Introduction
Breast cancer is the most prevalent cancer among
women, with an estimated 232,670 new cases diagnosed this year (29%
of all malignancies in women); breast cancer ranks second overall
in mortality (15%), with an estimated 40,000 deaths, behind lung
and bronchus carcinoma (1).
Diagnosis at an early stage is beneficial for patients (2,3).
Although there are prognostic factors for long-term survival
(4,5), breast cancer metastasis, especially
to the lungs, remains a significant clinical challenge; the median
survival of these patients is <2 years after diagnosis (6,7).
Therefore, there is an unmet need for prognostic biomarkers that
can predict breast cancer prognosis.
Metastasis of tumor cells involves the physical
translocation of cancer cells to a distant organ and the survival
of metastatic cancer cells at a distant site (8); during the invasion process, cancer
cells attempt to escape immune surveillance. The complement system
is the pillar of innate immunity (9,10);
the activation of complement leads to the efficient clearance of
‘non-self’ and unwanted ‘self’ antigens, including those on tumor
cells (11), via versatile
mechanisms, such as membrane attack complex (MAC)-mediated
cytolysis (12). To escape
complement attack, tumor cells express high levels of membrane
complement regulatory proteins (mCRPs) (13). CD59, the sole membrane regulator in
the terminal stage of complement activation, is considered to be
the most effective mCRP at protecting cancer cells by inhibiting
MAC formation (14). CD59 is
widely expressed, and its expression levels are higher in almost
all cancer cells than in adjacent normal cells (14–16),
which facilitates tumor cell escape from complement attack
(17,18). Although previous reports have
described CD59 as a prognostic biomarker in breast cancer (19–21)
playing important roles in immunotherapy of human malignancies
(22), the role of CD59 in breast
cancer growth and prognosis has not been fully elucidated.
Materials and methods
Ethical standards
The informed consent form was signed by each
participant, and appropriate ethics committee approval was
obtained. All the experiments complied with the current laws of
China.
Cell preparation
The MDA-MB-231 breast carcinoma cell line was
purchased from American Type Culture Collection (ATCC; Manassas,
VA, USA). The MDA-MB-231-HM breast carcinoma cell line, which
readily metastasizes to the lungs, was established previously
(23). Briefly, MDA-MB-231 cells
(2×107) were orthotopically injected into the exposed
axillary mammary fat pad of athymic mice after anesthetization.
Mice were sacrificed after 8–10 weeks under anesthetization and
pulmonary metastatic lesions tissue samples were removed for
primary culture in Leibovitz's L-15 (Gibco) medium supplemented
with 10% (v/v) fetal bovine serum (PAN Biotech) at 37°C in a
humidified atmosphere containing 5% CO2. Cells were
passaged continuously until there were no apparent mesenchymal
cells. Cells grown to ~80% confluence were reinoculated into the
mammary fat pad as described above and tissue samples from
metastatic lesions were again obtained and treated as above. The
procedures were repeated five to six times. A highly
lung-metastatic variant of parental MDA-MB-231 cell line, termed
MDA-MB-231-HM was established, followed by essential detections,
such as Mycoplasma contamination. These two cell lines were
cultured in Leibovitz's L-15 (Gibco) medium supplemented with 10%
(v/v) fetal bovine serum (PAN Biotech) at 37°C in a humidified
atmosphere containing 5% CO2. The pLKO.1-TRC cloning
vector (Addgene plasmid 10878) was utilized to construct two shRNA
plasmids, shCD59 and shCD59', via insertion of shCD59F
5′-CCGGCCGTCAATTGTTCATCTGATTCTCGAGAATCAGATGAACAATTGACGGTTTTTG-3′
and shCD59R
5′-AATTCAAAAACCGTCAATTGTTCATCTGATTCTCGAGAATCAGATGAACAATTGACGG-3′ or
shCD59′F
5′-CCGGGCTAACGTACTACTGCTGCAACTCGAGTTGCAGCAGTAGTACGTTAGCTTTTTG-3′
and shCD59'R
5′-AATTCAAAAAGCTAACGTACTACTGCTGCAACTCGAGTTGCAGCAGTAGTACGTTAGC-3′,
respectively. Lentiviruses containing one of 2 control vectors or
one of 2 CD59-targeting shRNAs were added to MDA-MB-231-HM culture
medium with 8 μg/ml polybrene (Sigma-Aldrich), and the cells were
incubated with the indicated lentiviruses for 24 h. The infected
cells were selected using 1 μg/ml puromycin (Sigma-Aldrich).
Antibodies and other reagents
Anti-β-actin (C4), anti-TFIIB (D-3), anti-CD59
(H-7), anti-CD46 (M177), anti-CD55 (H-7), anti-phospho-ERK1/2
(E-4), anti-p65 (F-6), anti-p50 (E-10), anti-cRel (B-6), anti-Sp1
(E-3), anti-CREB-1 (24H4B), and anti-phosphorylated CREB-1
(Ser-133) antibodies were purchased from Santa Cruz Biotechnology.
Anti-Akt (10176-2-AP) was purchased from ProteinTech Group.
Phospho-AktSer473 (2118-1) antibody was purchased from
Epitomics. FITC-conjugated mouse anti-human CD59 mAb (p282/H19) was
purchased from BD Pharmingen. A rabbit anti-CD59 polyclonal
antibody (10742-1-AP) was obtained from Proteintech Group.
Polybrene, puromycin, and CelLytic M (for mammalian cell lysis and
extraction) were purchased from Sigma-Aldrich. Cell Counting Kit-8
reagents were purchased from Dojindo. U0126 and MK2206 were
purchased from Selleck Chemicals and used at terminal
concentrations of 1 and 2 μM.
Immunoblotting
Total cell lysates were extracted using a mammalian
cell lysis/extraction reagent (Sigma-Aldrich). Nuclear and
cytoplasmic proteins were isolated using a nuclear protein
extraction kit (Active Motif). Immunoblotting was done, essentially
as described (24). The primary
antibodies used were as follows: anti-β-actin (C4; Santa Cruz
Biotechnology), anti-TFIIB (D-3; Santa Cruz Biotechnology),
anti-CD59 (H-7; Santa Cruz Biotechnology), anti-CD46 (M177; Santa
Cruz Biotechnology), anti-CD55 (H-7; Santa Cruz Biotechnology),
anti-ERK1/2 (C-9; Santa Cruz Biotechnology),
anti-anti-phospho-ERK1/2 (E-4; Santa Cruz Biotechnology), anti-p65
(F-6; Santa Cruz Biotechnology), anti-p50 (E-10; Santa Cruz
Biotechnology), anti-cRel (B-6; Santa Cruz Biotechnology), anti-Sp1
(E-3; Santa Cruz Biotechnology), anti-phosphorylated Sp1
(phospho-T453; Bioworld Technology), anti-CREB-1 (24H4B; Santa Cruz
Biotechnology), anti-phosphorylated CREB-1 (Ser-133; Santa Cruz
Biotechnology), rabbit anti-CD59 polyclonal antibody (10742-1-AP;
ProteinTech Group). The quality of loading and transferring was
assessed by immunostaining with the β-actin antibody. The dilutions
for the antibodies were as follows: 1:500 for CD59, 1:500 for CD55,
1:500 for CD46, 1:1,000 for TFIIB, 1:1,000 for p65, 1:500 for p50
1:1,000 for cRel, 1:1,000 for Sp1, 1:500 for phosphorylated Sp1,
1:1,000 for CREB-1, 1:1,000 for phosphorylated CREB-1, 1:500 for
ERK1/2, 1:500 for phospho-ERK1/2, 1:1,000 for rabbit anti CD59
polyclonal antibody and 1:2,000 for β-actin.
Cell proliferation analysis
CCK8 assays were used to measure cell proliferation.
The indicated cells were seeded at a density of 2×103
cells in a volume of 200 μl per well in 96-well plates (Nunc).
Before detection, 100 μl of fresh culture medium was added into
each well after the cells had been washed twice with PBS. Then, 10
μl of CCK8 solution (Dojindo) was added to each well, and the
plates were incubated for 7 days at 37°C. Cell viability was
measured as the absorbance at 450 nm with a microplate reader
(BioTek), and the mean optical density (OD450) values from
triplicate wells for each cell line were used as an index of cell
viability. All the experiments were performed in triplicate.
Flow cytometry analysis
CD59 expression at the cell membrane was detected by
flow cytometry as described previously (24). Briefly, the harvested cells were
incubated with a FITC-conjugated anti-human CD59 antibody (BD
Pharmingen) and then analyzed using a Cytomics FC 500 MPL (Beckman
Coulter).
For cell cycle analysis, cells were digested into
single-cell suspension and 1×106 cells were suspended by
1 ml PBS. Three milliliters Pre-cold ethanol (3 ml) at −20°C was
added into cell suspension with vortex and then the sample was
fixed at −20°C overnight. Fixed cells were collected by
centrifugation at 2,000 rpm for 5 min and the supernatant was
discarded. Cells were suspended by PBS for 15 min at room
temperature. Cells were then recollected by centrifugation at 800
rpm for 5 min and stained by Cell Cycle Staining kit [MultiSciences
(Lianke) Biotechnology] as instructed. Then cell cycle distribution
was analyzed by flow cytometry.
Quantitative real-time PCR
Total RNA was extracted from cells with or without
relative treatments using TRIzol reagent (Invitrogen) and reverse
transcribed into cDNA using PrimeScript™ RT Master Mix (Takara)
following the manufacturer's instructions. Standardized cDNA was
used as template for amplification of 40 cycles with SYBR Green
Master Mix and specific primers of different genes on ABI PRISM
7900HT (Applied Biosystems). β-actin was used as endogenous
control, and samples were analyzed in triplicate. The primers for
amplification of β-actin were 5′-ACC GAGCGCGGCTACAG-3′ (forward)
and 5′-CTTAATGTCACGCACGATTTCC-3′ (reverse). The primers for
amplification of CD59 were 5′-ATGGGAATCCAAGGAGGGTC-3′ (forward) and
5′-ATTGACGGCTGTTTTGCAGT-3′ (reverse).
Colony formation assay
Single-cell suspension was obtained by 0.25% trypsin
digestion and 500 cells were seeded in each well of 6-well plates.
After incubation at an atmosphere of 5% CO2 at 37°C for
15 days, cells were washed with PBS twice and fixed with 4%
paraformaldehyde/PBS (Solarbio) for 30 min. Then cells were stained
by 0.1% crystal violet staining solution (Solarbio) for 30 min and
washed with PBS 3 times. The number of colonies was counted and the
colony formation efficiency was calculated as number of
colonies/number of cells seeded x 100%.
Immunohistochemistry(IHC)staining
Immunohistochemistry for CD59, Akt, and phospho-Akt
was performed on the slides, using a two-step protocol (EnVision™
Detection Systems, Dako, Glostrup, Denmark). The slides were
dewaxed with xylene, gradually hydrated with gradient ethanol, and
washed with phosphate-buffered saline (PBS). Antigens were
retrieved by boiling the slides in citrate buffer (pH 6.0) at 121°C
for 15 min. Endogenous peroxidase was blocked by incubating the
slides in 3% H2O2 solution. Then, the slides
were washed in PBS for 10 min, blocked with 10% normal goat serum
(CW0130; CWBIO, Beijing, China) for 30 min at 37°C, and incubated
with anti-CD59 (H-7; Santa Cruz Biotechnology) at 4°C overnight.
The antibody dilution was 1:100 for the CD59 antibodies and 1:100
for Akt and phospho-Akt antibody. The immunoreactive products were
visualized by the catalysis of 3, 3-diaminobenzidine by horseradish
peroxidase (EnVision™ Detection Systems), following extensive
washing. The slides were then counterstained with Gill hematoxylin
(BA-5021, Zhuhai, China) and dehydrated in ascending grades of
ethanol, and finally, cleared in xylene and mounted under a
coverslip.
Evaluation of immunohistochemistry
analysis
As in our study, the expression of CD59 and
phospho-Akt was semi-quantitatively classified according to the
staining index (SI; range 0–9), which was calculated by multiplying
the staining intensity by the proportion score. The staining
intensities were classified according to four grades (0 denoting
negative; 1, weak; 2, moderate; and 3, strong), and the proportion
score was graded as the percentage of cells that were stained (0
denoting 0 to <10% of cells; 1, between 10 and 30% of cells; 2,
between 31 and 60% of cells; and 3, staining of >60% of cells).
In this study, SI 3 was defined as CD59-positive staining, whereas
SI <3 was defined as negative staining. The IHC scores were
independently determined by two experienced pathologists (Yiqun Du
and Yun Xia) as described previously with slight modifications
(25,26).
Xenograft animal studies
All the mouse experiments were performed according
to a protocol approved by the Fudan University Shanghai Cancer
Center (Shanghai Cancer Center Ethics Committee) Animal Care and
Use Committee. Female Balb/C nude mice, aged 8 weeks, were
purchased from SLAC Laboratory Animals, and were maintained under
specific pathogen-free conditions. The mice were acclimated
in-house for ≥7 days prior to use. A total of 1.0×107
cancer cells were suspended in complete culture medium containing
10% FBS and then injected into the left fourth mammary fat pad of
each female BALB/c nude mouse; a total of seven mice were included
in each group. Tumor size was measured twice weekly with calipers,
and tumor volume was calculated using the following formula:
(width)2 x length/2. At the endpoint of the experiment,
the tumor tissues were carefully excised and weighed after the mice
were anesthetized and sacrificed with CO2.
Collection of human breast cancer
specimens
Between May 2008 and July 2008, 120 women with
breast cancer, as confirmed by core needle biopsy, with no prior
treatment were treated by surgical resection at the Shanghai Cancer
Center, Fudan University. This study was approved by the
independent ethics committee/institutional review board of the
Fudan University Shanghai Cancer Center (Shanghai Cancer Center
Ethics Committee). All patients provided written informed consent
before they were included in this study.
Statistical analysis
All the data are presented as the mean and standard
deviation (SD). The χ2 test was used to evaluate the
relationship between patient characteristics and body mass index
(BMI) and to compare groups based on CD59 expression. Fisher's
exact test was performed when necessary. A multivariate logistic
regression model to predict metastasis was utilized based on both
categorical variables (CD59 expression, tumor size, lymph node
status, cancer embolus, ER status, PR status, and HER-2 status) and
continuous variables (age) that were evaluated at the time of
diagnosis. All the statistical tests were two-sided, and P-values
<0.05 were considered significant. All the analyses were
performed using SPSS (version 13.0; SPSS Inc., Chicago, IL,
USA).
Results
MDA-MB-231-HM cells express higher levels
of CD59, but not of CD46 or CD55, than MDA-MB-231 cells,
potentially via the ERK signaling pathway
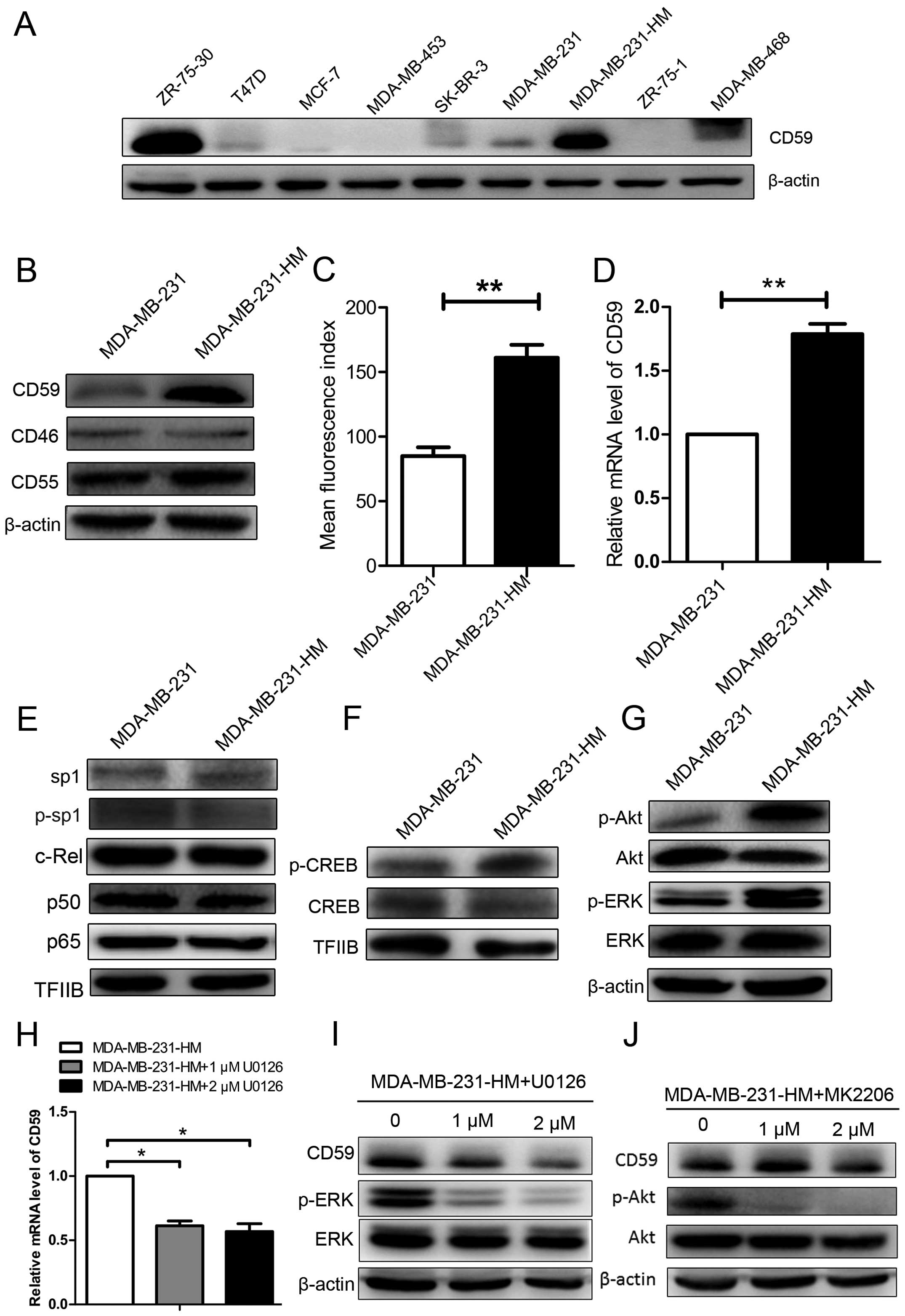
We performed immunoblotting assays with total cell
lysates to elucidate the expression levels of CD59 in breast cancer
cell lines, such as ZR-75-30, T47D, MCF-7, MDA-MB-453, SK-BR-3,
ZR-75-1, MDA-MB-468, MDA-MB-231 cells and the derived MDA-MB-231-HM
cells, which are prone to lung metastasis. CD59 expression was
significantly upregulated in MDA-MB-231-HM cells and ZR-75-30 cells
(Fig. 1A). CD59, but not that of
two other mCRPs (CD46 and CD55) was upregulated in MDA-MB-231-HM
compared with MDA-MB-231 (Fig.
1B). Flow cytometry assays and real-time PCR further confirmed
that CD59 expression was significantly higher in MDA-MB-231-HM
cells compared to the parental cells (Fig. 1C and D).
We recently reported that the constitutive
expression of CD59 is controlled by Sp1, whereas the inducible
expression of CD59 is regulated by NF-κB and enhancer-binding CREB
(24). To investigate the
underlying mechanisms by which MDA-MB-231-HM cells upregulate CD59,
we first examined the nuclear levels of total and phosphorylated
Sp1 and the nuclear translocation of NF-κB. There were no
significant differences between these two cell lines with respect
to Sp1 and NF-κB signaling (Fig.
1E). However, there was increased phosphorylation of CREB,
another transcription regulator, in MDA-MB-231-HM cells compared to
MDA-MB-231 cells (Fig. 1F). CREB
can be directly or indirectly phosphorylated by phospho-Akt and
phospho-ERK (27–29), so we determined the levels of Akt
and ERK phosphorylation. Both phospho-Akt and phospho-ERK were
significantly increased in MDA-MB-231-HM cells compared to
MDA-MB-231 cells (Fig. 1G). The
same result failed to apply to the other group of breast cancer
cells: ZR-75-1 and ZR-75-30 (data not shown), as the levels of
phosphorylated CREB between ZR-75-1 and ZR-75-30 showed no
significant difference. This is understandable considering the
complicated regulation of CD59 and that MDA-MB-231-HM was derived
from MDA-MB-231. To investigate the mechanism underlying the
upregulation of CD59 in cell MDA-MB-231-HM, we inhibited ERK
phosphorylation and Akt phosphorylation using U0126 and MK2206,
respectively. The inhibition of ERK phosphorylation downregulated
CD59 expression both at mRNA level and protein level (Fig. 1H and I), while the inhibition of
Akt phosphorylation failed (Fig.
1J). These findings suggested that CD59 was upregulated in
breast cancer MDA-MB-231-HM cells and that this upregulation may
occur via the ERK pathway.
CD59 knockdown suppresses the tumor
growth of MDA-MB-231-HM cells
Breast cancer lung metastasis leads to a poor
prognosis (6,7). We utilized MDA-MB-231-HM cells, which
are prone to lung metastasis, to explore the role of CD59 in the
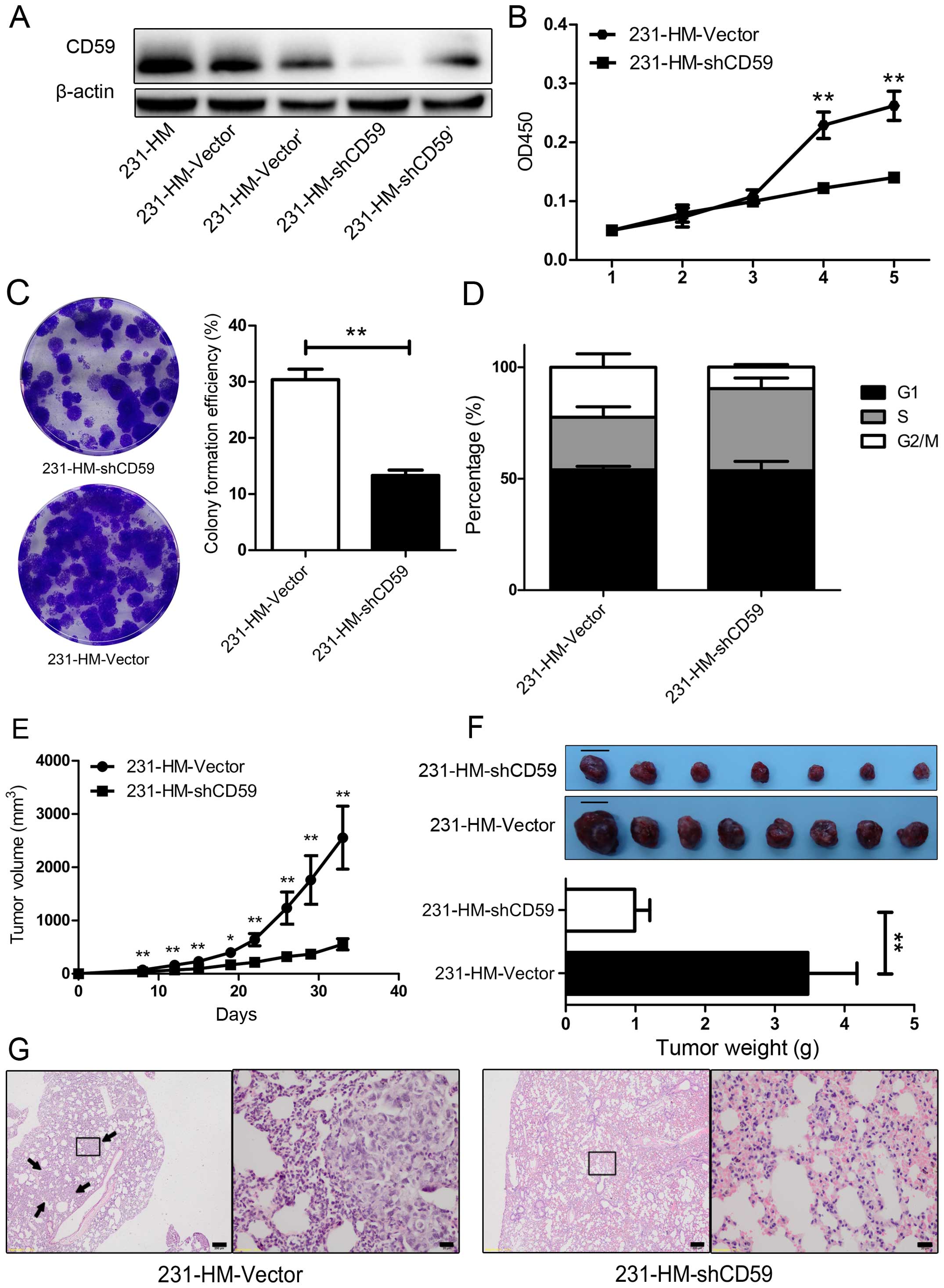
highly metastatic MDA-MB-231-HM cells. We knocked down CD59
expression in MDA-MB-231-HM cells using two vectors expressing
shRNA against CD59, referred to as 231-HM-shCD59 and 231-HM-shCD59'
(the corresponding controls were designated 231-HM-Vector and
231-HM-Vector', respectively). As shown in Fig. 2A, CD59 was robustly knocked down in
MDA-MB-231-HM cells. 231-HM-shCD59 was more efficient at knocking
down CD59 expression, and we therefore selected this construct for
use in subsequent studies.
We then used CCK8 assays to determine the effect of
CD59 knockdown on cell proliferation. As shown in Fig. 2B, 231-HM-shCD59 cells lacking CD59
expression grew more slowly than 231-HM-Vector cells beginning on
day 4 after cell plating (P<0.01). Colony formation assay showed
similar result (P<0.01) (Fig.
2C), and cell cycle analysis indicated that this growth arrest
may due to the cell cycle arrest in 231-HM-shCD59 cells (Fig. 2D). We next implanted 231-HM-Vector
and 231-HM-shCD59 cells into the left fourth mammary fat pads of
female athymic nude mice. Primary tumors developed successfully in
all mice, and CD59 knockdown markedly decreased tumor growth, as
evidenced by tumor size (Fig. 2E,
P<0.02, P<0.01). At 5 weeks after injection, all the mice
were sacrificed, and their tumors and lungs were resected, as
MDA-MB-231-HM is prone to lung metastasis. The tumors were weighed,
and the lungs were examined by hematoxylin and eosin (H&E)
staining. The tumors in the 231-HM-shCD59-injected mice were
clearly much smaller than those in the control mice, and the
corresponding tumor weights were consistent with this observation
(representative images are shown in Fig. 2F, P<0.01). In the
H&E-stained slices, we found metastases in 3/10 lungs from the
231-HM-Vector group, whereas none of the 10 lungs from the
231-HM-shCD59 group had meta-static lesions; representative images
are shown in Fig. 2G. These data
indicated that CD59 knockdown suppressed cell proliferation both
in vitro and in vivo and may decrease lung metastasis
in vivo.
High CD59 expression levels were
associated with lung metastasis and poor prognosis in breast cancer
patients
To explore the clinical significance of CD59
expression in prognosis, we analyzed a cohort of 120 breast cancer
patients, 58 with lung metastases and 62 without metastatic
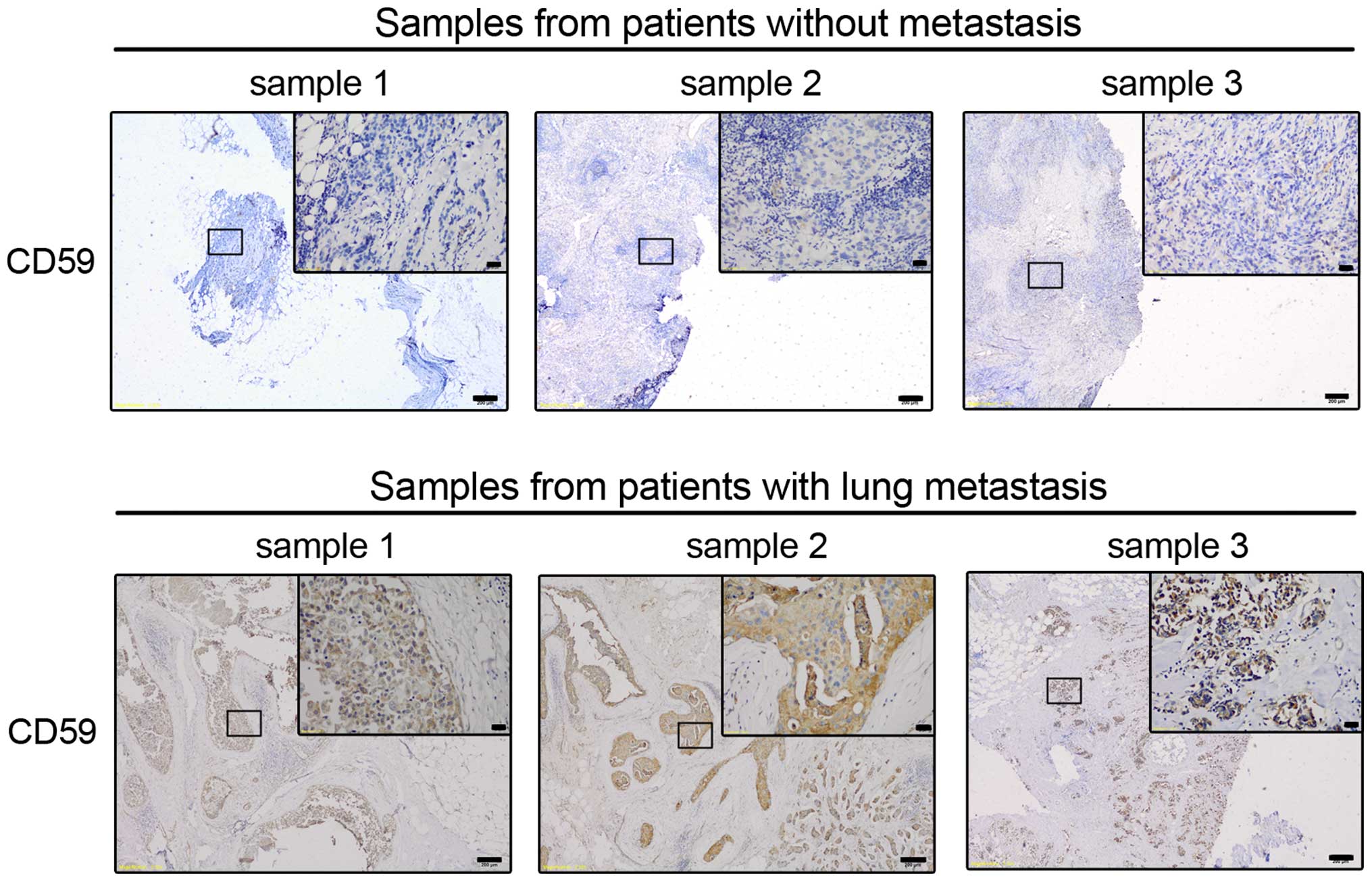
disease. We performed IHC staining for CD59 expression, and the
expression scores were ranked by two independent pathologists who
were blinded to the identity of each sample. Representative images
of CD59 in 3 patients with metastasis and in 3 patients without
metastasis are shown in Fig. 3. We
retrospectively collected complete clinical data, including age,
tumor size, pathological assessment and follow-up visit details.
Table I shows the characteristics
of the patients grouped based on CD59 expression. Only the presence
of metastasis was significantly associated with CD59 expression
(P=0.001). There was a higher percentage of HER-2-negative patients
among those with high CD59 expression. However, age, tumor size,
node status, tumor histology, ER status, PR status and HER-2 status
were not significantly correlated with CD59 expression.
 | Table IResults of the correlation analysis
of CD59 expression in 120 patients with breast cancer. |
Table I
Results of the correlation analysis
of CD59 expression in 120 patients with breast cancer.
| Characteristic | Patients (no.) | CD59 low | CD59 high | P-value |
|---|
| Age | | | | |
| ≤35 | 13 | 6 | 7 | 0.578a |
| >35 | 107 | 48 | 59 | |
| Size (cm) | | | | |
| <2 | 21 | 7 | 14 | 0.139a |
| ≥2, <5 | 90 | 45 | 45 | |
| ≥5 | 9 | 2 | 7 | |
| Grade | | | | |
| I–II | 81 | 35 | 46 | 0.354a |
| III | 39 | 19 | 20 | |
| Lymph node
status | | | | |
| − | 76 | 38 | 38 | 0.104a |
| + | 44 | 16 | 28 | |
| ER | | | | |
| − | 52 | 19 | 33 | 0.074a |
| + | 68 | 35 | 33 | |
| PR | | | | |
| − | 68 | 27 | 41 | 0.126a |
| + | 52 | 27 | 25 | |
| Her-2 | | | | |
| − | 91 | 39 | 52 | 0.267a |
| + | 28 | 15 | 14 | |
| Cancer embolus | | | | |
| − | 84 | 41 | 43 | 0.140a |
| + | 36 | 13 | 23 | |
| Metastasis | | | | |
| − | 65 | 40 | 25 |
0.001a |
| + | 55 | 14 | 41 | |
The metastasis rate was 48.3% (58/120) in the study
cohort. Table II shows the
univariate and multivariate analyses of the metastasis rate
predictors using a logistic regression model. In the univariate
model, there were no significant relationships between metastasis
rate and tumor size, tumor histology, ER status, PR status or HER-2
status (P>0.05). However, metastasis was significantly
associated with age, lymph node status, cancer embolus and CD59
expression (P<0.05). Patients 35 years or older, as well as
those with positive lymph node status, cancer embolus or high CD59
expression were more likely to develop metastases. In the
multivariate model, CD59 expression, lymph node status and cancer
embolus were independent predictors of metastasis, with P-values of
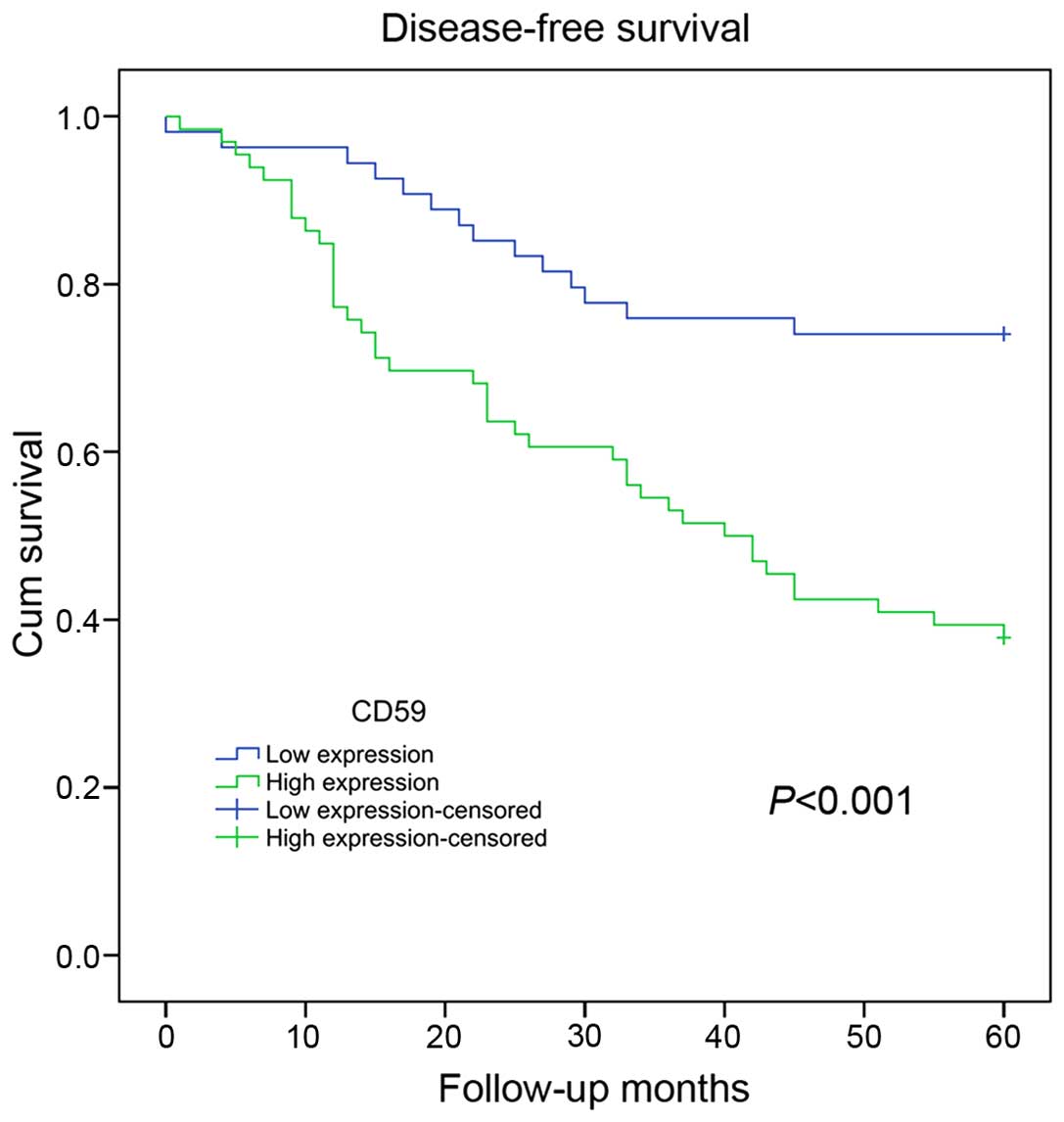
0.041, 0.032 and 0.008, respectively. Notably, a Kaplan-Meier
survival analysis revealed that CD59 expression levels were
negatively correlated with relapse-free survival among breast
cancer patients (P<0.001). The analysis indicated that patients
with higher CD59 expression would have a worse prognosis compared
to patients with lower CD59 expression (Fig. 4). These clinical data suggested
that CD59 expression may be a prognostic biomarker for metastasis
and outcome in breast cancer patients.
| Table IIResults of the univariate and
multivariate analyses of CD59 expression in 120 patients with
breast cancer. |
Table II
Results of the univariate and
multivariate analyses of CD59 expression in 120 patients with
breast cancer.
| Univariate
analysis | Multivariate
analysis |
|---|
|
|
|
|---|
| | 95% CI | | | 95% CI | |
|---|
| |
| | |
| |
|---|
| Variable | HR | Lower | Upper | P-value | HR | Lower | Upper | P-value |
|---|
| Age | 0.962 | 0.937 | 0.988 | 0.004 | 0.982 | 0.957 | 1.007 | 0.163 |
| Tumor size | 1.148 | 0.622 | 2.117 | 0.659 | | | | |
| ER | 0.692 | 0.408 | 1.176 | 0.174 | | | | |
| PR | 0.799 | 0.466 | 1.371 | 0.415 | | | | |
| Her-2 | 1.572 | 0.887 | 2.787 | 0.121 | | | | |
| Grade | 1.612 | 0.940 | 2.766 | 0.083 | | | | |
| Lymph node
status | 4.793 | 2.746 | 8.366 | 0.000 | 2.270 | 1.033 | 4.987 | 0.041 |
| Cancer embolus | 4.868 | 2.828 | 8.379 | 0.000 | 2.312 | 1.074 | 4.980 | 0.032 |
| CD59 | 3.074 | 1.673 | 5.648 | 0.000 | 2.328 | 1.253 | 4.326 | 0.008 |
Discussion
Human CD59 is a 18–20-kDa membrane complement
regulatory protein (mCRP) that is anchored to the cell membrane by
glycophosphatidylinositol (GPI) and protects innocent host cells
from activated complement destruction (15). In most solid tumors, CD59 is
overexpressed compared to adjacent normal cells, and inhibiting
CD59 function can improve breast cancer treatment (14,30,31).
Distant metastases are lethal to breast cancer patients (7). We hypothesized that CD59 may protect
tumor cells from complement attack, thus facilitating breast cancer
metastasis, leading to poor prognosis. Because lung metastasis is
unequivocally a clinical challenge in breast cancer treatment and
leads to poor prognosis in breast cancer patients (6,7), we
utilized the MDA-MB-231-HM cell line to elucidate the roles of
mCRPs; MDA-MB-231-HM cells are more prone to lung metastasis than
parental MDA-MB-231 cells. We discovered that among the three
mCRPs, only CD59 was upregulated in MDA-MB-231-HM cells. After
knocking down CD59 with a specific shRNA, we surprisingly found
that MDA-MB-231-HM cell growth was significantly suppressed both
in vitro and in vivo. CD59 knockdown in MDA-MB-231-HM
cells inhibited the formation of metastatic lesions in the
orthotopic xenograft model, although it was not statistically
significant. We also determined that CD59 over-expression was
associated with poor prognosis in a cohort of 120 breast cancer
patients. We first demonstrated that CD59 was associated with
MDA-MB-231-HM cell growth in vitro and in vivo and
then correlated CD59 levels with breast cancer poor prognosis.
CD59 transcriptional regulation involves many
pathways (24), including p-CREB,
as confirmed by Du et al in our laboratory research group,
by performing the CHIP assays. In this study, we showed that CD59
upregulation in MDA-MB-231-HM cells is related to p-CREB,
furthermore, CD59 upregulation in MDA-MB-231-HM cells may result
from ERK pathway activation which plays critical roles in cancer
(32); one of its functions may be
to upregulate CD59 through CREB (24,27–29).
The protective roles of mCRPs, especially CD59, may facilitate
tumor cell evasion from complement attack; the mCRPs are therefore
obstacles to antibody-based immunotherapy for cancer (13,14).
In addition, CD59 overexpression protects MCF-7 breast cancer cells
from complement-mediated cytolysis and promotes MCF-7 cell
proliferation by inhibiting BCL-2 expression; conversely, CD59
knockdown upregulates Fas and caspase-3 to induce apoptosis
(20). Besides, CD59 silencing in
NSCLC cancer cells can enhance complement-mediated cell apoptosis
and inhibit the growth of NSCLC (16), which shows accordance with our
study.
Inspiring studies by Madjd et al (19) and Terp et al (21) demonstrated that the loss of CD59
expression in breast tumors correlates with poor survival (19), and that CD59 is inversely
correlated with metastatic capability (21). Here, we showed that CD59
overexpression was associated with MDA-MB-231-HM cell growth as
well as with poor outcome for breast cancer patients. Though CD59
knock-down suppressed cell proliferation both in vitro and
in vivo, there were no significant relationships between
CD59 and tumor size in our clinical investigation. Corresponding to
the clinical-tumor size does not present positive correlation with
the prognosis of breast cancer. In the large study involving 520
patients reported by Madjd et al (19), lung metastasis was not evaluated,
whereas we focused on lung metastasis and established a cohort of
120 patients with or without lung metastasis. In addition, in our
study, a greater proportion of HR-negative and Her-2-negative
patients had high CD59 expression. These differences may explain
the discrepant conclusions. Terp et al (21) showed that CD59 is ‘associated’ with
the aggressiveness of metastasis rather than metastasis
colonization per se. This may be due to the inhibition of
cell growth; in our study, CD59 knockdown resulted in a better
‘outcome’ for the mice, so the failure to observe metastatic
lesions may due to cell growth inhibition during our experimental
time. However, it has been reported that CD59 can mediate signal
transmission through the Src family member Lck in T cells (33). We cannot eliminate the possibility
that in MDA-MB-231-HM cells, complex signals may be transmitted by
CD59 to promote cell proliferation and metastasis.
As a biomarker in breast cancer, CD59 may play
versatile roles. This study revealed that CD59 overexpression may
be a prognostic factor for poor survival in breast cancer patients.
However, the mechanism of action of CD59 and its controversial
roles in breast cancer, perhaps through Src family members or other
signaling pathways, remain to be elucidated. Besides, as an
immunotherapy ‘candidate’, CD59 may have other than the known roles
in breast cancer treatment (22).
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China (81201531), the 2012 Shanghai
Committee of Science and Technology Funds (12ZR1406200, 12DZ2260100
and 12140901502), and the Shanghai Committee of Science and
Technology Fund for 2013, Qimingxing Project (11QA1401400 to X.
Hu). We also would like to thank American Journal Experts editorial
team for their language polish.
References
|
1
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Elmore JG, Armstrong K, Lehman CD and
Fletcher SW: Screening for breast cancer. JAMA. 293:1245–1256.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tabár L, Vitak B, Chen HH, Duffy SW, Yen
MF, Chiang CF, Krusemo UB, Tot T and Smith RA: The Swedish
Two-County Trial twenty years later. Updated mortality results and
new insights from long-term follow-up. Radiol Clin North Am.
38:625–651. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Soerjomataram I, Louwman MW, Ribot JG,
Roukema JA and Coebergh JW: An overview of prognostic factors for
long-term survivors of breast cancer. Breast Cancer Res Treat.
107:309–330. 2008. View Article : Google Scholar :
|
|
5
|
Foulkes WD, Reis-Filho JS and Narod SA:
Tumor size and survival in breast cancer - a reappraisal. Nat Rev
Clin Oncol. 7:348–353. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Anan K, Mitsuyama S, Koga K, Tanabe R,
Saimura M, Tanabe Y, Watanabe M, Suehara N, Matsunaga H, Nishihara
K, et al: Disparities in the survival improvement of recurrent
breast cancer. Breast Cancer. 17:48–55. 2010. View Article : Google Scholar
|
|
7
|
Nguyen DX, Bos PD and Massagué J:
Metastasis: From dissemination to organ-specific colonization. Nat
Rev Cancer. 9:274–284. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chaffer CL and Weinberg RA: A perspective
on cancer cell metastasis. Science. 331:1559–1564. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Morgan BP, Marchbank KJ, Longhi MP, Harris
CL and Gallimore AM: Complement: Central to innate immunity and
bridging to adaptive responses. Immunol Lett. 97:171–179. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ricklin D, Hajishengallis G, Yang K and
Lambris JD: Complement: A key system for immune surveillance and
homeo-stasis. Nat Immunol. 11:785–797. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Matsumoto M, Takeda J, Inoue N, Hara T,
Hatanaka M, Takahashi K, Nagasawa S, Akedo H and Seya T: A novel
protein that participates in nonself discrimination of malignant
cells by homologous complement. Nat Med. 3:1266–1270. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dunkelberger JR and Song WC: Complement
and its role in innate and adaptive immune responses. Cell Res.
20:34–50. 2010. View Article : Google Scholar
|
|
13
|
Zhou X, Hu W and Qin X: The role of
complement in the mechanism of action of rituximab for B-cell
lymphoma: Implications for therapy. Oncologist. 13:954–966. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fishelson Z, Donin N, Zell S, Schultz S
and Kirschfink M: Obstacles to cancer immunotherapy: Expression of
membrane complement regulatory proteins (mCRPs) in tumors. Mol
Immunol. 40:109–123. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Davies A and Lachmann PJ: Membrane defence
against complement lysis: The structure and biological properties
of CD59. Immunol Res. 12:258–275. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li B, Lin H, Fan J, Lan J, Zhong Y, Yang
Y, Li H and Wang Z: CD59 is overexpressed in human lung cancer and
regulates apoptosis of human lung cancer cells. Int J Oncol.
43:850–858. 2013.PubMed/NCBI
|
|
17
|
Hu W, Ge X, You T, Xu T, Zhang J, Wu G,
Peng Z, Chorev M, Aktas BH, Halperin JA, et al: Human CD59
inhibitor sensitizes rituximab-resistant lymphoma cells to
complement-mediated cytolysis. Cancer Res. 71:2298–2307. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rus HG, Niculescu FI and Shin ML: Role of
the C5b-9 complement complex in cell cycle and apoptosis. Immunol
Rev. 180:49–55. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Madjd Z, Pinder SE, Paish C, Ellis IO,
Carmichael J and Durrant LG: Loss of CD59 expression in breast
tumours correlates with poor survival. J Pathol. 200:633–639. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li B, Chu X, Gao M and Xu Y: The effects
of CD59 gene as a target gene on breast cancer cells. Cell Immunol.
272:61–70. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Terp MG, Lund RR, Jensen ON, Leth-Larsen R
and Ditzel HJ: Identification of markers associated with highly
aggressive meta-static phenotypes using quantitative comparative
proteomics. Cancer Genomics Proteomics. 9:265–273. 2012.PubMed/NCBI
|
|
22
|
Maio M, Brasoveanu LI, Coral S, Sigalotti
L, Lamaj E, Gasparollo A, Visintin A, Altomonte M and Fonsatti E:
Structure, distribution, and functional role of protectin (CD59) in
complement-susceptibility and in immunotherapy of human
malignancies (Review). Int J Oncol. 13:305–318. 1998.PubMed/NCBI
|
|
23
|
Liu ZB, Hou YF, Min-Dong, Di GH, Wu J,
Shen ZZ and Shao ZM: PA-MSHA inhibits proliferation and induces
apoptosis through the up-regulation and activation of caspases in
the human breast cancer cell lines. J Cell Biochem. 108:195–206.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Du Y, Teng X, Wang N, Zhang X, Chen J,
Ding P, Qiao Q, Wang Q, Zhang L, Yang C, et al: NF-κB and
enhancer-binding CREB protein scaffolded by CREB-binding protein
(CBP)/p300 proteins regulate CD59 protein expression to protect
cells from complement attack. J Biol Chem. 289:2711–2724. 2014.
View Article : Google Scholar :
|
|
25
|
Detre S, Saclani Jotti G and Dowsett M: A
‘quickscore’ method for immunohistochemical semiquantitation:
Validation for oestrogen receptor in breast carcinomas. J Clin
Pathol. 48:876–878. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Harvey JM, Clark GM, Osborne CK and Allred
DC: Estrogen receptor status by immunohistochemistry is superior to
the ligand-binding assay for predicting response to adjuvant
endocrine therapy in breast cancer. J Clin Oncol. 17:1474–1481.
1999.PubMed/NCBI
|
|
27
|
Du K and Montminy M: CREB is a regulatory
target for the protein kinase Akt/PKB. J Biol Chem.
273:32377–32379. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Choi YH, Lee SN, Aoyagi H, Yamasaki Y, Yoo
JY, Park B, Shin DM, Yoon HG and Yoon JH: The extracellular
signal-regulated kinase mitogen-activated protein kinase/ribosomal
S6 protein kinase 1 cascade phosphorylates cAMP response
element-binding protein to induce MUC5B gene expression via
D-prostanoid receptor signaling. J Biol Chem. 286:34199–34214.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Boulware MI, Weick JP, Becklund BR, Kuo
SP, Groth RD and Mermelstein PG: Estradiol activates group I and II
metabotropic glutamate receptor signaling, leading to opposing
influences on cAMP response element-binding protein. J Neurosci.
25:5066–5078. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu M, Yang YJ, Zheng H, Zhong XR, Wang Y,
Wang Z, Wang YG and Wang YP: Membrane-bound complement regulatory
proteins are prognostic factors of operable breast cancer treated
with adjuvant trastuzumab: A retrospective study. Oncol Rep.
32:2619–2627. 2014.PubMed/NCBI
|
|
31
|
Mamidi S, Cinci M, Hasmann M, Fehring V
and Kirschfink M: Lipoplex mediated silencing of membrane
regulators (CD46, CD55 and CD59) enhances complement-dependent
anti-tumor activity of trastuzumab and pertuzumab. Mol Oncol.
7:580–594. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Samatar AA and Poulikakos PI: Targeting
RAS-ERK signalling in cancer: Promises and challenges. Nat Rev Drug
Discov. 13:928–942. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lipp AM, Juhasz K, Paar C, Ogris C,
Eckerstorfer P, Thuenauer R, Hesse J, Nimmervoll B, Stockinger H,
Schütz GJ, et al: Lck mediates signal transmission from CD59 to the
TCR/CD3 pathway in Jurkat T cells. PLoS One. 9:e859342014.
View Article : Google Scholar : PubMed/NCBI
|