaling
pathway can also regulate the neuroinflammatory response of
microglia. Aβ-induced production of inflammatory cytokines and ROS
can activate this pathway, leading to more severe inflammation. A
number of in vitro experiments have demonstrated that
inhibition of the MAPK signaling pathway can suppress
neuroinflammation in BV2 microglia (134,135), highlighting its potential as an
effective strategy for treating AD (Fig. 2 and Table II).
P38, a member of the P38 MAPK subfamily, has been
found to be activated in both AD brain tissue samples (136) and animal models (126) of AD. Additionally, study has
shown that the absence of P38 MAPK attenuates amyloid-like
pathology in AD models (137).
Specifically, P38α MAPK is thought to play a crucial role in the
dysregulation of microglia and neuroinflammation during AD
progression, making it a recognized target for AD treatment
(130,138,139). Thus, targeting P38α MAPK may
offer a promising therapeutic strategy to address the underlying
neuroinflammatory processes in AD.
Several inhibitors of the p38α MAPK signaling
pathway, including natural product extracts, and organic compounds,
have shown promise in reducing neuroinflammation and treating AD.
Both preclinical and clinical trials have evaluated the
pharmacological effects of these inhibitors in the brain. Selective
p38α MAPK inhibitors, such as MW01-2-069A-SRM (140) and MW181 (141), which are able to penetrate the
BBB, have demonstrated potent inhibitory effects on
neuroinflammation. Additionally, VX-745, a small molecule inhibitor
of p38α MAPK, has emerged as a promising candidate for
anti-inflammatory therapy and is currently undergoing pilot trials
for the treatment of rheumatoid arthritis (142). Notably, preclinical studies have
revealed that VX-745 exerts its anti-neuroinflammatory effects by
selectively targeting of p38α MAPK, resulting in a reduction of
IL-1β levels in the hippocampus of aged rats (143). This finding highlights the
potential utility of VX-745 as a therapeutic strategy for
neurological disorders characterized by neuroinflammation (143). MAPK-activated protein kinase II
(MK2), a downstream kinase of p38 MAPK (144), is activated and upregulated in
AD mouse models and is associated with Aβ deposition, microglia
activation, and the upregulation of pro-inflammatory cytokines
(145). Targeting MK2 may be a
promising therapeutic strategy for AD. MMI-0100, a cell-penetrating
peptide inhibitor of MK2 with anti-inflammatory activity (146), has been shown to inhibit
LPS-induced microglia activation and significantly reduce
pro-inflammatory cytokine production in mice by inhibiting MK2
phosphorylation (147).
Furthermore, intranasal administration of MMI-0100 can overcome the
challenge of failed AD treatments with large molecule protein or
peptide drugs due to its ability to penetrate the BBB (148,149).
Quinoline, a heterocyclic aromatic organic compound,
has attracted considerable attention for its antibacterial
properties (154) and its
ability to inhibit amyloid aggregation (155,156). Consequently, this framework is
widely utilized in the research and design of innovative
anti-inflammatory drugs. Cryptolepine, an indoloquinoline alkaloid
isolated from Cryptolepis sanguinolenta, has demonstrated
the ability to suppress LPS-induced microglia inflammation by
selectively targeting the NF-κB and p38 MAPK signaling pathways
(157). Similarly, VB-037
(155), a quinoline compound,
has been shown to effectively mitigate BV-2 microglial activation
induced by LPS/interferon-γ (IFN-γ). This attenuation is achieved
by inhibiting caspase 1 activation, IL-1β expression and P38
phosphorylation, as well as by affecting the JNK, Jun oncogene and
Jun signaling pathways. These findings substantiate that VB-037
selectively regulates the P38 and JNK/MAPK signaling pathways,
ameliorating neuronal damage and neuroinflammation and thereby
altering the progression of AD. The multifaceted mechanism of
quinoline derivatives offers several opportunities for the
development of AD therapeutics (155,157).
Numerous inhibitors of the MEK (mitogen-activated
extracellular signal-regulated kinase)/ERK signaling pathway, both
natural product extracts, and organic compounds, have displayed
potential in reducing neuroinflammation and treating AD. Recent
study has highlighted the efficacy of AZD6244 (163), an oral MEK1/2 inhibitor, in
suppressing acrolein-induced neuroinflammation by modulating of the
MEK/ERK signaling pathway in BV-2 cells, leading to its
neuroprotective effects (163).
Similarly, Dexmedetomidine (164), an α2 adrenergic receptor agonist
with sedative, analgesic and anxiolytic properties, was found to
upregulate anti-inflammatory cytokines and M2 phenotype markers,
while downregulating pro-inflammatory cytokines, M1 phenotype
markers, and p-ERK1/2 in LPS-stimulated BV2 microglia. This effect
has been shown to be reversed by LM22B-10, an ERK agonist,
supporting the notion that Dexmedetomidine promotes M2 polarization
in microglia through modulation of the ERK signaling pathway,
ultimately exerting its anti-inflammatory properties (164). Hominis placenta (HP) is a
dried placental extract from pregnant women after delivery that has
been shown to promote neural regeneration (165). Lee et al (166) demonstrated that pre-treatment
with HP significantly inhibited the expression of iNOS and COX2 in
LPS-induced BV2 cells. This anti-inflammatory effect was achieved,
at least in part, through the inhibition of the ERK pathway and the
phosphorylation of JNK and ERK. In addition, Bergamot juice (BJ)
was found to have antibacterial properties and to exert
anti-inflammatory effects (167)
through its flavonoid component (BJe) (168), which was shown to partly affect
the ERK signaling pathway. The critical role of monocytic cells in
neuroinflammation has been underlined by their ability to cross the
BBB and differentiate into microglia in the brain parenchyma
(169,170). In this context, a research team
found that pretreatment with BJe resulted in a
concentration-dependent reduction in the upregulation of
pro-inflammatory cytokine expression and a decrease in the
phosphorylation levels of JNK and ERK1/2 in
Aβ1-42-induced THP-1 monocytic cells. This effect was
associated with the disruption of DNA-binding activity of AP-1
(activator protein 1) and the MAPK/AP-1 pathway, thereby
counteracting the pro-inflammatory activation of
monocytic/microglia induced by Aβ and exerting an
anti-neuroinflammatory effect (168).
The NF-κB and MAPK signaling pathways have emerged
as key regulators of pro-inflammatory mediator expression and NLRP3
inflammasome formation, both of which play a role in
neuroinflammation. Therefore, targeting these signaling pathways
represents a potential therapeutic approach to alleviate
neuroinflammation. Notably, specific inhibitors or drugs have been
found to exhibit dual targeting of both NF-κB and MAPK signaling
pathways, which may provide a more robust anti-neuroinflammatory
effect. This highlights the possibility of developing a combination
therapy targeting multiple pathways for the treatment of
neuroinflammation.
Several synthetic drugs or inhibitors have been
discovered that have anti-neuroinflammatory effects by targeting
the signaling pathways of NF-κB and MAPK. For example, Tripterygium
(TG), a non-steroidal immunosuppressant, has been shown to have
anti-inflammatory, anti-tumor and immunosuppressive properties
(171). Research suggests that
TG can alleviate neuroinflammation by inhibiting the NF-κB and MAPK
signaling pathways, thereby reducing the expression of
Aβ25-35, p-Tau, CD11b and various pro-inflammatory
cytokines in an AD model. This implies the feasibility of TG
intervention in AD pathology (172). A compound called
4-[(5-bromo-3-chloro-2-hydroxybenzyl) amino]-2-hydroxybenzoic acid
(LX007) (173) has been
identified as a potent mitigator of microglia-induced inflammatory
responses. LX007 has demonstrated a significant anti-inflammatory
activity in LPS-stimulated primary microglia inflammation models by
inhibiting the phosphorylation of MAPK and NF-κB p65 nuclear
translocation, effectively inhibiting NO and prostaglandin E2
(PGE2) production and reducing pro-inflammatory cytokine gene and
protein expression (173). These
findings imply that LX007 may be a potential drug for treating
inflammatory reactions. Pseudane-VII, a secondary metabolite
derived from Pseudoalteromonas sp. M2, has been shown to
possess anti-inflammatory activity (173) by inhibiting the phosphorylation
of p38, ERK1/2, JNK1/2 and NF-κB. Similarly, diammonium
glycyrrhizinate (DG), the salt form of glycyrrhizin acid (174), has been found to play a critical
role in inhibiting Aβ1-42-induced neuroinflammation by
regulating the MAPK and NF-κB pathways (174). An in vivo study has
revealed that DG can alleviate memory impairment in mice, inhibit
activation of microglia in the hippocampus and reduce the
expression and production of pro-inflammatory mediators (175). Further investigation has
revealed that the anti-inflammatory effect of DG involves
inhibiting the translocation of NF-κB p65 to the nucleus, as well
as reducing the phosphorylation levels of ERK, JNK and p38 MAPK
(175). It is notably that
aldose reductase inhibitors (ARIs) exert their effects by
regulating the ROS/protein kinase C (PKC)-dependent NF-κB and MAPK
signaling pathways. Aldose reductase (AR), a rate-limiting enzyme
in the polyol pathway of glucose metabolism, is a molecular target
in various inflammatory diseases (176). An in vitro study was
conducted to investigate the effects of typical ARIs, sorbinil
(Sor) and zopolrestat (Zol) (177), on Aβ1-42-induced BV-2
microglia. The results demonstrated that both Sor and Zol
significantly inhibited TNF-α secretion, downregulated the
expression of pro-inflammatory genes and proteins via interference
with the NF-κB and MAPK pathways, in addition to inhibiting the
phosphorylation of several PKC subtypes (177). Notably, this inhibition of PKC
was demonstrated to be mediated by reducing intracellular ROS
generation (178). Taken
together, these findings suggest that the anti-neuroinflammatory
effects of ARIs are, at least in part, ROS/PKC dependent (177). However, further in vivo
studies are necessary to confirm the efficacy and safety of ARIs,
as well as to explore their potential for treating
neurodegenerative diseases.
Traditional medicines, natural products, and their
derivatives have demonstrated promising therapeutic properties for
the treatment of neuroinflammation. Artemisiae Iwayomogii Herba
(AIH), a traditional herb (179)
utilized for the treatment of inflammatory conditions, was found to
inhibit LPS-induced neuroinflammation in BV-2 microglia and mice
brains (180). This effect was
achieved by reducing NO production and the expression of
pro-inflammatory mediators, as well preventing the formation of the
NLRP3 inflammasome (180). The
anti-inflammatory effect of AIH is associated with the regulation
of the NF-κB and MAPK signaling pathways (180). Similarly, Ganoderma
lucidum extract (GLE) (181)
has been shown to possess neuroprotective properties (182) and has exhibited efficacy in the
treatment of inflammatory diseases (183). Pretreatment with GLE
downregulates the expression of pro-inflammatory genes in
LPS-stimulated BV-2 microglia by modulating NF-kB and MAPK
signaling pathways, thereby exerting an anti-neuroinflammatory
effect (181). Atractylodis
Rhizoma Alba (ARA) ethanolic extract (ARAE) (134) was also found to have
anti-neuroinflammatory effects in an in vitro inflammatory
model, associated with the inhibition of the NF-κB and MAPK
signaling pathways (184). ARAE
significantly decreased the production of NO and inflammatory
cytokines and inhibited the expression of iNOS and COX-2. Further
analysis indicated that the anti-inflammatory effects of ARAE were
mainly due to inhibition of IκBα degradation, phosphorylation, and
NF-κB p65 nuclear translocation, suggesting a multi-pathway
approach to reducing neuroinflammation (134). Similarly,
1-O-acetylbritannilactone (also termed Inulicin; ABL), a natural
product derived from Inula britannica L. (185) and its derivative 'compound 15'
were found to inhibit neuroinflammation in LPS-induced BV-2
microglia. Compound 15 was found to block NF-κB translocation,
reduce CD14 generation by TLR4 in a dose-dependent manner, and
significantly inhibit p38 MAPK phosphorylation, thereby
downregulating the p38 MAPK inflammatory signaling pathway.
Moreover, compound 15 was found to convert BV-2 microglia from M1
to M2 phenotypes, further enhancing its ability to inhibit
neuroinflammation (185).
Eucommia ulmoides Oliver (Du Zhong) is a renowned
traditional Chinese medicine containing therapeutic chemical
compounds for a variety of diseases (186,187). Its active compounds possess
anti-neuroinflammatory properties, with ulmoidol (ULM) (188) exhibiting the most potent
anti-inflammatory activity. By interfering with TLR4 signaling, ULM
inhibits downstream NF-κB and MAPK pathways, downregulates
pro-inflammatory cytokine expression and production in LPS-induced
BV-2 cells, thereby exerting its anti-neuroinflammatory effects
(188). Another active compound,
circumdatin D, extracted from Aspergillus ochraceus,
possesses dual activity in inhibiting AChE and promoting
anti-inflammatory reactions (189). It significantly inhibits NO
production, TNF-α, and IL-1β release, and reduces iNOS and COX-2
expression in LPS-induced BV-2 cells by inhibiting TLR4-mediated
NF-κB, MAPK, and JAK/STAT inflammatory signaling pathways.
Tectorigenin (TEC), an active ingredient in a number of traditional
medicines with anti-tumor (190)
and antibacterial effects (191), can also be used to treat
neuroinflammation. In in vitro experiments, TEC not only
reduces NF-κB p65 subunit levels but also inhibits ERK and JNK
phosphorylation (192). Notably,
TEC pre-treatment inhibited TLR4, MyD88, and LPS-induced
pro-inflammatory cytokine expression both in vivo and in
vitro, indicating that its anti-inflammatory mechanisms are
closely related to TLR4-MyD88-mediated inhibition of MAPK and NF-κB
(192). These findings suggest
that traditional Chinese herbal ingredients may be effective in
treating neuroinflammatory diseases by inhibiting TLR4 signaling
and downstream inflammatory pathways. Further studies are needed to
explore their potential clinical applications and mechanisms of
action in vivo. In summary, traditional medicines, natural
products, and their derivatives have shown promise in targeting
both NF-κB and MAPK signaling pathways and represent a promising
therapeutic approach for managing AD.
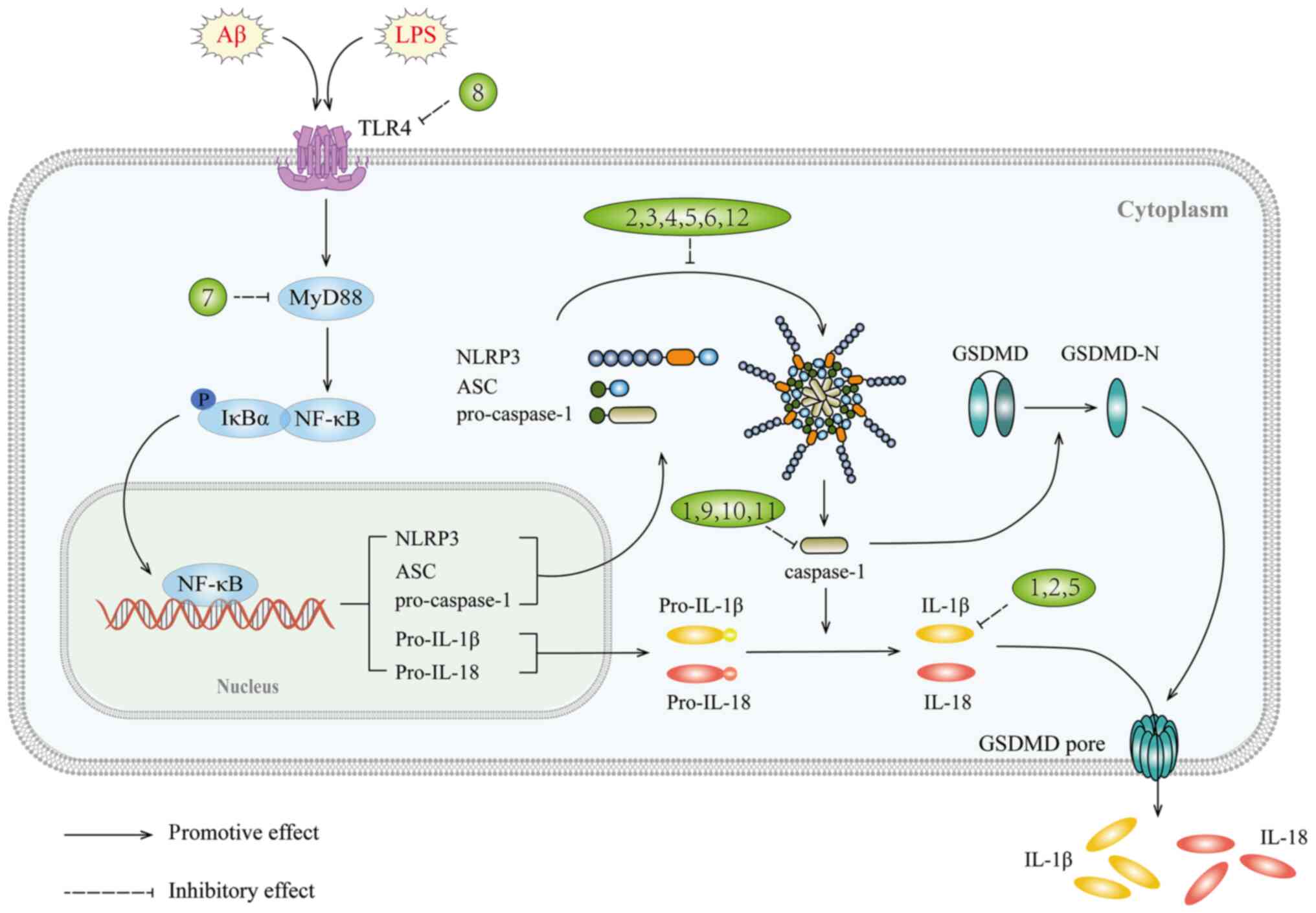
The NLRP3 inflammasome is a multi-protein complex
consisting of the regulatory subunit NLRP3, the adaptor protein
apoptosis-associated speck-like protein (ASC) and the effector
cysteine protease caspase-1 (192) that plays a central role in
sterile inflammatory diseases by regulating the cleavage of IL-1β
precursor (193). The
inflammasome requires two signals for activation: The first signal
triggers the synthesis of IL-1β precursor and other inflammasome
components such as NLRP3 and caspase-1; the second signal leads to
the assembly of the NLRP3 inflammasome, activation of caspase-1,
and secretion of IL-1β (194).
However, dysregulated signal transduction or excessive activation
of the NLRP3 inflammasome can lead to a chronic inflammatory
environment that promotes the pathogenesis and progression of
various diseases, including AD (195). Activated NLRP3 inflammasomes
have been observed in the brains of patients with AD and are
closely associated with microglia. Study has shown that NLRP3
inflammasomes affect Aβ pathology and behavioral deficits in animal
models of AD by modulating the phenotype and function of microglia
(196). Notably, Aβ can also
activate the NLRP3 inflammasome, leading to the release of
proinflammatory cytokines such as IL-1β by microglia, contributing
to neuroinflammation in AD (197). Thus, the NLRP3 inflammasome is a
crucial target in AD and drugs that inhibit its activation through
the inhibition of molecule formation, silencing of upstream
signals, or direct/indirect inhibition of inflammasome complex
formation may prove beneficial.
Inhibitors targeting the NLRP3 inflammasome have
shown efficacy in suppressing neuroinflammation and hold promise as
potential candidates for the prevention and treatment of AD. Among
these inhibitors, dapansutrile (OLT1177) (198), a novel oral agent that
selectively targets the NLRP3 inflammasome, has demonstrated the
ability to block caspase-1 activation and IL-1β maturation and
release. OLT1177 is currently in clinical trials for inflammatory
diseases and has been shown to be well tolerated in humans
(199,200). A study using a APP/PS1 mouse
model demonstrated that OLT1177 treatment can reduce microglia
activation and the number of Aβ plaques in the cortex (198). An in vitro study also
suggested that OLT1177 treatment can significantly reduce the
release of pro-inflammatory cytokines and improve the inflammatory
status of microglia (198).
Similarly, MCC950 (201), a
small molecule inhibitor specific for the NLRP3 inflammasome that
contains a diarylsulfonylurea structure, has shown promise as a
potential treatment for AD. MCC950 has been found to improve
cognitive impairment and reduce Aβ accumulation and microglia
activation in the APP/PS1 mouse model (201). An in vitro study has
shown that MCC950 can inhibit NLRP3 inflammasome activation and
IL-1β release while promoting the phagocytic effect of Aβ in
microglia (201). Similar
results were observed in middle-aged APPNL-F/NL-F mice, where
MCC950 blocked the NLRP3 inflammasome and attenuated the reactive
response of microglia induced by AβO, leading to improvements in
memory impairment (202).
Additionally, a lead compound, JC124 (203), based on sulfonamide-type NLRP3
inhibitors, has recently shown beneficial effects in the prevention
of AD. JC124 has been found to reduce Aβ plaques and microglia
activation in the brains of APP/PS1 mice and has demonstrated
certain anti-inflammatory properties (203).
In addition to specific inhibitors that target the
NLRP3 inflammasome, certain herbal extracts exhibit
anti-inflammatory effects on this pathway. Ginkgolide B (GB)
(204), a plant ester derived
from Ginkgo biloba, has been shown to possess
anti-inflammatory, antioxidant and anti-apoptotic properties, as
well as potent neuroprotective effects (205,206). In an in vitro study, GB
treatment prevented AD pathological processes and suppressed
neuroinflammation in Aβ1-42-induced BV2 microglia by
inhibiting NLRP3 inflammasome activation and promoting M2
polarization (204).
Paeoniflorin (PF) (207), a
natural neuroprotectant from Paeonia lactiflora Pall,
has shown significant therapeutic effects in experimental models of
Parkinson's disease (208) and
stroke (209). Research has
shown that PF significantly reduces the protein levels of the
pro-inflammatory cytokines TNF-α and IL-1β in APP/PS1 mice while
increasing the anti-inflammatory cytokines IL-10 and IL-4. Its
pharmacological effects are achieved by enhancing the activity of
AKT, inhibiting the activation of glycogen synthase kinase-3β
(GSK-3β) and NF-κB p65, and thereby reducing the NLRP3 expression
levels (207).
Controlling the activity of various kinases that
regulate NLRP3 inflammasome activity is another promising way to
suppress neuroinflammation by inhibiting NLRP3 inflammasome
activation. One such enzyme is hematopoietic cell kinase (HCK),
which is involved in a number of inflammatory responses (210). It is suggested that HCK is an
upstream regulator of the NLRP3 inflammasome and that the use of an
HCK inhibitor [A419259 (211), a
Src family kinase-specific inhibitor] can reduce NLRP3
inflammasome-mediated inflammation in microglia. Further
mechanistic studies have shown that the absence of HCK and
inhibition of HCK kinase activity directly affects NLRP3 function
by inhibiting ASC oligomerization and inflammasome assembly. In
vivo experiments confirm that A419259 intervention can
alleviate inflammation in a mouse model of LPS-induced inflammation
(211). Therefore, A419259 may
therefore be a promising drug candidate for the treatment of
diseases associated with NLRP3 inflammasome activation, such as
AD.
Targeting the initial signal for NLRP3 inflammasome
activation has emerged as an effective strategy for the treatment
of neuroinflammation. TAK-242 (212), a cyclohexene derivative, is a
specific small molecule inhibitor of TLR4 that is capable of
crossing the BBB and exerting neuroprotective effects (213). This effect may be mediated
through the modulation of the TLR4/MyD88/NF-κB/NLRP3 signaling
pathway. TAK-242 can reduce TLR4 expression and attenuate
inflammatory cytokine production in microglia from mice with AD
carrying APP/PS1 mutations (212). As a result, there is a
significant decrease in pro-inflammatory M1-type markers, such as
iNOS and TNFα, while M2-type markers, including Trem-2 and Arg-1
are increased (212). Further
investigation has also demonstrated that TAK-242 treatment can
improve the upregulation of inflammatory cytokines, as well as
MyD88, NF-κB p65 and NLRP3 (212). Similarly, the TLR4-specific
inhibitor, CLI-095 (214),
exerts similar anti-inflammatory effects on
LPS/Aβ1-42-induced BV-2 cells and primary microglia by
ameliorating neuroinflammation through the TLR4/NLRP3 pathway
(214).
Activation of the second signal of the inflammasome
is a mechanism by which certain drugs, such as Pterostilbene and
lignin-amides Datura metel seeds (LDS), can inhibit
neuroinflammation. Pterostilbene, a natural compound with
neuroprotective properties (215), has been found to inhibit
Aβ1-42-induced NO production, iNOS mRNA and protein
expression in BV-2 cells, while also reducing the expression and
secretion of inflammatory factors (216). Moreover, pterostilbene can
deactivate the NLRP3/caspase-1 inflammasome activated by
Aβ1-42, demonstrating its anti-inflammatory effects. The
caspase-1 inhibitor, Z-YVAD-FMK, effectively reduces
Aβ1-42-induced neuroinflammation in BV-2 cells,
providing further support for this hypothesis (216). In addition to pterostilbene, LDS
is also able to ameliorate neuroinflammation through the
NLRP3/caspase-1 pathway. Wang et al (217) found that LDS had
anti-inflammatory activity in LPS-induced BV2 cells. Additionally,
PPSR (PEG-PEI/siROCK2), a synthetic molecule used in gene therapy
for AD, was found to inhibit the increase in IL-1β induced by
LPS/Aβ in primary microglia through the NLRP3/caspase-1 pathway,
thus exhibiting anti-inflammatory effects (218). However, the specific mechanism
through which PPSR regulates the NLRP3/caspase-1 pathway remains to
be elucidated and requires further investigation (219).
Gasdermin D (GSDMD) plays a crucial role in
pyroptosis, whereby intracellular inflammasomes trigger
caspase-1-mediated cleavage of the effector protein GSDMD to form
p30-GSDMD, resulting in the formation of cell membrane pores and
release the inflammatory factors (220). Recently, two novel GSDMD
cleavage inhibitors, Sulfa-4 and Sulfa-22 (221), were shown to effectively
attenuate neuroinflammation and prevent AD by disrupting the
NLRP3/caspase-1/GSDMD classical pyroptosis pathway. The
investigation demonstrated that the administration of Sulfa-4 and
Sulfa-22 inhibited the activation of microglia in the brains of
APP/PS1 mice, reduced the expression of inflammatory factors and
suppressed the production of p30-GSDMD and upstream NLRP3
inflammasome and caspase-1 proteins. Furthermore, the study
revealed the specific binding relationship between Sulfa-4 and
Sulfa-22 and the GSDMD protein, establishing a valuable basis for
the development of drugs to target neuroinflammation in AD
(221).
AMPK is a vital molecule that plays a critical role
in regulating energy metabolism and mitochondrial function
(226). Mitochondrial dynamics
are primarily controlled by mitosis (227), which promotes the expression of
phosphate and tensin homolog deleted on chromosome 10
(PTEN)-induced kinase 1 (PINK1) on the damaged outer mitochondrial
membrane. This, in turn, elevates the activity of the E3 ubiquitin
ligase Parkin, modulating the autophagic process (228). Tetrahydroxy stilbene glycoside
(TSG) (229), the major
bioactive component of traditional Chinese medicine Polygoni
multiflori Radix, exhibits potent antioxidant and
anti-atherosclerotic properties (230) and has demonstrated a
neuroprotective in repairing brain injury (231). A recent study has found that TSG
can attenuate the LPS-induced inflammatory response in microglia by
inhibiting the NLRP3 signaling pathway while promoting the
autophagic process mediated by the AMPK/PINK1/Parkin pathway
(229). Notably, the
neuroprotective effect of TSG is abolished in PINK1 or Parkin
knockout models, underscoring the critical role of inhibition of
NLRP3 activation through the AMPK/PINK1/Parkin signaling pathway
for TSG to exert its neuroprotective effects (229).
In addition, RhoA, a member of the Rho family of
GTPases, forms the RhoA/ROCK signaling pathway with the downstream
effector Rho-dependent coiled-coil kinase (ROCK) (232). Activation of this pathway can
further activate NLRP3, leading to neuroinflammation (233) and increased Aβ production
(234) through APP
cleavage-dependent secretion, contributing to Aβ-induced
neurotoxicity. The RhoA/ROCK signaling pathway also affects the
phagocytic function (235) of
microglia and neuroinflammatory responses (236), as well as interactions with Aβ
and microglia (237). A recent
study has demonstrated that small molecule inhibitors, such as
Fasudil and Y27632, can alleviate AD pathogenesis by suppressing
the RhoA/ROCK/NLRP3 signaling pathway, thereby reducing LPS-induced
inflammatory responses (238).
In summary, targeting the NLRP3 inflammasome has
the potential to provide a multitude of effective therapeutic
avenues for managing neuroinflammation in AD (Fig. 3 and Table III).
Peroxisome proliferator-activated receptors (PPARs)
comprise three distinct forms, including PPARα, PPARβ/δ and PPARγ
(239), with a large body of
literature focusing on PPARγ (240-243). These receptors play a
significant role in regulating energy homeostasis and metabolism
(244) throughout the body
(245). In the brain, PPARs are
widely distributed in cognitive centers such as the prefrontal
cortex and hippocampus, which are vulnerable to neurodegeneration
in AD (246). Despite low
baseline expression of PPARγ in the brain, it has been observed to
increase in response to AD pathology (247). Studies have shown that PPARγ
agonists not only improve cognitive function in patients with AD
and animal models (248,249), but also reduce Aβ levels
(250). Furthermore, PPARγ is
highly expressed in microglia (251) and its activation induces
microglia to adopt an anti-inflammatory phenotype, thereby
suppressing neuroinflammatory responses (252,253). These findings highlight PPARγ as
an attractive therapeutic target for the treatment of AD, with the
potential to ameliorate disease pathology.
Current research has demonstrated the
anti-inflammatory effects of PPARγ agonists, particularly
pioglitazone (PIO), in various mouse models of AD. Berberine (BBR),
an alkaloid extracted from Coptidis Rhizoma (254) with similar binding affinity to
the PPARγ protein as PIO, has potentially overlapping effects
(255). BBR has been found to
partially improve neuroinflammation by reducing IL-6 and TNF-α
levels in LPS-induced BV-2 cells, indicating a potential preventive
or delayed onset of early AD (255). Rice bran extract (RBE), a novel
PPARγ regulator that enhances cognitive function in rats (256), also exerts anti-inflammatory
effects by regulating microglia phenotype in LPS-induced mice
(257). RBE and PIO can both
regulate microglia M1 to M2 phenotype, significantly reducing the
expression of NF-κB and pro-inflammatory microglia markers (CD45),
while increasing the expression of anti-inflammatory microglia
markers and PPARγ (257).
Additionally, RBE can reduce Aβ42 deposition and p-tau
protein levels, thereby effectively ameliorating AD pathology
(257).
AD is known to be closely associated with the
activation of inflammation, which can be exacerbated by obesity and
exacerbate cognitive impairment (258). Malva parviflora
extract (MpHE), with its hypoglycemic, anti-inflammatory and
antioxidant properties (258,259), has demonstrated the ability to
improve the adverse effects of a high-fat diet in an AD mouse model
through a PPARγ-dependent mechanism. MpHE not only improved spatial
learning deficits and reduced insoluble Aβ peptides in the
hippocampus of lean and obese 5XFAD mice but also inhibited the
accumulation of small glial cells around Aβ plaques and the
conversion to a pro-inflammatory M1 phenotype while promoting
phagocytic capacity (260).
Rescue of the phagocytic capacity of microglial cells by MpHE was
achieved through a PPARγ/CD36-dependent mechanism (260). Angiotensin II receptor blockers
(ARBs), used to treat metabolic disorders, have been found to
ameliorate inflammation in several brain disorders by blocking
angiotensin II type 1 receptors and activating PPARγ, thus exerting
a neuroprotective effect (261,262). Clinical trials have shown that
ARBs have a positive effect on cognitive decline (263). In addition, it has been shown
that telmisartan, a typical ARB, can ameliorate AβO-induced
inflammation in microglia (264). Telmisartan (264) has been shown to decrease the
expression of the pro-inflammatory cytokine IL-1β, while increasing
the expression of PTEN, a key lipid and protein phosphatase, and
the anti-inflammatory cytokine IL-10. Furthermore, Telmisartan has
also been shown to inhibit the activity of NF-κB, a key
transcription factor involved in inflammation and its upstream
regulators Akt and ERK (264).
These anti-inflammatory effects of telmisartan have been found to
be PPARγ dependent, with the PPARγ inhibitor GW9662 blocking the
expression of PTEN (264). Taken
together, telmisartan ameliorates AβO-induced microglial
inflammation via the PPARγ/PTEN pathways. Other compounds with
potential therapeutic benefit in AD through anti-inflammatory
mechanisms include Bis (ethylmaltolato) oxidovanadium (BEOV) and
platycodigenin. BEOV (265) has
demonstrated the ability to reduce levels of pro-inflammatory
cytokines and interfere with NF-κB signaling in Aβ-stimulated BV2
microglia and the hippocampus of APP/PS1 mice, and its effects have
been found to be PPARγ dependent. Platycodigenin (266), a triterpenoid compound found
mainly in Platycodon grandifloras, demonstrates
neuroprotective and anti-inflammatory activity. Study reveals that
platycodigenin can inhibit the secretion of pro-inflammatory
cytokines in Aβ-stimulated BV-2 microglia and induce M1-type
microglia to polarize towards the M2 type (266). Their anti-neuroinflammatory
effects have been attributed to the inhibition of p38 MAPK and
NF-κB p65 signaling while activating PPARγ. Although PPAR-γ
agonists have shown promising anti-inflammatory activity and their
potential use in the treatment of AD, long-term use of these drugs
often results in serious side effects, including congestive heart
failure, oedema, and weight gain (267,268). Therefore, there is an urgent
need to develop PPARγ-targeting drugs with improved tolerability
(Fig. 4 and Table IV).
STATs are a group of potential transcription
factors that are activated by cytokines and growth factors. When
stimulated by LPS, IFN-γ, and other cytokines, they can trigger
inflammatory signals that translocate STATs from the cytoplasm to
the nucleus and activate the expression of a number of
pro-inflammatory genes (269).
Of the seven types of STAT proteins found in humans, STAT3 has been
extensively studied for its involvement in acute stress responses,
cell growth, differentiation, and immune reactions (270). Previous studies have
demonstrated elevated activation of STAT3 in hippocampal slices in
patients with AD (271) and
mouse models (272).
Furthermore, STAT3 plays a crucial role in regulating the
reactivity of microglia and in mediating pro-inflammatory
responses, indicating a close functional interplay with microglia
(273). Given the dependence of
neuronal differentiation and cytokine signaling on STAT3, STAT3
phosphorylation is closely linked to cytokine secretion (274). Therefore, targeting the signal
network that activates STAT3 may be an effective therapeutic
strategy for the treatment of AD (275).
Janus kinase 2 (JAK2) is a non-receptor protein
tyrosine kinase that plays a critical role in the JAK2/STAT3
signaling pathway in the CNS (283). Activation of the JAK2/STAT3
pathway leads to the transcription and expression of inflammatory
genes, resulting in an excessive accumulation of inflammatory
mediators and subsequent inflammation (284). Therefore, inhibition of the
JAK2/STAT3 pathway may be a potential therapeutic approach for
neuroinflammatory injury. A promising compound, protosappanin A
(PTA), which is a major bioactive component isolated from
Caesalpinia sappan L., was found to regulate LPS-induced
neuroinflammation by inhibiting the JAK2/STAT3 pathway (285). In the LPS-induced BV2 cell
model, PTA treatment reduced the production of TNF-α, IL-1β and NO
in microglia, while also dose-dependently decreasing IL-6 and IL-1β
mRNA expression (285). A
further study demonstrated that PTA inhibited JAK2/STAT3-dependent
inflammatory pathways by downregulating JAK2 and STAT3
phosphorylation as well as STAT3 nuclear translocation (285). Furthermore, the Porro et
al (286) found that
curcumin, a pigment isolated from Curcuma longa (turmeric)
with anti-inflammatory, antioxidant, and anticancer activities
(287), regulates
neuroinflammation by inducing an anti-inflammatory response against
microglia through the JAK2/STAT/SOCS (suppressor of cytokine
signaling) signaling pathway. Curcumin treatment increased the
production of the anti-inflammatory cytokines IL-4 and IL-10,
upregulated the expression of the cytokine signaling suppressor
SOCS-1, blocked JAK2 and STAT3 phosphorylation and reduced the
M1/M2 ratio of microglia phenotype in the same LPS-induced BV2 cell
model, thereby ameliorating neuroinflammation from multiple
perspectives (286).
NF-κB and STAT3 are two key regulators of cytokine
production that can reciprocally modulate each other (288,289). Inhibition of STAT3 activation
has been shown to reduce NF-κB activation, thereby attenuating
amyloidogenesis and neuroinflammation (271,272). Ent-Sauchinone, a polyphenolic
compound from the lignan family, exerts inhibitory effects on
neuroinflammation and amyloidogenesis by blocking the STAT3/NF-κB
pathway (271). In
LPS-stimulated BV-2 microglia, ent-Sauchinone dose-dependently
reduces the production of ROS and NO, as well as the expression of
iNOS and COX-2, while inhibiting NF-κB activation and the elevated
DNA-binding activity of STAT3 induced by LPS. Inhibition of
neuroinflammation and prevention of neuroinflammation-induced Aβ
production were further confirmed using short interfering RNA and
pharmacological inhibitors of STAT3 (290). Sorafenib (291), an anti-cancer drug, also exerts
anti-neuroinflammatory effects by modulating the AKT/P38-linked
STAT3/NF-κB signaling pathway. It reduces the mRNA expression of
pro-inflammatory cytokines in LPS-induced BV-2 microglia and
inhibits the increase in STAT3 and NF-κB phosphorylation levels by
inhibiting AKT and P38 signaling. An in vivo study further
confirmed the anti-inflammatory effects of Sorafenib, suggesting
its potential as a therapeutic agent to inhibit neuroinflammatory
responses in the brain (291)
(Fig. 4 and Table IV).
The PI3K/Akt pathway is a vital signaling pathway
that regulates a variety of transcription factors and cellular
functions (292). Its
association with various pathogenic factors of AD, including aging,
Aβ and synaptic loss, has been uncovered (293). There are reports of reduced
expression of the PI3K/Akt pathway in the brains of patients with
AD, while upregulation of this pathway can alleviate tau-induced
neurotoxicity and Aβ deposition (294), improve learning and memory
capacity and reduce brain damage, and reduce inflammation and
oxidative stress in mice with AD (295). Therefore, the role of the
PI3K/Akt pathway in microglia has received increasing attention.
Studies indicate that PI3K/Akt phosphorylation directly regulates
NF-κB in microglia, suggesting a strong link between PI3K/Akt and
neuroinflammation (296). GSK-3β
signaling, which is involved in inflammation, oxidative stress, and
apoptosis, can be activated by Akt phosphorylation upstream
(297). Therefore, the
Akt/GSK-3β signaling pathway, an important mediator of the
inflammatory response, is closely linked to the PI3K/Akt pathway
and the role of microglia.
The cyclic AMP response element binding (CREB) is a
stimulus-inducible transcription factor that dimers with the
conserved cyclic AMP response element (CRE) (302) to activate CRE-responsive genes
in response to extracellular stimuli (303). In the CNS, CREB regulates
various protein kinases, including protein kinase A (PKA) and
MAPKs, which are involved in neuronal development, synaptic
plasticity, short-term to long-term memory conversion and
neuroprotection in the CNS (304,305). Furthermore, dysregulated CREB
phosphorylation has been identified in AD mouse models (306) and patients with AD (307), demonstrating the important role
of CREB in the pathogenesis of AD (308). Notably, CREB has been found to
be associated with neuroinflammation and may be an effective
therapeutic target for the treatment of AD (309). Phosphorylation of CREB has been
shown to reduce neuroinflammation by regulating NF-κB to block the
transcription of inflammatory mediators (310). Moreover, phosphorylation of CREB
promotes the production of anti-inflammatory cytokines in activated
microglia that induce microglia inactivation or polarization to the
M2 phenotype (311), thus
modulating neuroinflammation for neuroprotection in AD.
The cGMP-dependent protein kinase (PKG) plays an
important role in mediating the transcriptional regulation of CREB
by phosphorylating CREB and activating different downstream genes
(312). In an aged Tg APP/PS1
mouse model, sildenafil was found to be effective in reducing
neuroinflammation and Aβ levels in the brain. Specifically,
sildenafil suppressed Aβ-induced pro-inflammatory factors in the
hippocampus, and this effect was mediated through the PKG/CREB
signaling pathway (313).
Inhibition of PKG in the hippocampus prior to sildenafil injection
resulted in blocked CREB phosphorylation, resulting in a reduced
production of inflammatory factors and ultimately produced
anti-inflammatory effects. Furthermore, there is ample evidence in
the literature to support the crucial role of the PKA/CREB pathway
as a drug target in AD (314),
particularly in the context of downregulation of the
transcriptional cascade that contributes to the disease.
Additionally, inhibition of histamine H3 receptors (H3R) has been
shown to improve cognitive deficits in AD (315,316). The histamine H3R antagonist,
thioperamide (317), can
effectively inhibit inflammatory cell recruitment (318), further highlighting the
importance of the PKA/CREB pathway in modulating neuroinflammation
as a therapeutic target for AD. It has been found that thioperamide
exerts its effects on the PKA/CREB signaling pathway, suppressing
microglia activity and promoting their conversion from M1 to M2
phenotype, ultimately impeding LPS-induced neuroinflammation and
restoring cognitive function in mice (317). Mechanistically, the downstream
PKA/CREB pathway activated by H2R stimulation triggers CBP
(CREB-CREB binding protein) interactions that facilitated the
release of anti-inflammatory factors and brain-derived neurotrophic
factor, while simultaneously attenuating NF-κB-CBP interactions to
reduce the secretion of pro-inflammatory factors. These effects
were found to be reversible by cimetidine (H2R antagonist) but not
by piramine (H1R antagonist), indicating a novel H2R-dependent
histamine-mediated mechanism underlying the therapeutic effects of
thioperamide on neuroinflammation (317) (Table IV).
Nuclear factor erythroid 2-related factor 2 (Nrf2)
is a transcription factor that plays a crucial role in regulating
oxidative stress in various cell types, including glial cells and
neurons (319). Notably, a
reduction in Nrf2 expression has been detected in the brains of
patients with AD (320).
Moreover, a growing body of research indicates that augmenting Nrf2
signaling has the potential to improve Aβ-induced neurodegeneration
and oxidative stress in in vitro and in vivo models
of AD (321). Such
investigations have also revealed that enhancing Nrf2 signaling can
alleviate microglia-mediated inflammation in the brain (322), highlighting the potential for
therapeutic intervention targeting Nrf2 in the development of drugs
for the treatment of AD.
Studies have shown that certain herbs and natural
products contain active ingredients that can interfere with Nrf2,
thereby inhibiting neuroinflammation. For example, Methysticin
(323), a kavalactones derived
from the Piperaceae plant kava (324), has been demonstrated to inhibit
neuroinflammation and oxidative damage and to attenuate long-term
memory loss in APP/PS1 mice. These effects are attributed to its
ability to significantly reduce microglia activation and the
secretion of pro-inflammatory factors in the hippocampus and
cortex, possibly mediated by Nrf2. Similarly, a polyphenol extract
derived from Arabidopsis thaliana was found to have
anti-inflammatory activity in transgenic AD flies and
Aβ25-35-induced BV2 cells by influencing the nuclear
translocation of Nrf2 and NF-κB (325). Gracilin A, a natural product
isolated from the marine sponge Spongionella gracilis
(326), has been associated with
Nrf2-involved inflammation. An in vitro study has shown that
Gracilin A reduces the release of pro-inflammatory factors from BV2
cells induced by LPS by inhibiting the expression of iNOS and the
activation of p38 MAPK, which affects the translocation of NF-κB
p65 and Nrf2 (327).
Kelch-like ECH-associated protein 1 (Keap1), as an
adapter protein, inhibits the function of Nrf2 by degrading it in
the normal state of the cell (328). However, when cells are exposed
to external stimuli, the degradation of Keap1 (dependent on Nrf2),
is inhibited, leading to the accumulation of Nrf2 in the nucleus
and its regulatory role in the expression of various antioxidant
genes (329). Engeletin, a
flavonol glycoside derived from the leaves of Engelhardia
roxburghiana (330), has
demonstrated anti-inflammatory properties. Specifically, it has
been shown to inhibit the expression and secretion of
Aβ1-42-induced pro-inflammatory factors and to enhance
the activation of the Keap1/Nrf2 pathway in BV-2 cells. However,
when Nrf2 was knocked down, the inhibitory effect of Engeletin was
reversed. These findings further underscore the potential of
pharmacological intervention targeting the Keap1/Nrf2 pathway in
anti-AD therapy (331).
Haem oxygenase-1 (HO-1), a stress-inducible
protein, exerts a protective effect against inflammatory and
oxidative stress and has been shown to be beneficial in
neurodegenerative diseases including AD (332). The promotion of HO-1 expression
is mediated by Nrf2 (333).
Study has shown that Bambusae Caulis in Taeniam ethyl acetate
fraction (BCE) (334) as a
modulator of Nrf2 signaling, regulates the neuroprotective and
anti-neuroinflammatory effects of microglia BV2 by modulating the
expression of HO-1. BCE was shown to inhibit the production of
pro-inflammatory mediators and cytokines in LPS-induced BV2 cells,
while upregulating the mRNA and protein expression levels of HO-1,
and influencing the accumulation and transactivation of Nrf2 in the
cells (334). Further evidence
for the involvement of HO-1 in the observed anti-inflammatory
effects of BCE was obtained by using the selective HO-1 inhibitor,
SnPP, which reversed these effects (334).
Researchers have identified multiple inhibitors
that act on multiple pathways to inhibit neuroinflammation by
targeting Nrf2. Among these inhibitors, L-F001 (335), a newly developed ROCK inhibitor,
has shown promise in the treatment of AD by inhibiting NF-κB and
activating Nrf2. An in vitro study has demonstrated that
L-F001 significantly inhibits the expression of iNOS and COX-2 as
well as the secretion of pro-inflammatory mediators in BV-2 cells
following LPS induction (335).
This is accompanied by inhibition of NF-κB signaling and
upregulation the expression of HO-1 and glutamate cysteine ligase
modifier subunit, downstream effectors of Nrf2 (335). Similarly in vivo
experiments, on mice have confirmed that L-F001 significantly
reduces the levels of pro-inflammatory mediators induced by LPS, in
line with the in vitro findings (335). In addition, the researchers have
found that G protein-coupled receptor 17 was expressed in neurons
and microglia (336) and that
its antagonist, cangrelor (337), had an inhibitory effect on
neuroinflammation. In a mouse model of AD with
intracerebroventricular injection of Aβ1-42, cangrelor
reduced BACE1 activity as well as Aβ1-42 levels in the
hippocampus and frontal cortex of mice, while inhibiting microglia
activation and levels of pro-inflammatory factors through a
mechanism involving Nrf2/HO-1 and NF-κB signaling (337). Another inhibitor,
β-naphthoflavone (BNF) (338), a
derivative of a natural flavonoid widely used in the pharmaceutical
industry, has antioxidant and anti-inflammatory effects.
Pretreatment with BNF was found to inhibit activation of the NF-κB
pathway in LPS-treated BV-2 cells, promote AKT activation, enhance
the nuclear translocation of Nrf2, lead to an upregulation of the
HO-1 protein levels, and significantly reduce the expression of
pro-inflammatory mediators (338). The use of MK2206 (an AKT
inhibitor), RA (an Nrf2 inhibitor) and SnPP IX (an HO-1 inhibitor)
further confirms that BNF inhibits the production of
pro-inflammatory mediators by activating this pathway (338).
The pathogenesis of AD is a multifaceted process,
but studies ranging from cellular and animal models, as well as
studies involving patients with AD, have unequivocally established
the pivotal role of neuroinflammation. Excessive activation of
microglia releases inflammatory mediators that contribute to the
pathological features of AD. Thus, inhibiting microglia-mediated
inflammation is a promising approach to combat this disease.
Intracellular signaling pathways play a crucial
role in maintaining cellular function and metabolism and are
intricately associated with the pathogenesis of AD, including
neuroinflammation. It is worth noting that these signaling pathways
are complex, interconnected and capable of interacting with each
other. By intervening in the pertinent signaling pathways through
the use of drugs or inhibitors, it is possible to inhibit
neuroinflammation and exert an effect on AD. Therefore, this review
focused on neuroinflammation in AD and presented a comprehensive
synthesis and summary of the mechanisms of action and potential
signaling proteins linked with inhibitors, herbal medicines, and
their active ingredients and metabolites, from the standpoint of
signaling pathways (Fig. 5).
Various drugs or inhibitors can regulate various
signaling pathways, and multiple drugs can also target the same
pathway. It is worth noting that NF-κB, MAPK, and NLRP3 are key
signaling molecules targeted in neuroinflammation and have been
extensively studied in drug discovery. By interfering with one or
more of these signaling pathways, drugs can synergistically
modulate multiple targets, achieve a balance between antioxidant
and pro-inflammatory effects, and ultimately improve cognitive
impairment in patients with AD (refer to Fig. 1 for details). Given the complexity
of AD pathogenesis, drugs or inhibitors with multi-level and
multi-target potential hold promise as a breakthrough in AD drug
development. Exploring the anti-inflammatory effects of commonly
used clinical drugs may broaden their potential application.
However, current research is still primarily focused on animal and
cellular experiments, with a focus on LPSor Aβ-stimulated BV2
microglia. Although a number of problems have prevented a number of
drugs from entering clinical trials, inhibitors, that target
neuroinflammation remain a potentially promising therapeutic option
for AD.
In summary, the present review highlighted the
prominent role of neuroinflammation in AD pathology and reviewed
various anti-inflammatory inhibitors targeting molecular targets
and signaling pathways. These inhibitors have shown significant
potential as drug treatments for AD and have provided a foundation
for the further development of novel AD therapeutics. However, the
specificity, efficacy, safety, and availability of these
inhibitors, natural ingredients and metabolites, are critical
considerations for their clinical application. Moreover, further
studies on their pharmacokinetic profiles and underlying mechanisms
are necessary for the development of novel AD therapeutics. Despite
these challenges, the potential benefits of these drugs underscore
the need for continued research into their efficacy as treatments
for AD.
Data sharing is not applicable to this article, as
no data sets were generated or analyzed during the current
study.
YZ wrote the original draft of the manuscript. ZW,
RZ and XZ reviewed and edited the manuscript. QG, JG and PX
produced the diagrams and charts. XJ and LY contributed to the
conception, design and drafting of the manuscript. All authors read
and approved the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
Not applicable.
The present study was supported by the National Natural Science
Foundation of China (grant nos. 82174470 and 82274318) and Natural
Science Foundation of Tianjin City, China (grant no.
21JCQNJC01170).
|
1
|
Wang X, Iyaswamy A, Xu D, Krishnamoorthi
S, Sreenivasmurthy SG, Yang Y, Li Y, Chen C, Li M, Li HW and Wong
MS: Real-time detection and visualization of amyloid-β aggregates
induced by hydrogen peroxide in cell and mouse models of
Alzheimer's disease. ACS Appl Mater Interfaces. 15:39–47. 2023.
View Article : Google Scholar
|
|
2
|
Shih YH, Tu LH, Chang TY, Ganesan K, Chang
WW, Chang PS, Fang YS, Lin YT, Jin LW and Chen YR: TDP-43 interacts
with amyloid-β, inhibits fibrillization, and worsens pathology in a
model of Alzheimer's disease. Nat Commun. 11:59502020. View Article : Google Scholar
|
|
3
|
Nasaruddin ML, Pan X, McGuinness B,
Passmore P, Kehoe PG, Holscher C, Graham SF and Green BD: Evidence
that parietal lobe fatty acids may be more profoundly affected in
moderate Alzheimer's disease (AD) pathology than in severe AD
pathology. Metabolites. 8:692018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen YG: Research progress in the
pathogenesis of Alzheimer's disease. Chin Med J (Engl).
131:1618–1624. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ardura-Fabregat A, Boddeke EWGM,
Boza-Serrano A, Brioschi S, Castro-Gomez S, Ceyzériat K, Dansokho
C, Dierkes T, Gelders G, Heneka MT, et al: Targeting
neuroinflammation to treat Alzheimer's disease. CNS Drugs.
31:1057–1082. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Finneran DJ and Nash KR: Neuroinflammation
and fractalkine signaling in Alzheimer's disease. J
Neuroinflammation. 16:302019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Souza LC, Filho CB, Goes AT, Fabbro LD, de
Gomes MG, Savegnago L, Oliveira MS and Jesse CR: Neuroprotective
effect of physical exercise in a mouse model of Alzheimer's disease
induced by β-amyloid 2013. 1-40 peptide. Neurotox Res. 24:148–163.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Janelsins MC, Mastrangelo MA, Oddo S,
LaFerla FM, Federoff HJ and Bowers WJ: Early correlation of
microglial activation with enhanced tumor necrosis factor-alpha and
monocyte chemoattractant protein-1 expression specifically within
the entorhinal cortex of triple transgenic Alzheimer's disease
mice. J Neuroinflammation. 2:232005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ma K, Mount HTJ and McLaurin J:
Region-specific distribution of β-amyloid peptide and cytokine
expression in TgCRND8 mice. Neurosci Lett. 492:5–10. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wu YY, Hsu JL, Wang HC, Wu SJ, Hong CJ and
Cheng IHJ: Alterations of the neuroinflammatory markers IL-6 and
TRAIL in Alzheimer's disease. Dement Geriatr Cogn Dis Extra.
5:424–434. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Takeda S, Sato N, Ikimura K, Nishino H,
Rakugi H and Morishita R: Increased blood-brain barrier
vulnerability to systemic inflammation in an Alzheimer disease
mouse model. Neurobiol Aging. 34:2064–2070. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mann DM, Tucker CM and Yates PO:
Alzheimer's disease: An olfactory connection? Mech Ageing Dev.
42:1–15. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Christen-Zaech S, Kraftsik R, Pillevuit O,
Kiraly M, Martins R, Khalili K and Miklossy J: Early olfactory
involvement in Alzheimer's disease. Can J Neurol Sci. 30:20–25.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Abe Y, Ikegawa N, Yoshida K, Muramatsu K,
Hattori S, Kawai K, Murakami M, Tanaka T, Goda W, Goto M, et al:
Behavioral and electrophysiological evidence for a neuroprotective
role of aquaporin-4 in the 5xFAD transgenic mice model. Acta
Neuropathol Commun. 8:672020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Walker DG, Dalsing-Hernandez JE, Campbell
NA and Lue LF: Decreased expression of CD200 and CD200 receptor in
Alzheimer's disease: A potential mechanism leading to chronic
inflammation. Exp Neurol. 215:5–19. 2009. View Article : Google Scholar :
|
|
16
|
Ham HJ, Lee YS, Yun J, Son DJ, Lee HP, Han
SB and Hong JT: K284-6111 alleviates memory impairment and
neuroinflammation in Tg2576 mice by inhibition of chitinase-3-like
1 regulating ERK-dependent PTX3 pathway. J Neuroinflammation.
17:3502020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cullen NC, Malarstig AN, Stomrud E,
Hansson O and Mattsson-Carlgren N: Accelerated inflammatory aging
in Alzheimer's disease and its relation to amyloid, tau, and
cognition. Sci Rep. 11:19652021. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Voet S, Mc Guire C, Hagemeyer N, Martens
A, Schroeder A, Wieghofer P, Daems C, Staszewski O, Vande Walle L,
Jordao MJC, et al: A20 critically controls microglia activation and
inhibits inflammasome-dependent neuroinflammation. Nat Commun.
9:20362018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Erny D, Hrabě de Angelis AL, Jaitin D,
Wieghofer P, Staszewski O, David E, Keren-Shaul H, Mahlakoiv T,
Jakobshagen K, Buch T, et al: Host microbiota constantly control
maturation and function of microglia in the CNS. Nat Neurosci.
18:965–977. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sierra A, Beccari S, Diaz-Aparicio I,
Encinas JM, Comeau S and Tremblay MÈ: Surveillance, phagocytosis,
and inflammation: How never-resting microglia influence adult
hippocampal neurogenesis. Neural Plast. 2014:6103432014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ben-Yehuda H, Matcovitch-Natan O, Kertser
A, Spinrad A, Prinz M, Amit I and Schwartz M: Maternal Type-I
interferon signaling adversely affects the microglia and the
behavior of the offspring accompanied by increased sensitivity to
stress. Mol Psychiatr. 25:1050–1067. 2020. View Article : Google Scholar
|
|
22
|
Town T, Nikolic V and Tan J: The
microglial 'activation' continuum: From innate to adaptive
responses. J Neuroinflammation. 2:242005. View Article : Google Scholar
|
|
23
|
Leng F and Edison P: Neuroinflammation and
microglial activation in Alzheimer disease: Where do we go from
here? Nat Rev Neurol. 17:157–172. 2021. View Article : Google Scholar
|
|
24
|
Song WM, Joshita S, Zhou Y, Ulland TK,
Gilfillan S and Colonna M: Humanized TREM2 mice reveal
microglia-intrinsic and -extrinsic effects of R47H polymorphism. J
Exp Med. 215:745–760. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tansey MG, McCoy MK and Frank-Cannon TC:
Neuroinflammatory mechanisms in Parkinson's disease: Potential
environmental triggers, pathways, and targets for early therapeutic
intervention. Exp Neurol. 208:1–25. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jankowsky JL and Patterson PH: Cytokine
and growth factor involvement in long-term potentiation. Mol Cell
Neurosci. 14:273–286. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Das S and Basu A: Inflammation: A new
candidate in modulating adult neurogenesis. J Neurosci Res.
86:1199–1208. 2008. View Article : Google Scholar
|
|
28
|
Heneka MT, Carson MJ, El KJ, Landreth GE,
Brosseron F, Feinstein DL, Jacobs AH, Wyss-Coray T, Vitorica J,
Ransohoff RM, et al: Neuroinflammation in Alzheimer's disease.
Lancet Neurol. 14:388–405. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Giridharan S and Srinivasan M: Mechanisms
of NF-κB p65 and strategies for therapeutic manipulation. J Inflamm
Res. 11:407–419. 2018. View Article : Google Scholar :
|
|
30
|
Noort AR, Tak PP and Tas SW: Non-canonical
NF-κB signaling in rheumatoid arthritis: Dr Jekyll and Mr Hyde?
Arthritis Res Ther. 17:152015. View Article : Google Scholar
|
|
31
|
Mattson MP and Meffert MK: Roles for
NF-kappaB in nerve cell survival, plasticity, and disease. Cell
Death Differ. 13:852–860. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Miyauchi T, Uchida Y, Kadono K, Hirao H,
Kawasoe J, Watanabe T, Ueda S, Okajima H, Terajima H and Uemoto S:
Up-regulation of FOXO1 and reduced inflammation by β-hydroxybutyric
acid are essential diet restriction benefits against liver injury.
Proc Natl Acad Sci USA. 116:13533–13542. 2019. View Article : Google Scholar
|
|
33
|
Liu Y, Li D, Jiang Q, Zhang Q, Liu P, Wang
L, Zong M, Zhang Q, Li H, An Y, et al: (3R,
7R)-7-Acetoxyl-9-Oxo-de-O-methyllasiodiplodin, a secondary
metabolite of penicillium Sp., inhibits LPS-mediated inflammation
in RAW 264.7 macrophages through blocking ERK/MAPKs and NF-κB
signaling pathways. Inflammation. 42:1463–1473. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Christian F, Smith EL and Carmody RJ: The
regulation of NF-κB subunits by phosphorylation. Cells. 5:122016.
View Article : Google Scholar
|
|
35
|
Oeckinghaus A and Ghosh S: The NF-kappaB
family of transcription factors and its regulation. Cold Spring
Harb Perspect Biol. 1:a0000342009. View Article : Google Scholar
|
|
36
|
Yano H, Sakai M, Matsukawa T, Yagi T,
Naganuma T, Mitsushima M, Iida S, Inaba Y, Inoue H, Unoki-Kubota H,
et al: PHD3 regulates glucose metabolism by suppressing
stress-induced signalling and optimising gluconeogenesis and
insulin signalling in hepatocytes. Sci Rep. 8:142902018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Terai K, Matsuo A and McGeer PL:
Enhancement of immunoreactivity for NF-kappa B in the hippocampal
formation and cerebral cortex of Alzheimer's disease. Brain Res.
735:159–168. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kotilinek LA, Westerman MA, Wang Q,
Panizzon K, Lim GP, Simonyi A, Lesne S, Falinska A, Younkin LH,
Younkin SG, et al: Cyclooxygenase-2 inhibition improves
amyloid-beta-mediated suppression of memory and synaptic
plasticity. Brain. 131:651–664. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kolesnick R and Golde DW: The
sphingomyelin pathway in tumor necrosis factor and interleukin-1
signaling. Cell. 77:325–328. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kitamura Y, Shimohama S, Ota T, Matsuoka
Y, Nomura Y and Taniguchi T: Alteration of transcription factors
NF-kappaB and STAT1 in Alzheimer's disease brains. Neurosci Lett.
237:17–20. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chen CH, Zhou W, Liu S, Deng Y, Cai F,
Tone M, Tone Y, Tong Y and Song W: Increased NF-κB signalling
up-regulates BACE1 expression and its therapeutic potential in
Alzheimer's disease. Int J Neuropsychopharmacol. 15:77–90. 2012.
View Article : Google Scholar
|
|
42
|
Snow WM and Albensi BC: Neuronal gene
targets of NF-κB and their dysregulation in Alzheimer's disease.
Front Mol Neurosci. 9:1182016. View Article : Google Scholar
|
|
43
|
Wang C, Fan L, Khawaja RR, Liu B, Zhan L,
Kodama L, Chin M, Li Y, Le D, Zhou Y, et al: Microglial NF-κB
drives tau spreading and toxicity in a mouse model of tauopathy.
Nat Commun. 13:19692022. View Article : Google Scholar
|
|
44
|
López N, Tormo C, De Blas I, Llinares I
and Alom J: Oxidative stress in Alzheimer's disease and mild
cognitive impairment with high sensitivity and specificity. J
Alzheimers Dis. 33:823–829. 2013. View Article : Google Scholar
|
|
45
|
Wang SW, Yang SG, Liu W, Zhang YX, Xu PX,
Wang T, Ling TJ and Liu RT: Alpha-tocopherol quinine ameliorates
spatial memory deficits by reducing beta-amyloid oligomers,
neuroinflammation and oxidative stress in transgenic mice with
Alzheimer's disease. Behav Brain Res. 296:109–117. 2016. View Article : Google Scholar
|
|
46
|
Sun HJ, Xue DD, Lu BZ, Li Y, Sheng LX, Zhu
Z, Zhou YW, Zhang JX, Lin GJ, Lin SZ, et al: A novel synthetic
steroid of 2β,3α,5α-trihydroxy-androst-6-one alleviates the loss of
rat retinal ganglion cells caused by acute intraocular hypertension
via inhibiting the inflammatory activation of microglia. Molecules.
24:2522019. View Article : Google Scholar
|
|
47
|
Belkhelfa M, Rafa H, Medjeber O,
Arroul-Lammali A, Behairi N, Abada-Bendib M, Makrelouf M, Belarbi
S, Masmoudi AN, Tazir M and Touil-Boukoffa C: IFN-γ and TNF-α are
involved during Alzheimer disease progression and correlate with
nitric oxide production: a study in Algerian patients. J Interferon
Cytokine Res. 34:839–847. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sui Y, Stehno-Bittel L, Li S, Loganathan
R, Dhillon NK, Pinson D, Nath A, Kolson D, Narayan O and Buch S:
CXCL10-induced cell death in neurons: Role of calcium
dysregulation. Eur J Neurosci. 23:957–964. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Xu PX, Wang SW, Yu XL, Su YJ, Wang T, Zhou
WW, Zhang H, Wang YJ and Liu RT: Rutin improves spatial memory in
Alzheimer's disease transgenic mice by reducing Aβ oligomer level
and attenuating oxidative stress and neuroinflammation. Behav Brain
Res. 264:173–180. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Budzynska B, Faggio C, Kruk-Slomka M,
Samec D, Nabavi SF, Sureda A, Devi KP and Nabavi SM: Rutin as
neuroprotective agent: From bench to bedside. Curr Med Chem.
26:5152–5164. 2019. View Article : Google Scholar
|
|
51
|
Sun XY, Li LJ, Dong QX, Zhu J, Huang YR,
Hou SJ, Yu XL and Liu RT: Rutin prevents tau pathology and
neuroinflammation in a mouse model of Alzheimer's disease. J
Neuroinflammation. 18:1312021. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Bernard CB, Krishanmurty HG, Chauret D,
Durst T, Philogène BJ, Sánchez-Vindas P, Hasbun C, Poveda L, San
Román L and Arnason JT: Insecticidal defenses of piperaceae from
the neotropics. J Chem Ecol. 21:801–814. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Gu SM, Lee HP, Ham YW, Son DJ, Kim HY, Oh
KW, Han SB, Yun J and Hong JT: Piperlongumine improves
lipopolysaccharide-induced amyloidogenesis by suppressing NF-KappaB
pathway. Neuromolecular Med. 20:312–327. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Son DJ, Lee JW, Lee YH, Song HS, Lee CK
and Hong JT: Therapeutic application of anti-arthritis,
pain-releasing, and anti-cancer effects of bee venom and its
constituent compounds. Pharmacol Ther. 115:246–270. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Gu SM, Park MH, Hwang CJ, Song HS, Lee US,
Han SB, Oh KW, Ham YW, Song MJ, Son DJ and Hong JT: Bee venom
ameliorates lipopolysaccharide-induced memory loss by preventing
NF-kappaB pathway. J Neuroinflammation. 12:1242015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Lin CC, Hsu YF and Lin TC: Effects of
punicalagin and punicalin on carrageenan-induced inflammation in
rats. Am J Chin Med. 27:371–376. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Kim YE, Hwang CJ, Lee HP, Kim CS, Son DJ,
Ham YW, Hellström M, Han SB, Kim HS, Park EK and Hong JT:
Inhibitory effect of punicalagin on lipopolysaccharide-induced
neuroinflammation, oxidative stress and memory impairment via
inhibition of nuclear factor-kappaB. Neuropharmacology. 117:21–32.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Chen S and Jia J: Tenuifolin attenuates
amyloid-β42-induced neuroinflammation in microglia through the
NF-κB signaling pathway. J Alzheimers Dis. 76:195–205. 2020.
View Article : Google Scholar
|
|
59
|
Hammad AS, Ravindran S, Khalil A and
Munusamy S: Structure-activity relationship of piperine and its
synthetic amide analogs for therapeutic potential to prevent
experimentally induced ER stress in vitro. Cell Stress Chaperones.
22:417–428. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Shahbazi S, Zakerali T, Frycz B and Kaur
J: Impact of novel N-aryl substituted piperamide on NF-kappa B
translocation as a potent anti-neuroinflammatory agent. Biomed
Pharmacother. 127:1101992020. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Shahbazi S, Zakerali T, Frycz BA and Kaur
J: The critical role of piperamide derivative D4 in the regulation
of inflammatory response by the microglia and astrocytic glial
cells. Biomed Pharmacother. 132:1108952020. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Hsu YL, Kuo PL, Chiang LC and Lin CC:
Involvement of p53, nuclear factor kappaB and Fas/Fas ligand in
induction of apoptosis and cell cycle arrest by saikosaponin d in
human hepatoma cell lines. Cancer Lett. 213:213–221. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Park WH, Kang S, Piao Y, Pak CJ, Oh MS,
Kim J, Kang MS and Pak YK: Ethanol extract of Bupleurum falcatum
and saikosaponins inhibit neuroinflammation via inhibition of
NF-κB. J Ethnopharmacol. 174:37–44. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Leláková V, Béraud-Dufour S, Hošek J,
Šmejkal K, Prachyawarakorn V, Pailee P, Widmann C, Václavík J,
Coppola T, Mazella J, et al: Therapeutic potential of prenylated
stilbenoid macasiamenene F through its anti-inflammatory and
cytoprotective effects on LPS-challenged monocytes and microglia. J
Ethnopharmacol. 263:1131472020. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Najm FJ, Madhavan M, Zaremba A, Shick E,
Karl RT, Factor DC, Miller TE, Nevin ZS, Kantor C, Sargent A, et
al: Drug-based modulation of endogenous stem cells promotes
functional remyelination in vivo. Nature. 522:216–220. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Yeo IJ, Yun J, Son DJ, Han SB and Hong JT:
Antifungal drug miconazole ameliorated memory deficits in a mouse
model of LPS-induced memory loss through targeting iNOS. Cell Death
Dis. 11:6232020. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Solberg NO, Chamberlin R, Vigil JR, Deck
LM, Heidrich JE, Brown DC, Brady CI, Vander Jagt TA, Garwood M,
Bisoffi M, et al: Optical and SPION-enhanced MR imaging shows that
trans-stilbene inhibitors of NF-κB concomitantly lower Alzheimer's
disease plaque formation and microglial activation in AβPP/PS-1
transgenic mouse brain. J Alzheimers Dis. 40:191–212. 2014.
View Article : Google Scholar
|
|
68
|
Lindsay A, Hickman D and Srinivasan M: A
nuclear factor-kappa B inhibiting peptide suppresses innate immune
receptors and gliosis in a transgenic mouse model of Alzheimer's
disease. Biomed Pharmacother. 138:1114052021. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Choi JY, Yeo IJ, Kim KC, Choi WR, Jung JK,
Han SB and Hong JT: K284-6111 prevents the amyloid beta-induced
neuroinflammation and impairment of recognition memory through
inhibition of NF-κB-mediated CHI3L1 expression. J
Neuroinflammation. 15:2242018. View Article : Google Scholar
|
|
70
|
Lian W, Jia H, Xu L, Zhou W, Kang D, Liu A
and Du G: Multi-protection of DL0410 in ameliorating cognitive
defects in D-galactose induced aging mice. Front Aging Neurosci.
9:4092017. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Heynekamp JJ, Weber WM, Hunsaker LA,
Gonzales AM, Orlando RA, Deck LM and Jagt DL: Substituted
trans-stilbenes, including analogues of the natural product
resveratrol, inhibit the human tumor necrosis factor alpha-induced
activation of transcription factor nuclear factor kappaB. J Med
Chem. 49:7182–7189. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Ayroldi E and Riccardi C:
Glucocorticoid-induced leucine zipper (GILZ): A new important
mediator of glucocorticoid action. FASEB J. 23:3649–3658. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Cannarile L, Zollo O, D'Adamio F, Ayroldi
E, Marchetti C, Tabilio A, Bruscoli S and Riccardi C: Cloning,
chromosomal assignment and tissue distribution of human GILZ, a
glucocorticoid hormone-induced gene. Cell Death Differ. 8:201–203.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Bonneh-Barkay D, Wang G, Starkey A,
Hamilton RL and Wiley CA: In vivo CHI3L1 (YKL-40) expression in
astrocytes in acute and chronic neurological diseases. J
Neuroinflammation. 7:342010. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Muszyński P, Groblewska M,
Kulczyńska-Przybik A, Kułakowska A and Mroczko B: YKL-40 as a
potential biomarker and a possible target in therapeutic strategies
of Alzheimer's disease. Curr Neuropharmacol. 15:906–917. 2017.
View Article : Google Scholar
|
|
76
|
Di Rosa M and Malaguarnera L: Chitinase 3
like-1: An emerging molecule involved in diabetes and diabetic
complications. Pathobiology. 83:228–242. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Billod JM, Lacetera A, Guzmán-Caldentey J
and Martín-Santamaría S: Computational approaches to Toll-like
receptor 4 modulation. Molecules. 21:9942016. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Mertowski S, Grywalska E, Gosik K,
Smarz-Widelska I, Hymos A, Dworacki G, Niedźwiedzka-Rystwej P, Drop
B, Roliński J and Załuska W: TLR2 expression on select lymphocyte
subsets as a new marker in glomerulonephritis. J Clin Med.
9:5412020. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Elmaidomy AH, Alhadrami HA, Amin E, Aly
HF, Othman AM, Rateb ME, Hetta MH, Abdelmohsen UR and M Hassan H:
Anti-inflammatory and antioxidant activities of terpene- and
polyphenol-rich premna odorata leaves on alcohol-inflamed female
wistar albino rat liver. Molecules. 25:31162020. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Zhong X, Liu M, Yao W, Du K, He M, Jin X,
Jiao L, Ma G, Wei B and Wei M: Epigallocatechin-3-gallate
attenuates microglial inflammation and neurotoxicity by suppressing
the activation of canonical and noncanonical inflammasome via
TLR4/NF-κB pathway. Mol Nutr Food Res. 63:e18012302019. View Article : Google Scholar
|
|
81
|
Zhou X, Yuan L, Zhao X, Hou C, Ma W, Yu H
and Xiao R: Genistein antagonizes inflammatory damage induced by
β-amyloid peptide in microglia through TLR4 and NF-κB. Nutrition.
30:90–95. 2014. View Article : Google Scholar
|
|
82
|
Chen R, Wang Z, Zhi Z, Tian J, Zhao Y and
Sun J: Targeting the TLR4/NF-κB pathway in β-amyloid-stimulated
microglial cells: A possible mechanism that oxysophoridine exerts
anti-oxidative and anti-inflammatory effects in an in vitro model
of Alzheimer's disease. Brain Res Bull. 175:150–157. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Onishi S, Meguro S, Pervin M, Kitazawa H,
Yoto A, Ishino M, Shimba Y, Mochizuki Y, Miura S, Tokimitsu I and
Unno K: Green tea extracts attenuate brain dysfunction in
high-fat-diet-fed SAMP8 mice. Nutrients. 11:8212019. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Balázs A, Faisal Z, Csepregi R, Kőszegi T,
Kriszt B, Szabó I and Poór M: In Vitro evaluation of the individual
and combined cytotoxic and estrogenic effects of zearalenone, its
reduced metabolites, alternariol, and genistein. Int J Mol Sci.
22:62812021. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Wang R, Deng X, Gao Q, Wu X, Han L, Gao X,
Zhao S, Chen W, Zhou R, Li Z and Bai C: Sophora alopecuroides L.:
An ethnopharmacological, phytochemical, and pharmacological review.
J Ethnopharmacol. 248:1121722020. View Article : Google Scholar
|
|
86
|
Rui C, Yuxiang L, Ning J, Ningtian M,
Qingluan Z, Yinju H, Ru Z, Lin M, Tao S and Jianqiang Y:
Anti-apoptotic and neuroprotective effects of oxysophoridine on
cerebral ischemia both in vivo and in vitro. Planta Med.
79:916–923. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Wang Y, Wang Y, Jia S, Dong Q, Chen Y, Lu
S and Hou L: Effect of lipid-bound apolipoprotein A-I cysteine
mutant on ATF3 in RAW264.7 cells. Biosci Rep. 37:BSR201603982017.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Zeng KW, Zhao MB, Ma ZZ, Jiang Y and Tu
PF: Protosappanin A inhibits oxidative and nitrative stress via
interfering the interaction of transmembrane protein CD14 with
Toll-like receptor-4 in lipopolysaccharide-induced BV-2 microglia.
Int Immunopharmacol. 14:558–569. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Mo Y, Sun YY and Liu KY: Autophagy and
inflammation in ischemic stroke. Neural Regen Res. 15:1388–1396.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Capiralla H, Vingtdeux V, Zhao H,
Sankowski R, Al-Abed Y, Davies P and Marambaud P: Resveratrol
mitigates lipopolysaccharide- and Aβ-mediated microglial
inflammation by inhibiting the TLR4/NF-κB/STAT signaling cascade. J
Neurochem. 120:461–472. 2012. View Article : Google Scholar
|
|
91
|
Xu Y, Huang X, Huangfu B, Hu Y, Xu J, Gao
R, Huang K and He X: Sulforaphane ameliorates nonalcoholic fatty
liver disease induced by high-fat and high-fructose diet via
LPS/TLR4 in the gut-liver axis. Nutrients. 15:7432023. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Song J, Feng L, Zhong R, Xia Z, Zhang L,
Cui L, Yan H, Jia X and Zhang Z: Icariside II inhibits the EMT of
NSCLC cells in inflammatory microenvironment via down-regulation of
Akt/NF-κB signaling pathway. Mol Carcinogen. 56:36–48. 2017.
View Article : Google Scholar
|
|
93
|
Lee KS, Lee HJ, Ahn KS and Kim SH, Nam D,
Kim DK, Choi DY, Ahn KS, Lu J and Kim SH:
Cyclooxygenase-2/prostaglandin E2 pathway mediates icariside II
induced apoptosis in human PC-3 prostate cancer cells. Cancer Lett.
280:93–100. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Zhou J, Deng Y, Li F, Yin C, Shi J and
Gong Q: Icariside II attenuates lipopolysaccharide-induced
neuroinflammation through inhibiting TLR4/MyD88/NF-κB pathway in
rats. Biomed Pharmacother. 111:315–324. 2019. View Article : Google Scholar
|
|
95
|
Zhou D, Zhou W, Song JK, Feng ZY, Yang RY,
Wu S, Wang L, Liu AL and Du GH: DL0410, a novel dual cholinesterase
inhibitor, protects mouse brains against Aβ-induced neuronal damage
via the Akt/JNK signaling pathway. Acta Pharmacol Sin.
37:1401–1412. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Yang RY, Zhao G, Wang DM, Pang XC, Wang
SB, Fang JS, Li C, Liu AL, Wu S and Du GH: DL0410 can reverse
cognitive impairment, synaptic loss and reduce plaque load in
APP/PS1 transgenic mice. Pharmacol Biochem Behav. 139:15–26. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Zhang B, Lian W, Zhao J, Wang Z, Liu A and
Du G: DL0410 Alleviates memory impairment in D-galactose-induced
aging rats by suppressing neuroinflammation via the
TLR4/MyD88/NF-κB pathway. Oxid Med Cell Longev.
2021:65211462021.
|
|
98
|
Mao J, Huang S, Liu S, Feng XL, Yu M, Liu
J, Sun YE, Chen G, Yu Y, Zhao J and Pei G: A herbal medicine for
Alzheimer's disease and its active constituents promote neural
progenitor proliferation. Aging Cell. 14:784–796. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Geng Y, Li C, Liu J, Xing G, Zhou L, Dong
M, Li X and Niu Y: Beta-asarone improves cognitive function by
suppressing neuronal apoptosis in the beta-amyloid hippocampus
injection rats. Biol Pharm Bull. 33:836–843. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Zhong J, Qiu X, Yu Q, Chen H and Yan C: A
novel polysaccharide from Acorus tatarinowii protects against
LPS-induced neuroinflammation and neurotoxicity by inhibiting
TLR4-mediated MyD88/NF-κB and PI3K/Akt signaling pathways. Int J
Biol Macromol. 163:464–475. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Liu M, Guo H, Li Z, Zhang C, Zhang X, Cui
Q and Tian J: Molecular level insight into the benefit of myricetin
and dihydromyricetin uptake in patients with Alzheimer's diseases.
Front Aging Neurosci. 12:6016032020. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Jing N and Li X: Dihydromyricetin
attenuates inflammation through TLR4/NF-kappaB pathway. Open Med
(Wars). 14:719–725. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Gu RX, Gu H, Xie ZY, Wang JF, Arias HR,
Wei DQ and Chou KC: Possible drug candidates for Alzheimer's
disease deduced from studying their binding interactions with
alpha7 nicotinic acetylcholine receptor. Med Chem. 5:250–262. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Shi S, Liang D, Chen Y, Xie Y, Wang Y,
Wang L, Wang Z and Qiao Z: Gx-50 reduces β-amyloid-induced TNF-α,
IL-1β, NO, and PGE2 expression and inhibits NF-κB signaling in a
mouse model of Alzheimer's disease. Eur J Immunol. 46:665–676.
2016. View Article : Google Scholar
|
|
105
|
Zheng P, Huang C, Leng D, Sun B and Zhang
XD: Transcriptome analysis of peripheral whole blood identifies
crucial lncRNAs implicated in childhood asthma. Bmc Med Genomics.
13:1362020. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Zhong X, Feng L, Xu WH, Wu X, Ding YD,
Zhou Y, Lei CQ and Shu HB: The zinc-finger protein ZFYVE1 modulates
TLR3-mediated signaling by facilitating TLR3 ligand binding. Cell
Mol Immunol. 17:741–752. 2020. View Article : Google Scholar :
|
|
107
|
Wang Z, Xia Q, Liu X, Liu W, Huang W, Mei
X, Luo J, Shan M, Lin R, Zou D and Ma Z: Phytochemistry,
pharmacology, quality control and future research of Forsythia
suspensa (Thunb.) Vahl: A review. J Ethnopharmacol. 210:318–339.
2018. View Article : Google Scholar
|
|
108
|
Kong F, Jiang X, Wang R, Zhai S, Zhang Y
and Wang D: Forsythoside B attenuates memory impairment and
neuroinflammation via inhibition on NF-κB signaling in Alzheimer's
disease. J Neuroinflammation. 17:3052020. View Article : Google Scholar
|
|
109
|
Lu TC, Wu YH, Chen WY and Hung YC:
Targeting oxidative stress and endothelial dysfunction using
tanshinone IIA for the treatment of tissue inflammation and
fibrosis. Oxid Med Cell Longev. 2022:28117892022. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Lu BL, Li J, Zhou J, Li WW and Wu HF:
Tanshinone IIA decreases the levels of inflammation induced by
Aβ1-42 in brain tissues of Alzheimer's disease model rats.
Neuroreport. 27:883–893. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Jiang P, Li C, Xiang Z and Jiao B:
Tanshinone IIA reduces the risk of Alzheimer's disease by
inhibiting iNOS, MMP-2 and NF-κBp65 transcription and translation
in the temporal lobes of rat models of Alzheimer's disease. Mol Med
Rep. 10:689–694. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Maione F, Piccolo M, De Vita S, Chini MG,
Cristiano C, De Caro C, Lippiello P, Miniaci MC, Santamaria R,
Irace C, et al: Down regulation of pro-inflammatory pathways by
tanshinone IIA and cryptotanshinone in a non-genetic mouse model of
Alzheimer's disease. Pharmacol Res. 129:482–490. 2018. View Article : Google Scholar
|
|
113
|
Ding B, Lin C, Liu Q, He Y, Ruganzu JB,
Jin H, Peng X, Ji S, Ma Y and Yang W: Tanshinone IIA attenuates
neuroinflammation via inhibiting RAGE/NF-κB signaling pathway in
vivo and in vitro. J Neuroinflammation. 17:3022020. View Article : Google Scholar
|
|
114
|
Xing L, Tan ZR, Cheng JL, Huang WH, Zhang
W, Deng W, Yuan CS and Zhou HH: Bioavailability and pharmacokinetic
comparison of tanshinones between two formulations of Salvia
miltiorrhiza in healthy volunteers. Sci Rep. 7:47092017. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Hu X, Zhang J, Deng L, Hu H, Hu J and
Zheng G: Galactose-modified PH-sensitive niosomes for controlled
release and hepatocellular carcinoma target delivery of tanshinone
IIA. AAPS PharmSciTech. 22:962021. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Zhang X, Kang X, Du L, Zhang L, Huang Y,
Wang J, Wang S, Chang Y, Liu Y and Zhao Y: Tanshinone IIA loaded
chitosan nanoparticles decrease toxicity of β-amyloid peptide in a
Caenorhabditis elegans model of disease. Free Radical Bio Med.
193:81–94. 2022. View Article : Google Scholar
|
|
117
|
Thundyil J, Pavlovski D, Sobey CG and
Arumugam TV: Adiponectin receptor signalling in the brain. Brit J
Pharmacol. 165:313–327. 2012. View Article : Google Scholar
|
|
118
|
Ng RC, Cheng OY, Jian M, Kwan JS, Ho PW,
Cheng KK, Yeung PK, Zhou LL, Hoo RL, Chung SK, et al: Chronic
adiponectin deficiency leads to Alzheimer's disease-like cognitive
impairments and pathologies through AMPK inactivation and cerebral
insulin resistance in aged mice. Mol Neurodegener. 11:712016.
View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Boza-Serrano A, Yang Y, Paulus A and
Deierborg T: Innate immune alterations are elicited in microglial
cells before plaque deposition in the Alzheimer's disease mouse
model 5xFAD. Sci Rep. 8:15502018. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Jian M, Kwan JSC, Bunting M, Ng RCL and
Chan KH: Adiponectin suppresses amyloid-β oligomer (AβO)-induced
inflammatory response of microglia via AdipoR1-AMPK-NF-κB signaling
pathway. J Neuroinflammation. 16:1102019. View Article : Google Scholar
|
|
121
|
Chen X, Su J, Wang R, Hao R, Fu C, Chen J,
Li J and Wang X: Structural optimization of cannabidiol as
multifunctional cosmetic raw materials. Antioxidants (Basel).
12:3142023. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Malakoti F, Targhazeh N, Abadifard E,
Zarezadeh R, Samemaleki S, Asemi Z, Younesi S, Mohammadnejad R,
Hadi Hossini S, Karimian A, et al: DNA repair and damage pathways
in mesothelioma development and therapy. Cancer Cell Int.
22:1762022. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Schiffmann SN and Vanderhaeghen JJ:
Distribution of cells containing mRNA encoding cholecystokinin in
the rat central nervous system. J Comp Neurol. 304:219–233. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Hensley K, Floyd RA, Zheng NY, Nael R,
Robinson KA, Nguyen X, Pye QN, Stewart CA, Geddes J, Markesbery WR,
et al: p38 kinase is activated in the Alzheimer's disease brain. J
Neurochem. 72:2053–2058. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Zhu X, Raina AK, Rottkamp CA, Aliev G,
Perry G, Boux H and Smith MA: Activation and redistribution of
c-jun N-terminal kinase/stress activated protein kinase in
degenerating neurons in Alzheimer's disease. J Neurochem.
76:435–441. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Jin Y, Yan EZ, Fan Y, Zong ZH, Qi ZM and
Li Z: Sodium ferulate prevents amyloid-beta-induced neurotoxicity
through suppression of p38 MAPK and upregulation of ERK-1/2 and
Akt/protein kinase B in rat hippocampus. Acta Pharmacol Sin.
26:943–951. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
McDonald DR, Bamberger ME, Combs CK and
Landreth GE: beta-Amyloid fibrils activate parallel
mitogen-activated protein kinase pathways in microglia and THP1
monocytes. J Neurosci. 18:4451–4460. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Koistinaho M and Koistinaho J: Role of p38
and p44/42 mitogen-activated protein kinases in microglia. Glia.
40:175–183. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Kim SH, Smith CJ and Van Eldik LJ:
Importance of MAPK pathways for microglial pro-inflammatory
cytokine IL-1 beta production. Neurobiol Aging. 25:431–439. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Liang Z, Zhang B, Xu M, Morisseau C, Hwang
SH, Hammock BD and Li QX:
1-Trifluoromethoxyphenyl-3-(1-propionylpiperidin-4-yl) urea, a
selective and potent dual inhibitor of soluble epoxide hydrolase
and p38 kinase intervenes in Alzheimer's signaling in human nerve
cells. ACS Chem Neurosci. 10:4018–4030. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Chang D, Li H, Qian C and Wang Y: DiOHF
protects against doxorubicin-induced cardiotoxicity through ERK1
signaling pathway. Front Pharmacol. 10:10812019. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Lee HC, Yu HP, Liao CC, Chou AH and Liu
FC: Escin protects against acetaminophen-induced liver injury in
mice via attenuating inflammatory response and inhibiting ERK
signaling pathway. Am J Transl Res. 11:5170–5182. 2019.PubMed/NCBI
|
|
133
|
Sclip A, Tozzi A, Abaza A, Cardinetti D,
Colombo I, Calabresi P, Salmona M, Welker E and Borsello T: c-Jun
N-terminal kinase has a key role in Alzheimer disease synaptic
dysfunction in vivo. Cell Death Dis. 5:e10192014. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Jeong YH, Li W, Go Y and Oh YC:
Atractylodis rhizoma alba attenuates neuroinflammation in BV2
microglia upon LPS stimulation by inducing HO-1 activity and
inhibiting NF-κB and MAPK. Int J Mol Sci. 20:40152019. View Article : Google Scholar
|
|
135
|
Dang TK, Hong SM, Dao VT, Tran PTT, Tran
HT, Do GH, Hai TN, Nguyet Pham HT and Kim SY:
Anti-neuroinflammatory effects of alkaloid-enriched extract from
Huperzia serrata on lipopolysaccharide-stimulated BV-2 microglial
cells. Pharm Biol. 61:135–143. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Sun A, Liu M, Nguyen XV and Bing G: P38
MAP kinase is activated at early stages in Alzheimer's disease
brain. Exp Neurol. 183:394–405. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Schnöder L, Hao W, Qin Y, Liu S, Tomic I,
Liu X, Fassbender K and Liu Y: Deficiency of neuronal p38α MAPK
attenuates amyloid pathology in Alzheimer disease mouse and cell
models through facilitating lysosomal degradation of BACE1. J Biol
Chem. 291:2067–2079. 2016. View Article : Google Scholar
|
|
138
|
Wu H, Hu B, Zhou X, Zhou C, Meng J, Yang
Y, Zhao X, Shi Z and Yan S: Artemether attenuates LPS-induced
inflammatory bone loss by inhibiting osteoclastogenesis and bone
resorption via suppression of MAPK signaling pathway. Cell Death
Dis. 9:4982018. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Xing B, Bachstetter AD and Van Eldik LJ:
Microglial p38α MAPK is critical for LPS-induced neuron
degeneration, through a mechanism involving TNFα. Mol Neurodegener.
6:842011. View Article : Google Scholar
|
|
140
|
Munoz L, Ralay Ranaivo H, Roy SM, Hu W,
Craft JM, McNamara LK, Chico LW, Van Eldik LJ and Watterson DM: A
novel p38 alpha MAPK inhibitor suppresses brain proinflammatory
cytokine up-regulation and attenuates synaptic dysfunction and
behavioral deficits in an Alzheimer's disease mouse model. J
Neuroinflammation. 4:212007. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Maphis N, Jiang S, Xu G, Kokiko-Cochran
ON, Roy SM, Van Eldik LJ, Watterson DM, Lamb BT and Bhaskar K:
Selective suppression of the α isoform of p38 MAPK rescues
late-stage tau pathology. Alzheimers Res Ther. 8:542016. View Article : Google Scholar
|
|
142
|
Haddad JJ: VX-745. Vertex pharmaceuticals.
Curr Opin Investig Drugs. 2:1070–1076. 2001.
|
|
143
|
Alam JJ: Selective brain-targeted
antagonism of p38 MAPKα reduces hippocampal IL-1β levels and
improves morris water maze performance in aged rats. J Alzheimers
Dis. 48:219–227. 2015. View Article : Google Scholar
|
|
144
|
Hitti E, Iakovleva T, Brook M, Deppenmeier
S, Gruber AD, Radzioch D, Clark AR, Blackshear PJ, Kotlyarov A and
Gaestel M: Mitogen-activated protein kinase-activated protein
kinase 2 regulates tumor necrosis factor mRNA stability and
translation mainly by altering tristetraprolin expression,
stability, and binding to adenine/uridine-rich element. Mol Cell
Biol. 26:2399–2407. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Culbert AA, Skaper SD, Howlett DR, Evans
NA, Facci L, Soden PE, Seymour ZM, Guillot F, Gaestel M and
Richardson JC: MAPK-activated protein kinase 2 deficiency in
microglia inhibits pro-inflammatory mediator release and resultant
neurotoxicity. Relevance to neuroinflammation in a transgenic mouse
model of Alzheimer disease. J Biol Chem. 281:23658–23667. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Brugnano JL, Chan BK, Seal BL and Panitch
A: Cell-penetrating peptides can confer biological function:
regulation of inflammatory cytokines in human monocytes by MK2
inhibitor peptides. J Control Release. 155:128–133. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Jiang J, Wang Z, Liang X, Nie Y, Chang X,
Xue H, Li S and Min C: Intranasal MMI-0100 attenuates
Aβ1-42 - and LPS-induced neuroinflammation and memory
impairments via the MK2 signaling pathway. Front Immunol.
10:27072019. View Article : Google Scholar
|
|
148
|
Mittal D, Ali A, Md S, Baboota S, Sahni JK
and Ali J: Insights into direct nose to brain delivery: current
status and future perspective. Drug Deliv. 21:75–86. 2014.
View Article : Google Scholar
|
|
149
|
Mistry A, Stolnik S and Illum L:
Nose-to-brain delivery: Investigation of the transport of
nanoparticles with different surface characteristics and sizes in
excised porcine olfactory epithelium. Mol Pharm. 12:2755–2766.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Zakaria ZA, Patahuddin H, Mohamad AS,
Israf DA and Sulaiman MR: In vivo anti-nociceptive and
anti-inflammatory activities of the aqueous extract of the leaves
of Piper sarmentosum. J Ethnopharmacol. 128:42–48. 2010. View Article : Google Scholar
|
|
151
|
Chan EWL, Yeo ETY, Wong KWL, See ML, Wong
KY and Gan SY: Piper sarmentosum Roxb. Root extracts confer
neuroprotection by attenuating beta amyloid-induced
pro-inflammatory cytokines released from microglial cells. Curr
Alzheimer Res. 16:251–260. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Wang X, Zhao X and Tang S: Inhibitory
effects of EGb761 on the expression of matrix metalloproteinases
(MMPs) and cartilage matrix destruction. Cell Stress Chaperon.
20:781–786. 2015. View Article : Google Scholar
|
|
153
|
Meng M, Ai D, Sun L, Xu X and Cao X: EGb
761 inhibits Aβ1-42-induced neuroinflammatory response by
suppressing P38 MAPK signaling pathway in BV-2 microglial cells.
Neuroreport. 30:434–440. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Marchand G, Wambang N, Pellegrini S,
Molinaro C, Martoriati A, Bousquet T, Markey A, Lescuyer-Rousseau
A, Bodart JF, Cailliau K, et al: Effects of ferrocenyl
4-(Imino)-1,4-dihydro-quinolines on xenopus laevis prophase
I-arrested oocytes: Survival and hormonal-induced M-phase entry.
Int J Mol Sci. 21:30492020. View Article : Google Scholar
|
|
155
|
Chiu YJ, Hsieh YH, Lin TH, Lee GC,
Hsieh-Li HM, Sun YC, Chen CM, Chang KH and Lee-Chen GJ: Novel
compound VB-037 inhibits Aβ aggregation and promotes neurite
outgrowth through enhancement of HSP27 and reduction of P38 and
JNK-mediated inflammation in cell models for Alzheimer's disease.
Neurochem Int. 125:175–186. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Czarnecka K, Girek M, Maciejewska K,
Skibiński R, Jończyk J, Bajda M, Kabziński J, Sołowiej P, Majsterek
I and Szymański P: New cyclopentaquinoline hybrids with
multifunctional capacities for the treatment of Alzheimer's
disease. J Enzym Inhib Med Chem. 33:158–170. 2017. View Article : Google Scholar
|
|
157
|
Olajide OA, Bhatia HS, de Oliveira AC,
Wright CW and Fiebich BL: Inhibition of neuroinflammation in
LPS-activated microglia by cryptolepine. Evid Based Complement
Alternat Med. 2013:4597232013. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Castro-Torres RD, Busquets O, Parcerisas
A, Verdaguer E, Olloquequi J, Ettcheto M, Beas-Zarate C, Folch J,
Camins A and Auladell C: Involvement of JNK1 in neuronal
polarization during brain development. Cells. 9:18972020.
View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Ma Z, Zang T, Birnbaum SG, Wang Z, Johnson
JE, Zhang CL and Parada LF: TrkB dependent adult hippocampal
progenitor differentiation mediates sustained ketamine
antidepressant response. Nat Commun. 8:16682017. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Guo YJ, Pan WW, Liu SB, Shen ZF, Xu Y and
Hu LL: ERK/MAPK signalling pathway and tumorigenesis. Exp Ther Med.
19:1997–2007. 2020.PubMed/NCBI
|
|
161
|
Paquet C, Nicoll JA, Love S, Mouton-Liger
F, Holmes C, Hugon J and Boche D: Downregulated apoptosis and
autophagy after anti-Aβ immunotherapy in Alzheimer's disease. Brain
Pathol. 28:603–610. 2018. View Article : Google Scholar
|
|
162
|
Yuan Y, Chen J, Ge X, Deng J, Xu X, Zhao Y
and Wang H: Activation of ERK-Drp1 signaling promotes
hypoxia-induced Aβ accumulation by upregulating mitochondrial
fission and BACE1 activity. Febs Open Bio. 11:2740–2755. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Ho WC, Hsu CC, Huang HJ, Wang HT and Lin
AMY: Anti-inflammatory effect of AZD6244 on acrolein-induced
neuroinflammation. Mol Neurobiol. 57:88–95. 2020. View Article : Google Scholar
|
|
164
|
Qiu Z, Lu P, Wang K, Zhao X, Li Q, Wen J,
Zhang H, Li R, Wei H, Lv Y, et al: Dexmedetomidine inhibits
neuroinflammation by altering microglial M1/M2 polarization through
MAPK/ERK pathway. Neurochem Res. 45:345–353. 2020. View Article : Google Scholar
|
|
165
|
Goldfarb G, Doan Ba Tri and Duran A: Human
placenta for chronic leg ulcers. Lancet. 2:401980. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Lee KW, Ji HM, Kim DW, Choi SM, Kim S and
Yang EJ: Effects of Hominis placenta on LPS-induced cell toxicity
in BV2 microglial cells. J Ethnopharmacol. 147:286–292. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Filocamo A, Bisignano C, Ferlazzo N, Cirmi
S, Mandalari G and Navarra M: In vitro effect of bergamot (Citrus
bergamia) juice against cagA-positive and-negative clinical
isolates of Helicobacter pylori. BMC Complement Altern Med.
15:2562015. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Currò M, Risitano R, Ferlazzo N, Cirmi S,
Gangemi C, Caccamo D, Ientile R and Navarra M: Citrus bergamia
juice extract attenuates β-amyloid-induced pro-inflammatory
activation of THP-1 cells through MAPK and AP-1 pathways. Sci Rep.
6:208092016. View Article : Google Scholar
|
|
169
|
Eglitis MA and Mezey E: Hematopoietic
cells differentiate into both microglia and macroglia in the brains
of adult mice. Proc Natl Acad Sci USA. 94:4080–4085. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Hickey WF and Kimura H: Perivascular
microglial cells of the CNS are bone marrow-derived and present
antigen in vivo. Science. 239:290–292. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Tao M, Zheng D, Liang X, Wu D, Hu K, Jin J
and He Q: Tripterygium glycoside suppresses
epithelial-to-mesenchymal transition of diabetic kidney disease
podocytes by targeting autophagy through the mTOR/Twist1 pathway.
Mol Med Rep. 24:5922021. View Article : Google Scholar :
|
|
172
|
Tang L, Xiang Q, Xiang J, Zhang Y and Li
J: Tripterygium glycoside ameliorates neuroinflammation in a mouse
model of Aβ25-35-induced Alzheimer's disease by inhibiting the
phosphorylation of IκBα and p38. Bioengineered. 12:8540–8554. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Cao X, Jin Y, Zhang H, Yu L, Bao X, Li F
and Xu Y: The antiinflammatory effects of
4-[(5-Bromo-3-chloro-2-hydroxybenzyl) amino]-2-hydroxybenzoic acid
in lipopolysaccharide-activated primary microglial cells.
Inflammation. 41:530–540. 2018. View Article : Google Scholar
|
|
174
|
Kim ME, Jung I, Na JY, Lee Y, Lee J and
Lee JS and Lee JS: Pseudane-VII regulates LPS-induced
neuroinflammation in brain microglia cells through the inhibition
of iNOS expression. Molecules. 23:31962018. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Zhao H, Wang SL, Qian L, Jin JL, Li H, Xu
Y and Zhu XL: Diammonium glycyrrhizinate attenuates
Aβ(1-42)-induced neuroinflammation and regulates MAPK and NF-κB
pathways in vitro and in vivo. CNS Neurosci Ther. 19:117–124. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Ramana KV and Srivastava SK: Aldose
reductase: A novel therapeutic target for inflammatory pathologies.
Int J Biochem Cell Biol. 42:17–20. 2010. View Article : Google Scholar
|
|
177
|
Song XM, Yu Q, Dong X, Yang HO, Zeng KW,
Li J and Tu PF: Aldose reductase inhibitors attenuate
β-amyloid-induced TNF-α production in microlgia via
ROS-PKC-mediated NF-κB and MAPK pathways. Int Immunopharmacol.
50:30–37. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Meza CA, La Favor JD, Kim DH and Hickner
RC: Endothelial dysfunction: Is there a hyperglycemia-induced
imbalance of NOX and NOS? Int J Mol Sci. 20:37752019. View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Lee J, Narayan VP, Hong EY, Whang WK and
Park T: Artemisia iwayomogi extract attenuates high-fat
diet-induced hypertriglyceridemia in mice: Potential involvement of
the adiponectin-AMPK pathway and very low density lipoprotein
assembly in the liver. Int J Mol Sci. 18:17622017. View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Ju IG, Huh E, Kim N, Lee S, Choi JG, Hong
J and Oh MS: Artemisiae iwayomogii herba inhibits
lipopolysaccharide-induced neuroinflammation by regulating NF-κB
and MAPK signaling pathways. Phytomedicine. 84:1535012021.
View Article : Google Scholar
|
|
181
|
Hilliard A, Mendonca P and Soliman KFA:
Involvement of NFkB and MAPK signaling pathways in the preventive
effects of Ganoderma lucidum on the inflammation of BV-2 microglial
cells induced by LPS. J Neuroimmunol. 345:5772692020. View Article : Google Scholar
|
|
182
|
Cheung WM, Hui WS, Chu PW, Chiu SW and Ip
NY: Ganoderma extract activates MAP kinases and induces the
neuronal differentiation of rat pheochromocytoma PC12 cells. FEBS
Lett. 486:291–296. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Geng X, Zhong D, Su L and Yang B:
Preventive and therapeutic effect of ganoderma (Lingzhi) on renal
diseases and clinical applications. Adv Exp Med Biol. 1182:243–262.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Chen LG, Jan YS, Tsai PW, Norimoto H,
Michihara S, Murayama C and Wang CC: Anti-inflammatory and
antinociceptive constituents of atractylodes japonica koidzumi. J
Agric Food Chem. 64:2254–2262. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Tang JJ, Wang MR, Dong S, Huang LF, He QR
and Gao JM: 1,10-Seco-Eudesmane sesquiterpenoids as a new type of
anti-neuroinflammatory agents by suppressing TLR4/NF-κB/MAPK
pathways. Eur J Med Chem. 224:1137132021. View Article : Google Scholar
|
|
186
|
Choi MS, Jung UJ, Kim HJ, Do GM, Jeon SM,
Kim MJ and Lee MK: Du-zhong (Eucommia ulmoides Oliver) leaf extract
mediates hypolipidemic action in hamsters fed a high-fat diet. Am J
Chin Med. 36:81–93. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
187
|
He X, Wang J, Li M, Hao D, Yang Y, Zhang
C, He R and Tao R: Eucommia ulmoides Oliv.: Ethnopharmacology,
phytochemistry and pharmacology of an important traditional Chinese
medicine. J Ethnopharmacol. 151:78–92. 2014. View Article : Google Scholar
|
|
188
|
Han R, Yuan T, Yang Z, Zhang Q, Wang WW,
Lin LB, Zhu MQ and Gao JM: Ulmoidol, an unusual nortriterpenoid
from Eucommia ulmoides Oliv. Leaves prevents neuroinflammation by
targeting the PU.1 transcriptional signaling pathway. Bioorg Chem.
116:1053452021. View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Zhang C, Hu L, Liu D, Huang J and Lin W:
Circumdatin D exerts neuroprotective effects by attenuating
LPS-induced pro-inflammatory responses and downregulating
acetylcholinesterase activity in vitro and in vivo. Front
Pharmacol. 11:7602020. View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Gong P, Deng F, Zhang W, Ji J, Liu J, Sun
Y and Hu J: Tectorigenin attenuates the MPP+-induced
SH-SY5Y cell damage, indicating a potential beneficial role in
Parkinson's disease by oxidative stress inhibition. Exp Ther Med.
14:4431–4437. 2017.PubMed/NCBI
|
|
191
|
Oh KB, Kang H and Matsuoka H: Detection of
antifungal activity in Belamcanda chinensis by a single-cell
bioassay method and isolation of its active compound, tectorigenin.
Biosci Biotechnol Biochem. 65:939–942. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
192
|
Lim HS, Kim YJ, Kim BY, Park G and Jeong
SJ: The Anti-neuroinflammatory activity of tectorigenin
pretreatment via downregulated NF-κB and ERK/JNK pathways in BV-2
microglial and microglia inactivation in mice with
lipopolysaccharide. Front Pharmacol. 9:4622018. View Article : Google Scholar
|
|
193
|
Wang BR, Shi JQ, Ge NN, Ou Z, Tian YY,
Jiang T, Zhou JS, Xu J and Zhang YD: PM2.5 exposure aggravates
oligomeric amyloid beta-induced neuronal injury and promotes NLRP3
inflammasome activation in an in vitro model of Alzheimer's
disease. J Neuroinflammation. 15:1322018. View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Shao BZ, Xu ZQ, Han BZ, Su DF and Liu C:
NLRP3 inflammasome and its inhibitors: A review. Front Pharmacol.
6:2622015. View Article : Google Scholar : PubMed/NCBI
|
|
195
|
Feng YS, Tan ZX, Wu LY, Dong F and Zhang
F: The involvement of NLRP3 inflammasome in the treatment of
Alzheimer's disease. Ageing Res Rev. 64:1011922020. View Article : Google Scholar : PubMed/NCBI
|
|
196
|
Heneka MT, Kummer MP, Stutz A, Delekate A,
Schwartz S, Vieira-Saecker A, Griep A, Axt D, Remus A, Tzeng TC, et
al: NLRP3 is activated in Alzheimer's disease and contributes to
pathology in APP/PS1 mice. Nature. 493:674–678. 2013. View Article : Google Scholar
|
|
197
|
Halle A, Hornung V, Petzold GC, Stewart
CR, Monks BG, Reinheckel T, Fitzgerald KA, Latz E, Moore KJ and
Golenbock DT: The NALP3 inflammasome is involved in the innate
immune response to amyloid-beta. Nat Immunol. 9:857–865. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
198
|
Lonnemann N, Hosseini S, Marchetti C,
Skouras DB, Stefanoni D, D'Alessandro A, Dinarello CA and Korte M:
The NLRP3 inflammasome inhibitor OLT1177 rescues cognitive
impairment in a mouse model of Alzheimer's disease. Proc Natl Acad
Sci USA. 117:32145–32154. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
199
|
Marchetti C, Swartzwelter B, Gamboni F,
Neff CP, Richter K, Azam T, Carta S, Tengesdal I, Nemkov T,
D'Alessandro A, et al: OLT1177, a β-sulfonyl nitrile compound, safe
in humans, inhibits the NLRP3 inflammasome and reverses the
metabolic cost of inflammation. Proc Natl Acad Sci USA.
115:E1530–E1539. 2018. View Article : Google Scholar
|
|
200
|
Klück V, Jansen TLTA, Janssen M,
Comarniceanu A, Efdé M, Tengesdal IW, Schraa K, Cleophas MCP,
Scribner CL, Skouras DB, et al: Dapansutrile, an oral selective
NLRP3 inflammasome inhibitor, for treatment of gout flares: An
open-label, dose-adaptive, proof-of-concept, phase 2a trial. Lancet
Rheumatol. 2:e270–e280. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
201
|
Dempsey C, Rubio Araiz A, Bryson KJ,
Finucane O, Larkin C, Mills EL, Robertson AAB, Cooper MA, O'Neill
LAJ and Lynch MA: Inhibiting the NLRP3 inflammasome with MCC950
promotes non-phlogistic clearance of amyloid-β and cognitive
function in APP/PS1 mice. Brain Behav Immun. 61:306–316. 2017.
View Article : Google Scholar
|
|
202
|
Fekete C, Vastagh C, Dénes Á, Hrabovszky
E, Nyiri G, Kalló I, Liposits Z and Sárvári M: Chronic amyloid β
oligomer infusion evokes sustained inflammation and microglial
changes in the rat hippocampus via NLRP3. Neuroscience. 405:35–46.
2019. View Article : Google Scholar
|
|
203
|
Kuwar R, Rolfe A, Di L, Blevins H, Xu Y,
Sun X, Bloom GS, Zhang S and Sun D: A novel inhibitor targeting
NLRP3 inflammasome reduces neuropathology and improves cognitive
function in Alzheimer's disease transgenic mice. J Alzheimers Dis.
82:1769–1783. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
204
|
Zhang Y, Zhao Y, Zhang J, Gao Y, Li S,
Chang C, Yu D and Yang G: Ginkgolide B inhibits NLRP3 inflammasome
activation and promotes microglial M2 polarization in
Aβ1-42 -induced microglia cells. Neurosci Lett.
764:1362062021. View Article : Google Scholar
|
|
205
|
Gu JH, Ge JB, Li M, Wu F, Zhang W and Qin
ZH: Inhibition of NF-κB activation is associated with
anti-inflammatory and anti-apoptotic effects of ginkgolide B in a
mouse model of cerebral ischemia/reperfusion injury. Eur J Pharm
Sci. 47:652–660. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
206
|
Kaur N, Dhiman M, Perez-Polo JR and Mantha
AK: Ginkgolide B revamps neuroprotective role of
apurinic/apyrimidinic endonuclease 1 and mitochondrial oxidative
phosphorylation against Aβ25-35-induced neurotoxicity in human
neuroblastoma cells. J Neurosci Res. 93:938–947. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
207
|
Zhang HR, Peng JH, Cheng XB, Shi BZ, Zhang
MY and Xu RX: Paeoniflorin atttenuates amyloidogenesis and the
inflammatory responses in a transgenic mouse model of Alzheimer's
disease. Neurochem Res. 40:1583–1592. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
208
|
Liu HQ, Zhang WY, Luo XT, Ye Y and Zhu XZ:
Paeoniflorin attenuates neuroinflammation and dopaminergic
neurodegeneration in the MPTP model of Parkinson's disease by
activation of adenosine A1 receptor. Brit J Pharmacol. 148:314–325.
2006. View Article : Google Scholar
|
|
209
|
Liu DZ, Xie KQ, Ji XQ, Ye Y, Jiang CL and
Zhu XZ: Neuroprotective effect of paeoniflorin on cerebral ischemic
rat by activating adenosine A1 receptor in a manner different from
its classical agonists. Br J Pharmacol. 146:604–611. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
210
|
English BK, Ihle JN, Myracle A and Yi T:
Hck tyrosine kinase activity modulates tumor necrosis factor
production by murine macrophages. J Exp Med. 178:1017–1022. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
211
|
Kong X, Liao Y, Zhou L, Zhang Y, Cheng J,
Yuan Z and Wang S: Hematopoietic cell kinase (HCK) is essential for
NLRP3 inflammasome activation and lipopolysaccharide-induced
inflammatory response in vivo. Front Pharmacol. 11:5810112020.
View Article : Google Scholar : PubMed/NCBI
|
|
212
|
Cui W, Sun C, Ma Y, Wang S, Wang X and
Zhang Y: Inhibition of TLR4 induces M2 microglial polarization and
provides neuroprotection via the NLRP3 inflammasome in Alzheimer's
disease. Front Neurosci. 14:4442020. View Article : Google Scholar : PubMed/NCBI
|
|
213
|
Hua F, Tang H, Wang J, Prunty MC, Hua X,
Sayeed I and Stein DG: TAK-242, an antagonist for Toll-like
receptor 4, protects against acute cerebral ischemia/reperfusion
injury in mice. J Cereb Blood Flow Metab. 35:536–542. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
214
|
Liu Y, Dai Y, Li Q, Chen C, Chen H, Song
Y, Hua F and Zhang Z: Beta-amyloid activates NLRP3 inflammasome via
TLR4 in mouse microglia. Neurosci Lett. 736:1352792020. View Article : Google Scholar : PubMed/NCBI
|
|
215
|
Poulose SM, Thangthaeng N, Miller MG and
Shukitt-Hale B: Effects of pterostilbene and resveratrol on brain
and behavior. Neurochem Int. 89:227–233. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
216
|
Li Q, Chen L, Liu X, Li X, Cao Y, Bai Y
and Qi F: Pterostilbene inhibits amyloid-β-induced
neuroinflammation in a microglia cell line by inactivating the
NLRP3/caspase-1 inflammasome pathway. J Cell Biochem.
119:7053–7062. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
217
|
Wang SY, Liu Y, Li XM, Algradi AM, Jiang
H, Sun YP, Guan W, Pan J, Kuang HX and Yang BY: Discovery of active
ingredients targeted TREM2 by SPR biosensor-UPLC/MS recognition
system, and investigating the mechanism of anti-neuroinflammatory
activity on the lignin-amides from Datura metel seeds. Molecules.
26:59462021. View Article : Google Scholar : PubMed/NCBI
|
|
218
|
Liu Y, Yang X, Lei Q, Li Z, Hu J, Wen X,
Wang H and Liu Z: PEG-PEI/siROCK2 protects against Aβ42-induced
neurotoxicity in primary neuron cells for Alzheimer disease. Cell
Mol Neurobiol. 35:841–848. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
219
|
Liu Y, Zhang H, Peng A, Cai X, Wang Y,
Tang K, Wu X, Liang Y, Wang L and Li Z: PEG-PEI/siROCK2 inhibits
Aβ42-induced microglial inflammation via NLRP3/caspase 1 pathway.
Neuroreport. 33:26–32. 2022. View Article : Google Scholar
|
|
220
|
Schneider KS, Groß CJ, Dreier RF, Saller
BS, Mishra R, Gorka O, Heilig R, Meunier E, Dick MS, Ćiković T, et
al: The inflammasome drives GSDMD-independent secondary pyroptosis
and IL-1 release in the absence of caspase-1 protease activity.
Cell Rep. 21:3846–3859. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
221
|
Han C, Hu Q, Yu A, Jiao Q and Yang Y:
Mafenide derivatives inhibit neuroinflammation in Alzheimer's
disease by regulating pyroptosis. J Cell Mol Med. 25:10534–10542.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
222
|
Wang CZ, Du GJ, Zhen Z, Calway T and Yuan
CS: Significant dose differences in donepezil purchased from the
United States and Canada. Ann Intern Med. 155:279–280. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
223
|
Birks JS and Harvey RJ: Donepezil for
dementia due to Alzheimer's disease. Cochrane Database Syst Rev.
6:CD0011902018.PubMed/NCBI
|
|
224
|
Wang H, Zong Y, Han Y, Zhao J, Liu H and
Liu Y: Compared of efficacy and safety of high-dose donepezil vs
standard-dose donepezil among elderly patients with Alzheimer's
disease: A systematic review and meta-analysis. Expert Opin Drug
Saf. 21:407–415. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
225
|
Kim J, Lee HJ, Park SK, Park JH, Jeong HR,
Lee S, Lee H, Seol E and Hoe HS: Donepezil regulates LPS and
Aβ-stimulated neuroinflammation through MAPK/NLRP3
inflammasome/STAT3 signaling. Int J Mol Sci. 22:106372021.
View Article : Google Scholar
|
|
226
|
Kim HG, Moon M, Choi JG, Park G, Kim AJ,
Hur J, Lee KT and Oh MS: Donepezil inhibits the amyloid-beta
oligomer-induced microglial activation in vitro and in vivo.
Neurotoxicology. 40:23–32. 2014. View Article : Google Scholar
|
|
227
|
Chen M, Chen Z, Wang Y, Tan Z, Zhu C, Li
Y, Han Z, Chen L, Gao R, Liu L and Chen Q: Mitophagy receptor
FUNDC1 regulates mitochondrial dynamics and mitophagy. Autophagy.
12:689–702. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
228
|
Devi TS, Somayajulu M, Kowluru RA and
Singh LP: TXNIP regulates mitophagy in retinal Müller cells under
high-glucose conditions: Implications for diabetic retinopathy.
Cell Death Dis. 8:e27772017. View Article : Google Scholar
|
|
229
|
Gao Y, Li J, Li J, Hu C and Zhang L, Yan
J, Li L and Zhang L: Tetrahydroxy stilbene glycoside alleviated
inflammatory damage by mitophagy via AMPK related PINK1/Parkin
signaling pathway. Biochem Pharmacol. 177:1139972020. View Article : Google Scholar : PubMed/NCBI
|
|
230
|
Li F, Zhang T, He Y, Gu W, Yang X, Zhao R
and Yu J: Inflammation inhibition and gut microbiota regulation by
TSG to combat atherosclerosis in ApoE−/− mice. J
Ethnopharmacol. 247:1122322020. View Article : Google Scholar
|
|
231
|
Mu Y, Xu Z, Zhou X, Zhang H, Yang Q, Zhang
Y, Xie Y, Kang J, Li F and Wang S:
2,3,5,4'-Tetrahydroxystilbene-2-O-β-D-gluc oside attenuates
ischemia/reperfusion-induced brain injury in rats by promoting
angiogenesis. Planta Med. 83:676–683. 2017.
|
|
232
|
Zhou W, Yang Y, Mei C, Dong P, Mu S, Wu H,
Zhou Y, Zheng Y, Guo F and Yang JQ: Inhibition of rho-kinase
downregulates Th17 cells and ameliorates hepatic fibrosis by
schistosoma japonicum infection. Cells. 8:12622019. View Article : Google Scholar : PubMed/NCBI
|
|
233
|
Kimura T, Horikoshi Y, Kuriyagawa C and
Niiyama Y: Rho/ROCK pathway and noncoding RNAs: Implications in
ischemic stroke and spinal cord injury. Int J Mol Sci.
22:115732021. View Article : Google Scholar : PubMed/NCBI
|
|
234
|
Zhou Y, Su Y, Li B, Liu F, Ryder JW, Wu X,
Gonzalez-DeWhitt PA, Gelfanova V, Hale JE, May PC, et al:
Nonsteroidal anti-inflammatory drugs can lower amyloidogenic
Abeta42 by inhibiting Rho. Science. 302:1215–1217. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
235
|
Scheiblich H and Bicker G: Regulation of
microglial phagocytosis by RhoA/ROCK-inhibiting drugs. Cell Mol
Neurobiol. 37:461–473. 2017. View Article : Google Scholar
|
|
236
|
Alokam R, Singhal S, Srivathsav GS,
Garigipati S, Puppala S, Sriram D and Perumal Y: Design of dual
inhibitors of ROCK-I and NOX2 as potential leads for the treatment
of neuroinflammation associated with various neurological diseases
including autism spectrum disorder. Mol Biosyst. 11:607–617. 2015.
View Article : Google Scholar
|
|
237
|
Moon MY, Kim HJ, Li Y, Kim JG, Jeon YJ,
Won HY, Kim JS, Kwon HY, Choi IG, Ro E, et al: Involvement of small
GTPase RhoA in the regulation of superoxide production in BV2 cells
in response to fibrillar Aβ peptides. Cell Signal. 25:1861–1869.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
238
|
Zhang X, Ye P, Wang D, Liu Y, Cao L, Wang
Y, Xu Y and Zhu C: Involvement of RhoA/ROCK signaling in Aβ-induced
chemotaxis, cytotoxicity and inflammatory response of microglial
BV2 cells. Cell Mol Neurobiol. 39:637–650. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
239
|
van der Meer DLM, Degenhardt T, Väisänen
S, de Groot PJ, Heinäniemi M, de Vries SC, Müller M, Carlberg C and
Kersten S: Profiling of promoter occupancy by PPARalpha in human
hepatoma cells via ChIP-chip analysis. Nucleic Acids Res.
38:2839–2850. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
240
|
Xia P, Pan Y, Zhang F, Wang N, Wang E, Guo
Q and Ye Z: Pioglitazone confers neuroprotection against
ischemia-induced pyroptosis due to its inhibitory effects on
HMGB-1/RAGE and Rac1/ROS pathway by activating PPAR-ɤ. Cell Physiol
Biochem. 45:2351–2368. 2018. View Article : Google Scholar
|
|
241
|
Janani C and Ranjitha Kumari BD: PPAR
gamma gene-a review. Diabetes Metab Syndr. 9:46–50. 2015.
View Article : Google Scholar
|
|
242
|
Stark JM, Coquet JM and Tibbitt CA: The
role of PPAR-γ in allergic disease. Curr Allergy Asthma Rep.
21:452021. View Article : Google Scholar
|
|
243
|
Kumar AP, P P, Kumar BRP, Jeyarani V,
Dhanabal SP and Justin A: Glitazones, PPAR-γ and neuroprotection.
Mini Rev Med Chem. 21:1457–1464. 2021. View Article : Google Scholar
|
|
244
|
Furth N, Pateras IS, Rotkopf R, Vlachou V,
Rivkin I, Schmitt I, Bakaev D, Gershoni A, Ainbinder E, Leshkowitz
D, et al: LATS1 and LATS2 suppress breast cancer progression by
maintaining cell identity and metabolic state. Life Sci Alliance.
1:e2018001712018. View Article : Google Scholar : PubMed/NCBI
|
|
245
|
Villapol S: Roles of peroxisome
proliferator-activated receptor gamma on brain and peripheral
inflammation. Cell Mol Neurobiol. 38:121–132. 2018. View Article : Google Scholar
|
|
246
|
Moosecker S, Gomes P, Dioli C, Yu S,
Sotiropoulos I and Almeida OFX: Activated PPARγ abrogates
misprocessing of amyloid precursor protein, tau missorting and
synaptotoxicity. Front Cell Neurosci. 13:2392019. View Article : Google Scholar
|
|
247
|
de la Monte SM and Wands JR: Molecular
indices of oxidative stress and mitochondrial dysfunction occur
early and often progress with severity of Alzheimer's disease. J
Alzheimers Dis. 9:167–181. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
248
|
Watson GS, Cholerton BA, Reger MA, Baker
LD, Plymate SR, Asthana S, Fishel MA, Kulstad JJ, Green PS, Cook
DG, et al: Preserved cognition in patients with early Alzheimer
disease and amnestic mild cognitive impairment during treatment
with rosiglitazone: A preliminary study. Am J Geriatr Psychiatry.
13:950–958. 2005.PubMed/NCBI
|
|
249
|
Risner ME, Saunders AM, Altman JF, Ormandy
GC, Craft S, Foley IM, Zvartau-Hind ME, Hosford DA and Roses AD;
Rosiglitazone in Alzheimer's Disease Study Group: Efficacy of
rosiglitazone in a genetically defined population with
mild-to-moderate Alzheimer's disease. Pharmacogenomics J.
6:246–254. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
250
|
Mandrekar-Colucci S, Karlo JC and Landreth
GE: Mechanisms underlying the rapid peroxisome
proliferator-activated receptor-γ-mediated amyloid clearance and
reversal of cognitive deficits in a murine model of Alzheimer's
disease. J Neurosci. 32:10117–10128. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
251
|
Moreno S, Farioli-Vecchioli S and Cerù MP:
Immunolocalization of peroxisome proliferator-activated receptors
and retinoid X receptors in the adult rat CNS. Neuroscience.
123:131–145. 2004. View Article : Google Scholar
|
|
252
|
Liu ZJ, Liu W, Liu L, Xiao C, Wang Y and
Jiao JS: Curcumin protects neuron against cerebral ischemia-induced
inflammation through improving PPAR-gamma function. Evid Based
Complement Alternat Med. 2013:4709752013.PubMed/NCBI
|
|
253
|
Song GJ, Nam Y, Jo M, Jung M, Koo JY, Cho
W, Koh M, Park SB and Suk K: A novel small-molecule agonist of
PPAR-γ potentiates an anti-inflammatory M2 glial phenotype.
Neuropharmacology. 109:159–169. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
254
|
Qi Y, Zhang Q and Zhu H: Huang-Lian Jie-Du
decoction: A review on phytochemical, pharmacological and
pharmacokinetic investigations. Chin Med. 14:572019. View Article : Google Scholar : PubMed/NCBI
|
|
255
|
Wong LR, Tan EA, Lim MEJ, Shen W, Lian XL,
Wang Y, Chen L and Ho PCL: Functional effects of berberine in
modulating mitochondrial dysfunction and inflammatory response in
the respective amyloidogenic cells and activated microglial
cells-in vitro models simulating Alzheimer's disease pathology.
Life Sci. 282:1198242012. View Article : Google Scholar
|
|
256
|
Hagl S, Asseburg H, Heinrich M, Sus N,
Blumrich EM, Dringen R, Frank J and Eckert GP: Effects of long-term
rice bran extract supplementation on survival, cognition and brain
mitochondrial function in aged NMRI mice. Neuromolecular Med.
18:347–363. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
257
|
El-Din SS, Abd ES, Rashed L, Fayez S,
Aboulhoda BE, Heikal OA, Galal AF and Nour ZA: Possible role of
rice bran extract in microglial modulation through PPAR-gamma
receptors in alzheimer's disease mice model. Metab Brain Dis.
36:1903–1915. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
258
|
De Felice FG and Ferreira ST:
Inflammation, defective insulin signaling, and mitochondrial
dysfunction as common molecular denominators connecting type 2
diabetes to Alzheimer disease. Diabetes. 63:2262–2272. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
259
|
Bouriche H, Meziti H, Senator A and
Arnhold J: Anti-inflammatory, free radical-scavenging, and
metal-chelating activities of Malva parviflora. Pharm Biol.
49:942–946. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
260
|
Medrano-Jiménez E, Jiménez-Ferrer Carrillo
I, Pedraza-Escalona M, Ramírez-Serrano CE, Álvarez-Arellano L,
Cortés-Mendoza J, Herrera-Ruiz M, Jiménez-Ferrer E, Zamilpa A,
Tortoriello J, et al: Malva parviflora extract ameliorates the
deleterious effects of a high fat diet on the cognitive deficit in
a mouse model of Alzheimer's disease by restoring microglial
function via a PPAR-γ-dependent mechanism. J Neuroinflammation.
16:1432019. View Article : Google Scholar
|
|
261
|
Villapol S, Yaszemski AK, Logan TT,
Sanchez-Lemus E, Saavedra JM and Symes AJ: Candesartan, an
angiotensin II AT1-receptor blocker and PPAR-γ agonist,
reduces lesion volume and improves motor and memory function after
traumatic brain injury in mice. Neuropsychopharmacology.
37:2817–2829. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
262
|
Shindo T, Takasaki K, Uchida K, Onimura R,
Kubota K, Uchida N, Irie K, Katsurabayashi S, Mishima K, Nishimura
R, et al: Ameliorative effects of telmisartan on the inflammatory
response and impaired spatial memory in a rat model of Alzheimer's
disease incorporating additional cerebrovascular disease factors.
Biol Pharm Bull. 35:2141–2147. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
263
|
Li NC, Lee A, Whitmer RA, Kivipelto M,
Lawler E, Kazis LE and Wolozin B: Use of angiotensin receptor
blockers and risk of dementia in a predominantly male population:
Prospective cohort analysis. BMJ. 340:b54652010. View Article : Google Scholar : PubMed/NCBI
|
|
264
|
Wang ZF, Li J, Ma C, Huang C and Li ZQ:
Telmisartan ameliorates Aβ oligomer-induced inflammation via
PPARγ/PTEN pathway in BV2 microglial cells. Biochem Pharmacol.
171:1136742020. View Article : Google Scholar
|
|
265
|
He Z, Li X, Han S, Ren B, Hu X, Li N, Du
X, Ni J, Yang X and Liu Q: Bis(ethylmaltolato)oxidovanadium (IV)
attenuates amyloid-beta-mediated neuroinflammation by inhibiting
NF-κB signaling pathway via a PPARγ-dependent mechanism.
Metallomics. 13:mfab0362021. View Article : Google Scholar
|
|
266
|
Yang Z, Liu B, Yang LE and Zhang C:
Platycodigenin as potential drug candidate for Alzheimer's disease
via modulating microglial polarization and neurite regeneration.
Molecules. 24:32072019. View Article : Google Scholar : PubMed/NCBI
|
|
267
|
Pan D, Xu Y, Zhang L, Su Q, Chen M, Li B,
Xiao Q, Gao Q, Peng X, Jiang B, et al: Gene expression profile in
peripheral blood mononuclear cells of postpartum depression
patients. Sci Rep. 8:101392018. View Article : Google Scholar : PubMed/NCBI
|
|
268
|
Ahn S, Jang DM, Park SC, An S, Shin J, Han
BW and Noh M: Cyclin-dependent kinase 5 inhibitor butyrolactone I
elicits a partial agonist activity of peroxisome
proliferator-activated receptor γ. Biomolecules. 10:2752020.
View Article : Google Scholar
|
|
269
|
Wang J, Zhu G, Sun C, Xiong K, Yao T, Su Y
and Fang H: TAK-242 ameliorates DSS-induced colitis by regulating
the gut microbiota and the JAK2/STAT3 signaling pathway. Microb
Cell Fact. 19:1582020. View Article : Google Scholar : PubMed/NCBI
|
|
270
|
Kulesza DW, Ramji K, Maleszewska M,
Mieczkowski J, Dabrowski M, Chouaib S and Kaminska B: Search for
novel STAT3-dependent genes reveals SERPINA3 as a new STAT3 target
that regulates invasion of human melanoma cells. Lab Invest.
99:1607–1621. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
271
|
Lee DY, Hwang CJ, Choi JY, Park MH, Song
MJ, Oh KW, Han SB, Park WK, Cho HY, Cho SY, et al: KRICT-9 inhibits
neuroinflammation, amyloidogenesis and memory loss in Alzheimer's
disease models. Oncotarget. 8:68654–68667. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
272
|
Zhang ZH, Yu LJ, Hui XC, Wu ZZ, Yin KL,
Yang H and Xu Y: Hydroxy-safflor yellow A attenuates
Aβ1-42-induced inflammation by modulating the
JAK2/STAT3/NF-κB pathway. Brain Res. 1563:72–80. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
273
|
Zheng ZV, Chen J, Lyu H, Lam S, Lu G, Chan
WY and Wong GKC: Novel role of STAT3 in microglia-dependent
neuroinflammation after experimental subarachnoid haemorrhage.
Stroke Vasc Neurol. 7:62–70. 2022. View Article : Google Scholar :
|
|
274
|
Wan J, Fu AK, Ip FC, Ng HK, Hugon J, Page
G, Wang JH, Lai KO, Wu Z and Ip NY: Tyk2/STAT3 signaling mediates
beta-amyloid-induced neuronal cell death: Implications in
Alzheimer's disease. J Neurosci. 30:6873–6881. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
275
|
Carret-Rebillat AS, Pace C, Gourmaud S,
Ravasi L, Montagne-Stora S, Longueville S, Tible M, Sudol E, Chang
RC, Paquet C, et al: Neuroinflammation and Aβ accumulation linked
to systemic inflammation are decreased by genetic PKR
down-regulation. Sci Rep. 5:84892015. View Article : Google Scholar
|
|
276
|
Jin P, Kim JA, Choi DY, Lee YJ, Jung HS
and Hong JT: Anti-inflammatory and anti-amyloidogenic effects of a
small molecule, 2,4-bis(p-hydroxyphenyl)-2-butenal in Tg2576
Alzheimer's disease mice model. J Neuroinflammation. 10:22013.
View Article : Google Scholar : PubMed/NCBI
|
|
277
|
Choi JY, Hwang CJ, Lee DY, Gu SM, Lee HP,
Choi DY, Oh KW, Han SB and Hong JT:
(E)-2-methoxy-4-(3-(4-methoxyphenyl) prop-1-en-1-yl) phenol
ameliorates LPS-mediated memory impairment by inhibition of STAT3
pathway. Neuromolecular Med. 19:555–570. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
278
|
Hussein G, Sankawa U, Goto H, Matsumoto K
and Watanabe H: Astaxanthin, a carotenoid with potential in human
health and nutrition. J Nat Prod. 69:443–449. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
279
|
Han JH, Lee YS, Im JH, Ham YW, Lee HP, Han
SB and Hong JT: Astaxanthin ameliorates lipopolysaccharide-induced
neuroinflammation, oxidative stress and memory dysfunction through
inactivation of the signal transducer and activator of
transcription 3 pathway. Mar Drugs. 17:1232019. View Article : Google Scholar : PubMed/NCBI
|
|
280
|
Safar MM, Shahin NN, Mohamed AF and
Abdelkader NF: Suppression of BACE1 and amyloidogenic/RAGE axis by
sitagliptin ameliorates PTZ kindling-induced cognitive deficits in
rats. Chem Biol Interact. 328:1091442020. View Article : Google Scholar : PubMed/NCBI
|
|
281
|
Wen Y, Yu WH, Maloney B, Bailey J, Ma J,
Marié I, Maurin T, Wang L, Figueroa H, Herman M, et al:
Transcriptional regulation of beta-secretase by p25/cdk5 leads to
enhanced amyloidogenic processing. Neuron. 57:680–690. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
282
|
Millot P, San C, Bennana E, Porte B,
Vignal N, Hugon J, Paquet C, Hosten B and Mouton-Liger F: STAT3
inhibition protects against neuroinflammation and BACE1
upregulation induced by systemic inflammation. Immunol Lett.
228:129–134. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
283
|
Long QH, Wu YG, He LL, Ding L, Tan AH, Shi
HY and Wang P: Suan-Zao-Ren decoction ameliorates synaptic
plasticity through inhibition of the Aβ deposition and JAK2/STAT3
signaling pathway in AD model of APP/PS1 transgenic mice. Chin Med.
16:142021. View Article : Google Scholar
|
|
284
|
Jia T, Xing Z, Wang And H and Li G:
Protective effect of dexmedetomidine on intestinal mucosal barrier
function in rats after cardiopulmonary bypass. Exp Biol Med
(Maywood). 247:498–508. 2022. View Article : Google Scholar
|
|
285
|
Wang LC, Liao LX, Zhao MB, Dong X, Zeng KW
and Tu PF: Protosappanin A exerts anti-neuroinflammatory effect by
inhibiting JAK2-STAT3 pathway in lipopolysaccharide-induced BV2
microglia. Chin J Nat Med. 15:674–679. 2017.PubMed/NCBI
|
|
286
|
Porro C, Cianciulli A, Trotta T,
Lofrumento DD and Panaro MA: Curcumin regulates anti-inflammatory
responses by JAK/STAT/SOCS signaling pathway in BV-2 microglial
cells. Biology (Basel). 8:512019.PubMed/NCBI
|
|
287
|
Amalraj A, Pius A and Gopi S and Gopi S:
Biological activities of curcuminoids, other biomolecules from
turmeric and their derivatives-a review. J Tradit Complement Med.
7:205–233. 2016. View Article : Google Scholar
|
|
288
|
He G and Karin M: NF-κB and STAT3-key
players in liver inflammation and cancer. Cell Res. 21:159–168.
2011. View Article : Google Scholar
|
|
289
|
Sim DY, Lee HJ, Jung JH, Im E, Hwang J,
Kim DS and Kim SH: Suppression of STAT3 phosphorylation and
RelA/p65 acetylation mediated by MicroRNA134 plays a pivotal role
in the apoptotic effect of lambertianic acid. Int J Mol Sci.
20:29932019. View Article : Google Scholar : PubMed/NCBI
|
|
290
|
Song SY, Jung YY, Hwang CJ, Lee HP, Sok
CH, Kim JH, Lee SM, Seo HO, Hyun BK, Choi DY, et al: Inhibitory
effect of ent-Sauchinone on amyloidogenesis via inhibition of
STAT3-mediated NF-κB activation in cultured astrocytes and
microglial BV-2 cells. J Neuroinflammation. 11:1182014. View Article : Google Scholar
|
|
291
|
Kim J, Park JH, Park SK and Hoe HS:
Sorafenib modulates the LPS- and Aβ-induced neuroinflammatory
response in cells, wild-type mice, and 5xFAD mice. Front Immunol.
12:6843442021. View Article : Google Scholar
|
|
292
|
Hei YY, Xin M, Zhang H, Xie XX, Mao S and
Zhang SQ: Synthesis and antitumor activity evaluation of
4,6-disubstituted quinazoline derivatives as novel PI3K inhibitors.
Bioorg Med Chem Lett. 26:4408–4413. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
293
|
You JS, Li CY, Chen W, Wu XL, Huang LJ, Li
RK, Gao F, Zhang MY, Liu HL and Qu WL: A network pharmacology-based
study on Alzheimer disease prevention and treatment of Qiong Yu
Gao. BioData Min. 13:22020. View Article : Google Scholar : PubMed/NCBI
|
|
294
|
Yang S, Chen Z, Cao M, Li R, Wang Z and
Zhang M: Pioglitazone ameliorates Aβ42 deposition in rats with
diet-induced insulin resistance associated with AKT/GSK3β
activation. Mol Med Rep. 15:2588–2594. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
295
|
Liu X, Wang H, Bei J, Zhao J, Jiang G and
Liu X: The protective role of miR-132 targeting HMGA2 through the
PI3K/AKT pathway in mice with Alzheimer's disease. Am J Transl Res.
13:4632–4643. 2021.PubMed/NCBI
|
|
296
|
Hwang SY, Jung JS, Lim SJ, Kim JY, Kim TH,
Cho KH and Han IO: LY294002 inhibits interferon-gamma-stimulated
inducible nitric oxide synthase expression in BV2 microglial cells.
Biochem Biophys Res Commun. 318:691–697. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
297
|
He Y, Zhou A and Jiang W: Toll-like
receptor 4-mediated signaling participates in apoptosis of
hippocampal neurons. Neural Regen Res. 8:2744–2753. 2013.
|
|
298
|
Zu HB, Liu XY and Yao K: DHCR24
overexpression modulates microglia polarization and inflammatory
response via Akt/GSK3β signaling in Aβ25-35 treated BV-2
cells. Life Sci. 260:1184702020. View Article : Google Scholar
|
|
299
|
Sarajärvi T, Haapasalo A, Viswanathan J,
Mäkinen P, Laitinen M, Soininen H and Hiltunen M: Down-regulation
of seladin-1 increases BACE1 levels and activity through enhanced
GGA3 depletion during apoptosis. J Biol Chem. 284:34433–34443.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
300
|
Yang W, Liu Y, Xu QQ, Xian YF and Lin ZX:
Sulforaphene ameliorates neuroinflammation and hyperphosphorylated
tau protein via regulating the PI3K/Akt/GSK-3 β pathway in
experimental models of Alzheimer's disease. Oxid Med Cell Longev.
2020:47541952020. View Article : Google Scholar
|
|
301
|
Ivanovics G and Horvath S: Raphanin, an
antibacterial principle of the radish (Raphanus sativus). Nature.
160:2971947. View Article : Google Scholar : PubMed/NCBI
|
|
302
|
Lonze BE and Ginty DD: Function and
regulation of CREB family transcription factors in the nervous
system. Neuron. 35:605–623. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
303
|
Ahn SH, Suh JS, Jang YK, Kim HS, Choi GH,
Kim E and Kim TJ: Rhynchosia volubilis promotes cell survival via
cAMP-PKA/ERK-CREB pathway. Pharmaceuticals (Basel). 15:732022.
View Article : Google Scholar : PubMed/NCBI
|
|
304
|
Hu YJ, Sun Q, Zhang WH, Huo YJ, Xu CJ and
Liu JF: Specific activation of mGlu2 induced IGF-1R transactivation
in vitro through FAK phosphorylation. Acta Pharmacol Sin.
40:460–467. 2019. View Article : Google Scholar :
|
|
305
|
Viola H, Furman M, Izquierdo LA, Alonso M,
Barros DM, de Souza MM, Izquierdo I and Medina JH: Phosphorylated
cAMP response element-binding protein as a molecular marker of
memory processing in rat hippocampus: effect of novelty. J
Neurosci. 20:RC1122000. View Article : Google Scholar : PubMed/NCBI
|
|
306
|
Dineley KT, Westerman M, Bui D, Bell K,
Ashe KH and Sweatt JD: Beta-amyloid activates the mitogen-activated
protein kinase cascade via hippocampal alpha7 nicotinic
acetylcholine receptors: In vitro and in vivo mechanisms related to
Alzheimer's disease. J Neurosci. 21:4125–4133. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
307
|
Yamamoto-Sasaki M, Ozawa H, Saito T,
Rösler M and Riederer P: Impaired phosphorylation of cyclic AMP
response element binding protein in the hippocampus of dementia of
the Alzheimer type. Brain Res. 824:300–303. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
308
|
Mizuno M, Yamada K, Maekawa N, Saito K,
Seishima M and Nabeshima T: CREB phosphorylation as a molecular
marker of memory processing in the hippocampus for spatial
learning. Behav Brain Res. 133:135–141. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
309
|
Sharma VK and Singh TG: CREB: A
multifaceted target for Alzheimer's disease. Curr Alzheimer Res.
17:1280–1293. 2020. View Article : Google Scholar
|
|
310
|
Li C, Chen T, Zhou H, Feng Y, Hoi MPM, Ma
D, Zhao C, Zheng Y and Lee SMY: BHDPC is a novel neuroprotectant
that provides anti-neuroinflammatory and neuroprotective effects by
inactivating NF-κB and activating PKA/CREB. Front Pharmacol.
9:6142018. View Article : Google Scholar
|
|
311
|
Ghosh M, Xu Y and Pearse DD: Cyclic AMP is
a key regulator of M1 to M2a phenotypic conversion of microglia in
the presence of Th2 cytokines. J Neuroinflammation. 13:92016.
View Article : Google Scholar : PubMed/NCBI
|
|
312
|
Tripathi MK, Kartawy M and Amal H: The
role of nitric oxide in brain disorders: Autism spectrum disorder
and other psychiatric, neurological, and neurodegenerative
disorders. Redox Biol. 34:1015672020. View Article : Google Scholar : PubMed/NCBI
|
|
313
|
Zhang J, Guo J, Zhao X, Chen Z, Wang G,
Liu A, Wang Q, Zhou W, Xu Y and Wang C: Phosphodiesterase-5
inhibitor sildenafil prevents neuroinflammation, lowers
beta-amyloid levels and improves cognitive performance in APP/PS1
transgenic mice. Behav Brain Res. 250:230–237. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
314
|
Zeitlin R, Patel S, Burgess S, Arendash GW
and Echeverria V: Caffeine induces beneficial changes in PKA
signaling and JNK and ERK activities in the striatum and cortex of
Alzheimer's transgenic mice. Brain Res. 1417:127–136. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
315
|
Bitner RS, Markosyan S, Nikkel AL and
Brioni JD: In-vivo histamine H3 receptor antagonism activates
cellular signaling suggestive of symptomatic and disease modifying
efficacy in Alzheimer's disease. Neuropharmacology. 60:460–466.
2011. View Article : Google Scholar
|
|
316
|
Medhurst AD, Atkins AR, Beresford IJ,
Brackenborough K, Briggs MA, Calver AR, Cilia J, Cluderay JE, Crook
B, Davis JB, et al: GSK189254, a novel H3 receptor antagonist that
binds to histamine H3 receptors in Alzheimer's disease brain and
improves cognitive performance in preclinical models. J Pharmacol
Exp Ther. 321:1032–1045. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
317
|
Wang J, Liu B, Sun F, Xu Y, Luan H, Yang
M, Wang C, Zhang T, Zhou Z and Yan H: Histamine H3R antagonist
counteracts the impaired hippocampal neurogenesis in
lipopolysaccharide-induced neuroinflammation. Int Immunopharmacol.
110:1090452022. View Article : Google Scholar : PubMed/NCBI
|
|
318
|
Hiraga N, Adachi N, Liu K, Nagaro T and
Arai T: Suppression of inflammatory cell recruitment by histamine
receptor stimulation in ischemic rat brains. Eur J Pharmacol.
557:236–244. 2007. View Article : Google Scholar
|
|
319
|
Amin FU, Shah SA, Badshah H, Khan M and
Kim MO: Anthocyanins encapsulated by PLGA@PEG nanoparticles
potentially improved its free radical scavenging capabilities via
p38/JNK pathway against Aβ1-42-induced oxidative stress.
J Nanobiotechnology. 15:122017. View Article : Google Scholar
|
|
320
|
von Otter M, Landgren S, Nilsson S,
Zetterberg M, Celojevic D, Bergström P, Minthon L, Bogdanovic N,
Andreasen N, Gustafson DR, et al: Nrf2-encoding NFE2L2 haplotypes
influence disease progression but not risk in Alzheimer's disease
and age-related cataract. Mech Ageing Dev. 131:105–110. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
321
|
Kanninen K, Malm TM, Jyrkkänen HK,
Goldsteins G, Keksa-Goldsteine V, Tanila H, Yamamoto M,
Ylä-Herttuala S, Levonen AL and Koistinaho J: Nuclear factor
erythroid 2-related factor 2 protects against beta amyloid. Mol
Cell Neurosci. 39:302–313. 2008. View Article : Google Scholar
|
|
322
|
Li XX, Zheng X, Liu Z, Xu Q, Tang H, Feng
J, Yang S, Vong CT, Gao H and Wang Y: Cryptotanshinone from Salvia
miltiorrhiza Bunge (Danshen) inhibited inflammatory responses via
TLR4/MyD88 signaling pathway. Chin Med. 15:202020. View Article : Google Scholar : PubMed/NCBI
|
|
323
|
Fragoulis A, Siegl S, Fendt M, Jansen S,
Soppa U, Brandenburg LO, Pufe T, Weis J and Wruck CJ: Oral
administration of methysticin improves cognitive deficits in a
mouse model of Alzheimer's disease. Redox Biol. 12:843–853. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
324
|
Bilia AR, Scalise L, Bergonzi MC and
Vincieri FF: Analysis of kavalactones from Piper methysticum
(kava-kava). J Chromatogr B Analyt Technol Biomed Life Sci.
812:203–214. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
325
|
Mattioli R, Francioso A, d'Erme M, Trovato
M, Mancini P, Piacentini L, Casale AM, Wessjohann L, Gazzino R,
Costantino P and Mosca L: Anti-inflammatory activity of a
polyphenolic extract from Arabidopsis thaliana in in vitro and in
vivo models of Alzheimer's disease. Int J Mol Sci. 20:7082019.
View Article : Google Scholar : PubMed/NCBI
|
|
326
|
Rateb ME, Houssen WE, Schumacher M,
Harrison WT, Diederich M, Ebel R and Jaspars M: Bioactive diterpene
derivatives from the marine sponge Spongionella sp. J Nat Prod.
72:1471–1476. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
327
|
Alvariño R, Alonso E, Abbasov ME, Chaheine
CM, Conner ML, Romo D, Alfonso A and Botana LM: Gracilin A
derivatives target early events in Alzheimer's disease: In vitro
effects on neuroinflammation and oxidative stress. ACS Chem
Neurosci. 10:4102–4111. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
328
|
Wang X and Asghar M: Protein disulfide
isomerase regulates renal AT1 receptor function and blood pressure
in rats. Am J Physiol Renal Physiol. 313:F461–F466. 2017.
View Article : Google Scholar :
|
|
329
|
Itoh K, Mimura J and Yamamoto M: Discovery
of the negative regulator of Nrf2, Keap1: A historical overview.
Antioxid Redox Signal. 13:1665–1678. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
330
|
Wu H, Zhao G, Jiang K, Li C, Qiu C and
Deng G: Engeletin alleviates lipopolysaccharide-induced
endometritis in mice by inhibiting TLR4-mediated NF-κB activation.
J Agric Food Chem. 64:6171–6178. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
331
|
Huang Z, Ji H, Shi J, Zhu X and Zhi Z:
Engeletin attenuates Aβ1-42-induced oxidative stress and
neuroinflammation by Keap1/Nrf2 pathway. Inflammation.
43:1759–1771. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
332
|
Barone E, Di Domenico F, Sultana R, Coccia
R, Mancuso C, Perluigi M and Butterfield DA: Heme oxygenase-1
posttranslational modifications in the brain of subjects with
Alzheimer disease and mild cognitive impairment. Free Radic Biol
Med. 52:2292–2301. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
333
|
Zou Y, Hong B, Fan L, Zhou L, Liu Y, Wu Q,
Zhang X and Dong M: Protective effect of puerarin against
beta-amyloid-induced oxidative stress in neuronal cultures from rat
hippocampus: Involvement of the GSK-3β/Nrf2 signaling pathway. Free
Radic Res. 47:55–63. 2013. View Article : Google Scholar
|
|
334
|
Eom HW, Park SY, Kim YH, Seong SJ, Jin ML,
Ryu EY, Kim MJ and Lee SJ: Bambusae caulis in taeniam modulates
neuroprotective and anti-neuroinflammatory effects in hippocampal
and microglial cells via HO-1- and Nrf-2-mediated pathways. Int J
Mol Med. 30:1512–1520. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
335
|
Chen J, Yin W, Tu Y, Wang S, Yang X, Chen
Q, Zhang X, Han Y and Pi R: L-F001, a novel multifunctional ROCK
inhibitor, suppresses neuroinflammation in vitro and in vivo:
Involvement of NF-κB inhibition and Nrf2 pathway activation. Eur J
Pharmacol. 806:1–9. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
336
|
Zhan TW, Tian YX, Wang Q, Wu ZX, Zhang WP,
Lu YB and Wu M: Cangrelor alleviates pulmonary fibrosis by
inhibiting GPR17-mediated inflammation in mice. Int
Immunopharmacol. 62:261–269. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
337
|
Jin S, Wang X, Xiang X, Wu Y, Hu J, Li Y,
Lin DY, Tan Y and Wu X: Inhibition of GPR17 with cangrelor improves
cognitive impairment and synaptic deficits induced by
Aβ1-42 through Nrf2/HO-1 and NF-κB signaling pathway in
mice. Int Immunopharmacol. 101:1083352021. View Article : Google Scholar
|
|
338
|
Gao X, He D, Liu D, Hu G, Zhang Y, Meng T,
Su Y, Zhou A, Huang B, Du J and Fu S: Beta-naphthoflavone inhibits
LPS-induced inflammation in BV-2 cells via AKT/Nrf-2/HO-1-NF-kappaB
signaling axis. Immunobiology. 225:1519652020. View Article : Google Scholar
|
|
339
|
Just PA, Charawi S, Denis RGP, Savall M,
Traore M, Foretz M, Bastu S, Magassa S, Senni N, Sohier P, et al:
Lkb1 suppresses amino acid-driven gluconeogenesis in the liver. Nat
Commun. 11:61272020. View Article : Google Scholar : PubMed/NCBI
|
|
340
|
Marinangeli C, Didier S, Ahmed T,
Caillerez R, Domise M, Laloux C, Bégard S, Carrier S, Colin M,
Marchetti P, et al: AMP-activated protein kinase is essential for
the maintenance of energy levels during synaptic activation.
iScience. 9:1–13. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
341
|
Vingtdeux V, Davies P, Dickson DW and
Marambaud P: AMPK is abnormally activated in tangle- and
pre-tangle-bearing neurons in Alzheimer's disease and other
tauopathies. Acta Neuropathol. 121:337–349. 2011. View Article : Google Scholar :
|
|
342
|
Shah SA, Yoon GH, Chung SS, Abid MN, Kim
TH, Lee HY and Kim MO: Novel osmotin inhibits SREBP2 via the
AdipoR1/AMPK/SIRT1 pathway to improve Alzheimer's disease
neuropathological deficits. Mol Psychiatry. 22:407–416. 2017.
View Article : Google Scholar :
|
|
343
|
Vingtdeux V, Chandakkar P, Zhao H,
d'Abramo C, Davies P and Marambaud P: Novel synthetic
small-molecule activators of AMPK as enhancers of autophagy and
amyloid-β peptide degradation. FASEB J. 25:219–231. 2011.
View Article : Google Scholar :
|
|
344
|
Domise M, Didier S, Marinangeli C, Zhao H,
Chandakkar P, Buée L, Viollet B, Davies P, Marambaud P and
Vingtdeux V: AMP-activated protein kinase modulates tau
phosphorylation and tau pathology in vivo. Sci Rep. 6:267582016.
View Article : Google Scholar : PubMed/NCBI
|
|
345
|
Li C, Zhang C, Zhou H, Feng Y, Tang F, Hoi
M, He C, Ma D, Zhao C and Lee S: Inhibitory effects of betulinic
acid on LPS-induced neuroinflammation involve M2 microglial
polarization via CaMKKβ-Dependent AMPK activation. Front Mol
Neurosci. 11:982018. View Article : Google Scholar
|
|
346
|
Voss U and Ekblad E:
Lipopolysaccharide-induced loss of cultured rat myenteric
neurons-role of AMP-activated protein kinase. PLoS One.
9:e1140442014. View Article : Google Scholar
|
|
347
|
Zhou Z, Zhang L, Liu Y, Huang C, Xia W,
Zhou H, Zhou Z and Zhou X: Luteolin protects chondrocytes from H O
oxidative injury and attenuates osteoarthritis progression by 2
2-induced activating AMPK-Nrf2 signaling. Oxid Med Cell Longev.
2022:56357972022. View Article : Google Scholar
|
|
348
|
Zhou F, Wang M, Ju J, Wang Y, Liu Z, Zhao
X, Yan Y, Yan S, Luo X and Fang Y: Schizandrin A protects against
cerebral ischemia-reperfusion injury by suppressing inflammation
and oxidative stress and regulating the AMPK/Nrf2 pathway
regulation. Am J Transl Res. 11:199–209. 2019.PubMed/NCBI
|
|
349
|
Park SY, Choi MH, Park G and Choi YW:
Petasites japonicus bakkenolide B inhibits
lipopolysaccharide-induced pro-inflammatory cytokines via AMPK/Nrf2
induction in microglia. Int J Mol Med. 41:1683–1692. 2018.
|
|
350
|
Lee KP, Kang S, Park SJ, Choi YW, Lee YG
and Im DS: Anti-allergic and anti-inflammatory effects of
bakkenolide B isolated from Petasites japonicus leaves. J
Ethnopharmacol. 148:890–894. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
351
|
Li J, Wen PY, Li WW and Zhou J:
Upregulation effects of tanshinone IIA on the expressions of NeuN,
Nissl body, and IκB and downregulation effects on the expressions
of GFAP and NF-κB in the brain tissues of rat models of Alzheimer's
disease. Neuroreport. 26:758–766. 2015. View Article : Google Scholar : PubMed/NCBI
|